
Quantitative Analysis of the Human Semen Phosphorometabolome by 31P-NMR
 ,
,  , , , and
, , , and
Abstract
:1. Introduction
2. Results
2.1. Sperm Quality Measures
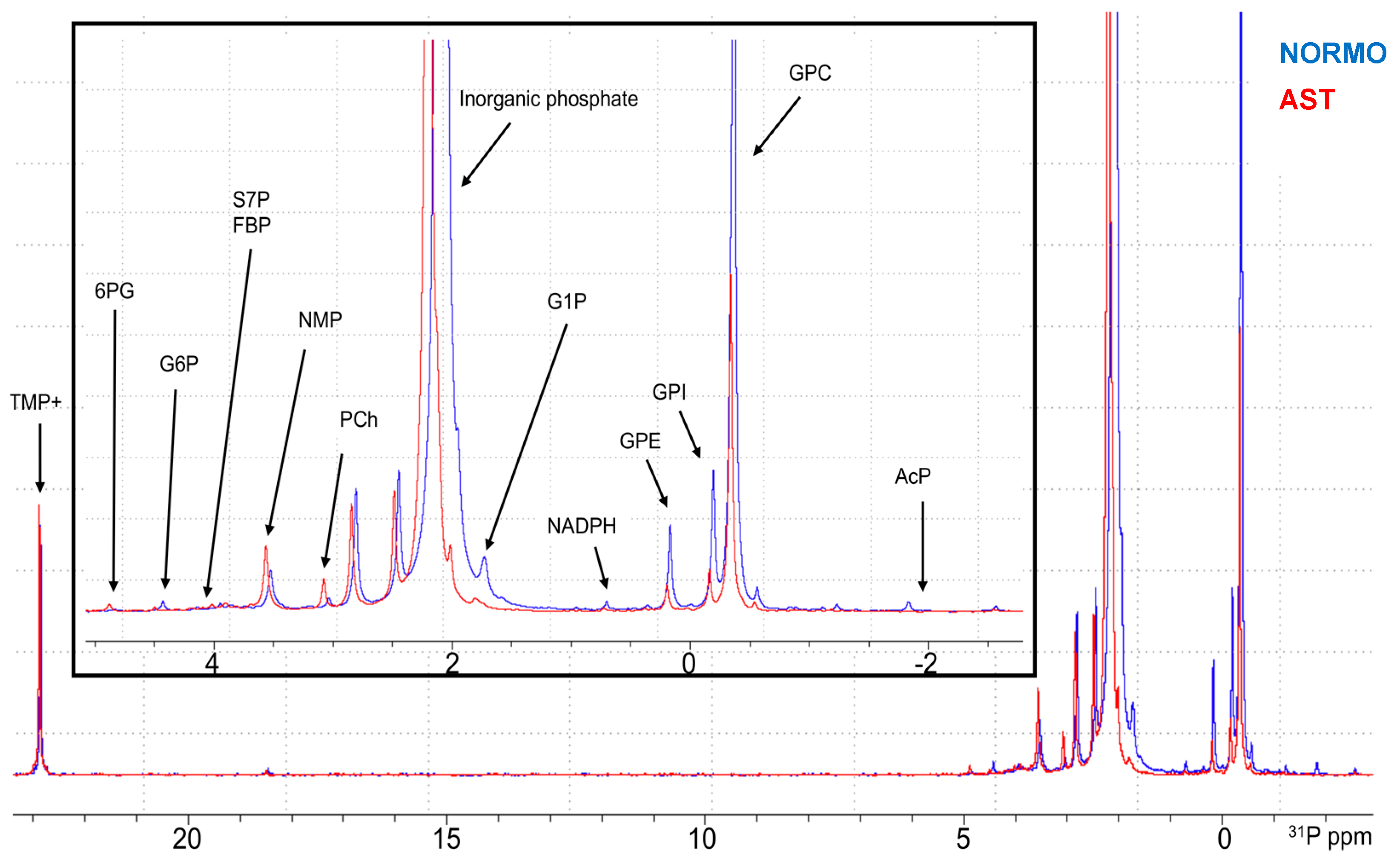
2.2. Phosphoromic Analysis of Semen: General View
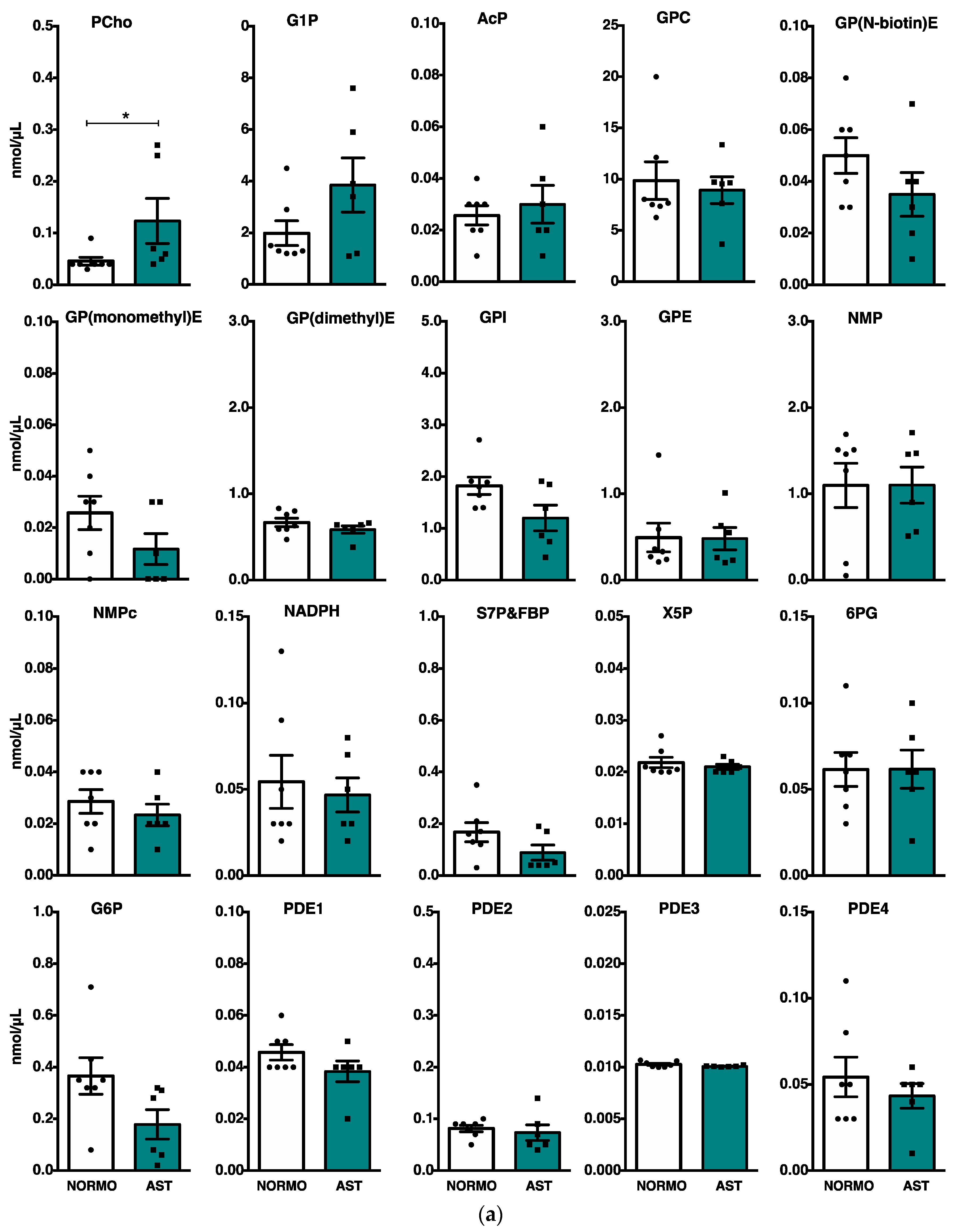
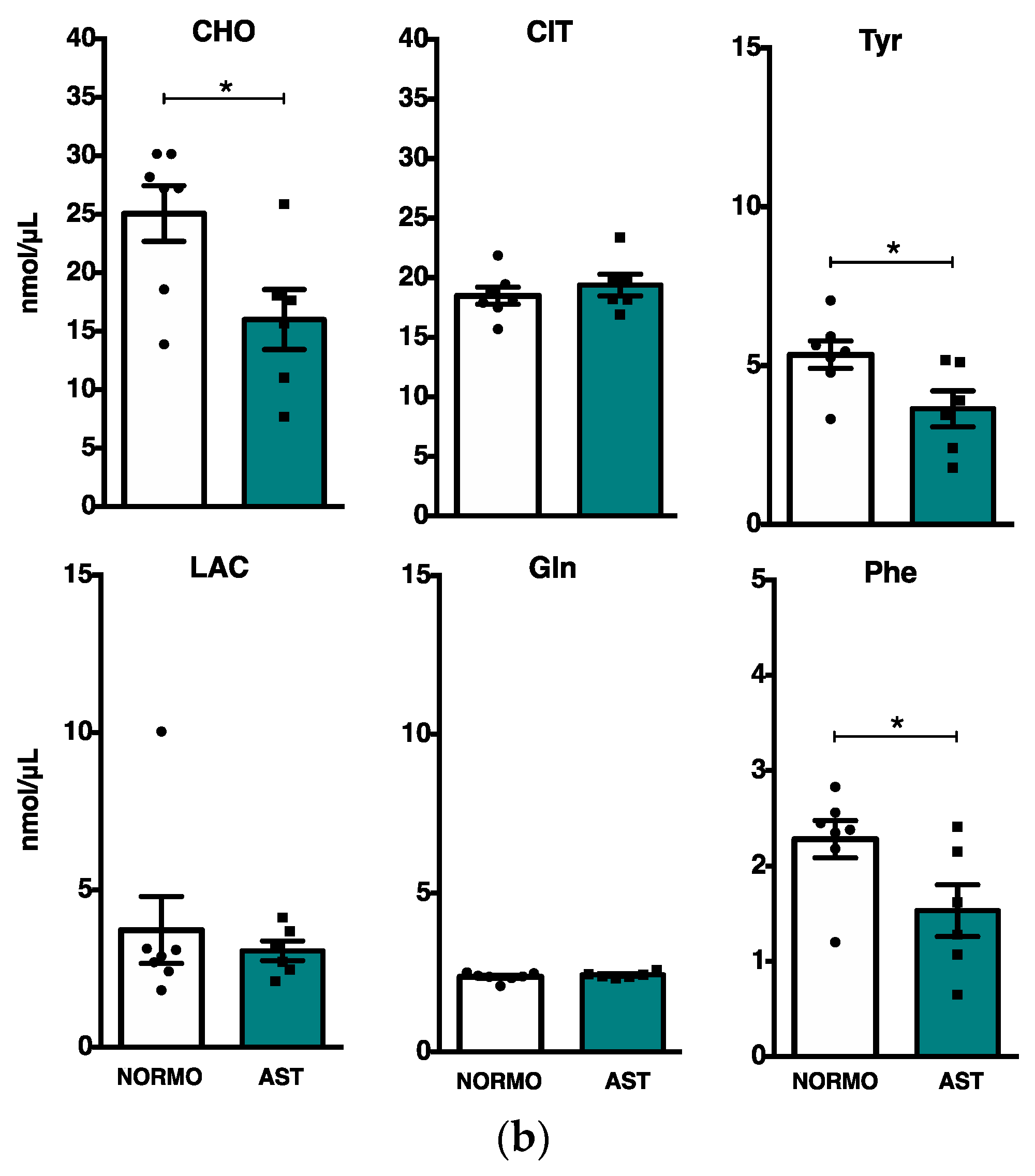
2.3. Phosphoromics Analysis in Seminal Plasma from AST and NORMO Groups
2.4. Metabolic Phosphorome Differences between Spermatozoa and Seminal Plasma from NORMO Group
3. Discussion
4. Materials and Methods
4.1. Human Semen Samples
4.2. Human Spermatozoa Motility Evaluation
4.3. Analysis of Human Sperm Functional Parameters by Flow Cytometry
4.4. Human Sperm Samples Preparation
4.5. NMR Sample Preparation
4.6. NMR Measurements
4.7. Identification and Quantification of Metabolites
4.8. Statistical Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Marshall, D.D.; Powers, R. Beyond the Paradigm: Combining Mass Spectrometry and Nuclear Magnetic Resonance for Metabolomics. Prog. Nucl. Magn. Reson. Spectrosc. 2017, 100, 1–16. [Google Scholar] [CrossRef]
- Cox, N.; Millard, P.; Charlier, C.; Lippens, G. Improved NMR Detection of Phospho-Metabolites in a Complex Mixture. Anal. Chem. 2021, 93, 4818–4824. [Google Scholar] [CrossRef]
- Blaurock, J.; Baumann, S.; Grunewald, S.; Schiller, J.; Engel, K.M. Metabolomics of Human Semen: A Review of Different Analytical Methods to Unravel Biomarkers for Male Fertility Disorders. Int. J. Mol. Sci. 2022, 23, 9031. [Google Scholar] [CrossRef]
- Qiao, S.; Wu, W.; Chen, M.; Tang, Q.; Xia, Y.; Jia, W.; Wang, X. Seminal Plasma Metabolomics Approach for the Diagnosis of Unexplained Male Infertility. PLoS ONE 2017, 12, e0181115. [Google Scholar] [CrossRef]
- Tang, B.; Shang, X.; Qi, H.; Li, J.; Ma, B.; An, G.; Zhang, Q. Metabonomic Analysis of Fatty Acids in Seminal Plasma between Healthy and Asthenozoospermic Men Based on Gas Chromatography Mass Spectrometry. Andrologia 2017, 49, e12744. [Google Scholar] [CrossRef]
- Gilany, K.; Jafarzadeh, N.; Mani-Varnosfaderani, A.; Minai-Tehrani, A.; Sadeghi, M.R.; Darbandi, M.; Darbandi, S.; Amini, M.; Arjmand, B.; Pahlevanzadeh, Z. Metabolic Fingerprinting of Seminal Plasma from Non-Obstructive Azoospermia Patients: Positive versus Negative Sperm Retrieval. J. Reprod. Infertil. 2018, 19, 109–114. [Google Scholar]
- Boguenet, M.; Bocca, C.; Bouet, P.E.; Serri, O.; Chupin, S.; Tessier, L.; Blanchet, O.; El Hachem, H.; Chao de la Barca, J.M.; Reynier, P.; et al. Metabolomic Signature of the Seminal Plasma in Men with Severe Oligoasthenospermia. Andrology 2020, 8, 1859–1866. [Google Scholar] [CrossRef]
- Chen, L.; Wen, C.W.; Deng, M.J.; Zhang, Z.D.; Zhou, Z.H.; Wang, X. Metabolic and Transcriptional Changes in Seminal Plasma of Asthenozoospermia Patients. Biomed. Chromatogr. 2020, 34, e4769. [Google Scholar] [CrossRef] [PubMed]
- Mehrparvar, B.; Chashmniam, S.; Nobakht, F.; Amini, M.; Javidi, A.; Minai-Tehrani, A.; Arjmand, B.; Gilany, K. Metabolic Profiling of Seminal Plasma from Teratozoospermia Patients. J. Pharm. Biomed. Anal. 2020, 178, 112903. [Google Scholar] [CrossRef] [PubMed]
- Mumcu, A.; Karaer, A.; Dogan, B.; Tuncay, G. Metabolomics Analysis of Seminal Plasma in Patients with Idiopathic Oligoasthenoteratozoospermia Using High-Resolution NMR Spectroscopy. Andrology 2020, 8, 450–456. [Google Scholar] [CrossRef] [PubMed]
- Murgia, F.; Corda, V.; Serrenti, M.; Usai, V.; Santoru, M.L.; Hurt, K.J.; Passaretti, M.; Monni, M.C.; Atzori, L.; Monni, G. Seminal Fluid Metabolomic Markers of Oligozoospermic Infertility in Humans. Metabolites 2020, 10, 64. [Google Scholar] [CrossRef]
- Xu, Y.; Lu, H.; Wang, Y.; Zhang, Z.; Wu, Q. Comprehensive Metabolic Profiles of Seminal Plasma with Different Forms of Male Infertility and Their Correlation with Sperm Parameters. J. Pharm. Biomed. Anal. 2020, 177, 112888. [Google Scholar] [CrossRef]
- Serri, O.; Boguenet, M.; Chao de la Barca, J.M.; Bouet, P.-E.; El Hachem, H.; Blanchet, O.; Reynier, P.; May-Panloup, P. A Metabolomic Profile of Seminal Fluid in Extremely Severe Oligozoopermia Suggesting an Epididymal Involvement. Metabolites 2022, 12, 1266. [Google Scholar] [CrossRef]
- Zhao, K.; Zhang, J.; Xu, Z.; Xu, Y.; Xu, A.; Chen, W.; Miao, C.; Liu, S.; Wang, Z.; Jia, R. Metabolomic Profiling of Human Spermatozoa in Idiopathic Asthenozoospermia Patients Using Gas Chromatography-Mass Spectrometry. BioMed Res. Int. 2018, 2018, 8327506. [Google Scholar] [CrossRef] [PubMed]
- Engel, K.M.; Baumann, S.; Blaurock, J.; Rolle-Kampczyk, U.; Schiller, J.; Von Bergen, M.; Grunewald, S. Differences in the Sperm Metabolomes of Smoking and Nonsmoking Men. Biol. Reprod. 2021, 105, 1484–1493. [Google Scholar] [CrossRef] [PubMed]
- Reynolds, S.; Calvert, S.J.; Walters, S.J.; Paley, M.N.; Pacey, A.A. NMR Spectroscopy of Live Human Asthenozoospermic and Normozoospermic Sperm Metabolism. Reprod. Fertil. 2022, 3, 77–89. [Google Scholar] [CrossRef] [PubMed]
- Paiva, C.; Amaral, A.; Rodriguez, M.; Canyellas, N.; Correig, X.; Ballescà, J.L.; Ramalho-Santos, J.; Oliva, R. Identification of Endogenous Metabolites in Human Sperm Cells Using Proton Nuclear Magnetic Resonance (1H-NMR) Spectroscopy and Gas Chromatography-Mass Spectrometry (GC-MS). Andrology 2015, 3, 496–505. [Google Scholar] [CrossRef] [PubMed]
- Reynolds, S.; Calvert, S.J.; Paley, M.N.; A Pacey, A. 1H Magnetic Resonance Spectroscopy of Live Human Sperm. Mol. Hum. Reprod. 2017, 23, 441–451. [Google Scholar] [CrossRef] [PubMed]
- Calvert, S.J.; Reynolds, S.; Paley, M.N.; Walters, S.J.; Pacey, A.A. Probing Human Sperm Metabolism Using 13C-Magnetic Resonance Spectroscopy. Mol. Hum. Reprod. 2019, 25, 30–41. [Google Scholar] [CrossRef]
- Engel, K.M.; Baumann, S.; Rolle-Kampczyk, U.; Schiller, J.; von Bergen, M.; Grunewald, S. Metabolomic Profiling Reveals Correlations between Spermiogram Parameters and the Metabolites Present in Human Spermatozoa and Seminal Plasma. PLoS ONE 2019, 14, e0211679. [Google Scholar] [CrossRef]
- Gamidov, S.I.; Shatylko, T.V.; Tambiev, A.K.; Tokareva, A.O.; Chagovets, V.V.; Bitsoev, T.B.; Starodubtseva, N.L.; Popova, A.Y.; Frankevich, V.E. Prognostic Ability of Seminal Plasma Lipidomic Analysis in Predicting the Success of microTESE in Men with Azoospermia. Urologiia 2022, 2022, 32–37. [Google Scholar] [CrossRef]
- Neto, F.T.L.; Marques, R.A.; Cavalcanti Filho, A.d.F.; da Fonte, J.E.F.; Lima, S.V.C.; Silva, R.O. Prediction of Semen Analysis Parameter Improvement after Varicocoelectomy Using 1 H NMR-Based Metabonomics Assays. Andrology 2022, 10, 1581–1592. [Google Scholar] [CrossRef]
- Wishart, D.S.; Guo, A.C.; Oler, E.; Wang, F.; Anjum, A.; Peters, H.; Dizon, R.; Sayeeda, Z.; Tian, S.; Lee, B.L.; et al. HMDB 5.0: The Human Metabolome Database for 2022. Nucleic Acids Res. 2022, 50, D622–D631. [Google Scholar] [CrossRef]
- Bhinderwala, F.; Evans, P.; Jones, K.; Laws, B.R.; Smith, T.G.; Morton, M.; Powers, R. Phosphorus NMR and Its Application to Metabolomics. Anal. Chem. 2020, 92, 9536–9545. [Google Scholar] [CrossRef] [PubMed]
- Bhinderwala, F.; Lonergan, S.; Woods, J.; Zhou, C.; Fey, P.D.; Powers, R. Expanding the Coverage of the Metabolome with Nitrogen-Based NMR. Anal. Chem. 2018, 90, 4521–4528. [Google Scholar] [CrossRef] [PubMed]
- Ragavan, M.; Merritt, M.E. Nuclear Magnetic Resonance Measurement of Metabolic Flux Using 13C and 1H Signals. In Methods in Molecular Biology; Humana Press: Totowa, NJ, USA, 2019; Volume 1996. [Google Scholar]
- Bernardo-Seisdedos, G.; Bilbao, J.; Fernández-Ramos, D.; Lopitz-Otsoa, F.; Gutierrez de Juan, V.; Bizkarguenaga, M.; Mateos, B.; Fondevila, M.F.; Abril-Fornaguera, J.; Diercks, T.; et al. Metabolic Landscape of the Mouse Liver by Quantitative 31P Nuclear Magnetic Resonance Analysis of the Phosphorome. Hepatology 2021, 74, 148–163. [Google Scholar] [CrossRef] [PubMed]
- Robitaille, P.-M.L.; Robitaille, P.A.; Martin, P.A.; Brown, G.G. Phosphorus-31 Nuclear Magnetic Resonance Studies of Spermatozoa from the Boar, Ram, Goat and Bull. Comp. Biochem. Physiol. Part B Comp. Biochem. 1987, 87, 285–296. [Google Scholar] [CrossRef]
- Leßig, J.; Gey, C.; Süß, R.; Schiller, J.; Glander, H.-J.; Arnhold, J. Analysis of the Lipid Composition of Human and Boar Spermatozoa by MALDI-TOF Mass Spectrometry, Thin Layer Chromatography and 31P NMR Spectroscopy. Comp. Biochem. Physiol. Part B Biochem. Mol. Biol. 2004, 137, 265–277. [Google Scholar] [CrossRef]
- Martin-Hidalgo, D.; Serrano, R.; Zaragoza, C.; Garcia-Marin, L.J.; Bragado, M.J. Human Sperm Phosphoproteome Reveals Differential Phosphoprotein Signatures That Regulate Human Sperm Motility. J. Proteom. 2020, 215, 103654. [Google Scholar] [CrossRef]
- World Health Organization. WHO Laboratory Manual for the Examination and Processing of Human Semen, 6th ed.; World Health Organization: Geneva, Switzerland, 2021. [Google Scholar]
- López-Ibáñez, J.; Pazos, F.; Chagoyen, M. MBROLE 2.0-Functional Enrichment of Chemical Compounds. Nucleic Acids Res. 2016, 44, W201–W204. [Google Scholar] [CrossRef]
- Chen, L.; Lu, W.; Wang, L.; Xing, X.; Chen, Z.; Teng, X.; Zeng, X.; Muscarella, A.D.; Shen, Y.; Cowan, A.; et al. Metabolite Discovery through Global Annotation of Untargeted Metabolomics Data. Nat. Methods 2021, 18, 1377–1385. [Google Scholar] [CrossRef]
- Alipour, H.; Duus, R.K.; Wimmer, R.; Dardmeh, F.; Du Plessis, S.S.; Jørgensen, N.; Christiansen, O.B.; Hnida, C.; Nielsen, H.I.; Van Der Horst, G. Seminal Plasma Metabolomics Profiles Following Long (4–7 Days) and Short (2 h) Sexual Abstinence Periods. Eur. J. Obstet. Gynecol. Reprod. Biol. 2021, 264, 178–183. [Google Scholar] [CrossRef] [PubMed]
- Zhang, X.; Diao, R.; Zhu, X.; Li, Z.; Cai, Z. Metabolic Characterization of Asthenozoospermia Using Nontargeted Seminal Plasma Metabolomics. Clin. Chim. Acta 2015, 450, 254–261. [Google Scholar] [CrossRef]
- Arrata, W.S.M.; Burt, T.; Corder, S. The Role of Phosphate Esters in Male Fertility. Fertil. Steril. 1978, 30, 329–333. [Google Scholar] [CrossRef]
- Lambert, I.H.; Kristensen, D.M.; Holm, J.B.; Mortensen, O.H. Physiological Role of Taurine—From Organism to Organelle. Acta Physiol. 2015, 213, 191–212. [Google Scholar] [CrossRef]
- Rand, S.; Winkelbauer, G.; Brinkmann, B. Investigations on PGM Activity and Electrophoretic Patterns from Ejaculates and Seminal Stains. In Proceedings of the 11th Congress of the Society for Forensic Haemogenetics (Gesellschaft für Forensische Blutgruppenkunde e.V.), Copenhagen, Denmark, 6–10 August 1985; Advances in Forensic Haemogenetics; Brinkmann, B., Henningsen, K., Eds.; Springer: Berlin/Heidelberg, Germany, 1986; Volume 1. [Google Scholar] [CrossRef]
- García-Rodríguez, A.; de la Casa, M.; Peinado, H.; Gosálvez, J.; Roy, R. Human Prostasomes from Normozoospermic and Non-Normozoospermic Men Show a Differential Protein Expression Pattern. Andrology 2018, 6, 585–596. [Google Scholar] [CrossRef]
- Banjoko, S.O.; Adeseolu, F.O. Seminal Plasma PH, Inorganic Phosphate, Total and Ionized Calcium Concentrations in the Assessment of Human Spermatozoa Function. J. Clin. Diagn. Res. 2013, 7, 2483–2486. [Google Scholar] [CrossRef]
- Asghari, A.; Marashi, S.-A.; Ansari-Pour, N. A Sperm-Specific Proteome-Scale Metabolic Network Model Identifies Non-Glycolytic Genes for Energy Deficiency in Asthenozoospermia. Syst. Biol. Reprod. Med. 2017, 63, 100–112. [Google Scholar] [CrossRef] [PubMed]
- Lane-Roberts, C.; Sharman, A.; Walker, K.; Barton, M. Sterility and Impaired Fertility: Pathogenesis, Investigation and Treatment; Hamish Hamilton: London, UK, 1948. [Google Scholar]
- Drabovich, A.P.; Saraon, P.; Jarvi, K.; Diamandis, E.P. Seminal Plasma as a Diagnostic Fluid for Male Reproductive System Disorders. Nat. Rev. Urol. 2014, 11, 278–288. [Google Scholar] [CrossRef]
- Lombó, M.; Ruiz-Díaz, S.; Gutiérrez-Adán, A.; Sánchez-Calabuig, M.-J. Sperm Metabolomics through Nuclear Magnetic Resonance Spectroscopy. Animals 2021, 11, 1669. [Google Scholar] [CrossRef]
- Furse, S.; Kusinski, L.C.; Ray, A.; Glenn-Sansum, C.; Williams, H.E.L.; Koulman, A.; Meek, C.L. Relative Abundance of Lipid Metabolites in Spermatozoa across Three Compartments. Int. J. Mol. Sci. 2022, 23, 11655. [Google Scholar] [CrossRef]
- Nevalainen, T.J.; Meri, K.-M.; Niemi, M. Synovial-type (Group II) Phospholipase A2 Human Seminal Plasma. Andrologia 1993, 25, 355–358. [Google Scholar] [CrossRef]
- Kallajoki, M.; Alanen, K.A.; Nevalainen, M.; Nevalainen, T.J. Group II Phospholipase A2 in Human Male Reproductive Organs and Genital Tumors. Prostate 1998, 35, 263–272. [Google Scholar] [CrossRef]
- Sato, H.; Taketomi, Y.; Isogai, Y.; Miki, Y.; Yamamoto, K.; Masuda, S.; Hosono, T.; Arata, S.; Ishikawa, Y.; Ishii, T.; et al. Group III Secreted Phospholipase A2 Regulates Epididymal Sperm Maturation and Fertility in Mice. J. Clin. Investig. 2010, 120, 1400–1414. [Google Scholar] [CrossRef]
- Hamamah, S.; Seguin, F.; Barthelemy, C.; Akoka, S.; Le Pape, A.; Lansac, J.; Royere, D. 1H Nuclear Magnetic Resonance Studies of Seminal Plasma from Fertile and Infertile Men. Reproduction 1993, 97, 51–55. [Google Scholar] [CrossRef]
- Gupta, A.; Mahdi, A.A.; Ahmad, M.K.; Shukla, K.K.; Jaiswer, S.P.; Shankhwar, S.N. 1H NMR Spectroscopic Studies on Human Seminal Plasma: A Probative Discriminant Function Analysis Classification Model. J. Pharm. Biomed. Anal. 2011, 54, 106–113. [Google Scholar] [CrossRef]




{kind=link}
{kind=link}
{kind=link}
| NORMO | AST | |
|---|---|---|
| Semen volume (mL) | 3 ± 1 | 3 ± 0 |
| Sperm concentration (106 per mL) | 167 ± 29 | 47 ± 11 ** |
| Total sperm motility (%) | 71 ± 2 | 27 ± 5 ** |
| Progressive motility (%) | 45 ± 4 | 17 ± 4 ** |
| Rapid progressive (%) | 28 ± 3 | 11 ± 3 ** |
| Sperm viability (%) | 70 ± 3 | 46 ± 2 ** |
| hMMP (%) | 49 ± 4 | 44 ± 9 |
| SOP (%) | 31 ± 7 | 37 ± 2 |
| (a) | (b) | ||||||
|---|---|---|---|---|---|---|---|
| NMR | Metabolites | ↑ CELL | ↑ SP | Metabolic Pathway | p Value | Metabolites | |
| 1H | Citrate | 1.28 ** | CELL | ||||
| CHO | 1.30 ** | Signal transduction | 1.32 × 10−5 | NMP|NMPc | |||
| Tyr | 1.25 | Purine metabolism | 3.34 × 10−4 | NMP|NMPc | |||
| LAC | 1.07 | Ether lipid metabolism | 3.42 × 10−3 | GP(N-biotin)E|GP(monomethyl)E | |||
| Gln | 1.57 ** | Metabolic pathways | 4.39 × 10−3 | NMP|G1P|PCh|AcP|X5P FBP|S7P|GP(monomethyl)E GP(N-biotin)E | |||
| Phe | 1.01 | ||||||
| 31P | GPC | 1.11 | |||||
| NMP | 2.12 * | PPP | 6.57 × 10−3 | X5P|S7P | |||
| S7P and FBP | 4.95 | GPL metabolism | 1.33 × 10−2 | PCh|GP(monomethyl)E GP(N-biotin)E | |||
| GPE | 1.33 | ||||||
| G1P | 2.84 ** | Pentose and glucuronate interconversions | 1.74 × 10−2 | G1P|X5P | |||
| GPI | 3.35 ** | ||||||
| G6P | 1.38 | SP | |||||
| PCh | 3.01 ** | GPL metabolism | 6.67 × 10−5 | GPC|GPI|GP(dimethyl)E | |||
| PDE4 | 2.31 ** | Ether lipid metabolism | 8.11 × 10−4 | GP(dimethyl)E | |||
| PDE3 | 12.54 ** | Inositol phosphate metabolism | 2.34 × 10−3 | G6P|GPI | |||
| PDE2 | 1.17 | ||||||
| GP(dimethyl)E | 6.86 ** | GPI-anchor biosynthesis | 5.85 × 10−3 | GPI | |||
| GP(N-biotin)E | 1.33 | Regulation of autophagy | 9.73 × 10−3 | GPI | |||
| X5P | 3.14 ** | ||||||
| PDE1 | 1.37 | ||||||
| 6PG | 1.32 | ||||||
| NMPc | 1.26 | ||||||
| AcP | 1.35 | ||||||
| NADPH | 1.50 | ||||||
| GP(monomethyl)E | 1.06 | ||||||
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Serrano, R.; Martin-Hidalgo, D.; Bilbao, J.; Bernardo-Seisdedos, G.; Millet, O.; Garcia-Marin, L.J.; Bragado, M.J. Quantitative Analysis of the Human Semen Phosphorometabolome by 31P-NMR. Int. J. Mol. Sci. 2024, 25, 1682. https://doi.org/10.3390/ijms25031682
Serrano R, Martin-Hidalgo D, Bilbao J, Bernardo-Seisdedos G, Millet O, Garcia-Marin LJ, Bragado MJ. Quantitative Analysis of the Human Semen Phosphorometabolome by 31P-NMR. International Journal of Molecular Sciences. 2024; 25(3):1682. https://doi.org/10.3390/ijms25031682
Chicago/Turabian StyleSerrano, Rebeca, David Martin-Hidalgo, Jon Bilbao, Ganeko Bernardo-Seisdedos, Oscar Millet, Luis J. Garcia-Marin, and Maria Julia Bragado. 2024. "Quantitative Analysis of the Human Semen Phosphorometabolome by 31P-NMR" International Journal of Molecular Sciences 25, no. 3: 1682. https://doi.org/10.3390/ijms25031682