
Exploring the Regulatory Dynamics of BrFLC-Associated lncRNA in Modulating the Flowering Response of Chinese Cabbage
 , ,
, , {kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results
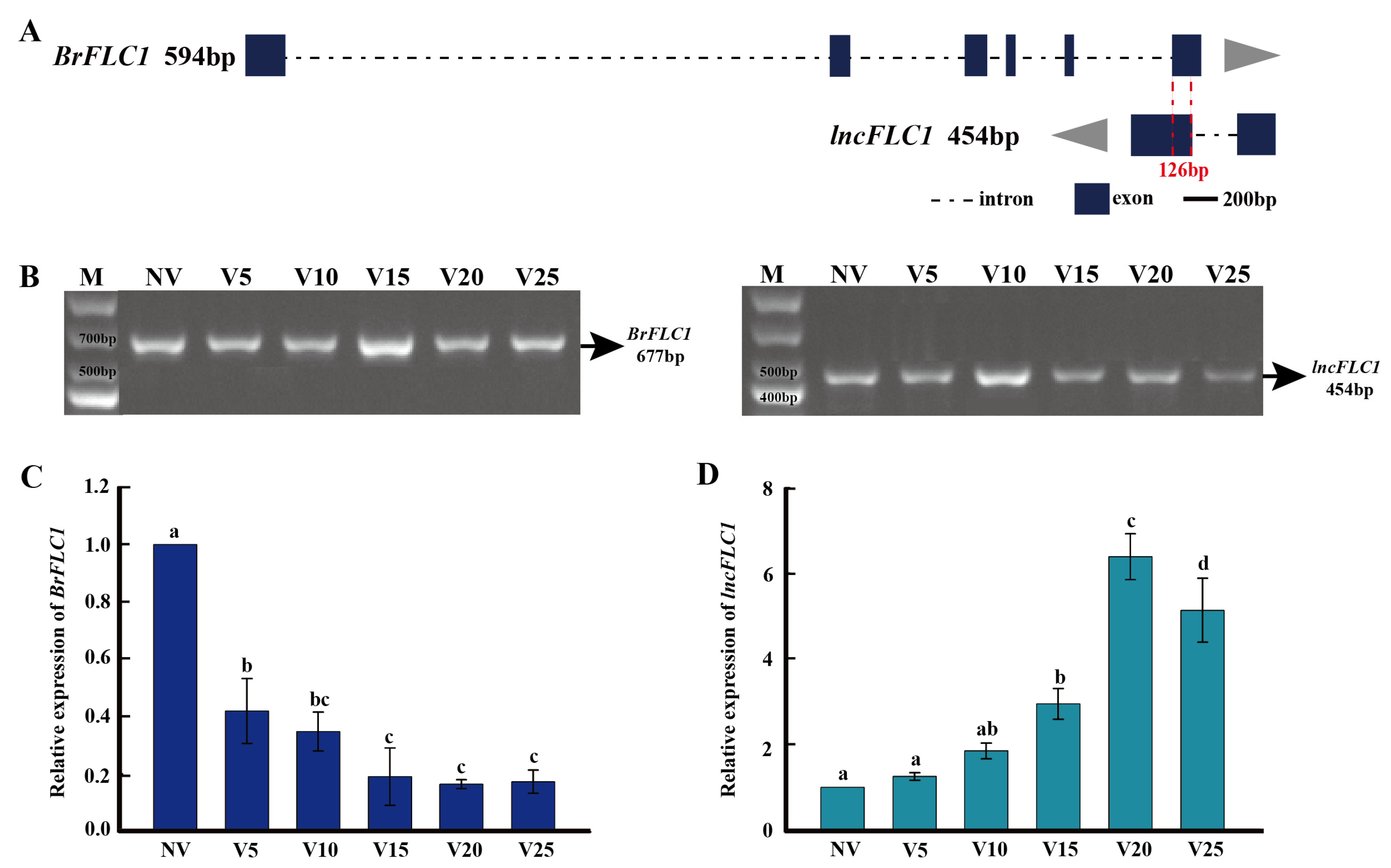
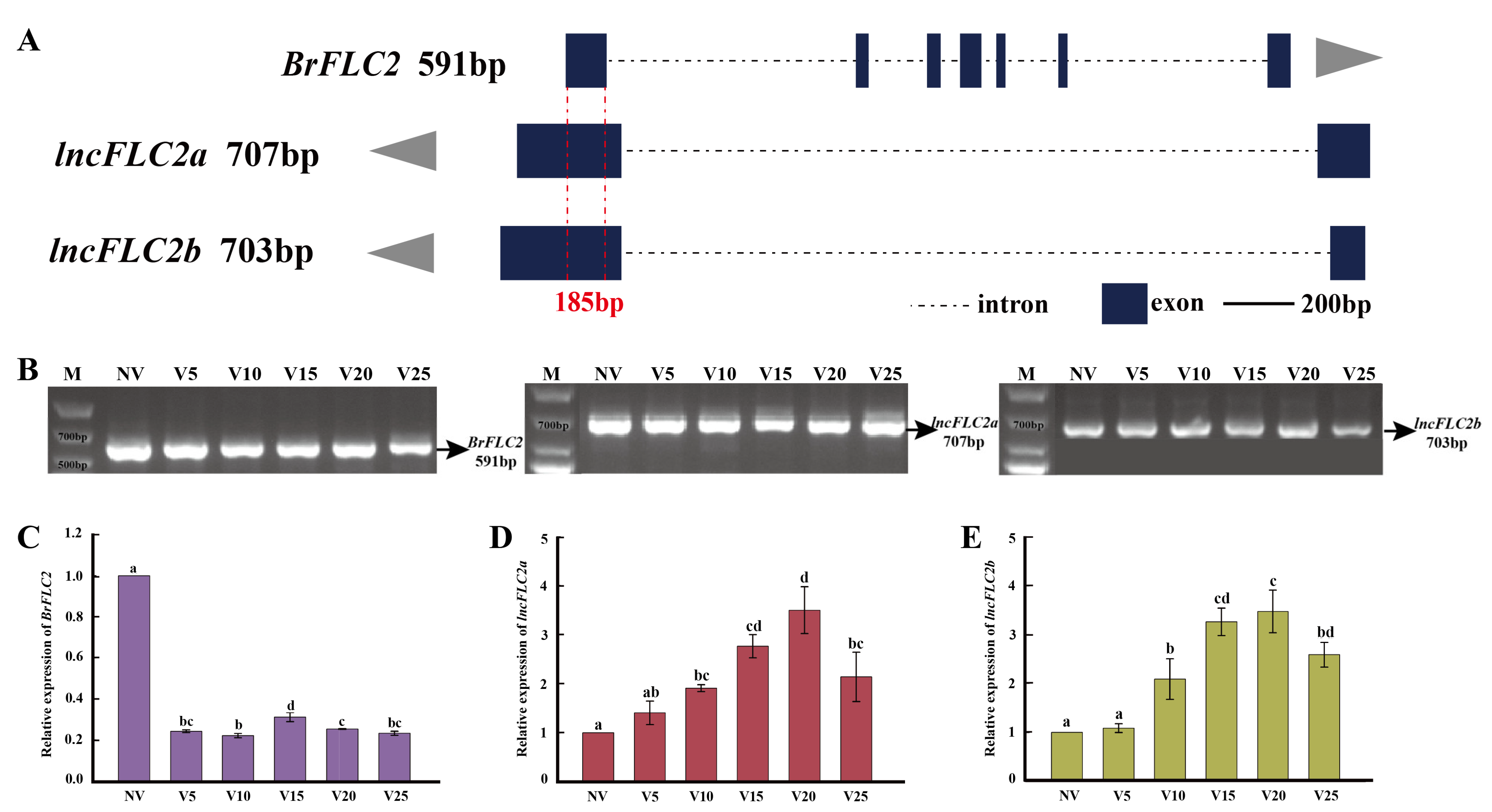
2.1. Identification of lncFLC1, lncFLC2a and lncFLC2b in Chinese Cabbage
2.2. Vernalization-Induced Upregulation of lncFLC1, lncFLC2a, and lncFLC2b
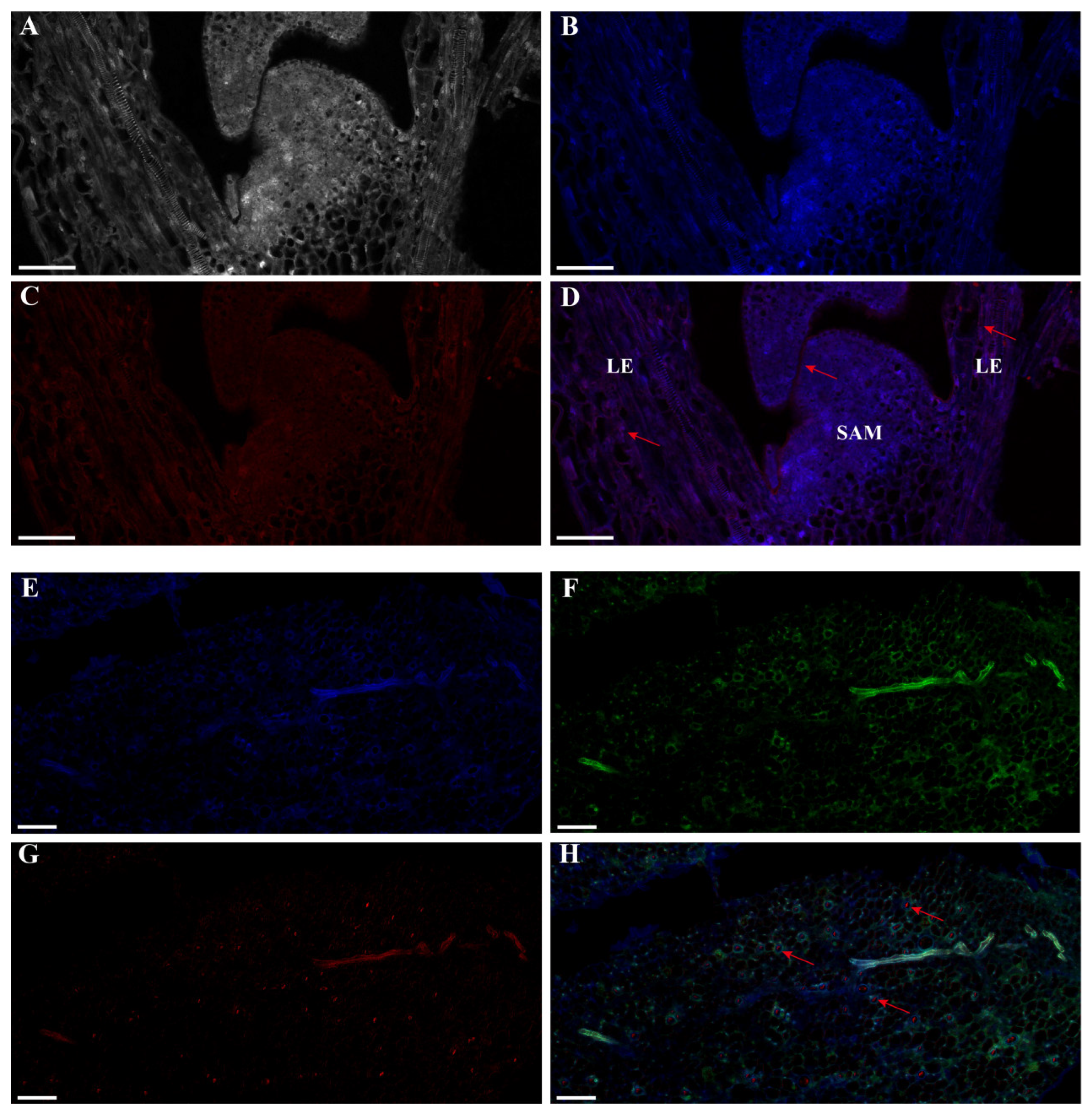
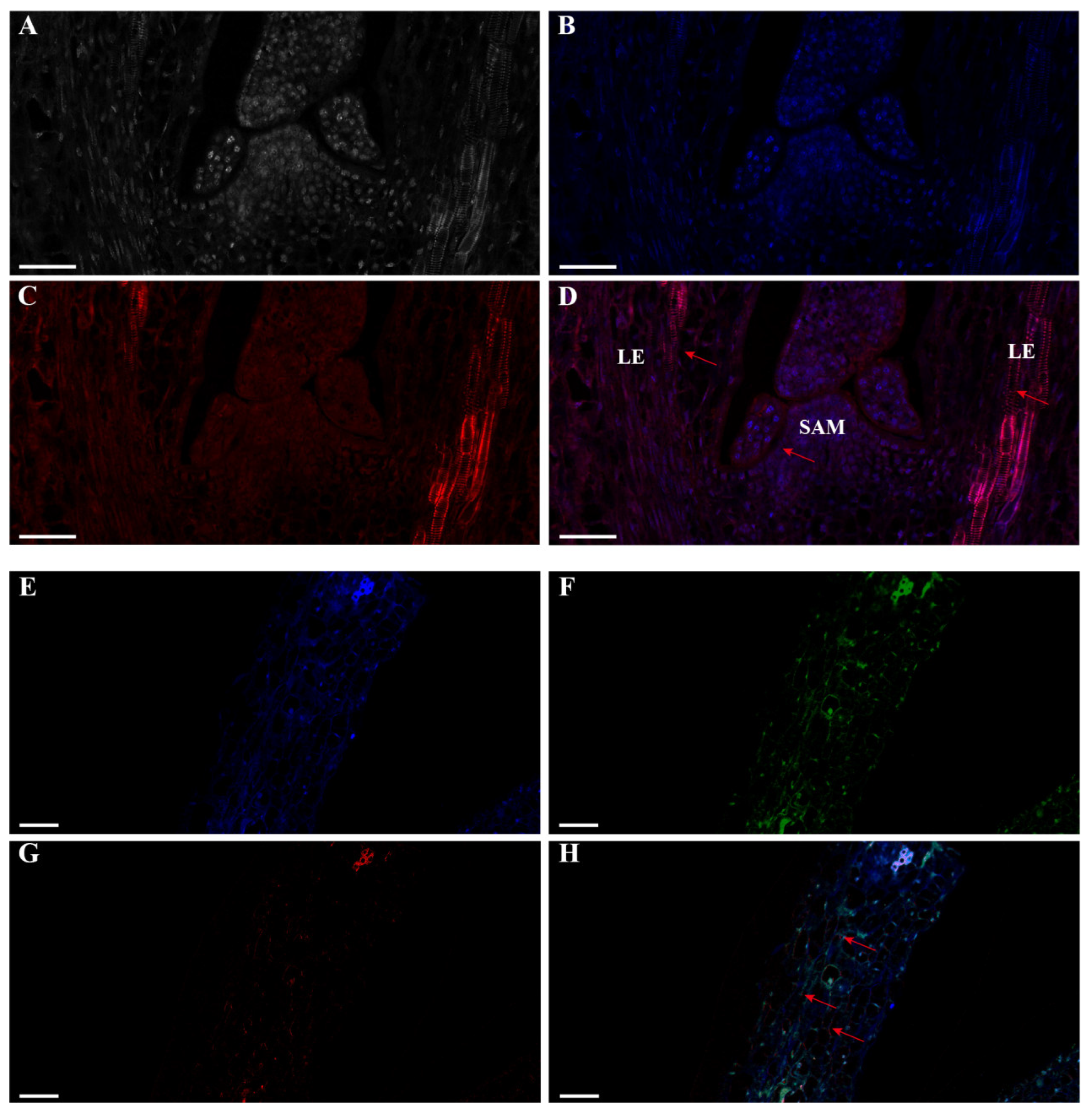
2.3. Expression Profiles of lncFLC1 and lncFLC2
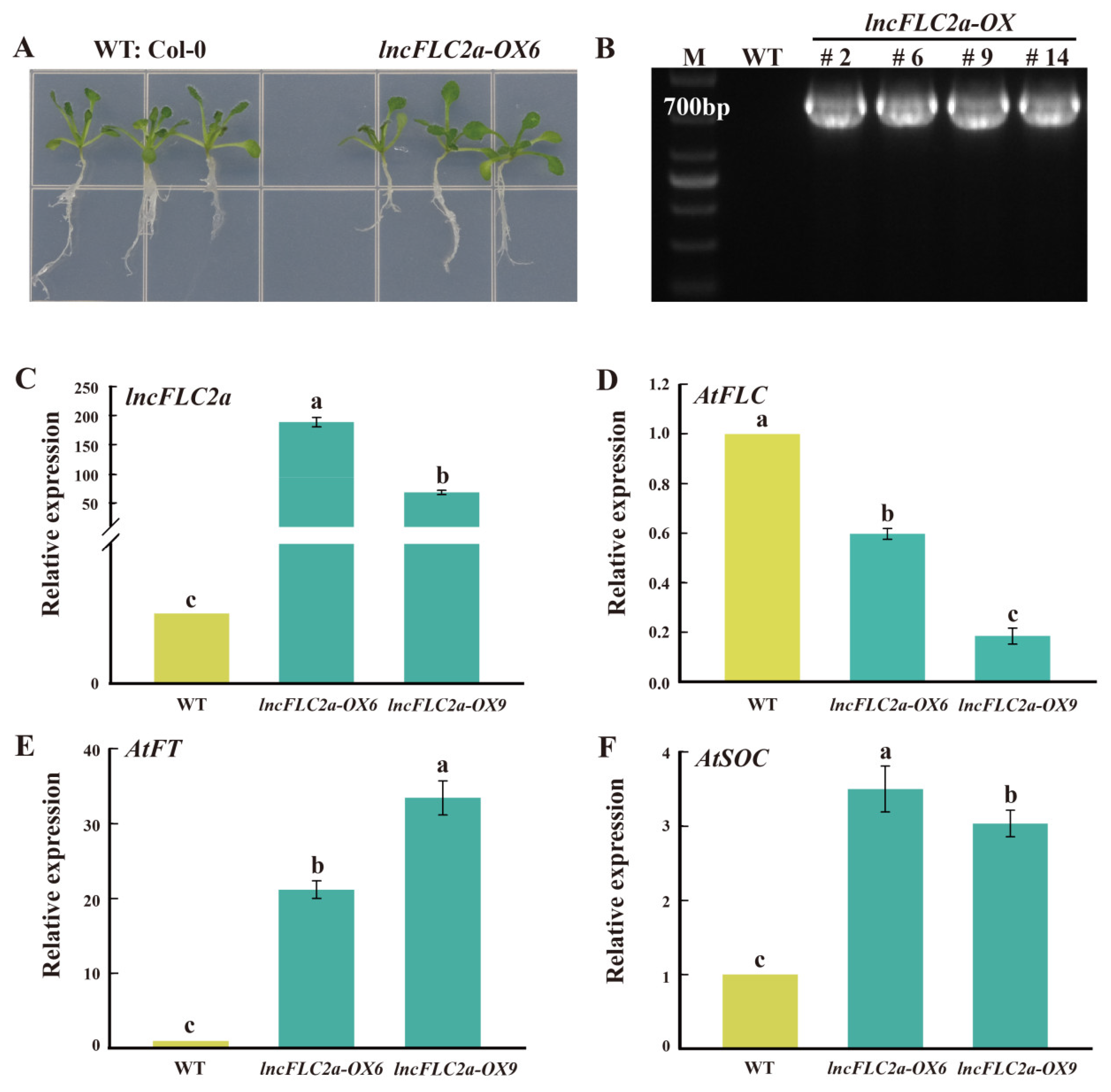
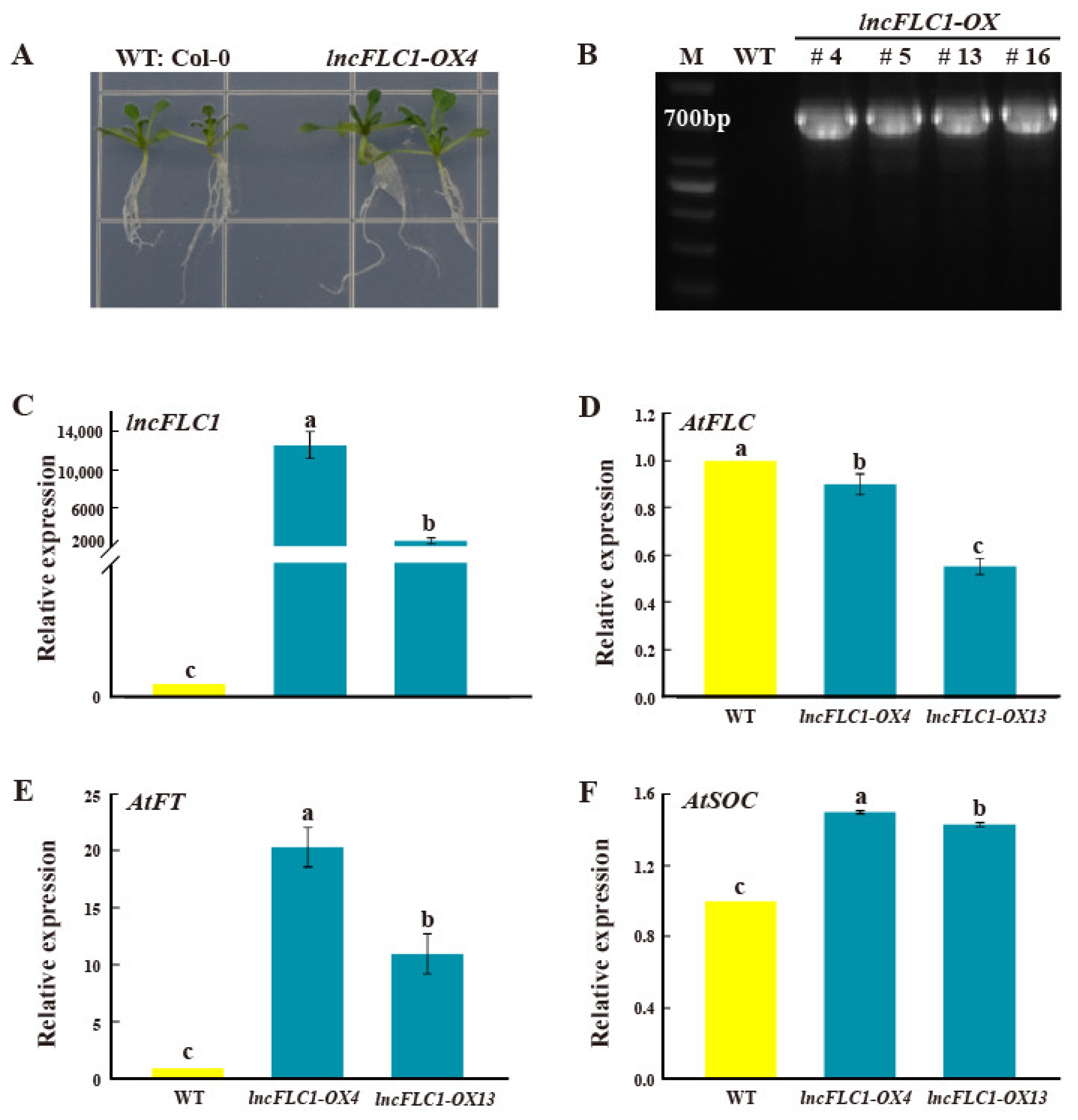
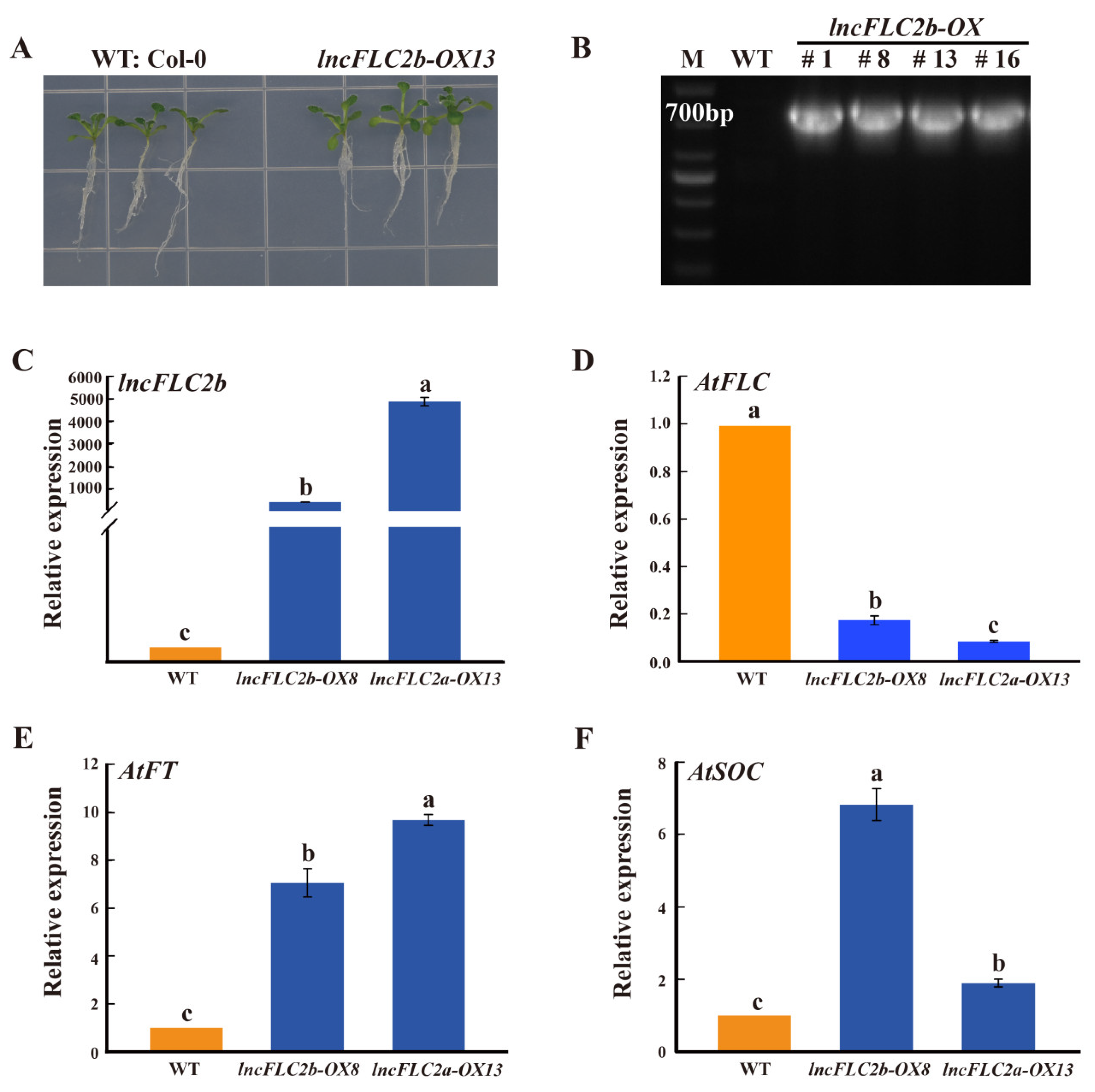
2.4. Expression Patterns of lncFLC1, lncFLC2a and lncFLC2b in Arabidopsis Transgenic Lines
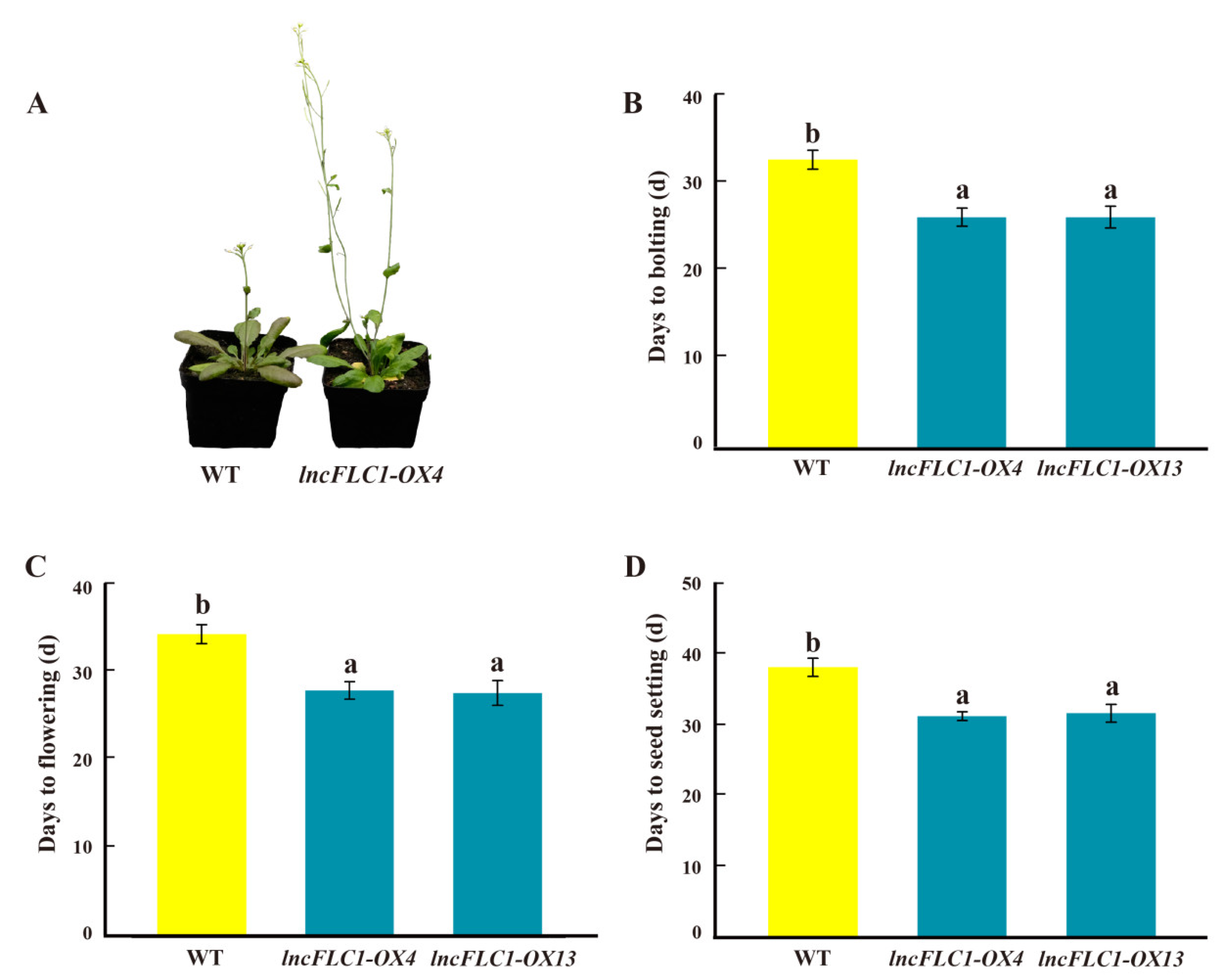
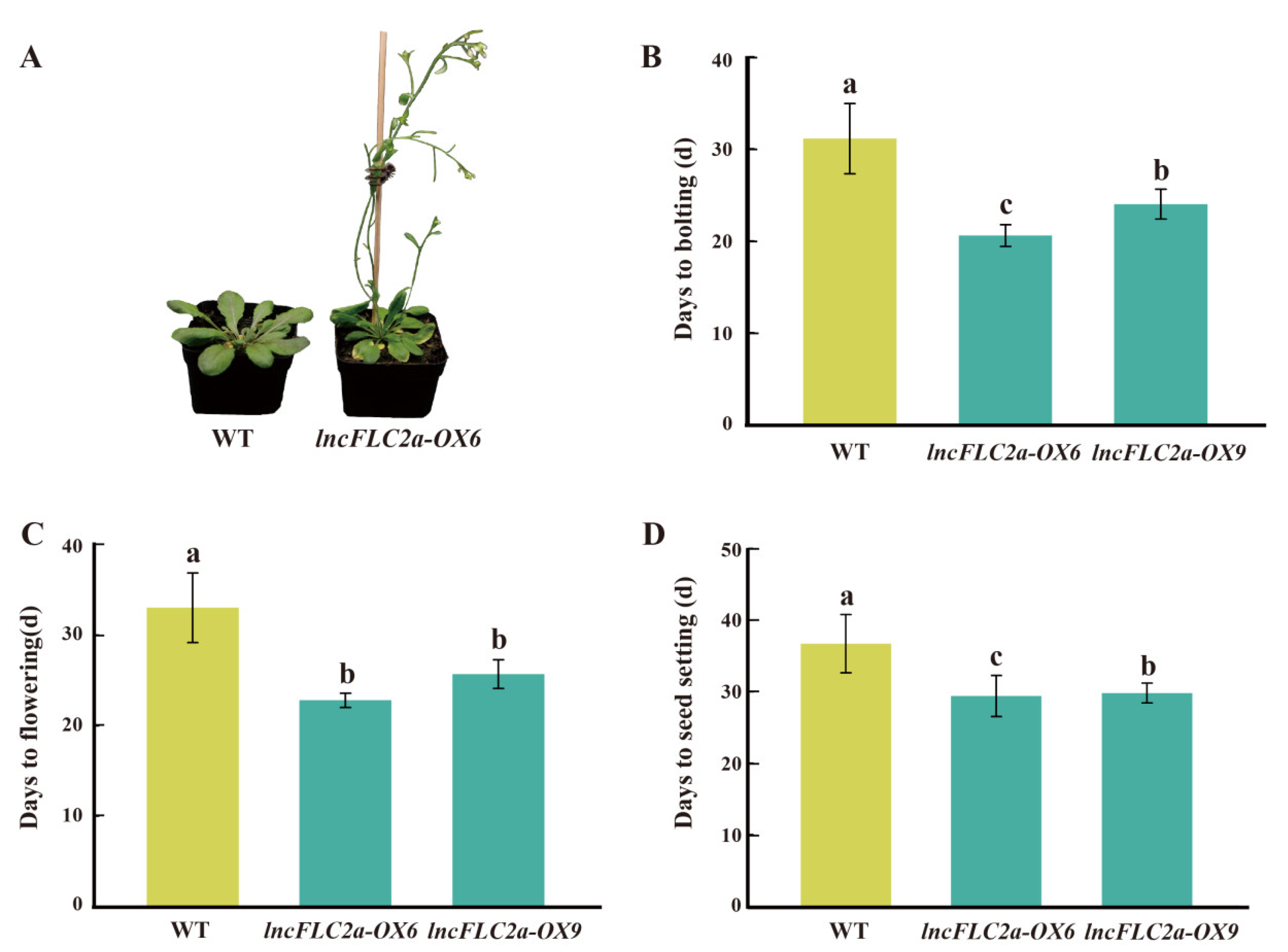
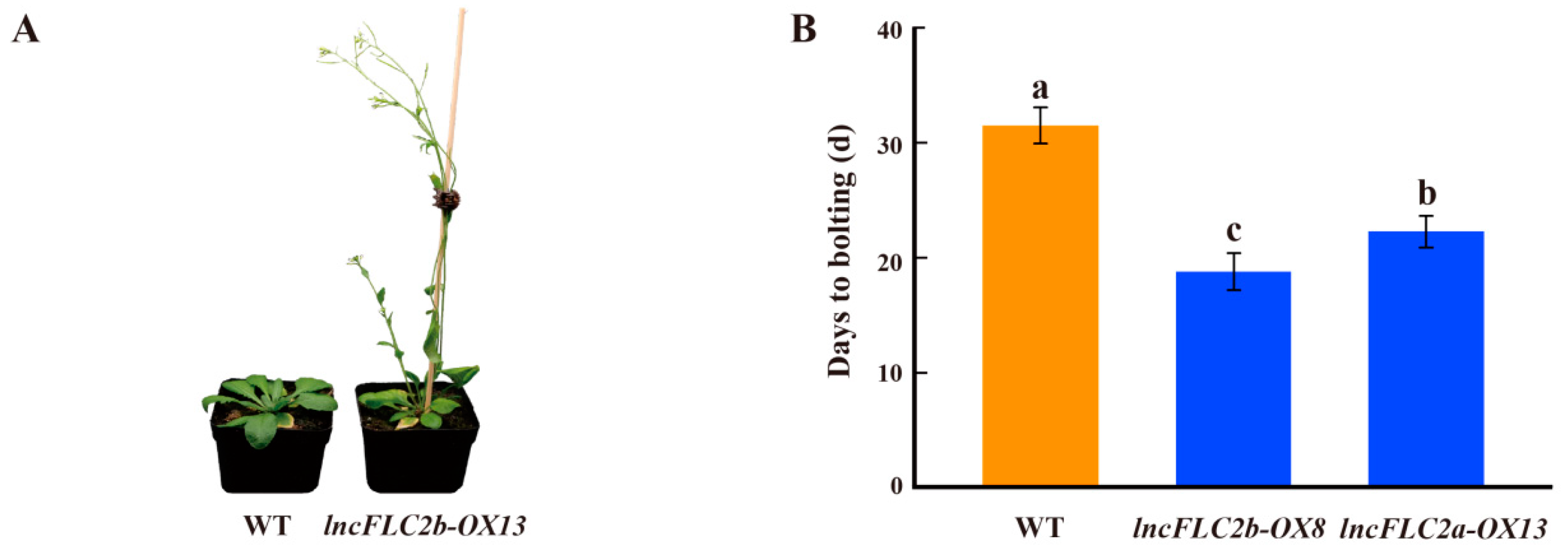
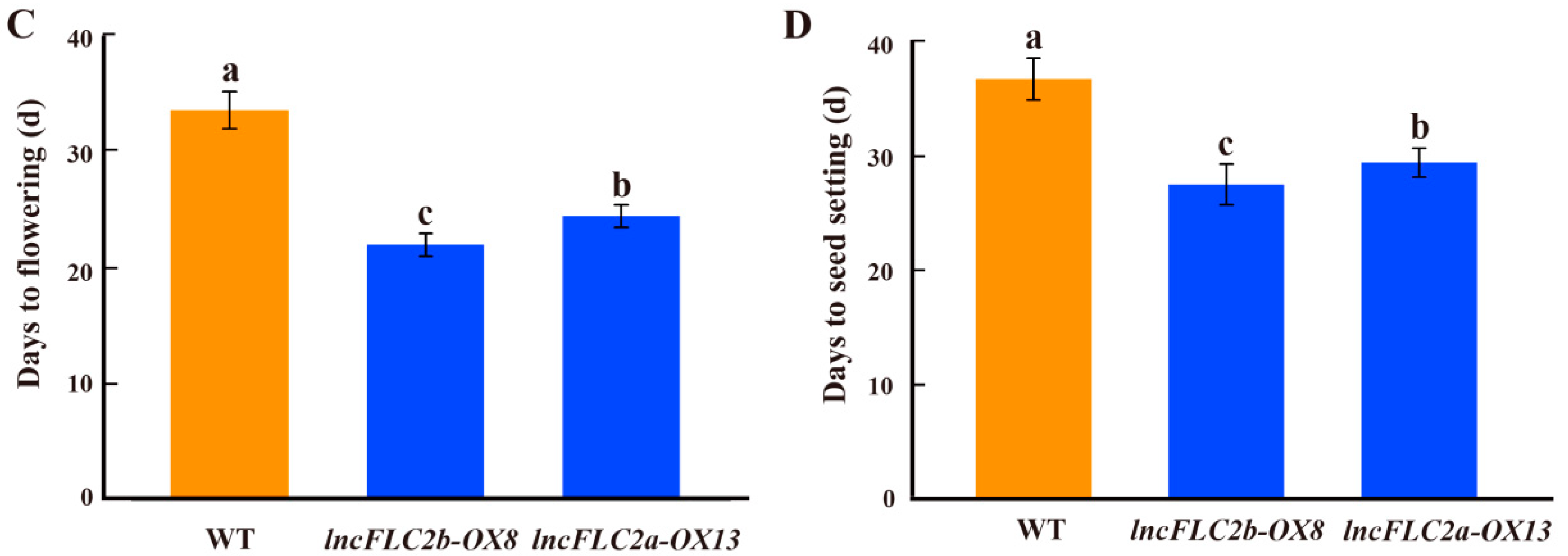
2.5. Promotion of Early Flowering in Arabidopsis by Overexpression of lncFLC1, lncFLC2a and lncFLC2b
3. Discussion
4. Materials and Methods
4.1. Plant Material and Growth Conditions
4.2. RNA Fluorescence In Situ Hybridization (FISH) Technique
4.3. Construction of Plasmids and Generation of Transgenic Arabidopsis Lines
4.4. Phenotypic Investigation of Transgenic Arabidopsis Plants
4.5. Gene Expression Analysis
4.6. Statistical Analysis
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Chekanova, J.A. Long non-coding RNAs and their functions in plants. Curr. Opin. Plant Biol. 2015, 27, 207–216. [Google Scholar] [CrossRef] [PubMed]
- Yamada, M. Functions of long intergenic non-coding (linc) RNAs in plants. J. Plant Res. 2017, 130, 67–73. [Google Scholar] [CrossRef] [PubMed]
- Das, P.; Agarwala, N.; Gill, S.S.; Varshney, R.K. Emerging role of plant long non coding RNAs (lncRNAs) in salinity stress response. Plant Stress 2023, 10, 93–105. [Google Scholar] [CrossRef]
- Mattick, J.S. The genetic signatures of noncoding RNAs. PLoS Genet. 2009, 5, e1000459. [Google Scholar] [CrossRef]
- Spizzo, R.; Almeida, M.I.; Colombatti, A.; Calin, G.A. Long non-coding RNAs and cancer: A new frontier of translational research? Oncogene 2012, 31, 4577–4587. [Google Scholar] [CrossRef] [PubMed]
- Dykes, I.M.; Emanueli, C. Transcriptional and post-transcriptional gene regulation by long non-coding RNA. Genom. Proteom. Bioinf. 2017, 15, 177–186. [Google Scholar] [CrossRef]
- Jin, Y.; Ivanov, M.; Dittrich, A.N.; Nelson, A.D.L.; Marquardt, S. LncRNA flail affects alternative splicing and represses flowering in Arabidopsis. Embo J. 2023, 42, e110921. [Google Scholar] [CrossRef]
- Shin, W.J.; Nam, A.H.; Kim, J.Y.; Kwak, J.S.; Song, J.T.; Seo, H.S. Intronic long noncoding RNA, RICE FLOWERING ASSOCIATED (RIFLA) regulates OsMADS56-mediated flowering in rice. Plant Sci. 2022, 320, 111278. [Google Scholar] [CrossRef]
- Zhou, H.L.; Yan, R.Z.; He, H.; Wei, X.L.; Liu, S.C.; Guo, B.T.; Zhang, Y.H.; Liu, X.Y.; Rahman, S.U.; Zhou, C.; et al. Transcriptional profiling of long noncoding RNAs associated with flower color formation in ipomoea nil. Planta 2023, 258, 6. [Google Scholar]
- Yamaguchi, A.; Abe, M. Regulation of reproductive development by non-coding RNA in Arabidopsis: To flower or not to flower. J. Plant Res. 2012, 152, 693–704. [Google Scholar] [CrossRef]
- Liu, T.K.; Wu, P.; Wang, Q.; Wang, W.L.; Zhang, C.W.; Sun, F.F.; Liu, Z.K.; Li, Y.; Hou, X.L. Comparative transcriptome discovery and elucidation of the mechanism of long noncoding RNAs during vernalization in Brassica rapa. Plant Growth Regul. 2018, 85, 27–39. [Google Scholar] [CrossRef]
- Severing, E.; Faino, L.; Jamge, S.; Busscher, M.; Kuijer-Zhang, Y.; Bellinazzo, F.; Busscher-Lange, J.; Fernández, V.; Angenent, G.C.; Immink, R.G.H.; et al. Arabidopsis thaliana ambient temperature responsive lncRNAs. BMC Plant Biol. 2018, 18, 145. [Google Scholar] [CrossRef] [PubMed]
- Swiezewski, S.; Liu, F.Q.; Magusin, A.; Dean, C. Cold-induced silencing by long antisense transcripts of an Arabidopsis polycomb target. Nature 2009, 462, 799–802. [Google Scholar] [CrossRef] [PubMed]
- Csorba, T.; Questa, J.I.; Sun, Q.W.; Dean, C. Antisense COOLAIR mediates the coordinated switching of chromatin states at FLC during vernalization. Proc. Natl. Acad. Sci. USA 2014, 111, 16160–16165. [Google Scholar] [CrossRef] [PubMed]
- Heo, J.B.; Sung, S. Vernalization-mediated epigenetic silencing by a long intronic noncoding RNA. Science 2011, 331, 76–79. [Google Scholar] [CrossRef] [PubMed]
- Henriques, R.; Wang, H.; Liu, J.; Boix, M.; Huang, L.F.; Chua, N.H. The antiphasic regulatory module comprising CDF5 and its antisense RNA FLORE links the circadian clock to photoperiodic flowering. New Phytol. 2017, 3, 854–867. [Google Scholar] [CrossRef] [PubMed]
- Dai, Y.; Li, G.L.; Gao, X.Y.; Wang, S.X.; Li, Z.; Song, C.; Zhang, S.F.; Li, F.; Fang, Z.Y.; Sun, R.F.; et al. Identification of long noncoding RNAs involved in plumule-vernalization of Chinese cabbage. Front. Plant Sci. 2023, 14, 1147494. [Google Scholar] [CrossRef]
- Zhao, M.G.; Wang, T.Z.; Sun, T.Y.; Yu, X.X.; Tian, R.; Zhang, W.H. Identification of tissue-specific and cold-responsive lncRNAs in Medicago truncatula by high-throughput RNA sequencing. BMC Plant Biol. 2020, 20, 230. [Google Scholar] [CrossRef]
- Ding, Z.H.; Tie, W.W.; Fu, L.L.; Yan, Y.; Liu, G.H.; Yan, W.; Li, Y.A.; Wu, C.L.; Zhang, J.M.; Hu, W. Strand-specific RNA-seq based identification and functional prediction of drought-responsive lncRNAs in cassava. BMC Genomics 2019, 20, 214. [Google Scholar] [CrossRef]
- Jin, J.J.; Ohama, N.; He, X.J.; Wu, H.W.; Chua, N.H. Tissue-specific transcriptomic analysis uncovers potential roles of natural antisense transcripts in Arabidopsis heat stress response. Front. Plant Sci. 2022, 13, 997967. [Google Scholar] [CrossRef]
- Ariel, F.; Romero-Barrios, N.; Jégu, T.; Benhamed, M.; Crespi, M. Battles and hijacks: Noncoding transcription in plants. Trends Plant Sci. 2015, 6, 362–371. [Google Scholar] [CrossRef] [PubMed]
- Pang, J.L.; Zhang, X.; Ma, X.H.; Zhao, J. Spatio-temporal transcriptional dynamics of maize long non-coding RNAs responsive to drought stress. Genes 2019, 2, 138. [Google Scholar] [CrossRef] [PubMed]
- Wang, G.N.; Wang, X.F.; Zhang, Y.; Yang, J.; Li, Z.K.; Wu, L.Z.; Wu, J.H.; Wu, N.; Liu, L.X.; Liu, Z.W.; et al. Dynamic characteristics and functional analysis provide new insights into long non-coding RNA responsive to Verticillium dahliae infection in Gossypium. BMC Plant Biol. 2021, 21, 68. [Google Scholar] [CrossRef]
- Burghardt, L.T.; Runcie, D.E.; Wilczek, A.M.; Cooper, M.D.; Roe, J.L.; Welch, S.M.; Schmitt, J. Fluctuating, warm temperatures decrease the effect of a key floral repressor on flowering time in Arabidopsis thaliana. New Phytol. 2016, 210, 564–576. [Google Scholar] [CrossRef] [PubMed]
- Ortuño-Miquel, S.; Rodríguez-Cazorla, E.; Zavala-Gonzalez, E.A.; Martínez-Laborda, A.; Vera, A. Arabidopsis HUA ENHANCER 4 delays flowering by upregulating the MADS-box repressor genes FLC and MAF4. Sci. Rep. 2019, 9, 1478. [Google Scholar] [CrossRef]
- Kimura, Y.; Aoki, S.; Ando, E.; Kitatsuji, A.; Watanabe, A.; Ohnishi, M.; Takahashi, K.; Inoue, S.; Nakamichi, N.; Tamada, Y.; et al. A Flowering Integrator, SOC1, Affects Stomatal Opening in Arabidopsis thaliana. Plant Cell Physiol. 2015, 56, 640–649. [Google Scholar] [CrossRef]
- Kinmonth-Schultz, H.; Lewandowska-Sabat, A.M.; Imaizumi, T.; Ward, J.K.; Rognli, O.A.; Fjellheim, S. Flowering Times of Wild Arabidopsis Accessions From Across Norway Correlate With Expression Levels of FT, CO, and FLC Genes. Front. Plant Sci. 2021, 12, 747740. [Google Scholar] [CrossRef]
- Quiroz, S.; Yustis, J.C.; Chávez-Hernández, E.C.; Martínez, T.; Sanchez, M.D.; Garay-Arroyo, A.; Alvarez-Buylla, E.R.; García-Ponce, B. Beyond the Genetic Pathways, Flowering Regulation Complexity in Arabidopsis thaliana. Int. J. Mol. Sci. 2021, 22, 5716. [Google Scholar] [CrossRef]
- Li, X.R.; Zhang, S.F.; Bai, J.J.; He, Y.K. Tuning growth cycles of Brassica crops via natural antisense transcripts of BrFLC. Plant Biotechnol. J. 2016, 14, 905–914. [Google Scholar] [CrossRef]
- Shea, D.J.; Nishida, N.; Takada, S.; Itabashi, E.; Takahashi, S.; Akter, A.; Miyaji, N.; Osabe, K.; Mehraj, H.; Shimizu, M.; et al. Long noncoding RNAs in Brassica rapa L. following vernalization. Sci. Rep. 2019, 9, 9302. [Google Scholar] [CrossRef]
- Zhang, Z.C.; Guo, J.; Cai, X.; Li, Y.F.; Xi, X.; Lin, R.M.; Liang, J.L.; Wang, X.W.; Wu, J. Improved reference genome annotation of Brassica rapa by pacific biosciences RNA sequencing. Front. Plant Sci. 2019, 13, 841618. [Google Scholar] [CrossRef] [PubMed]
- Luo, X.; He, Y.H. Experiencing winter for spring flowering: A molecular epigenetic perspective on vernalization. J. Integr. Plant. Biol. 2020, 62, 104–117. [Google Scholar] [CrossRef] [PubMed]
- Schranz, M.E.; Quijada, P.; Sung, S.B.; Lukens, L.; Amasino, R.; Osborn, T.C. Characterization and effects of the replicated flowering time gene FLC in Brassica rapa. Genetics 2002, 162, 1457–1468. [Google Scholar] [CrossRef] [PubMed]
- Xiao, D.; Zhao, J.J.; Hou, X.L.; Basnet, R.K.; Carpio, D.P.D.; Zhang, N.W.; Bucher, J.; Lin, K.; Cheng, F.; Wang, X.W.; et al. The Brassica rapa FLC homologue FLC2 is a key regulator of flowering time, identified through transcriptional co-expression networks. J. Exp. Bot. 2013, 64, 4503–4516. [Google Scholar] [CrossRef] [PubMed]
- Wang, X.Y.; Xu, X.T.; Mo, X.W.; Zhong, L.Y.; Zhang, J.C.; Mo, B.X.; Kuai, B.K. Overexpression of TCP8 delays Arabidopsis flowering through a FLOWERING LOCUS C-dependent pathway. BMC Plant Biolo. 2019, 19, 534. [Google Scholar] [CrossRef] [PubMed]
- Abeyawardana, O.A.J.; Moravec, T.; Krüger, M.; Belz, C.; Gutierrez-Larruscain, D.; Vondráková, Z.; Eliásová, K.; Storchová, H. The FLOWERING LOCUS T LIKE 2–1 gene of chenopodium triggers precocious flowering in Arabidopsis seedlings. Plant Signal. Behav. 2023, 18, 2239420. [Google Scholar] [CrossRef] [PubMed]
- Deng, P.C.; Liu, S.; Nie, X.J.; Weining, S.; Wu, L. Conservation analysis of long non-coding RNAs in plants. Sci. China Life Sci. 2018, 61, 190–198. [Google Scholar] [CrossRef] [PubMed]
- Rai, M.I.; Alam, M.; Lightfoot, D.A.; Gurha, P.; Afzal, A.J. Classification and experimental identification of plant long non-coding RNAs. Genomics 2019, 111, 997–1005. [Google Scholar] [CrossRef]
- Liu, F.Q.; Marquardt, S.; Lister, C.; Swiezewski, S.; Dean, C. Targeted 3′ processing of antisense transcripts triggers Arabidopsis flc chromatin silencing. Science 2010, 327, 94–97. [Google Scholar] [CrossRef]
- Zhao, Y.S.; Zhu, P.; Hepworth, J.; Bloomer, R.; Antoniou-Kourounioti, R.L.; Doughty, J.; Heckmann, A.; Xu, C.Y.; Yang, H.C.; Dean, C. Natural temperature fluctuations promote COOLAIR regulation of FLC. Gene. Dev. 2021, 35, 888–898. [Google Scholar] [CrossRef]
- Tian, Y.K.; Zheng, H.; Zhang, F.; Wang, S.L.; Ji, X.R.; Xu, C.; He, Y.H.; Ding, Y. PRC2 recruitment and H3K27me3 deposition at FLCrequire FCA binding of COOLAIR. Sci. Dev. 2019, 5, eaau7246. [Google Scholar]
- Zhang, X.R.; Henriques, R.; Lin, S.S.; Niu, Q.W.; Chua, N.H. Agrobacterium-mediated transformation of Arabidopsis thaliana using the floral dip method. Nat. Protoc. 2006, 1, 641–646. [Google Scholar] [CrossRef] [PubMed]
- Nolan, T.; Hands, R.E.; Bustin, S.A. Quantification of mRNA using real-time RT-PCR. Nat. Protoc. 2006, 1, 1559–1582. [Google Scholar] [CrossRef] [PubMed]











Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Dai, Y.; Gao, X.; Zhang, S.; Li, F.; Zhang, H.; Li, G.; Sun, R.; Zhang, S.; Hou, X. Exploring the Regulatory Dynamics of BrFLC-Associated lncRNA in Modulating the Flowering Response of Chinese Cabbage. Int. J. Mol. Sci. 2024, 25, 1924. https://doi.org/10.3390/ijms25031924
Dai Y, Gao X, Zhang S, Li F, Zhang H, Li G, Sun R, Zhang S, Hou X. Exploring the Regulatory Dynamics of BrFLC-Associated lncRNA in Modulating the Flowering Response of Chinese Cabbage. International Journal of Molecular Sciences. 2024; 25(3):1924. https://doi.org/10.3390/ijms25031924
Chicago/Turabian StyleDai, Yun, Xinyu Gao, Shifan Zhang, Fei Li, Hui Zhang, Guoliang Li, Rifei Sun, Shujiang Zhang, and Xilin Hou. 2024. "Exploring the Regulatory Dynamics of BrFLC-Associated lncRNA in Modulating the Flowering Response of Chinese Cabbage" International Journal of Molecular Sciences 25, no. 3: 1924. https://doi.org/10.3390/ijms25031924