
Do DEEPER ROOTING 1 Homologs Regulate the Lateral Root Slope Angle in Cucumber (Cucumis sativus)?

 ,
,  and
and 
Abstract
:1. Introduction
2. Results
2.1. Identification of the Putative Ortholog(s) of Arabidopsis and Rice DEEPER ROOTING1 (DRO1) in Cucumis sativus by Phylogenetic Analysis
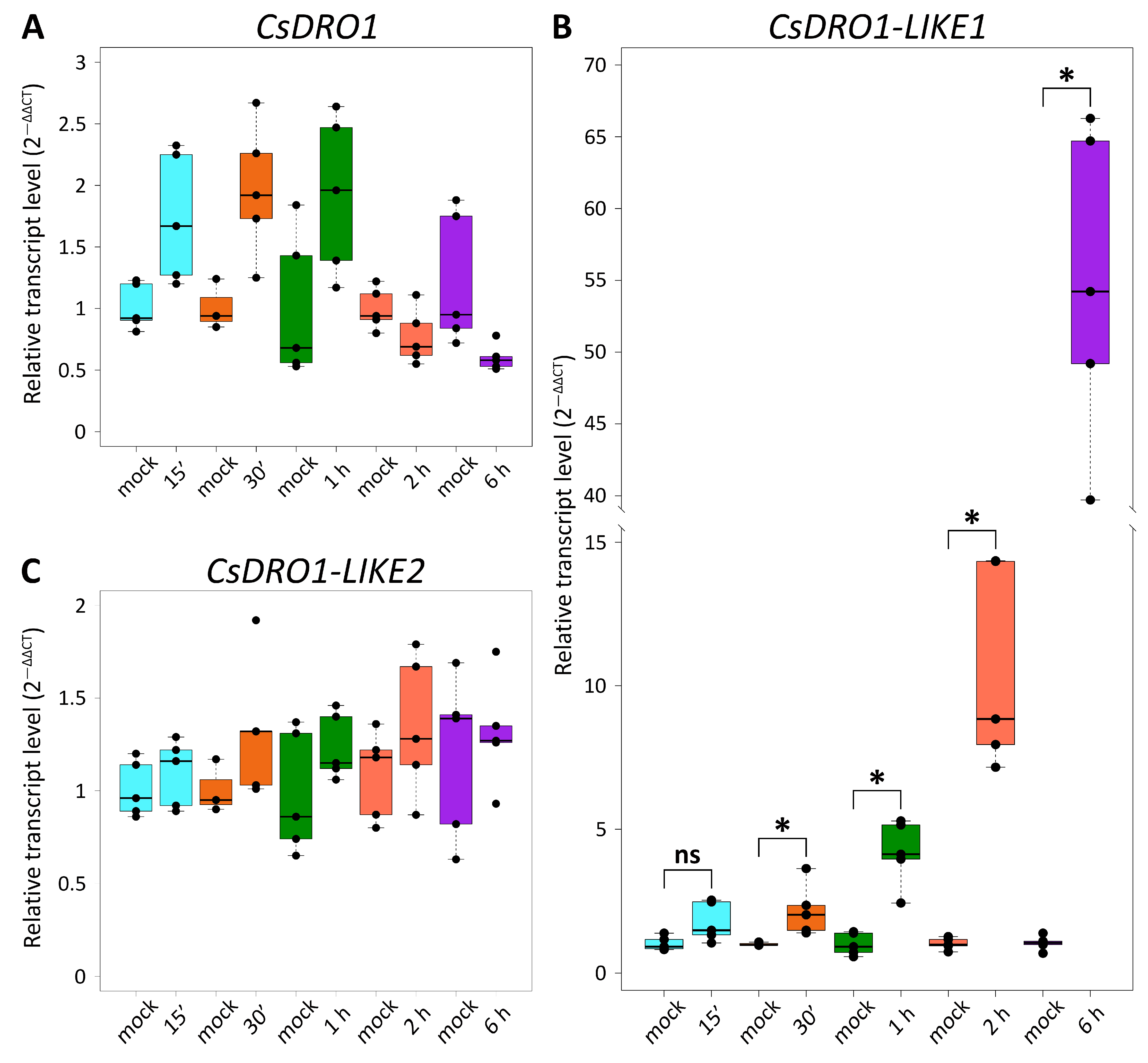
2.2. Expression Levels of Cucumber DRO Genes Differ between Organs and in Their Response to Exogenous Auxin
2.3. Expression Patterns of Cucumis sativus DRO Genes in the Root Tip and during Lateral Root Emergence Based on Promoter–Reporter Fusions in Transgenic Hairy Roots
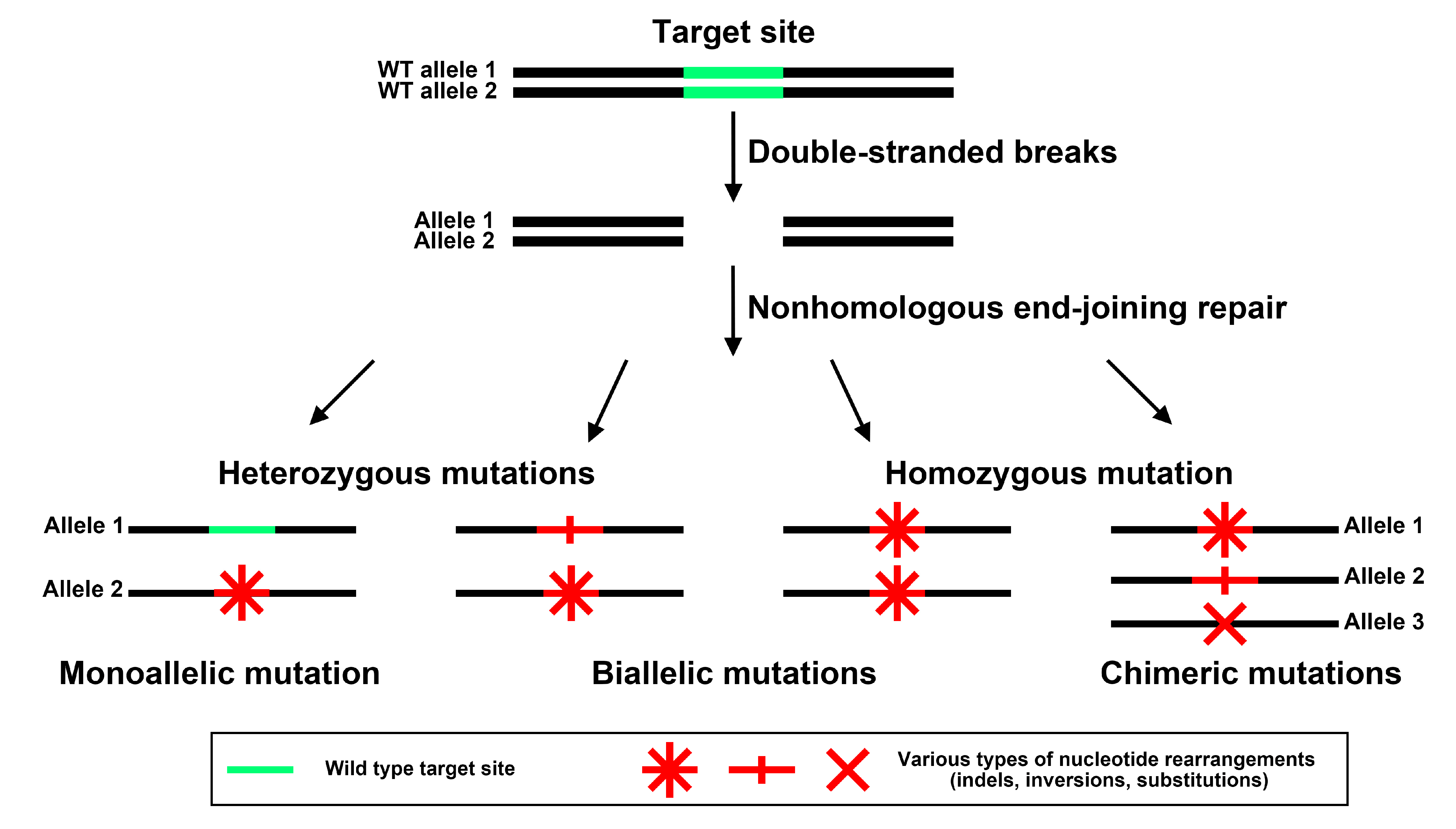
2.4. CRISPR/Cas9-Mediated Genome Editing in Transgenic Cucumber Roots
2.4.1. Identification of Mutations in a Single Cucumber DRO1, DRO1L1 or DRO1L2 Gene
2.4.2. Knockout of Any Individual DRO Gene Does Not Change the Lateral Root Slope Angle
3. Discussion
4. Materials and Methods
4.1. Plant Material and Bacterial Strains
4.2. Phylogeny and Bioinformatics
4.3. Molecular Cloning, Plasmid Construction, and Plant Transformation
4.3.1. Genetic Constructs for Promoter–Reporter Fusions
4.3.2. Construction of Vectors for the CRISPR-Cas9-Mediated Genome Editing
4.3.3. Plant Transformation
4.3.4. Preparation of Transgenic Cucumber Roots for Detection of CRISPR/Cas9-Mediated Mutations
4.4. Treatments with Exogenous Auxin
4.5. RT-qPCR Assays
4.6. Fluorescence Protein Reporter Assays and Microscopy
4.7. Statistical Analyses
5. Conclusions and Perspectives
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Abbreviations
References
- Koevoets, I.T.; Venema, J.H.; Elzenga, J.T.M.; Testerink, C. Roots Withstanding their Environment: Exploiting Root System Architecture Responses to Abiotic Stress to Improve Crop Tolerance. Front. Plant Sci. 2016, 7, 1335. [Google Scholar] [CrossRef]
- Lynch, J.P. Harnessing root architecture to address global challenges. Plant J. 2022, 109, 415–431. [Google Scholar] [CrossRef]
- Pierret, A. Will deeper roots be enough? Engineering drought-resistant crops will entail in-depth understanding of root hydraulic architecture. A Commentary on ‘Root and xylem anatomy varies with root length, root order, soil depth and environment’. Ann. Bot. 2022, 130, XV–XVII. [Google Scholar] [CrossRef]
- Osmont, K.S.; Sibout, R.; Hardtke, C.S. Hidden Branches: Developments in Root System Architecture. Annu. Rev. Plant Biol. 2007, 58, 93–113. [Google Scholar] [CrossRef] [PubMed]
- Motte, H.; Vanneste, S.; Beeckman, T. Molecular and Environmental Regulation of Root Development. Annu. Rev. Plant Biol. 2019, 70, 465–488. [Google Scholar] [CrossRef] [PubMed]
- Satbhai, S.B.; Ristova, D.; Busch, W. Underground tuning: Quantitative regulation of root growth. J. Exp. Bot. 2015, 66, 1099–1112. [Google Scholar] [CrossRef] [PubMed]
- Uga, Y.; Sugimoto, K.; Ogawa, S.; Rane, J.; Ishitani, M.; Hara, N.; Kitomi, Y.; Inukai, Y.; Ono, K.; Kanno, N.; et al. Control of root system architecture by DEEPER ROOTING 1 increases rice yield under drought conditions. Nat. Genet. 2013, 45, 1097–1102. [Google Scholar] [CrossRef] [PubMed]
- Kitomi, Y.; Hanzawa, E.; Kuya, N.; Inoue, H.; Hara, N.; Kawai, S.; Kanno, N.; Endo, M.; Sugimoto, K.; Yamazaki, T.; et al. Root angle modifications by the DRO1 homolog improve rice yields in saline paddy fields. Proc. Natl. Acad. Sci. USA 2020, 117, 21242–21250. [Google Scholar] [CrossRef] [PubMed]
- Guseman, J.M.; Webb, K.; Srinivasan, C.; Dardick, C. DRO1 influences root system architecture in Arabidopsis and Prunus species. Plant J. 2017, 89, 1093–1105. [Google Scholar] [CrossRef] [PubMed]
- Yoshihara, T.; Spalding, E.P. LAZY Genes Mediate the Effects of Gravity on Auxin Gradients and Plant Architecture. Plant Physiol. 2017, 175, 959–969. [Google Scholar] [CrossRef]
- Sun, C.; Liang, W.; Yan, K.; Xu, D.; Qin, T.; Fiaz, S.; Kear, P.; Bi, Z.; Liu, Y.; Liu, Z.; et al. Expression of Potato StDRO1 in Arabidopsis Alters Root Architecture and Drought Tolerance. Front. Plant Sci. 2022, 13, 836063. [Google Scholar] [CrossRef]
- Dardick, C.; Callahan, A.; Horn, R.; Ruiz, K.B.; Zhebentyayeva, T.; Hollender, C.; Whitaker, M.; Abbott, A.; Scorza, R. PpeTAC1 promotes the horizontal growth of branches in peach trees and is a member of a functionally conserved gene family found in diverse plants species. Plant J. 2013, 75, 618–630. [Google Scholar] [CrossRef]
- Waite, J.M.; Dardick, C. The roles of the IGT gene family in plant architecture: Past, present, and future. Curr. Opin. Plant Biol. 2021, 59, 101983. [Google Scholar] [CrossRef]
- Ashraf, A.; Rehman, O.U.; Muzammil, S.; Léon, J.; Naz, A.A.; Rasool, F.; Ali, G.M.; Zafar, Y.; Khan, M.R. Evolution of Deeper Rooting 1-like homoeologs in wheat entails the C-terminus mutations as well as gain and loss of auxin response elements. PLoS ONE 2019, 14, e0214145. [Google Scholar] [CrossRef]
- Li, Z.; Liang, Y.; Yuan, Y.; Wang, L.; Meng, X.; Xiong, G.; Zhou, J.; Cai, Y.; Han, N.; Hua, L.; et al. OsBRXL4 Regulates Shoot Gravitropism and Rice Tiller Angle through Affecting LAZY1 Nuclear Localization. Mol. Plant 2019, 12, 1143–1156. [Google Scholar] [CrossRef]
- Furutani, M.; Hirano, Y.; Nishimura, T.; Nakamura, M.; Taniguchi, M.; Suzuki, K.; Oshida, R.; Kondo, C.; Sun, S.; Kato, K.; et al. Polar recruitment of RLD by LAZY1-like protein during gravity signaling in root branch angle control. Nat. Commun. 2020, 11, 76. [Google Scholar] [CrossRef]
- Yang, P.; Wen, Q.; Yu, R.; Han, X.; Deng, X.W.; Chen, H. Light modulates the gravitropic responses through organ-specific PIFs and HY5 regulation of LAZY4 expression in Arabidopsis. Proc. Natl. Acad. Sci. USA 2020, 117, 18840–18848. [Google Scholar] [CrossRef] [PubMed]
- Chen, J.; Yu, R.; Li, N.; Deng, Z.; Zhang, X.; Zhao, Y.; Qu, C.; Yuan, Y.; Pan, Z.; Zhou, Y.; et al. Amyloplast sedimentation repolarizes LAZYs to achieve gravity sensing in plants. Cell 2023, 186, 4788–4802.e15. [Google Scholar] [CrossRef] [PubMed]
- Szemenyei, H.; Hannon, M.; Long, J.A. TOPLESS mediates auxin-dependent transcriptional repression during Arabidopsis embryogenesis. Science 2008, 319, 1384–1386. [Google Scholar] [CrossRef] [PubMed]
- Mullen, J.; Hangarter, R. Genetic analysis of the gravitropic set-point angle in lateral roots of arabidopsis. Adv. Space Res. 2003, 31, 2229–2236. [Google Scholar] [CrossRef] [PubMed]
- Guyomarc’H, S.; Léran, S.; Auzon-Cape, M.; Perrine-Walker, F.; Lucas, M.; Laplaze, L. Early development and gravitropic response of lateral roots in Arabidopsis thaliana. Philos. Trans. R. Soc. B Biol. Sci. 2012, 367, 1509–1516. [Google Scholar] [CrossRef] [PubMed]
- Demchenko, K.N.; Demchenko, N.P. Changes of root structure in connection with the development of lateral root primordia in wheat and pumpkins. In Recent Advances of Plant Root Structure and Function. Developments in Plant and Soil Sciences; Gašparíková, O., Čiamporová, M., Mistrík, I., Baluška, F., Eds.; Springer: Dordrecht, The Netherlands, 2001; Volume 90, pp. 39–47. [Google Scholar] [CrossRef]
- Ilina, E.L.; Kiryushkin, A.S.; Semenova, A.V.; Demchenko, N.P.; Pawlowski, K.; Demchenko, K.N. Lateral root initiation and formation within the parental root meristem of Cucurbita pepo: Is auxin a key player? Ann. Bot. 2018, 122, 873–888. [Google Scholar] [CrossRef] [PubMed]
- Xiao, T.T.; van Velzen, R.; Kulikova, O.; Franken, C.; Bisseling, T. Lateral root formation involving cell division in both pericycle, cortex and endodermis is a common and ancestral trait in seed plants. Development 2019, 146, dev.182592. [Google Scholar] [CrossRef] [PubMed]
- Chiang, Y.-L. Macro- and microscopic structure of the root of Ceratopteris pteridoides (Hook.) Hieron. Taiwania 1970, 15, 31–49. [Google Scholar] [CrossRef]
- Chen, J.-F.; Staub, J.E.; Jiang, J. A reevaluation of karyotype in cucumber (Cucumis sativus L.). Genet. Resour. Crop Evol. 1998, 45, 301–305. [Google Scholar] [CrossRef]
- Gasteiger, E.; Gattiker, A.; Hoogland, C.; Ivanyi, I.; Appel, R.D.; Bairoch, A. ExPASy: The proteomics server for in-depth protein knowledge and analysis. Nucleic Acids Res. 2003, 31, 3784–3788. [Google Scholar] [CrossRef]
- Huang, L.; Schiefelbein, J. Conserved gene expression programs in developing roots from diverse plants. Plant Cell 2015, 27, 2119–2132. [Google Scholar] [CrossRef]
- Li, Z.; Zhang, Z.; Yan, P.; Huang, S.; Fei, Z.; Lin, K. RNA-Seq improves annotation of protein-coding genes in the cucumber genome. BMC Genom. 2011, 12, 540. [Google Scholar] [CrossRef]
- Fukaki, H.; Okushima, Y.; Tasaka, M. Regulation of lateral root formation by auxin signaling in Arabidopsis. Plant Biotechnol. 2005, 22, 393–399. [Google Scholar] [CrossRef]
- Rosquete, M.R.; von Wangenheim, D.; Marhavý, P.; Barbez, E.; Stelzer, E.H.; Benková, E.; Maizel, A.; Kleine-Vehn, J. An Auxin Transport Mechanism Restricts Positive Orthogravitropism in Lateral Roots. Curr. Biol. 2013, 23, 817–822. [Google Scholar] [CrossRef]
- Omelyanchuk, N.A.; Wiebe, D.S.; Novikova, D.D.; Levitsky, V.G.; Klimova, N.; Gorelova, V.; Weinholdt, C.; Vasiliev, G.V.; Zemlyanskaya, E.V.; Kolchanov, N.A.; et al. Auxin regulates functional gene groups in a fold-change-specific manner in Arabidopsis thaliana roots. Sci. Rep. 2017, 7, 2489. [Google Scholar] [CrossRef]
- Stigliani, A.; Martin-Arevalillo, R.; Lucas, J.; Bessy, A.; Vinos-Poyo, T.; Mironova, V.; Vernoux, T.; Dumas, R.; Parcy, F. Capturing Auxin Response Factors Syntax Using DNA Binding Models. Mol. Plant 2018, 12, 822–832. [Google Scholar] [CrossRef] [PubMed]
- Waite, J.M.; Collum, T.D.; Dardick, C. AtDRO1 is nuclear localized in root tips under native conditions and impacts auxin localization. Plant Mol. Biol. 2020, 103, 197–210. [Google Scholar] [CrossRef] [PubMed]
- Ge, L.; Chen, R. Negative gravitropic response of roots directs auxin flow to control root gravitropism. Plant Cell Environ. 2019, 42, 2372–2383. [Google Scholar] [CrossRef]
- Wardhani, T.A.; Roswanjaya, Y.P.; Dupin, S.; Li, H.; Linders, S.; Hartog, M.; Geurts, R.; Van Zeijl, A. Transforming, Genome Editing and Phenotyping the Nitrogen-fixing Tropical Cannabaceae Tree Parasponia andersonii. J. Vis. Exp. 2019, 150, e59971. [Google Scholar] [CrossRef]
- Ma, X.; Zhu, Q.; Chen, Y.; Liu, Y.-G. CRISPR/Cas9 Platforms for Genome Editing in Plants: Developments and Applications. Mol. Plant 2016, 9, 961–974. [Google Scholar] [CrossRef] [PubMed]
- Shan, S.; Mavrodiev, E.V.; Li, R.; Zhang, Z.; Hauser, B.A.; Soltis, P.S.; Soltis, D.E.; Yang, B. Application of CRISPR/Cas9 to Tragopogon (Asteraceae), an evolutionary model for the study of polyploidy. Mol. Ecol. Resour. 2018, 18, 1427–1443. [Google Scholar] [CrossRef] [PubMed]
- Aoki, H.; Teramura, H.; Schepetilnikov, M.; Ryabova, L.A.; Kusano, H.; Shimada, H. Enhanced translation of the downstream ORF attributed to a long 5′ untranslated region in the OsMac1 gene family members, OsMac2 and OsMac3. Plant Biotechnol. 2014, 31, 221–228. [Google Scholar] [CrossRef]
- Onodera, H.; Shingu, S.; Ohnuma, M.; Horie, T.; Kihira, M.; Kusano, H.; Teramura, H.; Shimada, H. Establishment of a conditional TALEN system using the translational enhancer dMac3 and an inducible promoter activated by glucocorticoid treatment to increase the frequency of targeted mutagenesis in plants. PLoS ONE 2018, 13, e0208959. [Google Scholar] [CrossRef]
- Kusano, H.; Ohnuma, M.; Mutsuro-Aoki, H.; Asahi, T.; Ichinosawa, D.; Onodera, H.; Asano, K.; Noda, T.; Horie, T.; Fukumoto, K.; et al. Establishment of a modified CRISPR/Cas9 system with increased mutagenesis frequency using the translational enhancer dMac3 and multiple guide RNAs in potato. Sci. Rep. 2018, 8, 13753. [Google Scholar] [CrossRef]
- Kusano, H.; Takeuchi, A.; Shimada, H. Efficiency of potato genome editing: Targeted mutation on the genes involved in starch biosynthesis using the CRISPR/dMac3-Cas9 system. Plant Biotechnol. 2023, 40, 201–209. [Google Scholar] [CrossRef]
- Bullock, W.O.; Fernandez, J.M.; Short, J.M. XL1-blue: A high efficiency plasmid transforming recA Escherichia coli strain with β-galactosidase selection. BioTechniques 1987, 5, 376–379. [Google Scholar]
- White, F.F.; Taylor, B.H.; Huffman, G.A.; Gordon, M.P.; Nester, E.W. Molecular and genetic analysis of the transferred DNA regions of the root-inducing plasmid of Agrobacterium rhizogenes. J. Bacteriol. 1985, 164, 33–44. [Google Scholar] [CrossRef]
- Owens, L.D.; Cress, D.E. Genotypic Variability of Soybean Response to Agrobacterium Strains Harboring the Ti or Ri Plasmids. Plant Physiol. 1985, 77, 87–94. [Google Scholar] [CrossRef]
- Berardini, T.Z.; Reiser, L.; Li, D.; Mezheritsky, Y.; Muller, R.; Strait, E.; Huala, E. The arabidopsis information resource: Making and mining the “gold standard” annotated reference plant genome. Genesis 2015, 53, 474–485. [Google Scholar] [CrossRef]
- Goodstein, D.M.; Shu, S.; Howson, R.; Neupane, R.; Hayes, R.D.; Fazo, J.; Mitros, T.; Dirks, W.; Hellsten, U.; Putnam, N.; et al. Phytozome: A comparative platform for green plant genomics. Nucleic Acids Res. 2012, 40, D1178–D1186. [Google Scholar] [CrossRef] [PubMed]
- Zheng, Y.; Wu, S.; Bai, Y.; Sun, H.; Jiao, C.; Guo, S.; Zhao, K.; Blanca, J.; Zhang, Z.; Huang, S.; et al. Cucurbit Genomics Database (CuGenDB): A central portal for comparative and functional genomics of cucurbit crops. Nucleic Acids Res. 2019, 47, D1128–D1136. [Google Scholar] [CrossRef] [PubMed]
- Clark, K.; Karsch-Mizrachi, I.; Lipman, D.J.; Ostell, J.; Sayers, E.W. GenBank. Nucleic Acids Res. 2016, 44, D67–D72. [Google Scholar] [CrossRef]
- Sievers, F.; Wilm, A.; Dineen, D.; Gibson, T.J.; Karplus, K.; Li, W.; Lopez, R.; McWilliam, H.; Remmert, M.; Söding, J.; et al. Fast, scalable generation of high-quality protein multiple sequence alignments using Clustal Omega. Mol. Syst. Biol. 2011, 7, 539. [Google Scholar] [CrossRef] [PubMed]
- Kumar, S.; Stecher, G.; Tamura, K. MEGA7: Molecular Evolutionary Genetics Analysis Version 7.0 for Bigger Datasets. Mol. Biol. Evol. 2016, 33, 1870–1874. [Google Scholar] [CrossRef] [PubMed]
- Yang, Z. Maximum likelihood phylogenetic estimation from DNA sequences with variable rates over sites: Approximate methods. J. Mol. Evol. 1994, 39, 306–314. [Google Scholar] [CrossRef]
- Jones, D.T.; Taylor, W.R.; Thornton, J.M. The rapid generation of mutation data matrices from protein sequences. Comput. Appl. Biosci. 1992, 8, 275–282. [Google Scholar] [CrossRef]
- Shoemaker, J.S.; Fitch, W.M. Evidence from nuclear sequences that invariable sites should be considered when sequence divergence is calculated. Mol. Biol. Evol. 1989, 6, 270–289. [Google Scholar] [CrossRef]
- Kiryushkin, A.S.; Ilina, E.L.; Puchkova, V.A.; Guseva, E.D.; Pawlowski, K.; Demchenko, K.N. Lateral Root Initiation in the Parental Root Meristem of Cucurbits: Old Players in a New Position. Front. Plant Sci. 2019, 10, 365. [Google Scholar] [CrossRef] [PubMed]
- Camp, R.H.O.D.; De Mita, S.; Lillo, A.; Cao, Q.; Limpens, E.; Bisseling, T.; Geurts, R. A Phylogenetic Strategy Based on a Legume-Specific Whole Genome Duplication Yields Symbiotic Cytokinin Type-A Response Regulators. Plant Physiol. 2011, 157, 2013–2022. [Google Scholar] [CrossRef] [PubMed]
- Limpens, E.; Ramos, J.; Franken, C.; Raz, V.; Compaan, B.; Franssen, H.; Bisseling, T.; Geurts, R. RNA interference in Agrobacterium rhizogenes-transformed roots of Arabidopsis and Medicago truncatula. J. Exp. Bot. 2004, 55, 983–992. [Google Scholar] [CrossRef] [PubMed]
- Xie, M.; He, Y.; Gan, S. Bidirectionalization of polar promoters in plants. Nat. Biotechnol. 2001, 19, 677–679. [Google Scholar] [CrossRef]
- Wang, Q.; Wan, L.; Li, D.; Zhu, L.; Qian, M.; Deng, M. Searching for bidirectional promoters in Arabidopsis thaliana. BMC Bioinform. 2009, 10, S29. [Google Scholar] [CrossRef] [PubMed]
- Untergasser, A. Cloning—Multiple Gateway Reaction II without a Mastermix. Available online: http://www.untergasser.de/lab/protocols/lr_multiple_gateway_reaction_ii_v1_0.htm (accessed on 13 December 2023).
- Kiryushkin, A.S.; Ilina, E.L.; Guseva, E.D.; Pawlowski, K.; Demchenko, K.N. Hairy CRISPR: Genome Editing in Plants Using Hairy Root Transformation. Plants 2022, 11, 51. [Google Scholar] [CrossRef]
- Xing, H.-L.; Dong, L.; Wang, Z.-P.; Zhang, H.-Y.; Han, C.-Y.; Liu, B.; Wang, X.-C.; Chen, Q.-J. A CRISPR/Cas9 toolkit for multiplex genome editing in plants. BMC Plant Biol. 2014, 14, 327. [Google Scholar] [CrossRef]
- Gibson, D.G.; Young, L.; Chuang, R.-Y.; Venter, J.C.; Hutchison, C.A., III; Smith, H.O. Enzymatic assembly of DNA molecules up to several hundred kilobases. Nat. Methods 2009, 6, 343–345. [Google Scholar] [CrossRef]
- New England Biolabs. Gibson Assembly® Protocol (E5510). Available online: https://international.neb.com/protocols/2012/12/11/gibson-assembly-protocol-e5510 (accessed on 13 December 2023).
- Haeussler, M.; Schönig, K.; Eckert, H.; Eschstruth, A.; Mianné, J.; Renaud, J.-B.; Schneider-Maunoury, S.; Shkumatava, A.; Teboul, L.; Kent, J.; et al. Evaluation of off-target and on-target scoring algorithms and integration into the guide RNA selection tool CRISPOR. Genome Biol. 2016, 17, 148. [Google Scholar] [CrossRef] [PubMed]
- Minkenberg, B.; Zhang, J.; Xie, K.; Yang, Y. CRISPR-PLANT v2: An online resource for highly specific guide RNA spacers based on improved off-target analysis. Plant Biotechnol. J. 2019, 17, 5–8. [Google Scholar] [CrossRef] [PubMed]
- Kerpedjiev, P.; Hammer, S.; Hofacker, I.L. Forna (force-directed RNA): Simple and effective online RNA secondary structure diagrams. Bioinformatics 2015, 31, 3377–3379. [Google Scholar] [CrossRef] [PubMed]
- Ilina, E.L.; Logachov, A.A.; Laplaze, L.; Demchenko, N.P.; Pawlowski, K.; Demchenko, K.N. Composite Cucurbita pepo plants with transgenic roots as a tool to study root development. Ann. Bot. 2012, 110, 479–489. [Google Scholar] [CrossRef] [PubMed]
- Ilina, E.I.; Kiryushkin, A.S.; Demchenko, K.N. Features of Fluorescent Protein Application to Study the Root System Development of Cucurbits (Cucurbitaceae). Agric. Biol. (Sel’skokhozyaistvennaya Biol.) 2020, 55, 1040–1055. [Google Scholar] [CrossRef]
- Kiryushkin, A.S.; Ilina, E.L.; Guseva, E.D.; Pawlowski, K.; Demchenko, K.N. Lateral Root Initiation in Cucumber (Cucumis sativus): What Does the Expression Pattern of Rapid Alkalinization Factor 34 (RALF34) Tell Us? Int. J. Mol. Sci. 2023, 24, 8440. [Google Scholar] [CrossRef]
- Stumpe, M.; Göbel, C.; Demchenko, K.; Hoffmann, M.; Klösgen, R.B.; Pawlowski, K.; Feussner, I. Identification of an allene oxide synthase (CYP74C) that leads to formation of α-ketols from 9-hydroperoxides of linoleic and linolenic acid in below-ground organs of potato. Plant J. 2006, 47, 883–896. [Google Scholar] [CrossRef]
- Chomczynski, P.; Mackey, K.; Drews, R.; Wilfinger, W. DNAzol®: A Reagent for the Rapid Isolation of Genomic DNA. BioTechniques 1997, 22, 550–553. [Google Scholar] [CrossRef]
- Hoagland, D.R.; Arnon, D.I. The Water-Culture Method for Growing Plants without Soil; University of California, College of Agriculture, Agricultural Experiment Station: Berkeley, CA, USA, 1938; pp. 1–40. [Google Scholar]
- R Core Team. R: A Language and Environment for Statistical Computing. 2020; R Foundation for Statistical Computing: Vienna, Austria. Available online: https://www.R-project.org/ (accessed on 13 December 2023).
- RStudio Team. RStudio: Integrated Development Environment for R. Available online: https://posit.co/ (accessed on 13 December 2023).
- Stuttmann, J.; Barthel, K.; Martin, P.; Ordon, J.; Erickson, J.L.; Herr, R.; Ferik, F.; Kretschmer, C.; Berner, T.; Keilwagen, J.; et al. Highly efficient multiplex editing: One-shot generation of 8× Nicotiana benthamiana and 12× Arabidopsis mutants. Plant J. 2021, 106, 8–22. [Google Scholar] [CrossRef]
- Gogolev, Y.V.; Ahmar, S.; Akpinar, B.A.; Budak, H.; Kiryushkin, A.S.; Gorshkov, V.Y.; Hensel, G.; Demchenko, K.N.; Kovalchuk, I.; Mora-Poblete, F.; et al. OMICs, Epigenetics, and Genome Editing Techniques for Food and Nutritional Security. Plants 2021, 10, 1423. [Google Scholar] [CrossRef] [PubMed]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Protein Names (Gene IDs of C. sativus cv Chinese Long v. 2) | Similarity (Identity) with AtDRO1 Protein | Similarity (Identity) with AtDRO2 Protein | Similarity (Identity) with AtDRO3 Protein |
|---|---|---|---|
| CsDRO1 (Csa7G448840) | 63.5% (51.5%) | 35.9% (27.4%) | 64.5% (52%) |
| CsDRO1-LIKE1 (CsDRO1L1, Csa1G597130) | 43.7% (32.5%) | 30.5% (23.9%) | 43.5% (31.2%) |
| CsDRO1-LIKE2 (CsDRO1L2, Csa3G645860) | 25.7% (15%) | 25.4% (16%) | 27% (17%) |
| Protein Names (Gene IDs of C. sativus cv Chinese Long v. 2) | Similarity (Identity) with OsDRO1 Protein | Similarity (Identity) with OsDRL1 Protein | Similarity (Identity) with OsDRL2 Protein | Similarity (Identity) with OsDRL3 Protein |
|---|---|---|---|---|
| CsDRO1 (Csa7G448840) | 51.7% (37.2%) | 48.2% (36.2%) | 31.5% (18.9%) | 36.9% (29.4%) |
| CsDRO1-LIKE1 (CsDRO1L1, Csa1G597130) | 45.3% (32.5%) | 43% (27%) | 31% (20.4%) | 32.1% (22.2%) |
| CsDRO1-LIKE2 (CsDRO1L2, Csa3G645860) | 22% (13%) | 24% (13.2%) | 24.3% (10.8%) | 20.3% (9.3%) |
| Gene Name | The Position of Cloning Region before the Predicted Translational Start Site | Size of Putative Promoter, bp |
|---|---|---|
| CsDRO1 | from −3295 bp to −5 bp | 3291 |
| CsDRO1L1 | from −808 bp to −1 bp | 808 |
| CsDRO1L2 | from −2965 bp to −1 bp | 2965 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kiryushkin, A.S.; Ilina, E.L.; Kiikova, T.Y.; Pawlowski, K.; Demchenko, K.N. Do DEEPER ROOTING 1 Homologs Regulate the Lateral Root Slope Angle in Cucumber (Cucumis sativus)? Int. J. Mol. Sci. 2024, 25, 1975. https://doi.org/10.3390/ijms25041975
Kiryushkin AS, Ilina EL, Kiikova TY, Pawlowski K, Demchenko KN. Do DEEPER ROOTING 1 Homologs Regulate the Lateral Root Slope Angle in Cucumber (Cucumis sativus)? International Journal of Molecular Sciences. 2024; 25(4):1975. https://doi.org/10.3390/ijms25041975
Chicago/Turabian StyleKiryushkin, Alexey S., Elena L. Ilina, Tatyana Y. Kiikova, Katharina Pawlowski, and Kirill N. Demchenko. 2024. "Do DEEPER ROOTING 1 Homologs Regulate the Lateral Root Slope Angle in Cucumber (Cucumis sativus)?" International Journal of Molecular Sciences 25, no. 4: 1975. https://doi.org/10.3390/ijms25041975