
Genome-Wide Identification of DUF668 Gene Family and Expression Analysis under F. solani, Chilling, and Waterlogging Stresses in Zingiber officinale

 ,
,
Abstract
:1. Introduction
2. Results
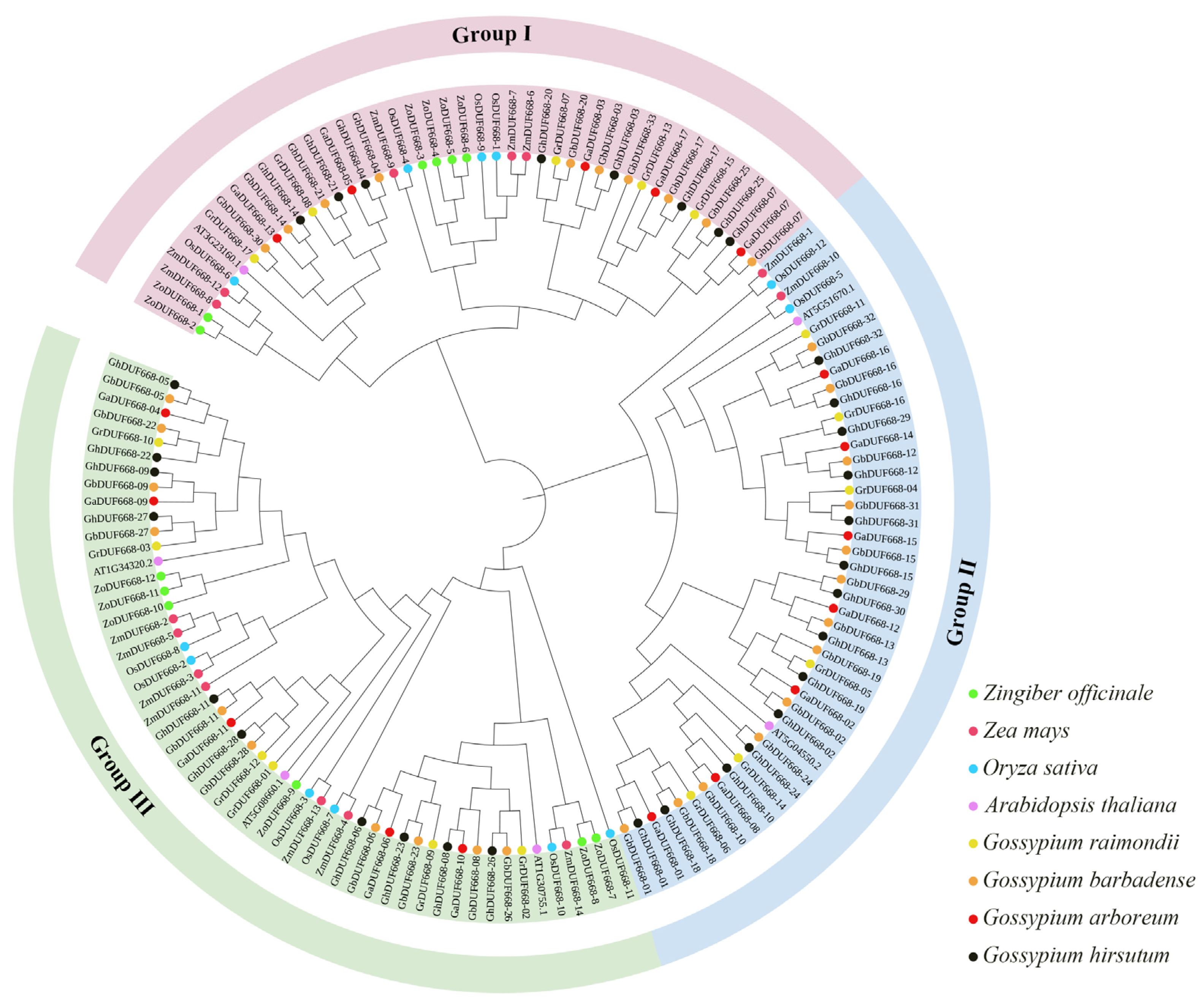
2.1. Identification of ZoDUF668s
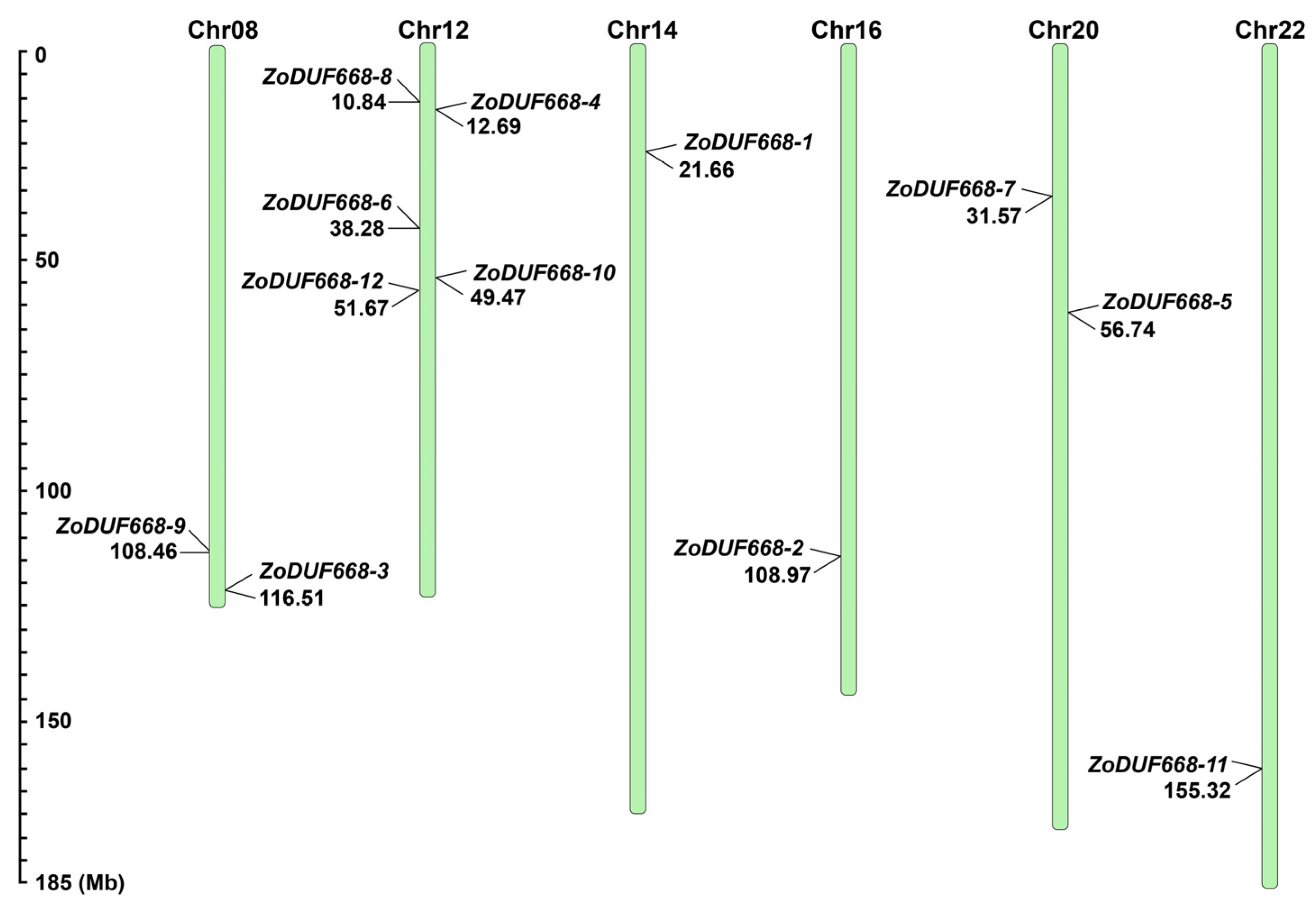
2.2. Chromosomal Distribution of ZoDUF668s
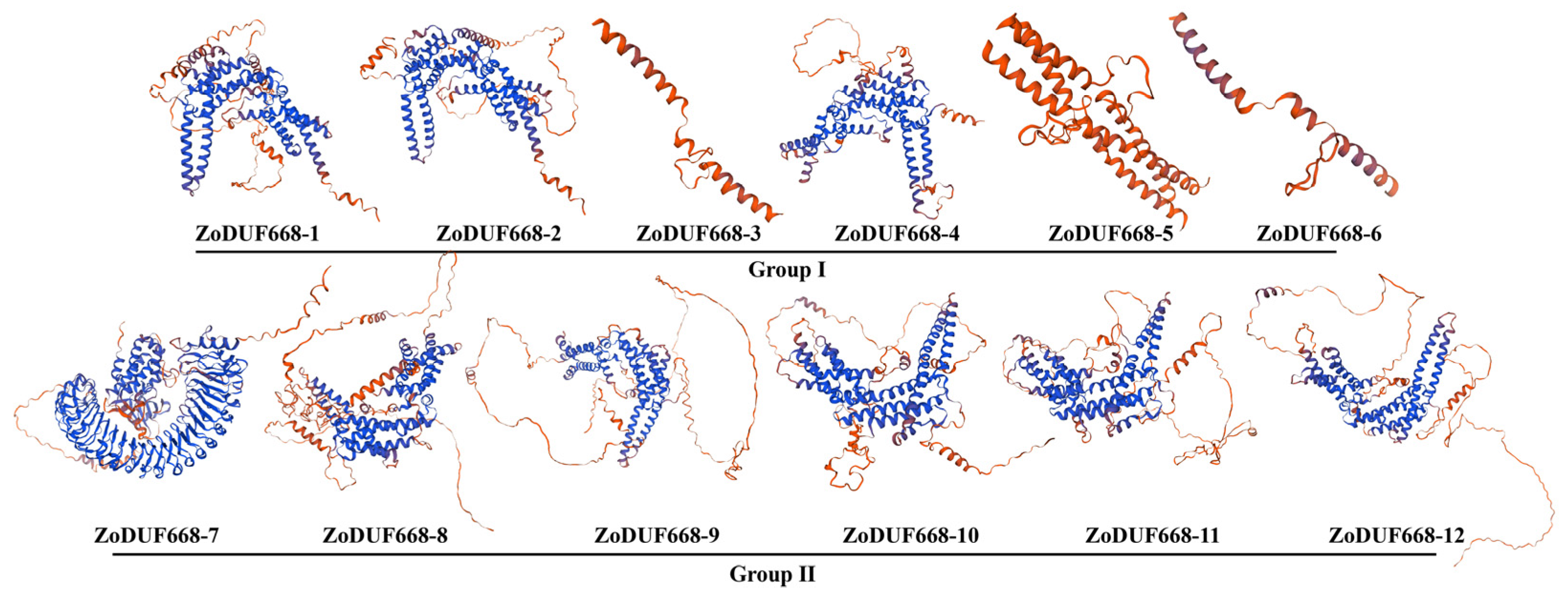
2.3. Characterization Analysis of ZoDUF668 Proteins Sequence
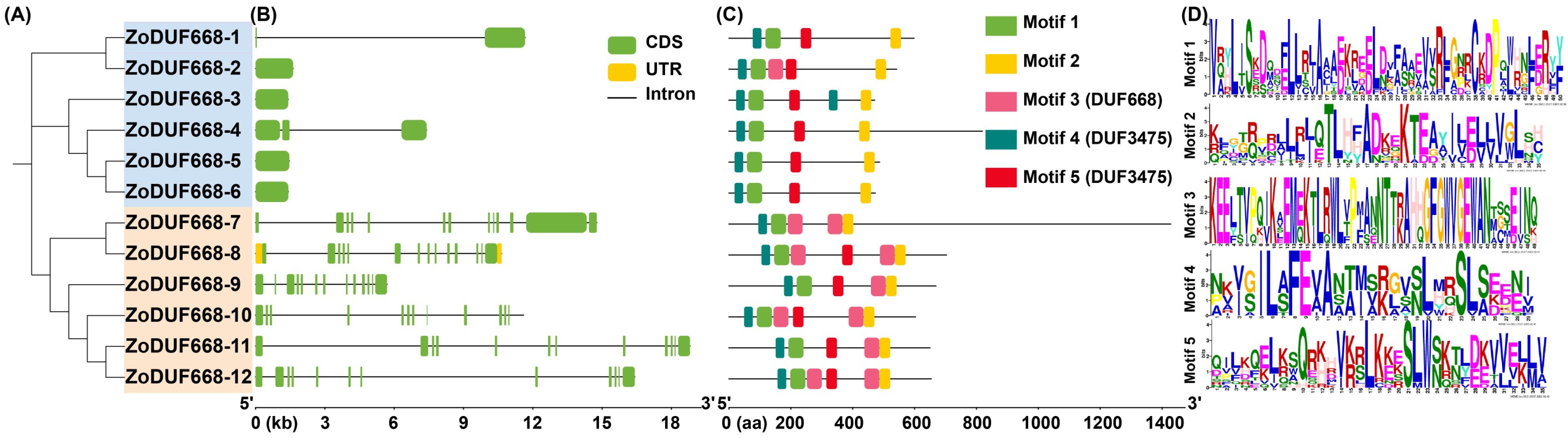
2.4. Gene Structure and Conservative Motif Analysis
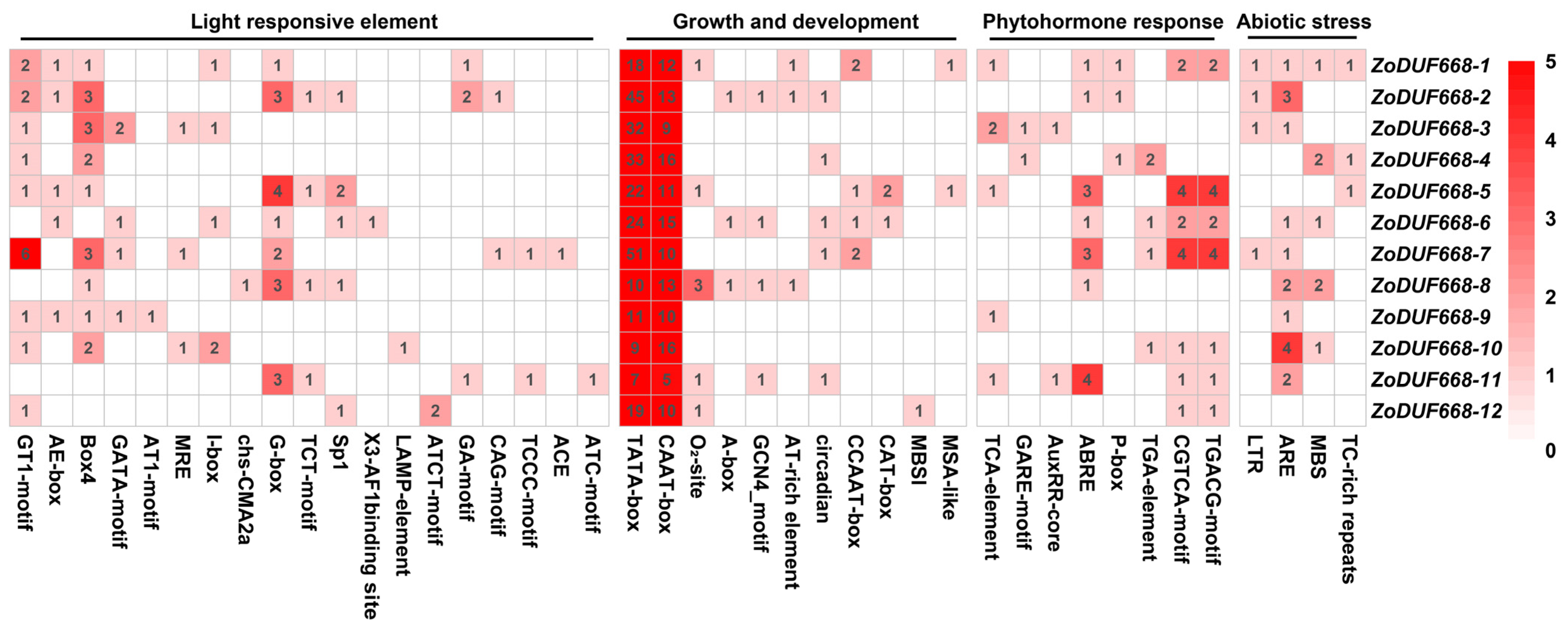
2.5. Promoter Analysis of ZoDUF668s
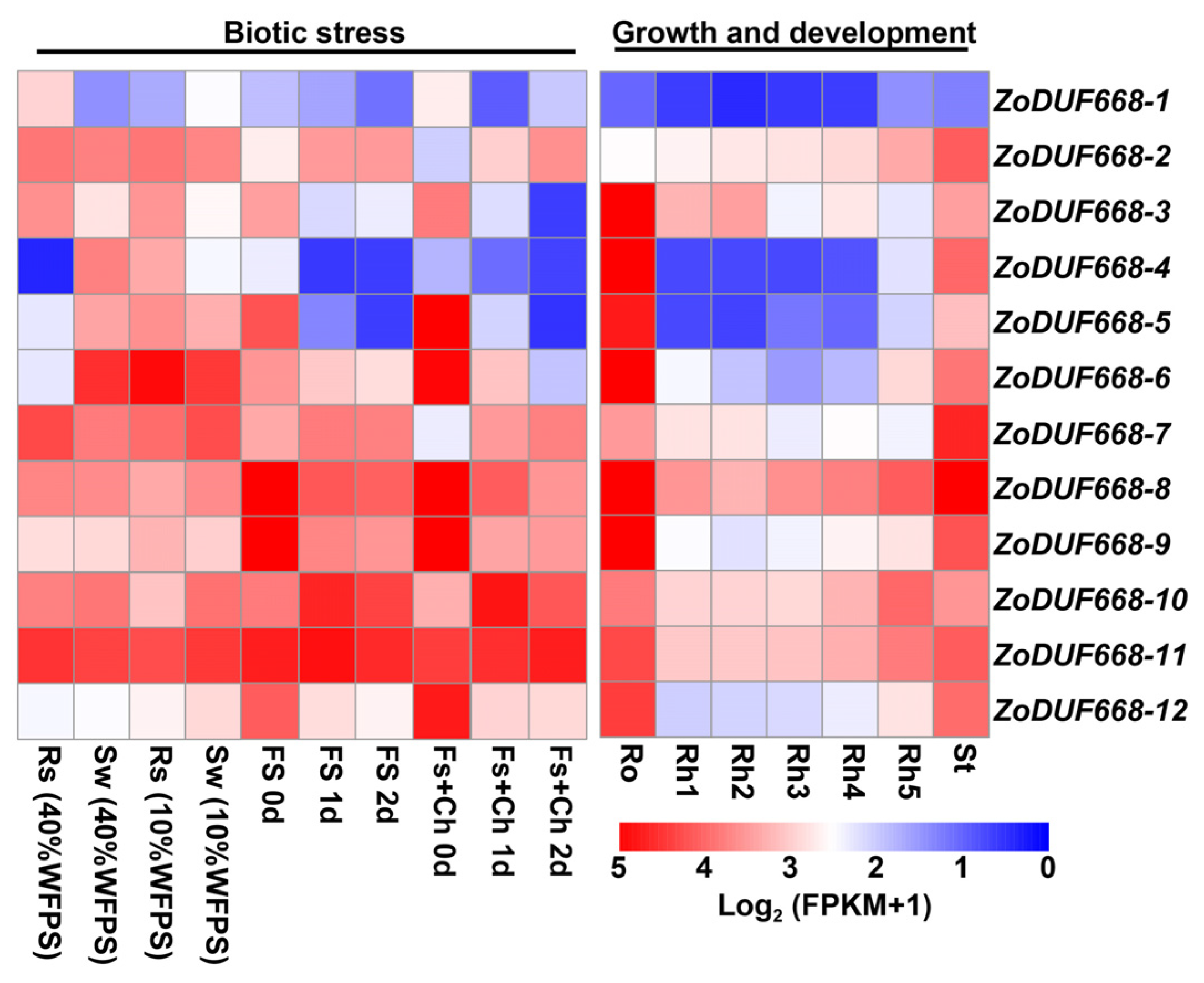
2.6. Expression Profile of ZoDUF668 Gene under Multiple Growth Stages and Stress Conditions
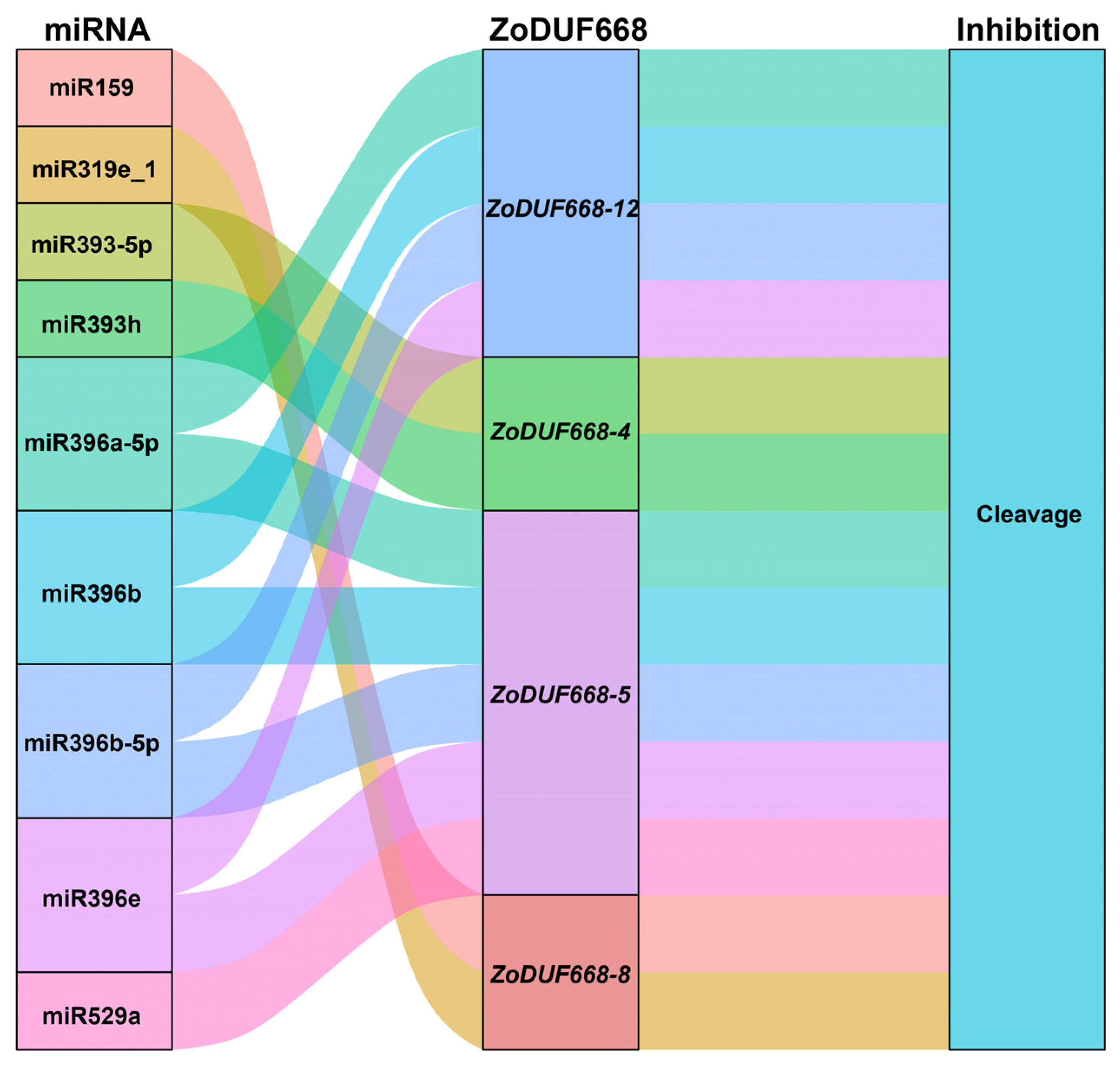
2.7. Post-Transcription Regulation of miRNA to ZoDUF668s
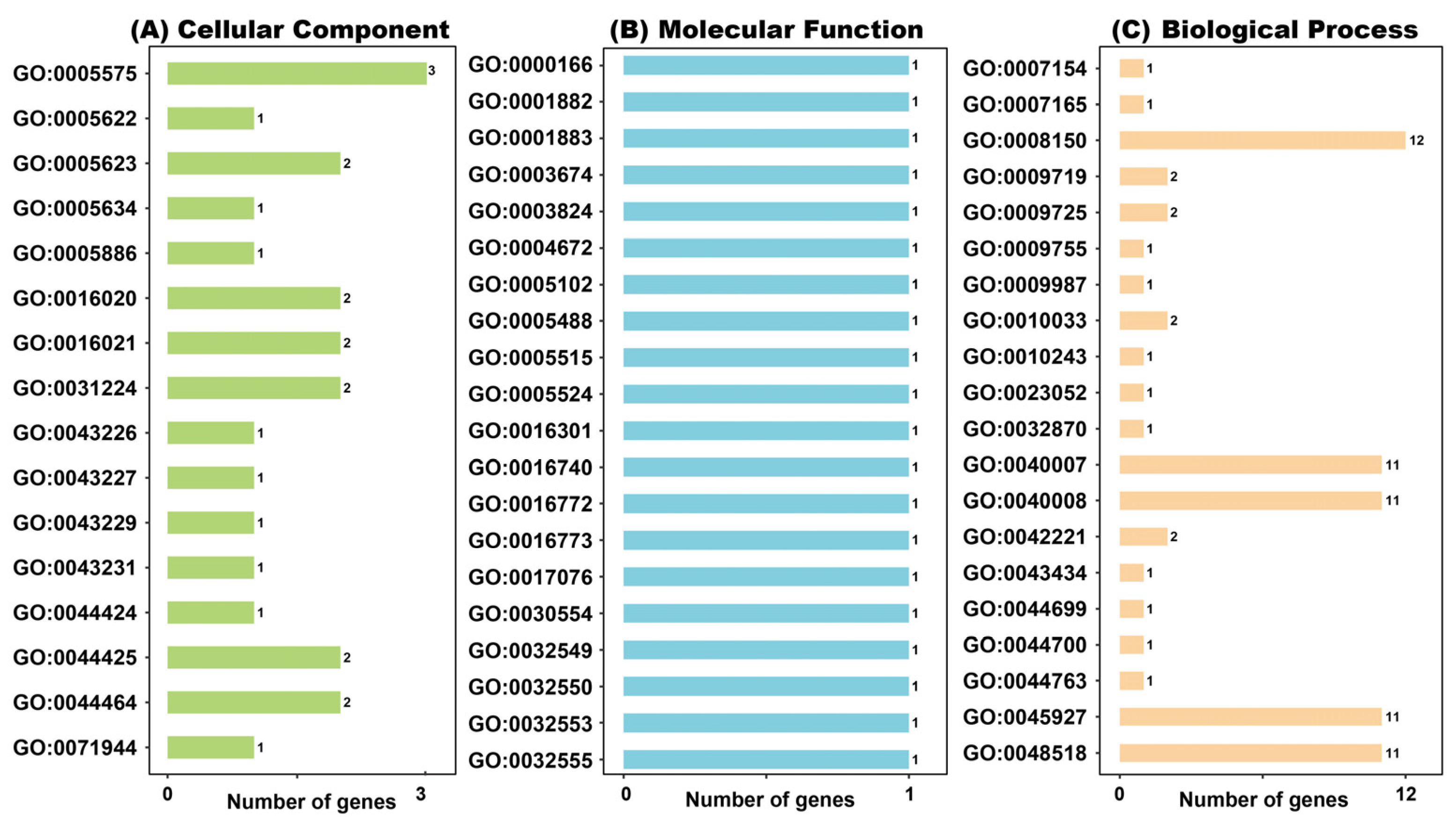
2.8. GO Function Enrichment Analysis
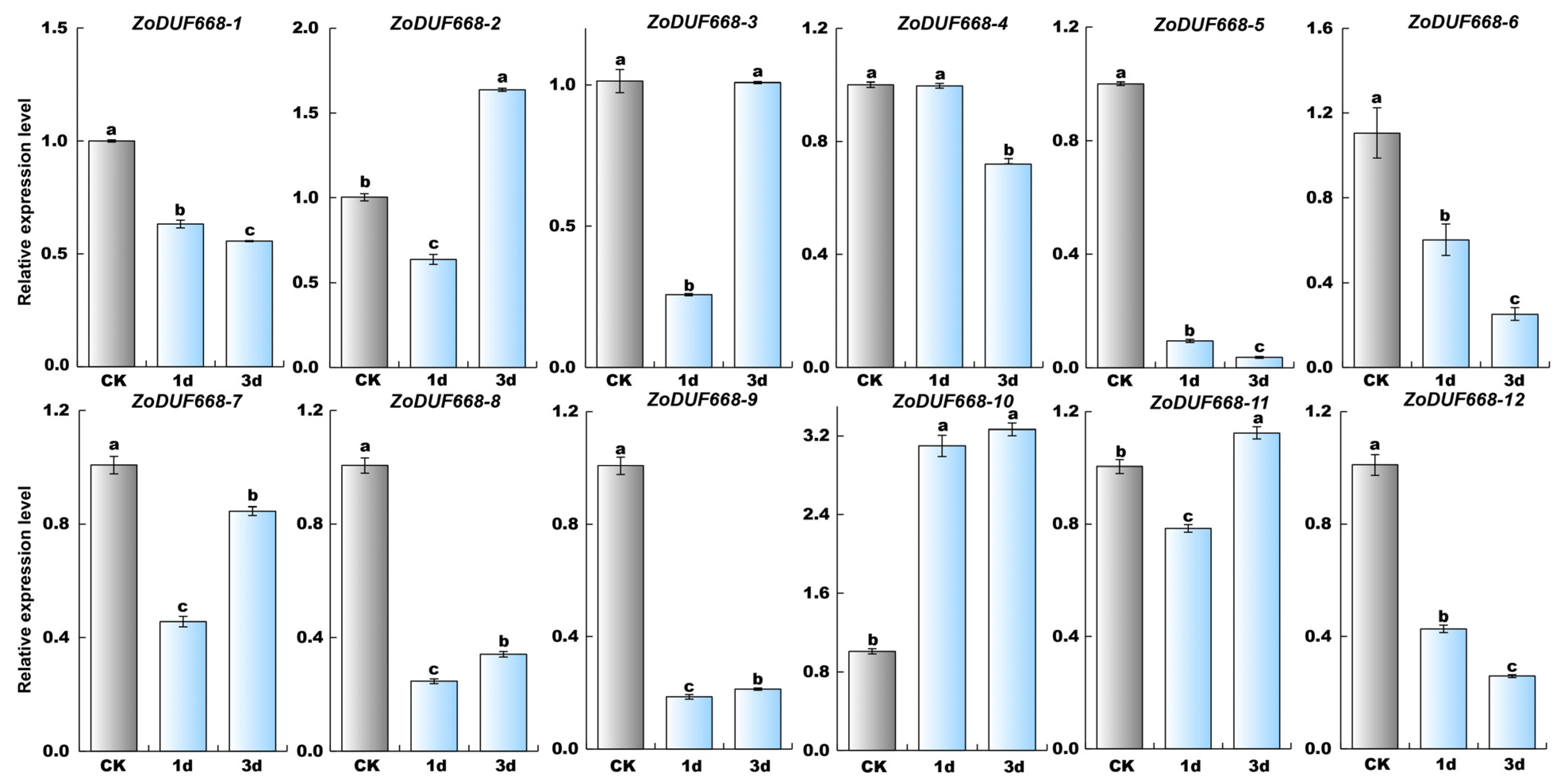
2.9. ZoDUF668s Respond to Biotic Stress
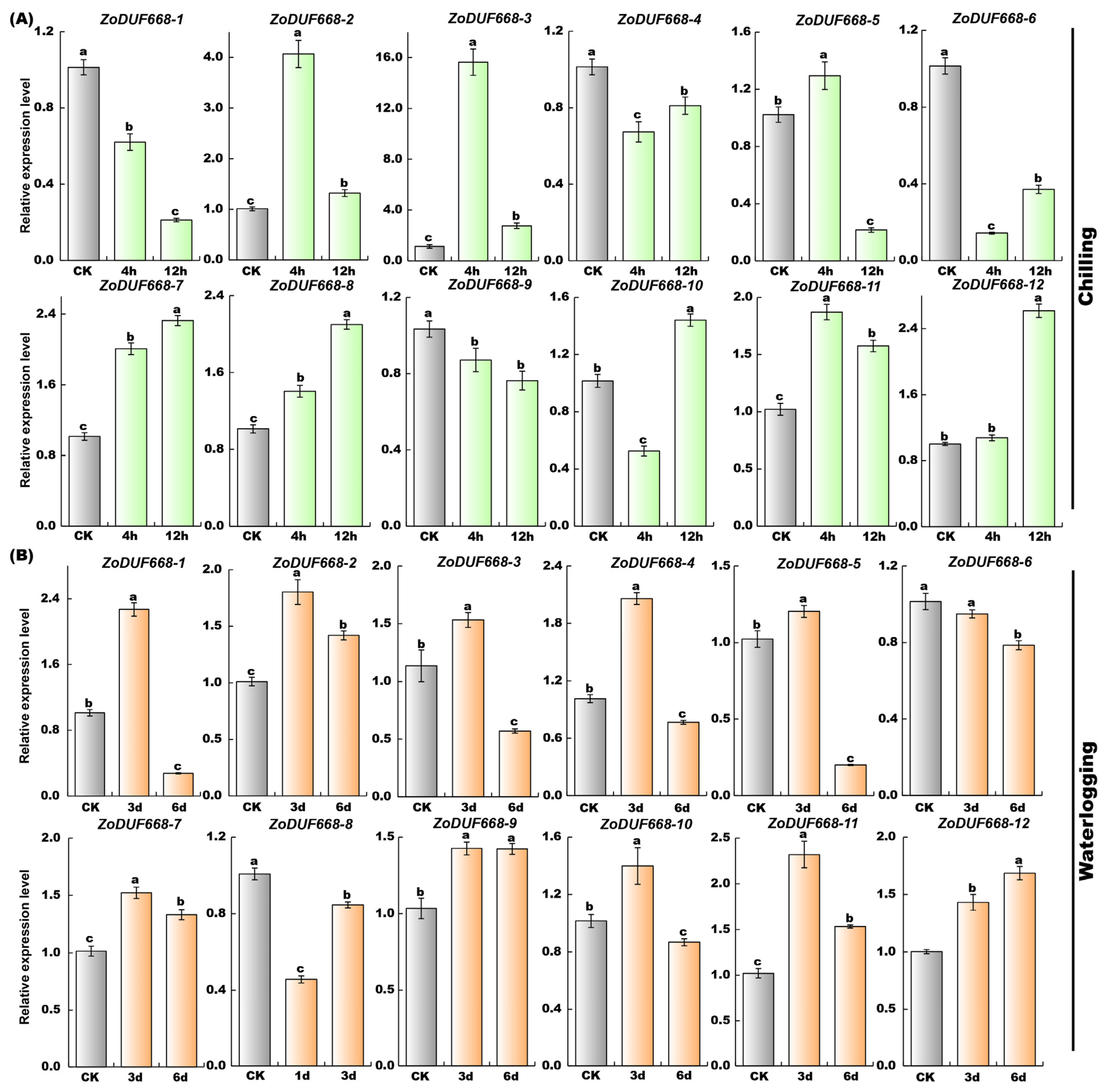
2.10. ZoDUF668s Respond to Abiotic Stress
3. Discussion
4. Materials and Methods
4.1. Identification and Phylogenetic Analysis of ZoDUF668 Gene
4.2. Chromosome Location Analysis
4.3. Gene Structure and Conserved Motifs Analysis of the ZoDUF668 Gene Family
4.4. Prediction of ZoDUF668 Physical and Chemical Properties
4.5. Cis-Element Analysis
4.6. Transcriptomic Analysis of ZoDUF668s
4.7. Prediction of miRNAs Targeting Relationship
4.8. Gene Ontology (GO) Data Analysis and Prediction of ZoDUF668s
4.9. Plant Materials
4.10. RNA Extraction and Quantitative Real-Time PCR Analysis
4.11. Statistical Analysis
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Santosh, D.T.; Sagar, M. Effect of drip irrigation and plastic mulch on yield and quality of ginger (Zingiber officinale). Res. Crops 2022, 23, 211–219. [Google Scholar] [CrossRef]
- Sharma, V.; Thakur, M.; Tomar, M. In vitro selection of gamma irradiated shoots of ginger (Zingiber officinale Rosc.) against Fusarium oxysporum f. sp. zingiberi and molecular analysis of the resistant plants. Plant Cell Tissue Organ Cult. 2020, 143, 319–330. [Google Scholar] [CrossRef]
- Zhong, H.; Zhang, H.Y.; Guo, R.; Wang, Q.; Huang, X.P.; Jianglin, L.; Li, Y.S.; Huang, Y.J.; Wang, Z.H. Characterization and functional divergence of a novel DUF668 gene family in rice based on comprehensive expression patterns. Genes 2019, 10, 980. [Google Scholar] [CrossRef] [PubMed]
- Bateman, A.; Coggill, P.; Finn, R. DUFs: Families in search of function. Acta Crystallogr. Sect. F 2010, 66, 1148–1152. [Google Scholar] [CrossRef] [PubMed]
- Mistry, J.; Chuguransky, S.; Williams, L.; Qureshi, M.; Salazar Gustavo, A.; Sonnhammer, E.L.L.; Tosatto, S.C.E.; Paladin, L.; Raj, S.; Richardson, L.J.; et al. Pfam: The protein families database in 2021. Nucleic Acids Res. 2021, 49, D412–D419. [Google Scholar] [CrossRef] [PubMed]
- Zhao, J.Y.; Wang, P.; Gao, W.J.; Long, Y.L.; Wang, Y.X.; Geng, S.W.; Su, X.N.; Jiao, Y.; Chen, Q.J.; Qu, Y.Y. Genome-wide identification of the DUF668 gene family in cotton and expression profiling analysis of GhDUF668 in Gossypium hirsutum under adverse stress. BMC Genom. 2021, 22, 395. [Google Scholar] [CrossRef] [PubMed]
- Yuan, G.Q.; Zou, T.; He, Z.Y.; Xiao, Q.; Li, G.W.; Liu, S.J.; Xiong, P.P.; Chen, H.; Peng, K.; Zhang, X.; et al. SWOLLEN TAPETUM AND STERILITY 1 is required for tapetum degeneration and pollen wall formation in rice. Plant Physiol. 2022, 190, 352–370. [Google Scholar] [CrossRef] [PubMed]
- Nabi, R.B.S.; Tayade, R.; Imran, Q.M.; Hussain, A.; Shahid, M.; Yun, B.-W. Functional insight of nitric-oxide induced DUF genes in Arabidopsis thaliana. Front. Plant Sci. 2020, 11, 1041. [Google Scholar] [CrossRef] [PubMed]
- Zhou, X.Y.; Zhu, X.G.; Shao, W.N.; Song, J.H.; Jiang, W.Q.; He, Y.Q.; Yin, J.L.; Ma, D.F.; Qiao, Y.L. Genome-wide mining of wheat DUF966 gene family provides new insights into salt stress responses. Front. Plant Sci. 2020, 11, 569838. [Google Scholar] [CrossRef]
- Yin, X.H.; Yuan, Y.; Han, X.W.; Han, S.; Li, Y.T.; Ma, D.F.; Fang, Z.W.; Gong, S.J.; Yin, J.L. Genome-wide identification, characterization, and expression profiling of TaDUF668 gene family in Triticum aestivum. Agronomy 2023, 13, 2178. [Google Scholar] [CrossRef]
- Stührwohldt, N.; Hartmann, J.; Dahlke, R.I.; Oecking, C.; Sauter, M. The PSI family of nuclear proteins is required for growth in arabidopsis. Plant Mol. Biol. 2014, 86, 289–302. [Google Scholar] [CrossRef]
- Zaynab, M.; Sharif, Y.; Xu, Z.S.; Fiaz, S.; Al-Yahyai, R.; Yadikar, H.A.; Al Kashgry, N.A.T.; Qari, S.H.; Sadder, M.; Li, S.F. Genome-wide analysis and expression profiling of DUF668 genes in Glycine max under salt stress. Plants 2023, 12, 2923. [Google Scholar] [CrossRef] [PubMed]
- Liu, E.L.; Li, Z.Q.; Luo, Z.Q.; Xu, L.L.; Jin, P.; Ji, S.; Zhou, G.H.; Wang, Z.Y.; Zhou, Z.L.; Zhang, H. Genome-wide identification of DUF668 gene family and expression analysis under drought and salt stresses in sweet potato [Ipomoea batatas (L.) Lam]. Genes 2023, 14, 217. [Google Scholar] [CrossRef]
- Biju, C.N.; Krishnamurthy, K.S.; Bhat, A.I. Physiological and biochemical response of ginger varieties to virus infection. Plant Physiol. Rep. 2022, 27, 171–179. [Google Scholar] [CrossRef]
- Shiu, S.H.; Bleecker, A.B. Expansion of the receptor-like kinase/pelle gene family and receptor-like proteins in Arabidopsis. Plant Physiol. 2003, 132, 530–543. [Google Scholar] [CrossRef] [PubMed]
- Xu, G.X.; Guo, C.C.; Shan, H.Y.; Kong, H.Z. Divergence of duplicate genes in exon–intron structure. Proc. Natl. Acad. Sci. USA 2012, 109, 1187–1192. [Google Scholar] [CrossRef] [PubMed]
- Yanagisawa, S. Dof domain proteins: Plant-specific transcription factors associated with diverse phenomena unique to plants. Plant Cell Physiol. 2004, 45, 386–391. [Google Scholar] [CrossRef]
- Walther, D.; Brunnemann, R.; Selbig, J. The regulatory code for transcriptional response diversity and its relation to genome structural properties in A. thaliana. PLOS Genet. 2007, 3, e11. [Google Scholar] [CrossRef]
- Han, X.W.; Han, S.; Li, Y.T.; Li, K.K.; Yang, L.J.; Ma, D.F.; Fang, Z.W.; Yin, J.L.; Zhu, Y.X.; Gong, S.J. Double roles of light-harvesting chlorophyll a/b binding protein TaLhc2 in wheat stress tolerance and photosynthesis. Int. J. Biol. Macromol. 2023, 253, 127215. [Google Scholar] [CrossRef]
- Han, R.; Jian, C.; Lv, J.Y.; Yan, Y.; Chi, Q.; Li, Z.J.; Wang, Q.; Zhang, J.; Liu, X.L.; Zhao, H.X. Identification and characterization of microRNAs in the flag leaf and developing seed of wheat (Triticum aestivum L.). BMC Genom. 2014, 15, 289. [Google Scholar] [CrossRef]
- Asefpour Vakilian, K. Machine learning improves our knowledge about miRNA functions towards plant abiotic stresses. Sci. Rep. 2020, 10, 3041. [Google Scholar] [CrossRef]
- Kuczyński, J.; Gracz, J.; Twardowski, T.; Karlowski, W.; Tyczewska, A. Cold stress-induced miRNA and degradome changes in four soybean varieties differing in chilling resistance. J. Agron. Crop Sci. 2021, 208, 777–794. [Google Scholar] [CrossRef]
- Xu, X.W.; Wang, K.X.; Pan, J.W.; Chen, X.H. Small RNA sequencing identifies cucumber miRNA roles in waterlogging-triggered adventitious root primordia formation. Mol. Biol. Rep. 2019, 46, 6381–6389. [Google Scholar] [CrossRef] [PubMed]
- Finn, R.D.; Bateman, A.; Clements, J.; Coggill, P.; Eberhardt, R.Y.; Eddy, S.R.; Heger, A.; Hetherington, K.; Holm, L.; Mistry, J.; et al. Pfam: The protein families database. Nucleic Acids Res. 2014, 42, D222–D230. [Google Scholar] [CrossRef] [PubMed]
- Zou, Z.; Huang, Q.X.; An, F. Genome-wide identification, classification and expression analysis of Lhc supergene family in castor bean (Ricinus communis L.). Agric. Biotechnol. 2013, 2, 44. [Google Scholar] [CrossRef]
- Thompson, J.D.; Higgins, D.G.; Gibson, T.J. CLUSTAL W: Improving the sensitivity of progressive multiple sequence alignment through sequence weighting, position-specific gap penalties and weight matrix choice. Nucleic Acids Res. 1994, 22, 4673–4680. [Google Scholar] [CrossRef]
- Yuan, Y.; Yin, X.H.; Han, X.W.; Han, S.; Li, Y.T.; Ma, D.F.; Fang, Z.W.; Yin, J.L.; Gong, S.J. Genome-wide identification, characterization and expression analysis of the TaDUF724 gene family in wheat (Triticum aestivum). Int. J. Mol. Sci. 2023, 24, 14248. [Google Scholar] [CrossRef]
- Duvaud, S.; Gabella, C.; Lisacek, F.; Stockinger, H.; Ioannidis, V.; Durinx, C. Expasy, the swiss bioinformatics resource portal, as designed by its users. Nucleic Acids Res. 2021, 49, W216–W227. [Google Scholar] [CrossRef]
- Schwede, T.; Kopp, J.r.; Guex, N.; Peitsch, M.C. SWISS-MODEL: An automated protein homology-modeling server. Nucleic Acids Res. 2003, 31, 3381–3385. [Google Scholar] [CrossRef]
- Chou, K.C.; Shen, H.B. Plant-mPLoc: A top-down strategy to augment the power for predicting plant protein subcellular localization. PLoS ONE 2010, 5, e11335. [Google Scholar] [CrossRef]
- Nguyen, H.C.; Lin, K.H.; Ho, S.L.; Chiang, C.M.; Yang, C.M. Enhancing the abiotic stress tolerance of plants: From chemical treatment to biotechnological approaches. Physiol. Plant. 2018, 164, 452–466. [Google Scholar] [CrossRef]
- Li, H.L.; Wu, L.; Dong, Z.; Jiang, Y.; Jiang, S.; Xing, H.; Li, Q.; Liu, G.; Tian, S.; Wu, Z.; et al. Haplotype-resolved genome of diploid ginger (Zingiber officinale) and its unique gingerol biosynthetic pathway. Hortic. Res. 2021, 8, 189. [Google Scholar] [CrossRef]
- Xing, H.T.; Jiang, Y.S.; Zou, Y.; Long, X.L.; Wu, X.L.; Ren, Y.; Li, Y.; Li, H.L. Genome-wide investigation of the AP2/ERF gene family in ginger: Evolution and expression profiling during development and abiotic stresses. BMC Plant Biol. 2021, 21, 561. [Google Scholar] [CrossRef]
- Xing, H.T.; Li, Y.; Ren, Y.; Zhao, Y.; Wu, X.L.; Li, H.L. Genome-wide investigation of microRNAs and expression profiles during rhizome development in ginger (Zingiber officinale Roscoe). BMC Genom. 2022, 23, 1–13. [Google Scholar] [CrossRef]
- Jiang, Y.S.; Huang, M.J.; Wisniewski, M.; Li, H.L.; Zhang, M.X.; Tao, X.; Liu, Y.Q.; Zou, Y. Transcriptome analysis provides insights into gingerol biosynthesis in ginger (Zingiber officinale). Plant Genome 2018, 11, 180034. [Google Scholar] [CrossRef]
- Li, G.; Ma, J.W.; Yin, J.L.; Guo, F.L.; Xi, K.Y.; Yang, P.H.; Cai, X.D.; Jia, Q.; Li, L.; Liu, Y.Q.; et al. Identification of reference genes for reverse transcription-quantitative PCR analysis of ginger under abiotic stress and for postharvest biology studies. Front. Plant Sci. 2022, 13, 1587. [Google Scholar] [CrossRef]
- Livak, K.J.; Schmittgen, T.D. Analysis of relative gene expression data using real-time quantitative PCR and the 2−ΔΔCT method. Methods 2001, 25, 402–408. [Google Scholar] [CrossRef]











{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Name | Gene ID | Len (aa) | MW (kDa) | pI | Atoms | Ins | Sta. | GRAVY | Sub. |
|---|---|---|---|---|---|---|---|---|---|
| ZoDUF668-1 | Maker00038732 | 598 | 67.18 | 8.19 | 9461 | 53.19 | unstable | −0.153 | Nucleus |
| ZoDUF668-2 | Maker00008971 | 542 | 60.70 | 7.55 | 8536 | 56.39 | unstable | −0.151 | Nucleus |
| ZoDUF668-3 | Maker00015846 | 471 | 52.91 | 8.99 | 7477 | 61.19 | unstable | −0.325 | Chloroplast |
| ZoDUF668-4 | Maker00022604 | 819 | 91.12 | 9.15 | 12,756 | 65.43 | unstable | −0.351 | Nucleus |
| ZoDUF668-5 | Maker00068009 | 487 | 54.85 | 9.59 | 7777 | 61.04 | unstable | −0.29 | Chloroplast |
| ZoDUF668-6 | Maker00077672 | 473 | 53.11 | 9.68 | 7508 | 63.83 | unstable | −0.269 | Chloroplast |
| ZoDUF668-7 | Maker00061506 | 1428 | 156.83 | 6.64 | 22,128 | 36.44 | stable | −0.12 | Cell membrane |
| ZoDUF668-8 | Maker00022935 | 703 | 79.57 | 8.57 | 11,217 | 50.84 | unstable | −0.464 | Chloroplast |
| ZoDUF668-9 | Maker00055763 | 669 | 74.67 | 9.32 | 10,544 | 48.28 | unstable | −0.456 | Chloroplast |
| ZoDUF668-10 | Maker00078796 | 603 | 67.72 | 9.13 | 9586 | 44.7 | unstable | −0.431 | Chloroplast |
| ZoDUF668-11 | Maker00034739 | 650 | 72.14 | 9.07 | 10,148 | 55.51 | unstable | −0.59 | Nucleus |
| ZoDUF668-12 | Maker00078495 | 654 | 72.53 | 9.37 | 10,205 | 46.08 | unstable | −0.598 | Nucleus |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Han, S.; Han, X.; Qi, C.; Guo, F.; Yin, J.; Liu, Y.; Zhu, Y. Genome-Wide Identification of DUF668 Gene Family and Expression Analysis under F. solani, Chilling, and Waterlogging Stresses in Zingiber officinale. Int. J. Mol. Sci. 2024, 25, 929. https://doi.org/10.3390/ijms25020929
Han S, Han X, Qi C, Guo F, Yin J, Liu Y, Zhu Y. Genome-Wide Identification of DUF668 Gene Family and Expression Analysis under F. solani, Chilling, and Waterlogging Stresses in Zingiber officinale. International Journal of Molecular Sciences. 2024; 25(2):929. https://doi.org/10.3390/ijms25020929
Chicago/Turabian StyleHan, Shuo, Xiaowen Han, Chuandong Qi, Fengling Guo, Junliang Yin, Yiqing Liu, and Yongxing Zhu. 2024. "Genome-Wide Identification of DUF668 Gene Family and Expression Analysis under F. solani, Chilling, and Waterlogging Stresses in Zingiber officinale" International Journal of Molecular Sciences 25, no. 2: 929. https://doi.org/10.3390/ijms25020929