
Harnessing the Stem Cell Niche in Regenerative Medicine: Innovative Avenue to Combat Neurodegenerative Diseases
 , , , ,
, , , ,
Abstract
:1. Introduction
2. Stem Cell Niches: Important Postulations
2.1. Definition
2.2. Stemness and the Niche
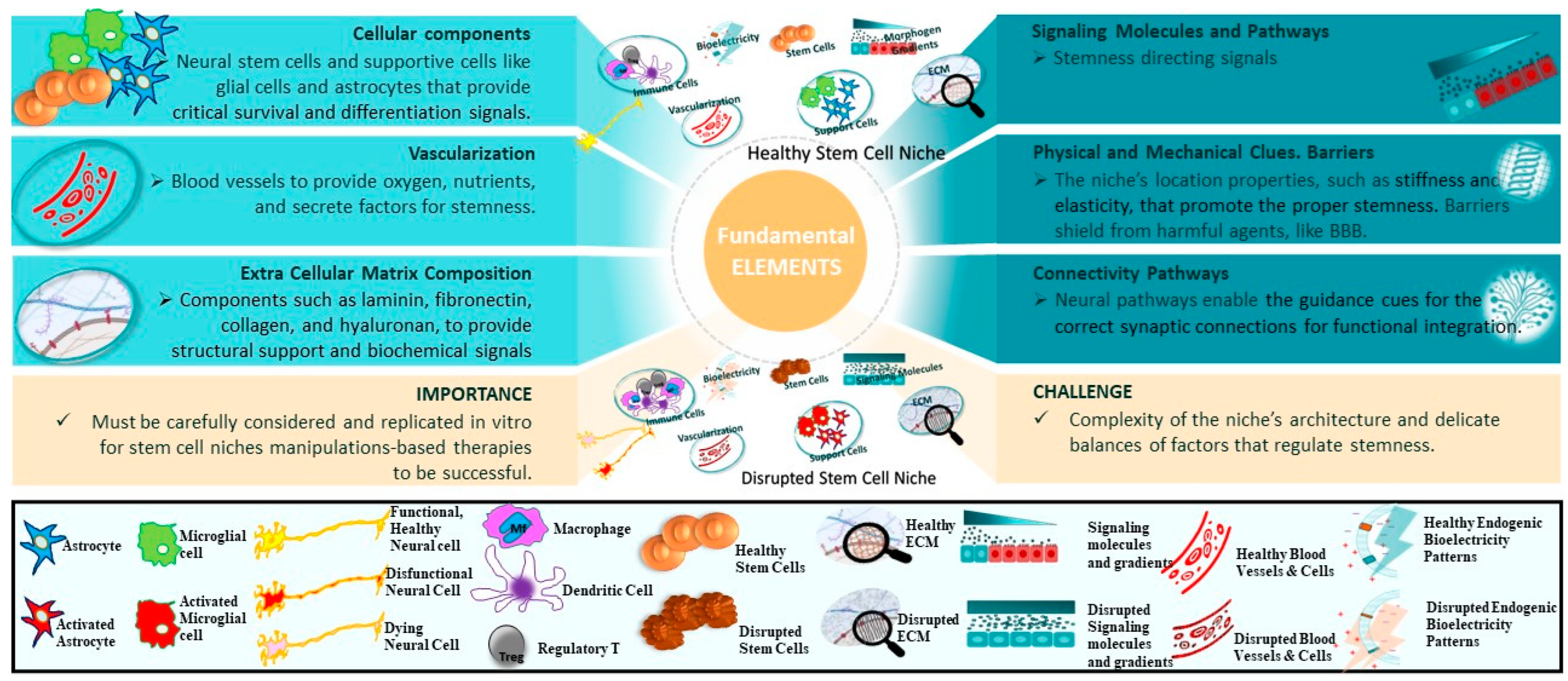
2.3. Elements

2.3.1. Cellular Components
2.3.2. Signaling Molecules
2.3.3. Extracellular Matrix
2.3.4. Blood Vessels
2.3.5. Other Factors
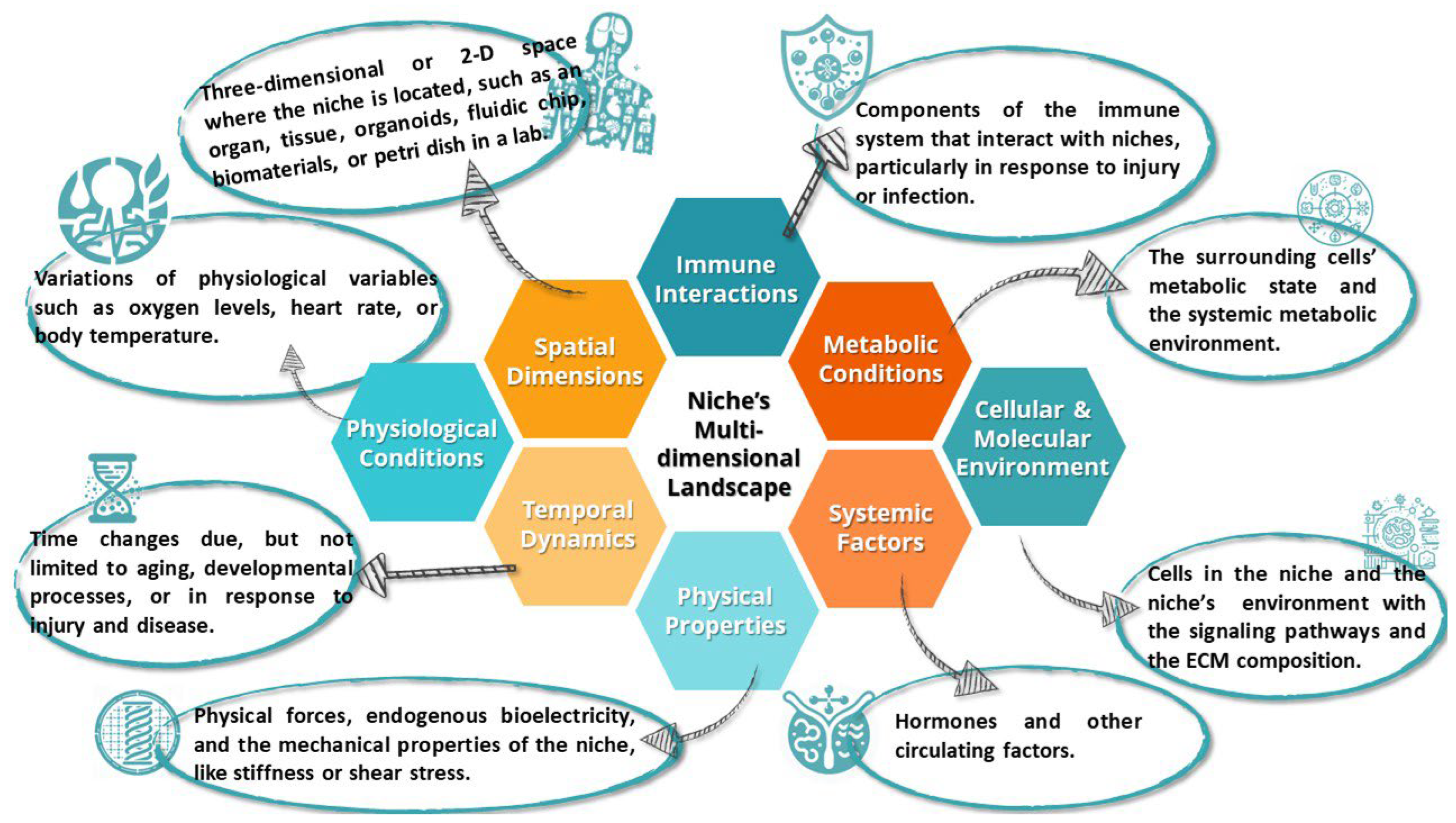
2.4. Location and Dynamics
3. The Multidimensional Landscape of Stem Cell Niches: Interplay, Influences, and Implications Inside and Out
3.1. The Dual Role of Neural Stem Cell Niches: Sustaining Neurogenesis and Contributing to Neurodegenerative Pathologies
3.2. Sophisticated Balance between Immune System and Stem Cell Niche in Neurodegenerative Diseases
3.3. The Intricate Relationship between Natural Stem Cell Niches and Transplanted Stem Cells
3.3.1. Mimicking the Niche for Improved Engraftment
3.3.2. Regulation of Stem Cell Survival, Proliferation, and Differentiation
3.3.3. Promoting Integration with Host Tissue
3.3.4. Avoiding Overgrowth and Tumorigenesis
3.3.5. Modulating the Immune Response
3.3.6. Ex Vivo Niche Recreation
3.3.7. Understanding Stem Cell Homing Mechanisms
3.4. Engineering Artificial Niches through Biomaterials
3.5. Interactions with Induced Stem Cells
4. The Power of the Stem Cell Niche in Advanced Regeneration
4.1. The Role of ACA in Stem Cell Reprogramming and Niche Modulation
4.2. Endogenous Electrical Cues: Directing Stem Cell Dynamics and Tissue Repair
4.3. Morphogenetic Fields: Shaping Niche Dynamics and Stem Cell Behavior
4.4. Decoding Nature’s Masters of Regeneration: Insights into Niche Dynamics and Cellular Renewal
4.5. Cross-Niche Collaborations and Systemic Influences
4.6. Manipulating Epigenetic Signature in Stem Cell Niches and Its Implications for Neurodegenerative Diseases
4.7. Noncoding RNAs: Cell-Specific and Nonspecific Key Gene Regulators Implicated in Neurodegenerative Diseases
4.8. Exosomes—Messengers and Directors
5. From Niche Understanding to Deployment: Tools and Clinical Implications of Stem Cell Niche Dynamics
5.1. Direct In Vivo Reprograming: A Paradigm Shift in Cell Fate Regulation
5.2. Single-Cell Transcriptomics: Unraveling Niche Heterogeneity at Unprecedented Resolution
5.3. 3D Organoids and Microfluidic Models
5.4. Neural Niche Dynamics in Disesases
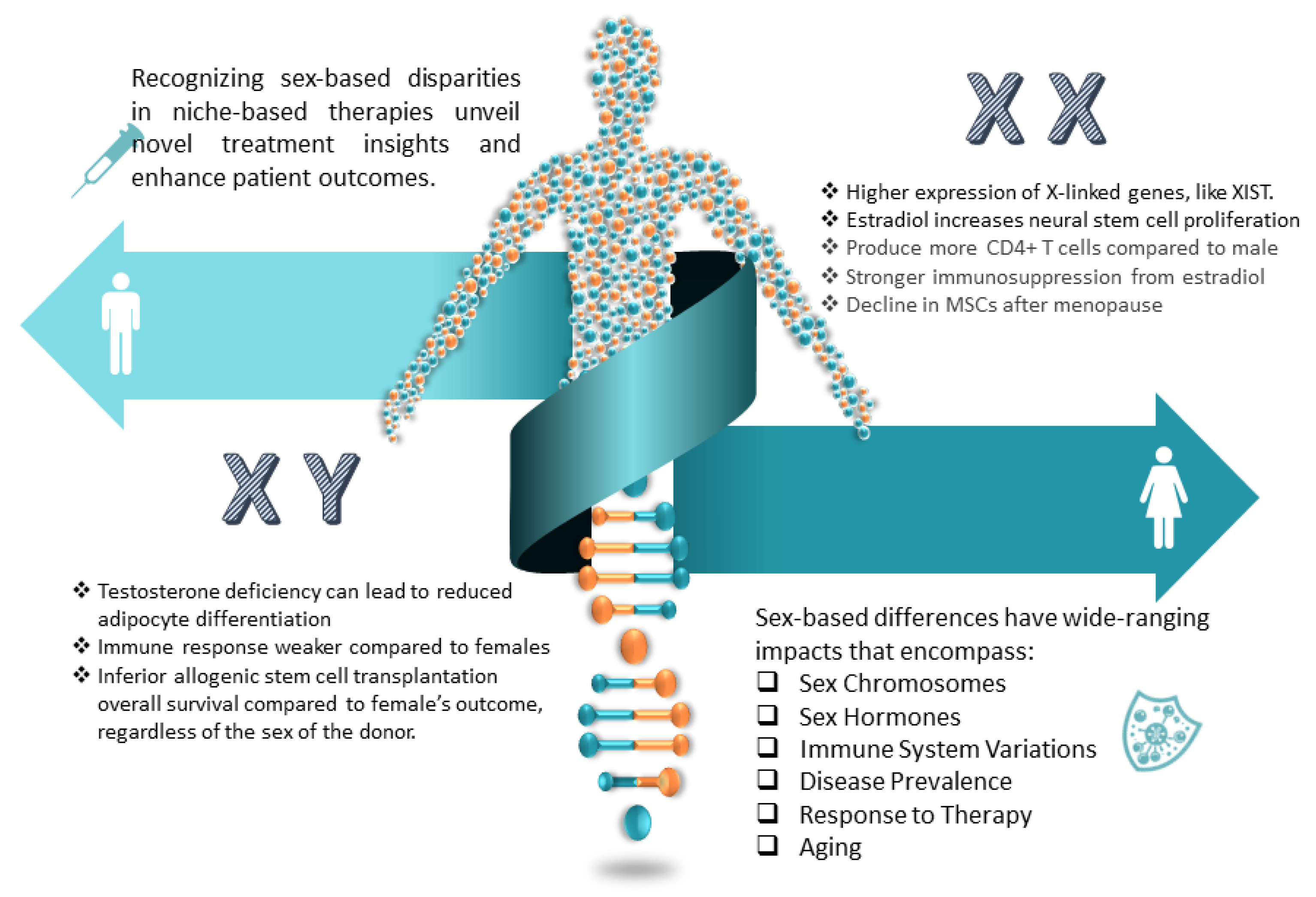
5.4.1. His and Hers: Sex Differences in Neurogenic Niches
5.4.2. Aging: A Need for Rejuvenation
5.4.3. Translational Insights into Stem Cell Niche Disruptions in Neurodegenerative Diseases
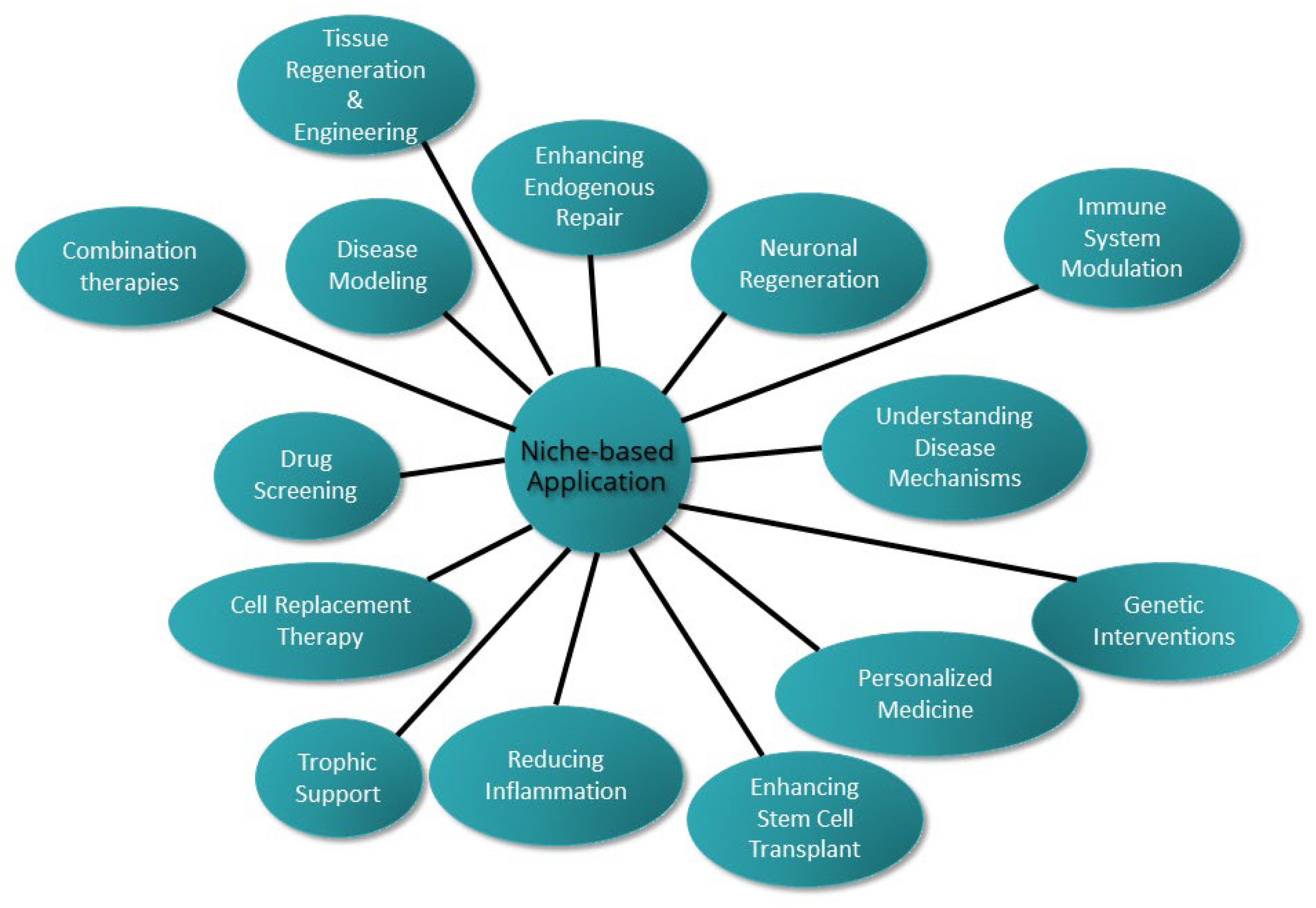
6. Niche Hacking: Breaking the Box
6.1. System-of-Niches Models: A Synergistic Approach for Understanding Neurodegenerative Diseases
6.2. From Bench to Bedside: The Challenges in Translating
7. Deciphering Niche Complexity: The Core Link in Advancing Regenerative Medicine
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Martino, G.; Pluchino, S.; Bonfanti, L.; Schwartz, M. Brain regeneration in physiology and pathology: The immune signature driving therapeutic plasticity of neural stem cells. Physiol. Rev. 2011, 91, 1281–1304. [Google Scholar] [CrossRef] [PubMed]
- Nelson, C.G.; Elta, T.; Bannister, J.; Dzandu, J.; Mangram, A.; Zach, V. Severe Traumatic Brain Injury: A Case Report. Am. J. Case Rep. 2016, 17, 186–191. [Google Scholar] [CrossRef]
- Lindvall, O.; Kokaia, Z. Neurogenesis following Stroke Affecting the Adult Brain. Cold Spring Harb. Perspect. Biol. 2015, 7, a019034. [Google Scholar] [CrossRef] [PubMed]
- Sampson, T.R.; Debelius, J.W.; Thron, T.; Janssen, S.; Shastri, G.G.; Ilhan, Z.E.; Challis, C.; Schretter, C.E.; Rocha, S.; Gradinaru, V.; et al. Gut Microbiota Regulate Motor Deficits and Neuroinflammation in a Model of Parkinson’s Disease. Cell 2016, 167, 1469–1480. [Google Scholar] [CrossRef] [PubMed]
- Lazarov, O.; Marr, R.A. Neurogenesis and Alzheimer’s disease: At the crossroads. Exp. Neurol. 2010, 223, 267–281. [Google Scholar] [CrossRef] [PubMed]
- Kandasamy, M.; Lehner, B.; Kraus, S.; Sander, P.R.; Marschallinger, J.; Rivera, F.J.; Aigner, L. TGF-β signalling in the adult neurogenic niche promotes stem cell quiescence as well as generation of new neurons. J. Cell Mol. Med. 2010, 14, 943–956. [Google Scholar] [CrossRef] [PubMed]
- Itasaki, N.; Hoppler, S. Crosstalk between Wnt and bone morphogenic protein signaling: A turbulent relationship. Dev. Dyn. 2010, 239, 16–33. [Google Scholar] [CrossRef] [PubMed]
- Fekri Aval, S.; Lotfi, H.; Sheervalilou, R.; Zarghami, N.; Mohammadi, H. Tuning of major signaling networks (TGF-β, Wnt, Notch and Hedgehog) by miRNAs in human stem cells commitment to different lineages: Possible clinical application. Biomed. Pharmacother. 2017, 91, 849–860. [Google Scholar] [CrossRef]
- Shen, Q.; Wang, Y.; Kokovay, E.; Lin, G.; Chuang, S.M.; Goderie, S.K.; Roysam, B.; Temple, S. Adult SVZ stem cells lie in a vascular niche: A quantitative analysis of niche cell-cell interactions. Cell Stem Cell 2008, 3, 289–300. [Google Scholar] [CrossRef] [PubMed]
- Voog, J.; Jones, D.L. Stem cells and the niche: A dynamic duo. Cell Stem Cell 2010, 6, 103–115. [Google Scholar] [CrossRef]
- Sonners, J. Hyperbaric Oxygen and Regenerative Medicine: Non-Traditional Uses to Help Reduce Inflammation, Stimulate Cell Regeneration and Improve Healing. Integr. Complement. Ther. 2022, 28, 166–171. [Google Scholar] [CrossRef]
- Mercier, F. Fractones: Extracellular matrix niche controlling stem cell fate and growth factor activity in the brain in health and disease. Cell Mol. Life Sci. 2016, 73, 4661–4674. [Google Scholar] [CrossRef] [PubMed]
- Rundberg Nilsson, A.; Hidalgo, I.; Bryder, D.; Pronk, C.J. Temporal dynamics of TNF-mediated changes in hematopoietic stem cell function and recovery. iScience 2023, 26, 106341. [Google Scholar] [CrossRef]
- Morales-Mantilla, D.E.; King, K.Y. The Role of Interferon-Gamma in Hematopoietic Stem Cell Development, Homeostasis, and Disease. Curr. Stem Cell Rep. 2018, 4, 264–271. [Google Scholar] [CrossRef]
- Inestrosa, N.C.; Varela-Nallar, L. Wnt signaling in the nervous system and in Alzheimer’s disease. J. Mol. Cell Biol. 2014, 6, 64–74. [Google Scholar] [CrossRef] [PubMed]
- Alkailani, M.I.; Aittaleb, M.; Tissir, F. WNT signaling at the intersection between neurogenesis and brain tumorigenesis. Front. Mol. Neurosci. 2022, 15, 1017568. [Google Scholar] [CrossRef]
- Nian, F.-S.; Hou, P.-S. Evolving Roles of Notch Signaling in Cortical Development. Front. Neurosci. 2022, 16, 844410. [Google Scholar] [CrossRef]
- Lampada, A.; Taylor, V. Notch signaling as a master regulator of adult neurogenesis. Front. Neurosci. 2023, 17, 1179011. [Google Scholar] [CrossRef]
- Ferent, J.; Cochard, L.; Faure, H.; Taddei, M.; Hahn, H.; Ruat, M.; Traiffort, E. Genetic Activation of Hedgehog Signaling Unbalances the Rate of Neural Stem Cell Renewal by Increasing Symmetric Divisions. Stem Cell Rep. 2014, 3, 312–323. [Google Scholar] [CrossRef]
- Casas Gimeno, G.; Paridaen, J.T.M. The Symmetry of Neural Stem Cell and Progenitor Divisions in the Vertebrate Brain. Front. Cell Dev. Biol. 2022, 10, 885269. [Google Scholar] [CrossRef]
- Shqirat, M.; Kinoshita, A.; Kageyama, R.; Ohtsuka, T. Sonic hedgehog expands neural stem cells in the neocortical region leading to an expanded and wrinkled neocortical surface. Genes Cells 2021, 26, 399–410. [Google Scholar] [CrossRef] [PubMed]
- Zhang, J.; Uchiyama, J.; Imami, K.; Ishihama, Y.; Kageyama, R.; Kobayashi, T. Novel Roles of Small Extracellular Vesicles in Regulating the Quiescence and Proliferation of Neural Stem Cells. Front. Cell Dev. Biol. 2021, 9, 762293. [Google Scholar] [CrossRef] [PubMed]
- Maric, D.M.; Velikic, G.; Maric, D.L.; Supic, G.; Vojvodic, D.; Petric, V.; Abazovic, D. Stem Cell Homing in Intrathecal Applications and Inspirations for Improvement Paths. Int. J. Mol. Sci. 2022, 23, 4290. [Google Scholar] [CrossRef]
- Ming, G.L.; Song, H. Adult neurogenesis in the mammalian brain: Significant answers and significant questions. Neuron 2011, 70, 687–702. [Google Scholar] [CrossRef] [PubMed]
- Bernstock, J.; Verheyen, J.; Huang, B.; Hallenbeck, J.; Pluchino, S. Typical and Atypical Stem Cell Niches of the Adult Nervous System in Health and Inflammatory Brain and Spinal Cord Diseases. In Adult Stem Cell Niches; InTech: London, UK, 2014. [Google Scholar]
- Llorente, V.; Velarde, P.; Desco, M.; Gómez-Gaviro, M.V. Current Understanding of the Neural Stem Cell Niches. Cells 2022, 11, 3002. [Google Scholar] [CrossRef] [PubMed]
- Schneider, J.; Karpf, J.; Beckervordersandforth, R. Role of Astrocytes in the Neurogenic Niches. In Astrocytes: Methods in Molecular Biology; Di Benedetto, B., Ed.; Humana Press: New York, NY, USA, 2019; Volume 1938. [Google Scholar] [CrossRef]
- Chintamen, S.; Imessadouene, F.; Kernie, S.G. Immune Regulation of Adult Neu-rogenic Niches in Health and Disease. Front. Cell Neurosci. 2021, 14, 571071. [Google Scholar] [CrossRef]
- Lei, Z.; Hu, X.; Wu, Y.; Fu, L.; Lai, S.; Lin, J.; Li, X.; Lv, Y. The Role and Mechanism of the Vascular Endothelial Niche in Diseases: A Review. Front. Physiol. 2022, 13, 863265. [Google Scholar] [CrossRef]
- Riquelme, P.A.; Drapeau, E.; Doetsch, F. Brain micro-ecologies: Neural stem cell niches in the adult mammalian brain. Philos. Trans. R. Soc. Lond. B Biol. Sci. 2008, 363, 123–137. [Google Scholar] [CrossRef]
- Lin, R.; Iacovitti, L. Classic and novel stem cell niches in brain homeostasis and repair. Brain Res. 2015, 1628, 327–342. [Google Scholar] [CrossRef]
- Winner, B.; Kohl, Z.; Gage, F.H. Neurodegenerative disease and adult neurogenesis. Eur. J. Neurosci. 2011, 33, 1139–1151. [Google Scholar] [CrossRef]
- Mu, Y.; Gage, F.H. Adult hippocampal neurogenesis and its role in Alzheimer’s disease. Mol. Neurodegener. 2011, 6, 85. [Google Scholar] [CrossRef] [PubMed]
- Pallotto, M.; Deprez, F. Regulation of adult neurogenesis by GABAergic transmission: Signaling beyond GABAA-receptors. Front. Cell. Neurosci. 2014, 8, 166. [Google Scholar] [CrossRef] [PubMed]
- Dietz, A.; Senf, K.; Karius, J.; Stumm, R.; Neuhaus, E.M. Glia Cells Control Olfactory Neurogenesis by Fine-Tuning CXCL12. Cells 2023, 12, 2164. [Google Scholar] [CrossRef] [PubMed]
- Calsolaro, V.; Edison, P. Neuroinflammation in Alzheimer’s disease: Current evidence and future directions. Alzheimers Dement. 2016, 12, 719–732. [Google Scholar] [CrossRef]
- Becher, B.; Spath, S.; Goverman, J. Cytokine networks in neuroinflammation. Nat. Rev. Immunol. 2017, 17, 49–59. [Google Scholar] [CrossRef]
- Ramesh, G.; MacLean, A.G.; Philipp, M.T. Cytokines and Chemokines at the Crossroads of Neuroinflammation, Neurodegeneration, and Neuropathic Pain. Mediat. Inflamm. 2013, 2013, 480739. [Google Scholar] [CrossRef] [PubMed]
- Koronyo-Hamaoui, M.; Gaire, B.P.; Frautschy, S.A.; Alvarez, J.I. Editorial: Role of Inflammation in Neurodegenerative Diseases. Front. Immunol. 2022, 13, 958487. [Google Scholar] [CrossRef]
- Kwon, H.S.; Koh, S.H. Neuroinflammation in neurodegenerative disorders: The roles of microglia and astrocytes. Transl. Neurodegener. 2020, 9, 42. [Google Scholar] [CrossRef]
- Kerever, A.; Arikawa-Hirasawa, E. Optimal Extracellular Matrix Niches for Neurogenesis: Identifying Glycosaminoglycan Chain Composition in the Subventricular Neurogenic Zone. Front. Neuroanat. 2021, 15, 764458. [Google Scholar] [CrossRef]
- Sun, Y.; Xu, S.; Jiang, M.; Liu, X.; Yang, L.; Bai, Z.; Yang, Q. Role of the Extracellular Matrix in Alzheimer’s Disease. Front. Aging Neurosci. 2021, 13, 707466. [Google Scholar] [CrossRef] [PubMed]
- Falnikar, A.; Stratton, J.; Lin, R.; Andrews, C.; Tyburski, A.; Trovillion, V.; Gottschalk, C.; Ghosh, B.; Iacovitti, L.; Elliott, M.; et al. Differential Response in Novel Stem Cell Niches of the Brain after Cervical Spinal Cord Injury and Traumatic Brain Injury. J. Neurotrauma 2018, 35, 2195–2209. [Google Scholar] [CrossRef]
- Winner, B.; Winkler, J. Adult neurogenesis in neurodegenerative diseases. Cold Spring Harb. Perspect. Biol. 2015, 7, a021287. [Google Scholar] [CrossRef] [PubMed]
- Karakatsani, A.; Shah, B.; Ruiz de Almodovar, C. Blood Vessels as Regulators of Neural Stem Cell Properties. Front. Mol. Neurosci. 2019, 12, 85. [Google Scholar] [CrossRef] [PubMed]
- Peña-Villalobos, I.; Casanova-Maldonado, I.; Lois, P.; Prieto, C.; Pizarro, C.; Lattus, J.; Osorio, G.; Palma, V. Hyperbaric Oxygen Increases Stem Cell Proliferation, Angiogenesis and Wound-Healing Ability of WJ-MSCs in Diabetic Mice. Front. Physiol. 2018, 9, 995. [Google Scholar] [CrossRef] [PubMed]
- MacLaughlin, K.J.; Barton, G.P.; Braun, R.K.; MacLaughlin, J.E.; Lamers, J.J.; Marcou, M.D.; Eldridge, M.W. Hyperbaric air mobilizes stem cells in humans; a new perspective on the hormetic dose curve. Front. Neurol. 2023, 14, 1192793. [Google Scholar] [CrossRef]
- Lindenmann, J.; Kamolz, L.; Graier, W.; Smolle, J.; Smolle-Juettner, F.M. Hyperbaric Oxygen Therapy and Tissue Regeneration: A Literature Survey. Biomedicines 2022, 10, 3145. [Google Scholar] [CrossRef] [PubMed]
- Kempermann, G.; Song, H.; Gage, F.H. Neurogenesis in the adult hippocampus. Cold Spring Harb. Perspect. Biol. 2015, 7, a018812. [Google Scholar]
- Zlokovic, B.V. Neurovascular pathways to neurodegeneration in Alzheimer’s disease and other disorders. Nat. Rev. Neurosci. 2011, 12, 723–738. [Google Scholar] [CrossRef]
- De La Monte, S.M.; Wands, J.R. Alzheimer’s disease is type 3 diabetes-evidence reviewed. J. Diabetes Sci. Technol. 2008, 2, 1101–1113. [Google Scholar] [CrossRef]
- Mas-Bargues, C.; Sanz-Ros, J.; Román-Domínguez, A.; Inglés, M.; Gimeno-Mallench, L.; El Alami, M.; Viña-Almunia, J.; Gambini, J.; Viña, J.; Borrás, C. Relevance of Oxygen Concentration in Stem Cell Culture for Regenerative Medicine. Int. J. Mol. Sci. 2019, 20, 1195. [Google Scholar] [CrossRef] [PubMed]
- Kashiwagi, T.; Takazawa, Y.; Kagawa, T.; Taga, T. Organization of self-advantageous niche by neural stem/progenitor cells during development via autocrine VEGF-A under hypoxia. Inflamm. Regener. 2023, 43, 8. [Google Scholar] [CrossRef] [PubMed]
- Lim, D.A.; Alvarez-Buylla, A. The Adult Ventricular-Subventricular Zone (V-SVZ) and Olfactory Bulb (OB) Neurogenesis. Cold Spring Harb. Perspect. Biol. 2016, 8, a018820. [Google Scholar] [CrossRef] [PubMed]
- Hicks, M.R.; Pyle, A.D. The emergence of the stem cell niche. Trends Cell Biol. 2023, 33, 112–123. [Google Scholar] [CrossRef]
- Petrik, D.; Jörgensen, S.; Eftychidis, V.; Siebzehnrubl, F.A. Singular Adult Neural Stem Cells Do Not Exist. Cells 2022, 11, 722. [Google Scholar] [CrossRef] [PubMed]
- Welner, R.S.; Kincade, P.W. Stem Cells on Patrol. Cell 2007, 131, 842–844. [Google Scholar] [CrossRef] [PubMed]
- Laird, D.J.; von Andrian, U.H.; Wagers, A.J. Stem Cell Trafficking in Tissue Development, Growth, and Disease. Cell 2008, 132, 612–630. [Google Scholar] [CrossRef] [PubMed]
- Sirko, S.; Behrendt, G.; Johansson, P.A.; Tripathi, P.; Costa, M.R.; Bek, S.; Heinrich, C.; Tiedt, S.; Colak, D.; Dichgans, M.; et al. Reactive Glia in the Injured Brain Acquire Stem Cell Properties in Response to Sonic Hedgehog. Cell Stem Cell 2013, 12, 426–439. [Google Scholar] [CrossRef]
- Purro, S.A.; Dickins, E.M.; Salinas, P.C. The secreted Wnt antagonist Dickkopf-1 is required for amyloid β-mediated synaptic loss. J. Neurosci. 2012, 32, 3492–3498. [Google Scholar] [CrossRef]
- Heneka, M.T.; Carson, M.J.; El Khoury, J.; Landreth, G.E.; Brosseron, F.; Feinstein, D.L.; Jacobs, A.H.; Wyss-Coray, T.; Vitorica, J.; Ransohoff, R.M.; et al. Neuroinflammation in Alzheimer’s disease. Lancet Neurol. 2015, 14, 388–405. [Google Scholar] [CrossRef]
- Lindholm, D.; Mäkelä, J.; Di Liberto, V.; Mudò, G.; Belluardo, N.; Eriksson, O.; Saarma, M. Current disease modifying approaches to treat Parkinson’s disease. Cell Mol. Life Sci. 2016, 73, 1365–1379. [Google Scholar] [CrossRef]
- Jurkowski, M.P.; Bettio, L.; Woo, E.K.; Patten, A.; Yau, S.-Y.; Gil-Mohapel, J. Beyond the Hippocampus and the SVZ: Adult Neurogenesis Throughout the Brain. Front. Cell Neurosci. 2020, 14, 576444. [Google Scholar] [CrossRef] [PubMed]
- Katsimpardi, L.; Litterman, N.K. Vascular and neurogenic rejuvenation of the aging mouse brain by young systemic factors. Science 2014, 344, 630–634. [Google Scholar] [CrossRef]
- Villeda, S.A.; Luo, J.; Mosher, K.I.; Zou, B.; Britschgi, M.; Bieri, G.; Stan, T.M.; Fainberg, N.; Ding, Z.; Eggel, A.; et al. The ageing systemic milieu negatively regulates neurogenesis and cognitive function. Nature 2011, 477, 90–94. [Google Scholar] [CrossRef] [PubMed]
- Inestrosa, N.C.; Toledo, E.M. The role of Wnt signaling in neuronal dysfunction in Alzheimer’s Disease. Mol. Neurodegener. 2008, 3, 9. [Google Scholar] [CrossRef] [PubMed]
- Toda, T.; Parylak, S.L.; Linker, S.B.; Gage, F.H. The role of adult hippocampal neurogenesis in brain health and disease. Mol. Psychiatry 2019, 24, 67–87. [Google Scholar] [CrossRef] [PubMed]
- Yang, J.; Liang, X.; Liang, Z.; Li, Y.; Zhang, Y.; Wang, P.; Zhang, C.; Zhang, W.; Lu, Y. GDNF Enhances Therapeutic Efficiency of Neural Stem Cells-Based Therapy in Chronic Experimental Allergic Encephalomyelitis in Rat. Mol. Neurobiol. 2016, 53, 6868–6881. [Google Scholar]
- Ransohoff, R.M.; Brown, M.A. Innate immunity in the central nervous system. J. Clin. Investig. 2012, 122, 1164–1171. [Google Scholar] [CrossRef] [PubMed]
- Ekdahl, C.T.; Claasen, J.-H.; Bonde, S.; Kokaia, Z.; Lindvall, O. Inflammation is detrimental for neurogenesis in adult brain. Proc. Natl. Acad. Sci. USA 2003, 100, 13632–13637. [Google Scholar] [CrossRef]
- Gemma, C.; Bachstetter, A.D. The role of microglia in adult hippocampal neurogenesis. Front. Cell Neurosci. 2013, 7, 229. [Google Scholar] [CrossRef]
- Sheng, J.; Chen, D.; Van Laar, V.S.; Li, Z.; Prabhakaran, V.; Patel, M.; Xie, Y.; Zhang, H.; Alexander, J.C.; Kozlowski, M.T.; et al. Human stem cell-derived astrocytes replicate human prions in a PRNP genotype-dependent manner. J. Exp. Med. 2015, 212, 351–367. [Google Scholar]
- Russo, I.; Barlati, S.; Bosetti, F. Effects of neuroinflammation on the regenerative capacity of brain stem cells. J. Neurochem. 2011, 116, 947–956. [Google Scholar] [CrossRef]
- Cho, S.-H.; Sun, B.; Zhou, Y.; Kauppinen, T.M.; Halabisky, B.; Wes, P.; Ransohoff, R.M.; Gan, L. CX3CR1 protein signaling modulates microglial activation and protects against plaque-independent cognitive deficits in a mouse model of Alzheimer disease. J. Biol. Chem. 2011, 286, 32713–32722. [Google Scholar] [CrossRef]
- Faridar, A.; Vasquez, M.; Thome, A.D.; Yin, Z.; Xuan, H.; Wang, J.H.; Wen, S.; Li, X.; Thonhoff, J.R.; Zhao, W.; et al. Ex Vivo Expanded Human Regulatory T Cells Modify Neuroinflammation in a Preclinical Model of Alzheimer’s Disease. Acta Neuropathol. Commun. 2022, 10, 144. [Google Scholar] [CrossRef]
- Leyns, C.E.G.; Holtzman, D.M. Glial Contributions to Neurodegeneration in Tauopathies. Mol. Neurodegener. 2017, 12, 50. [Google Scholar] [CrossRef]
- Vandervelde, S.; van Luyn, M.J.A.; Tio, R.A.; Harmsen, M.C. Signaling factors in stem cell-mediated repair of infarcted myocardium. J. Mol. Cell Cardiol. 2005, 39, 363–376. [Google Scholar] [CrossRef]
- Willerth, S.M.; Sakiyama-Elbert, S.E. Approaches to neural tissue engineering using scaffolds for drug delivery. Adv. Drug Deliv. Rev. 2007, 59, 325–338. [Google Scholar] [CrossRef]
- Shen, Q.; Goderie, S.K.; Jin, L.; Karanth, N.; Sun, Y.; Abramova, N.; Vincent, P.; Pumiglia, K.; Temple, S. Endothelial cells stimulate self-renewal and expand neurogenesis of neural stem cells. Science 2004, 304, 1338–1340. [Google Scholar] [CrossRef]
- Watt, F.M.; Hogan, B.L.M. Out of Eden: Stem cells and their niches. Science 2000, 287, 1427–1430. [Google Scholar] [CrossRef] [PubMed]
- Morrison, S.J.; Spradling, A.C. Stem cells and niches: Mechanisms that promote stem cell maintenance throughout life. Cell 2008, 132, 598–611. [Google Scholar] [CrossRef]
- Williams, C.A.; Lavik, E.B. Engineering the CNS stem cell microenvironment. Regen. Med. 2009, 4, 865–877. [Google Scholar] [CrossRef] [PubMed]
- Magnusson, J.P.; Zamboni, M.; Santopolo, G.; Mold, J.E.; Barrientos-Somarribas, M.; Talavera-Lopez, C.; Andersson, B.; Frisén, J. Activation of a neural stem cell transcriptional program in parenchymal astrocytes. ELife 2020, 9, e59733. [Google Scholar] [CrossRef]
- Beyer, F.; Samper Agrelo, I.; Küry, P. Do Neural Stem Cells Have a Choice? Heterogenic Outcome of Cell Fate Acquisition in Different Injury Models. Int. J. Mol. Sci. 2019, 20, 455. [Google Scholar] [CrossRef]
- Lim, D.A.; Alvarez-Buylla, A. Noggin antagonizes BMP signaling to create a niche for adult neurogenesis. Neuron 2000, 28, 713–726. [Google Scholar] [CrossRef] [PubMed]
- Fujisaki, J.; Wu, J.; Carlson, A.L.; Silberstein, L.; Putheti, P.; Larocca, R.; Gao, W.; Saito, T.I.; Lo Celso, C.; Tsuyuzaki, H.; et al. In vivo imaging of Treg cells providing immune privilege to the haematopoietic stem-cell niche. Nature 2011, 474, 216–219. [Google Scholar] [CrossRef] [PubMed]
- Peerani, R.; Zandstra, P.W. Enabling stem cell therapies through synthetic stem cell-niche engineering. J. Clin. Investig. 2010, 120, 60–70. [Google Scholar] [CrossRef]
- Morrison, S.J.; Scadden, D.T. The bone marrow niche for haematopoietic stem cells. Nature 2014, 505, 327–334. [Google Scholar] [CrossRef]
- Chaker, Z.; Codega, P.; Doetsch, F. A mosaic world: Puzzles revealed by adult neural stem cell heterogeneity. WIREs Dev. Biol. 2016, 5, 640–658. [Google Scholar] [CrossRef] [PubMed]
- Sugiyama, T.; Kohara, H.; Noda, M.; Nagasawa, T. Maintenance of the hematopoietic stem cell pool by CXCL12-CXCR4 chemokine signaling in bone marrow stromal cell niches. Immunity 2006, 25, 977–988. [Google Scholar] [CrossRef] [PubMed]
- Schantz, J.-T.; Chim, H.; Whiteman, M. Cell guidance in tissue engineering: SDF-1 mediates site-directed homing of mesenchymal stem cells within three-dimensional polycaprolactone scaffolds. Tissue Eng. 2007, 13, 2615–2624. [Google Scholar] [CrossRef]
- Wang, Y.; Yao, M.; Zhou, J.; Zheng, W.; Zhou, C.; Dong, D.; Liu, Y.; Teng, Z.; Jiang, Y.; Wei, G.; et al. The promotion of neural progenitor cells proliferation by aligned and randomly oriented collagen nanofibers through β1 integrin/MAPK signaling pathway. Biomaterials 2011, 32, 6737–6744. [Google Scholar] [CrossRef] [PubMed]
- Mahoney, M.J.; Anseth, K.S. Three-dimensional growth and function of neural tissue in degradable polyethylene glycol hydrogels. Biomaterials 2006, 27, 2265–2274. [Google Scholar] [CrossRef] [PubMed]
- De Witte, T.M.; Bramono, D.S.; Lamontagne, C.A.; Evseenko, D.N.; Tuan, R.S. Bone tissue engineering via growth factor delivery: From scaffolds to complex matrices. Regen. Biomater. 2018, 5, 197–211. [Google Scholar] [CrossRef]
- Lu, X.; Guo, H.; Li, J.; Sun, T.; Xiong, M. Recombinant Human Bone Morphogenic Protein-2 Immobilized Fabrication of Magnesium Functionalized Injectable Hydrogels for Controlled-Delivery and Osteogenic Differentiation of Rat Bone Marrow-Derived Mesenchymal Stem Cells in Femoral Head Necrosis Repair. Front. Cell Dev. Biol. 2021, 9, 723789. [Google Scholar] [CrossRef]
- Bible, E.; Chau, D.Y.S.; Alexander, M.R.; Price, J.; Shakesheff, K.M.; Modo, M. The support of neural stem cells transplanted into stroke-induced brain cavities by PLGA particles. Biomaterials 2009, 30, 2985–2994. [Google Scholar] [CrossRef]
- Zhao, Y.; Rafatian, N.; Feric, N.T.; Cox, B.J.; Aschar-Sobbi, R.; Wang, E.Y.; Aggarwal, P.; Zhang, B.; Conant, G.; Ronaldson-Bouchard, K.; et al. A Platform for Generation of Chamber-Specific Cardiac Tissues and Disease Modeling. Cell 2019, 176, 913–927.e18. [Google Scholar] [CrossRef] [PubMed]
- Takahashi, K.; Yamanaka, S. Induction of pluripotent stem cells from mouse embryonic and adult fibroblast cultures by defined factors. Cell 2006, 126, 663–676. [Google Scholar] [CrossRef] [PubMed]
- Kondo, T.; Asai, M.; Tsukita, K.; Kutoku, Y.; Ohsawa, Y.; Sunada, Y.; Imamura, K.; Egawa, N.; Yahata, N.; Okita, K.; et al. Modeling Alzheimer’s disease with iPSCs reveals stress phenotypes associated with intracellular Aβ and differential drug responsiveness. Cell Stem Cell 2013, 12, 487–496. [Google Scholar] [CrossRef]
- Kikuchi, T.; Morizane, A.; Doi, D.; Magotani, H.; Onoe, H.; Hayashi, T.; Mizuma, H.; Takara, S.; Takahashi, R.; Inoue, H.; et al. Human iPS cell-derived dopaminergic neurons function in a primate Parkinson’s disease model. Nature 2017, 548, 592–596. [Google Scholar] [CrossRef]
- Chen, Y.; Blair, K.; Smith, A. Robust self-renewal of rat embryonic stem cells requires fine-tuning of glycogen synthase kinase-3 inhibition. Stem Cell Rep. 2013, 1, 209–217. [Google Scholar] [CrossRef]
- Lee, A.; Tang, C.; Rao, M.; Cayrou, C.; Jiang, E.; Helms, J.A.; Chang, S.; Cui, X.; Crabtree, G.R. Tumorigenicity as a clinical hurdle for pluripotent stem cell therapies. Nat. Med. 2013, 19, 998–1004. [Google Scholar] [CrossRef]
- Daley, G.Q. Stem cells: Roadmap to the clinic. J. Clin. Investig. 2010, 120, 8–10. [Google Scholar] [CrossRef]
- Scadden, D.T. The stem-cell niche as an entity of action. Nature 2006, 441, 1075–1079. [Google Scholar] [CrossRef] [PubMed]
- Lane, S.W.; Williams, D.A.; Watt, F.M. Modulating the stem cell niche for tissue regeneration. Nat. Biotechnol. 2014, 32, 795–803. [Google Scholar] [CrossRef] [PubMed]
- Murphy, W.L.; McDevitt, T.C.; Engler, A.J. Materials as stem cell regulators. Nat. Mater. 2014, 13, 547–557. [Google Scholar] [CrossRef] [PubMed]
- Marturano-Kruik, A.; Nava, M.M.; Yeager, K.; Chramiec, A.; Hao, L.; Robinson, S.; Guo, E.; Raimondi, M.T.; Vunjak-Novakovic, G. Human bone perivascular niche-on-a-chip for studying metastatic colonization. Proc. Natl. Acad. Sci. USA 2018, 115, 1256–1261. [Google Scholar] [CrossRef]
- Becker-Kojic, Z.A. GPI Anchored Glycoprotein ACA as a Novel Tumor Marker. Patent EP1391463A1, 25 February 2004. [Google Scholar]
- Becker-Kojić, Z.A.; Terness, P. A Novel Human Erythrocyte Glycosylphosphatidylinositol (GPI)-anchored Glycoprotein ACA. J. Biol. Chem. 2002, 277, 40472–40478. [Google Scholar] [CrossRef] [PubMed]
- Becker-Kojić, Z.A.; Ureña-Peralta, J.R.; Zipančić, I.; Rodriguez-Jiménez, F.J.; Rubio, M.P.; Stojković, P.; Roselló, M.G.; Stojković, M. A Novel Human Glycoprotein ACA is an Upstream Regulator of Human Hematopoiesis. Cell Technol. Biol. Med. 2013, 2, 69–84. [Google Scholar] [CrossRef]
- Becker-Kojić, Z.A.; Ureña-Peralta, J.R.; Zipančić, I.; Rodriguez-Jiménez, F.J.; Rubio, M.P.; Stojković, P.; Roselló, M.G.; Stojković, M. Activation by ACA Induces Pluripotency in Human Blood Progenitor Cells. Cell Technol. Biol. Med. 2013, 2, 85–101. [Google Scholar] [CrossRef]
- Sun, Y.S. Studying Electrotaxis in Microfluidic Devices. Sensors 2017, 17, 2048. [Google Scholar] [CrossRef]
- Mehta, A.S.; Ha, P.; Zhu, K.; Li, S.Y.; Ting, K.; Soo, C.; Zhang, X.; Zhao, M. Physiological electric fields induce directional migration of mammalian cranial neural crest cells. Dev. Biol. 2021, 471, 97–105. [Google Scholar] [CrossRef]
- Kamimura, Y.; Mogilner, A.; Devreotes, P.N.; Zhao, M. Different Roles of Membrane Potentials in Electrotaxis and Chemotaxis of Dictyostelium Cells. Eukaryot. Cell 2011, 10, 1251–1256. [Google Scholar]
- Lange, F.; Venus, J.; Shams Esfand Abady, D.; Porath, K.; Einsle, A.; Sellmann, T.; Neubert, V.; Reichart, G.; Linnebacher, M.; Köhling, R. Galvanotactic Migration of Glioblastoma and Brain Metastases Cells. Life 2022, 12, 580. [Google Scholar] [CrossRef] [PubMed]
- Huang, Y.J.; Hoffmann, G.; Wheeler, B.; Schäfer, R.; Barrett, C.; Rudolph, M.; Günther, S.; Yondorf, M.; Koch, D.; Schröder, T.; et al. Cellular microenvironment modulates the galvanotaxis of brain tumor initiating cells. Sci. Rep. 2016, 6, 21583. [Google Scholar] [CrossRef] [PubMed]
- Allen, G.M.; Mogilner, A.; Theriot, J.A. Electrophoresis of cellular membrane components creates the directional cue guiding keratocyte galvanotaxis. Curr. Biol. 2013, 23, 560–568. [Google Scholar] [CrossRef]
- Brockes, J.P.; Kumar, A. Comparative aspects of animal regeneration. Annu. Rev. Cell Dev. Biol. 2008, 24, 525–549. [Google Scholar] [CrossRef] [PubMed]
- Kumar, A.; Brockes, J.P. Nerve dependence in tissue, organ, and appendage regeneration. Trends Neurosci. 2012, 35, 691–699. [Google Scholar] [CrossRef]
- Beane, W.S.; Morokuma, J.; Lemire, J.M.; Levin, M. Bioelectric signaling regulates head and organ size during planarian regeneration. Development 2013, 140, 313–322. [Google Scholar] [CrossRef]
- Chernet, B.T.; Levin, M. Transmembrane voltage potential of somatic cells controls oncogene-mediated tumorigenesis at long-range. Oncotarget 2014, 5, 3287–3306. [Google Scholar] [CrossRef]
- Nuccitelli, R. A role for endogenous electric fields in wound healing. Curr. Top. Dev. Biol. 2003, 58, 1–26. [Google Scholar]
- Rodriguez, G.; Arnaldi, D.; Picco, A. Brain Functional Network in Alzheimer’s Disease: Diagnostic Markers for Diagnosis and Monitoring. Int. J. Alzheimers Dis. 2011, 2011, 481903. [Google Scholar] [CrossRef] [PubMed]
- Ning, S.; Jorfi, M.; Patel, S.R.; Kim, D.Y.; Tanzi, R.E. Neurotechnological Approaches to the Diagnosis and Treatment of Alzheimer’s Disease. Front. Neurosci. 2022, 16, 854992. [Google Scholar] [CrossRef]
- Karumbaiah, L.; Saxena, T.; Carlson, D.; Patil, K.; Patkar, R.; Gaupp, E.A.; Betancur, M.; Stanley, G.B.; Carin, L.; Bellamkonda, R.V. Relationship between intracortical electrode design and chronic recording function. Biomaterials 2013, 34, 8061–8074. [Google Scholar] [CrossRef] [PubMed]
- Urrego, D.; Tomczak, A.P.; Zahed, F.; Stühmer, W.; Pardo, L.A. Potassium channels in cell cycle and cell proliferation. Philos. Trans. R. Soc. B Biol. Sci. 2014, 369, 20130094. [Google Scholar] [CrossRef] [PubMed]
- Mengchu, C.; Hongfei, G.; Zhao, H.; Yongjie, Z.; Yujie, C.; Hua, F. Electromagnetic Fields for the Regulation of Neural Stem Cells. Stem Cells Int. 2017, 2017, 9898439. [Google Scholar]
- McNeill, J.; Rudyk, C.; Hildebrand, M.E.; Salmaso, N. Ion Channels and Electrophysiological Properties of Astrocytes: Implications for Emergent Stimulation Technologies. Front. Cell Neurosci. 2021, 15, 644126. [Google Scholar] [CrossRef] [PubMed]
- Citri, A.; Malenka, R.C. Synaptic plasticity: Multiple forms, functions, and mechanisms. Neuropsychopharmacology 2008, 33, 18–41. [Google Scholar] [CrossRef] [PubMed]
- Fregni, F.; Pascual-Leone, A. Technology insight: Noninvasive brain stimulation in neurology-perspectives on the therapeutic potential of rTMS and tDCS. Nat. Clin. Pract. Neurol. 2007, 3, 383–393. [Google Scholar] [CrossRef]
- Wu, A.D.; Fregni, F.; Simon, D.K.; Deblieck, C.; Pascual-Leone, A. Noninvasive brain stimulation for Parkinson’s disease and dystonia. Neurotherapeutics 2008, 5, 345–361. [Google Scholar] [CrossRef]
- Spagnuolo, G.; Genovese, F.; Fortunato, L.; Simeone, M.; Rengo, C.; Tatullo, M. The Impact of Optogenetics on Regenerative Medicine. Appl. Sci. 2020, 10, 173. [Google Scholar] [CrossRef]
- Albrecht, S.; Grothe, C.; Pistorius, A.; Braun, C.; Marschallinger, J.; Adler, K.; Meissirel, C.; Götz, M.; Markl, J. Tracking stem cell differentiation in the setting of automated optogenetic stimulation. Stem Cells 2011, 29, 78–88. [Google Scholar]
- Ma, T.; Ding, Q.; Liu, C.; Zhang, Z.J.; Shao, L.; Chen, J.; Gao, J.; Gu, N.; Zhang, X. Electromagnetic fields regulate calcium-mediated cell fate of stem cells: Osteogenesis, chondrogenesis and apoptosis. Stem Cell Res. Ther. 2023, 14, 133. [Google Scholar] [CrossRef] [PubMed]
- Rong, Z.; Sun, Z.; Li, C.; Ramakrishna, S.; Chiu, K.; He, L. Electrical stimulation affects neural stem cell fate and function in vitro. Exp. Neurol. 2019, 319, 112963. [Google Scholar]
- Moschetta, M.; Chiacchiaretta, M.; Cesca, F.; Roy, I.; Athanassiou, A.; Benfenati, F.; Papadopoulou, E.L.; Bramini, M. Graphene Nanoplatelets Render Poly(3-Hydroxybutyrate) a Suitable Scaffold to Promote Neuronal Network Development. Front. Neurosci. 2021, 15, 731198. [Google Scholar] [CrossRef] [PubMed]
- Cheng, H.; Huang, Y.; Yue, H.; Fan, Y. Electrical Stimulation Promotes Stem Cell Neural Differentiation in Tissue Engineering. Stem Cells Int. 2021, 2021, 6697574. [Google Scholar] [CrossRef] [PubMed]
- Liu, Q.; Telezhkin, V.; Jiang, W.; Marshall, S.; Yung, S.; Woodhall, G.L.; Leigh, P.N.; Brown, R.; Sinden, J.D.; Riccardi, D.; et al. Electric field stimulation boosts neuronal differentiation of neural stem cells for spinal cord injury treatment via PI3K/Akt/GSK-3β/β-catenin activation. Cell Biosci. 2023, 13, 4. [Google Scholar] [CrossRef] [PubMed]
- Cheng, H.; Huang, Y.; Chen, W.; Che, J.; Liu, T.; Na, J.; Wang, R.; Fan, Y. Cyclic Strain and Electrical Co-stimulation Improve Neural Differentiation of Marrow-Derived Mesenchymal Stem Cells. Front. Cell Dev. Biol. 2021, 9, 624755. [Google Scholar] [CrossRef]
- Maziarz, A.; Kocan, B.; Bester, M.; Budzik, S.; Cholewa, M.; Ochiya, T.; Banas, A. How electromagnetic fields can influence adult stem cells: Positive and negative impacts. Stem Cell Res. Ther. 2016, 7, 54. [Google Scholar] [CrossRef]
- Gurdon, J.B.; Harger, P.; Mitchell, A.; Lemaire, P. Activin signalling and response to a morphogen gradient. Nature 1994, 371, 487–492. [Google Scholar] [CrossRef]
- Dessaud, E.; McMahon, A.P.; Briscoe, J. Pattern formation in the vertebrate neural tube: A sonic hedgehog morphogen-regulated transcriptional network. Development 2008, 135, 2489–2503. [Google Scholar] [CrossRef]
- Zon, L.I. Intrinsic and extrinsic control of haematopoietic stem-cell self-renewal. Nature 2008, 453, 306–313. [Google Scholar] [CrossRef]
- Gudas, L.J.; Wagner, J.A. Retinoids regulate stem cell differentiation. J. Cell Physiol. 2011, 226, 322–330. [Google Scholar] [CrossRef]
- Grace, C.S.; Mikkola, H.K.A.; Dou, D.R.; Calvanese, V.; Rönn, R.E.; Purton, L.E. Protagonist or antagonist? The complex roles of retinoids in the regulation of hematopoietic stem cells and their specification from pluripotent stem cells. Exp. Hematol. 2018, 65, 1–16. [Google Scholar] [CrossRef] [PubMed]
- Mezquita, B.; Mezquita, C. Two Opposing Faces of Retinoic Acid: Induction of Stemness or Induction of Differentiation Depending on Cell-Type. Biomolecules 2019, 9, 567. [Google Scholar] [CrossRef]
- Cabezas-Wallscheid, N.; Buettner, F.; Sommerkamp, P.; Klimmeck, D.; Ladel, L.; Thalheimer, F.B.; Pastor-Flores, D.; Roma, L.P.; Renders, S.; Zeisberger, P.; et al. Vitamin A-Retinoic Acid Signaling Regulates Hematopoietic Stem Cell Dormancy. Cell 2017, 169, 807–823. [Google Scholar] [CrossRef] [PubMed]
- McCusker, C.D.; Gardiner, D.M. Understanding positional cues in salamander limb regeneration: Implications for optimizing cell-based regenerative therapies. Dis. Models Mech. 2014, 7, 593–599. [Google Scholar] [CrossRef] [PubMed]
- Pathak, M.M.; Nourse, J.L.; Tran, T.; Hwe, J.; Arulmoli, J.; Le, D.T.; Bernardis, E.; Flanagan, L.A.; Tombola, F. Stretch-activated ion channel Piezo1 directs lineage choice in human neural stem cells. Proc. Natl. Acad. Sci. USA 2014, 111, 16148–16153. [Google Scholar] [CrossRef] [PubMed]
- Ambler, C.A.; Watt, F.M. Adult epidermal Notch activity induces dermal accumulation of T cells and neural crest derivatives through upregulation of jagged 1. Development 2010, 137, 3569–3579. [Google Scholar] [CrossRef] [PubMed]
- Szarek, D.; Marycz, K.; Lis, A.; Zawada, Z.; Tabakow, P.; Laska, J.; Jarmundowicz, W. Lizard tail spinal cord: A new experimental model of spinal cord injury without limb paralysis. FASEB J. 2016, 30, 1391–1403. [Google Scholar] [CrossRef] [PubMed]
- Alibardi, L. Introduction to the Study on Regeneration in Lizards as an Amniote Model of Organ Regeneration. J. Dev. Biol. 2021, 9, 51. [Google Scholar] [CrossRef]
- Tokuyama, M.A.; Xu, C.; Fisher, R.E.; Wilson-Rawls, J.; Kusumi, K.; Newbern, J.M. Developmental and adult-specific processes contribute to de novo neuromuscular regeneration in the lizard tail. Dev. Biol. 2018, 433, 287–296. [Google Scholar] [CrossRef] [PubMed]
- Ross Stewart, K.M.; Walker, S.L.; Baker, A.H.; Riley, P.R.; Brittan, M. Hooked on heart regeneration: The zebrafish guide to recovery. Cardiovasc. Res. 2022, 118, 1667–1679. [Google Scholar] [CrossRef]
- Roberts-Galbraith, R.H.; Newmark, P.A. On the organ trail: Insights into organ regeneration in the planarian. Curr. Opin. Genet. Dev. 2015, 32, 37–46. [Google Scholar] [CrossRef]
- McCusker, C.; Bryant, S.V.; Gardiner, D.M. The axolotl limb blastema: Cellular and molecular mechanisms driving blastema formation and limb regeneration in tetrapods. Regeneration 2015, 2, 54–71. [Google Scholar] [CrossRef] [PubMed]
- Tseng, S.; Beane, W.S.; Lemire, J.M.; Masi, A.; Levin, M. Induction of vertebrate regeneration by a transient sodium current. J. Neurosci. 2010, 30, 13192–13200. [Google Scholar] [CrossRef]
- Amamoto, R.; Lopez Huerta, V.G.; Takahashi, E.; Dai, G.; Grant, A.K.; Fu, Z.; Arlotta, P. Adult axolotls can regenerate original neuronal diversity in response to brain injury. eLife 2016, 5, e13998. [Google Scholar] [CrossRef]
- Dimou, L.; Götz, M. Glial cells as progenitors and stem cells: New roles in the healthy and diseased brain. Physiol. Rev. 2014, 94, 709–737. [Google Scholar] [CrossRef] [PubMed]
- Khyeam, S.; Lee, S.; Huang, G.N. Genetic, epigenetic, and post-transcriptional basis of divergent tissue regenerative capacities among vertebrates. Adv. Genet. 2021, 2, e10042. [Google Scholar] [CrossRef] [PubMed]
- Encinas, J. Radial glia and radial glia-like cells: Their role in neurogenesis and regeneration. Front. Neurosci. 2022, 16, 1006037. [Google Scholar]
- Cryan, J.F.; O’Riordan, K.J.; Cowan, C.S.M.; Sandhu, K.V.; Bastiaanssen, T.F.S.; Boehme, M.; Codagnone, M.G.; Cussotto, S.; Fulling, C.; Golubeva, A.V.; et al. The Microbiota-Gut-Brain Axis. Physiol. Rev. 2019, 99, 1877–2013. [Google Scholar] [CrossRef] [PubMed]
- Yoo, B.B.; Mazmanian, S.K. The Enteric Network: Interactions between the Immune and Nervous Systems of the Gut. Immunity 2017, 46, 910. [Google Scholar] [CrossRef]
- Bátiz, L.F.; Castro, M.A.; Burgos, P.V.; Velásquez, Z.D.; Muñoz, R.I.; Lafourcade, C.A.; Troncoso-Escudero, P.; Wyneken, U. Exosomes as Novel Regulators of Adult Neurogenic Niches. Front. Cell Neurosci. 2016, 9, 501. [Google Scholar] [CrossRef] [PubMed]
- Sweeney, M.D.; Kisler, K.; Montagne, A.; Toga, A.W.; Zlokovic, B.V. The role of brain vasculature in neurodegenerative disorders. Nat. Neurosci. 2018, 21, 1318. [Google Scholar] [CrossRef]
- Müller, S.J.; Teckentrup, V.; Rebollo, I.; Hallschmid, M.; Kroemer, N.B. Vagus nerve stimulation increases stomach-brain coupling via a vagal afferent pathway. Brain Stimul. 2022, 15, 1279–1289. [Google Scholar] [CrossRef]
- Zhou, H.; Wang, B.; Sun, H.; Xu, X.; Wang, Y. Epigenetic regulations in neural stem cells and neurological diseases. Stem Cells Int. 2018, 2018, 6087143. [Google Scholar] [CrossRef]
- Meissner, A.; Mikkelsen, T.S.; Gu, H.; Wernig, M.; Hanna, J.; Sivachenko, A.; Zhang, X.; Bernstein, B.E.; Nusbaum, C.; Jaffe, D.B.; et al. Genome-scale DNA methylation maps of pluripotent and differentiated cells. Nature 2008, 454, 766–770. [Google Scholar] [CrossRef]
- Chouliaras, L.; Rutten, B.P.; Kenis, G.; Peerbooms, O.; Visser, P.J.; Verhey, F.; van Os, J.; Steinbusch, H.W.; van den Hove, D.L. Epigenetic regulation in the pathophysiology of Alzheimer’s disease. Prog. Neurobiol. 2010, 90, 498–510. [Google Scholar] [CrossRef] [PubMed]
- Gräff, J.; Rei, D.; Guan, J.S.; Wang, W.Y.; Seo, J.; Hennig, K.M.; Nieland, T.J.; Fass, D.M.; Kao, P.F.; Kahn, M.; et al. An epigenetic blockade of cognitive functions in the neurodegenerating brain. Nature 2012, 483, 222–226. [Google Scholar] [CrossRef] [PubMed]
- Coppedè, F. Genetics and epigenetics of Parkinson’s disease. Sci. World J. 2012, 2012, 489830. [Google Scholar] [CrossRef]
- Kilgore, M.; Miller, C.A.; Fass, D.M.; Hennig, K.M.; Haggarty, S.J.; Sweatt, J.D.; Rumbaugh, G. Inhibitors of class 1 histone deacetylases reverse contextual memory deficits in a mouse model of Alzheimer’s disease. Neuropsychopharmacology 2010, 35, 870–880. [Google Scholar] [CrossRef] [PubMed]
- Qiu, L.; Tan, E.K.; Zeng, L. MicroRNAs and neurodegenerative diseases. Adv. Exp. Med. Biol. 2015, 888, 85–105. [Google Scholar] [PubMed]
- Nguyen, T.P.N.; Kumar, M.; Fedele, E.; Bonanno, G.; Bonifacino, T. MicroRNA Alteration, Application as Biomarkers, and Therapeutic Approaches in Neurodegenerative Diseases. Int. J. Mol. Sci. 2022, 23, 4718. [Google Scholar] [CrossRef] [PubMed]
- Panio, A.; Cava, C.; D’Antona, S.; Bertoli, G.; Porro, D. Diagnostic Circulating miRNAs in Sporadic Amyotrophic Lateral Sclerosis. Front. Med. 2022, 9, 861960. [Google Scholar] [CrossRef] [PubMed]
- Miñones-Moyano, E.; Porta, S.; Escaramís, G.; Rabionet, R.; Iraola, S.; Kagerbauer, B.; Espinosa-Parrilla, Y.; Ferrer, I.; Estivill, X.; Martí, E. MicroRNA profiling of Parkinson’s disease brains identifies early downregulation of miR-34b/c which modulate mitochondrial function. Hum. Mol. Genet. 2011, 20, 3067–3078. [Google Scholar] [CrossRef] [PubMed]
- Caggiu, E.; Paulus, K.; Mameli, G.; Arru, G.; Sechi, G.P.; Leonardo, A. Sechi, Differential expression of miRNA 155 and miRNA 146a in Parkinson’s disease patients. eNeurologicalSci 2018, 13, 1–4. [Google Scholar] [CrossRef] [PubMed]
- Hamzei Taj, S.; Kho, W.; Riou, A.; Wiedermann, D.; Hoehn, M. MiRNA-124 induces neuroprotection and functional improvement after focal cerebral ischemia. Biomaterials 2016, 91, 151–165. [Google Scholar] [CrossRef]
- Yang, Z.; Xu, J.; Zhu, R.; Liu, L. Down-regulation of miRNA-128 contributes to neuropathic pain following spinal cord injury via activation of P38. Med. Sci. Monit. 2017, 23, 405–411. [Google Scholar] [CrossRef] [PubMed]
- Lin, B.; Lu, L.; Wang, Y.; Zhang, Q.; Wang, Z.; Cheng, G.; Duan, X.; Zhang, F.; Xie, M.; Le, H.; et al. Nanomedicine Directs Neuronal Differentiation of Neural Stem Cells via Silencing Long Noncoding RNA for Stroke Therapy. Nano Lett. 2021, 21, 806–815. [Google Scholar] [CrossRef]
- Zhang, L.; Mao, L.; Wang, H. The Neuroprotection Effects of Exosome in Central Nervous System Injuries: A New Target for Therapeutic Intervention. Mol. Neurobiol. 2022, 59, 7152–7169. [Google Scholar] [CrossRef]
- Yuan, P.; Ding, L.; Chen, H.; Wang, Y.; Li, C.; Zhao, S.; Yang, X.; Ma, Y.; Zhu, J.; Qi, X.; et al. Neural Stem Cell-Derived Exosomes Regulate Neural Stem Cell Differentiation Through miR-9-Hes1 Axis. Front. Cell Dev. Biol. 2021, 9, 601600. [Google Scholar] [CrossRef] [PubMed]
- Fan, Y.; Chen, Z.; Zhang, M. Role of exosomes in the pathogenesis, diagnosis, and treatment of central nervous system diseases. J. Transl. Med. 2022, 20, 291. [Google Scholar] [CrossRef]
- Gao, P.; Li, X.; Du, X.; Liu, S.; Xu, Y. Diagnostic and Therapeutic Potential of Exosomes in Neurodegenerative Diseases. Front. Aging Neurosci. 2021, 13, 790863. [Google Scholar] [CrossRef] [PubMed]
- Wang, H.; Yang, Y.; Liu, J.; Qian, L. Direct cell reprogramming: Approaches, mechanisms and progress. Nat. Rev. Mol. Cell Biol. 2021, 22, 410–424. [Google Scholar] [CrossRef] [PubMed]
- Richard, J.P.; Zuryn, S.; Fischer, N.; Pavet, V.; Vaucamps, N.; Jarriault, S. Direct in vivo cellular reprogramming involves transition through discrete, non-pluripotent steps. Development 2011, 138, 1483–1492. [Google Scholar] [CrossRef] [PubMed]
- Wang, C.; Rabadan Ros, R.; Martinez-Redondo, P.; Ma, Z.; Shi, L.; Xue, Y.; Guillen-Guillen, I.; Huang, L.; Hishida, T.; Liao, H.-K.; et al. In vivo partial reprogramming of myofibers promotes muscle regeneration by remodeling the stem cell niche. Nat. Commun. 2021, 12, 3094. [Google Scholar] [CrossRef] [PubMed]
- Raredon, M.S.B.; Yang, J.; Kothapalli, N.; Lewis, W.; Kaminski, N.; Niklason, L.E.; Kluger, Y. Comprehensive visualization of cell–cell interactions in single-cell and spatial transcriptomics with NICHES. Bioinformatics 2023, 39, btac775. [Google Scholar] [CrossRef] [PubMed]
- Haas, S.; Baccin, C.; Al-Sabah, J.; Velten, L.; Lars, S.; Trumpp, A. Combined Single-Cell and Spatial Transcriptomics to Deconvolute the Hematopoietic Stem Cell Niche. Blood 2018, 132 (Suppl. S1), 876. [Google Scholar] [CrossRef]
- Al-Sabah, J.; Baccin, C.; Haas, S. Single-cell and spatial transcriptomics approaches of the bone marrow microenvironment. Curr. Opin. Oncol. 2020, 32, 146–153. [Google Scholar] [CrossRef]
- Hadland, B.; Varnum-Finney, B.; Dozono, S.; Dignum, T.; Nourigat-McKay, C.; Heck, A.M.; Ishida, T.; Jackson, D.L.; Itkin, T.; Butler, J.M.; et al. Engineering a niche supporting haematopoietic stem cell development using integrated single cell transcriptomics. bioRxiv 2021. [Google Scholar] [CrossRef]
- Carrano, A.; Juarez, J.J.; Incontri, D.; Ibarra, A.; Guerrero Cazares, H. Sex-Specific Differences in Glioblastoma. Cells 2021, 10, 1783. [Google Scholar] [CrossRef]
- O’Leary, T.P.; Askari, B.; Lee, B.H.; Darby, K.; Knudson, C.; Ash, A.M.; Seib, D.R.; Espinueva, D.F.; Snyder, J.S. Sex Differences in the Spatial Behavior Functions of Adult-Born Neurons in Rats. eNeuro 2022, 9, ENEURO.0054-22.2022. [Google Scholar] [CrossRef]
- Hillerer, K.M.; Neumann, I.D.; Couillard-Despres, S.; Aigner, L.; Slattery, D.A. Sex-dependent regulation of hippocampal neurogenesis under basal and chronic stress conditions in rats. Hippocampus 2013, 23, 476–487. [Google Scholar] [CrossRef] [PubMed]
- Yagi, S.; Splinter, J.E.; Tai, D.; Wong, S.; Wen, Y.; Galea, L.A. Sex Differences in Maturation and Attrition of Adult Neurogenesis in the Hippocampus. eNeuro 2020, 7, ENEURO.0468-19.2020. [Google Scholar] [CrossRef] [PubMed]
- Kim, H.T.; Zhang, M.-J.; Woolfrey, A.E.; Martin, A.S.; Chen, J.; Saber, W.; Perales, M.-A.; Armand, P.; Eapen, M. Donor and recipient sex in allogeneic stem cell transplantation: What really matters. Haematologica 2016, 101, 1260–1266. [Google Scholar] [CrossRef] [PubMed]
- Islam, P.; Tang, H.; Jin, H.; Cao, F.; Bohannon, L.M.; Ren, Y.; Chao, N.J.; Choi, T.; Gasparetto, C.; Horwitz, M.E.; et al. Female Sex Is Associated with Improved Long-Term Survival Following Allogeneic Hematopoietic Stem Cell Transplantation. Transpl. Cell Ther. 2021, 27, e784.e1–e784.e7. [Google Scholar] [CrossRef]
- Klein, S.; Flanagan, K. Sex differences in immune responses. Nat. Rev. Immunol. 2016, 16, 626–638. [Google Scholar] [CrossRef] [PubMed]
- Prins, G.S.; Calderon-Gierszal, E.L.; Hu, W.-Y. Stem Cells as Hormone Targets That Lead to Increased Cancer Susceptibility. Endocrinology 2015, 156, 3451–3457. [Google Scholar] [CrossRef] [PubMed]
- Okada, M.; Makino, A.; Nakajima, M.; Okuyama, S.; Furukawa, S.; Furukawa, Y. Estrogen stimulates proliferation and differentiation of neural stem/progenitor cells through different signal transduction pathways. Int. J. Mol. Sci. 2010, 11, 4114–4123. [Google Scholar] [CrossRef]
- Rae, J.M.; Lippman, M.E. The role of estrogen receptor signaling in suppressing the immune response to cancer. J. Clin. Invest. 2021, 131, e155476. [Google Scholar] [CrossRef]
- Harding, A.T.; Heaton, N.S. The Impact of Estrogens and Their Receptors on Immunity and Inflammation during Infection. Cancers 2022, 14, 909. [Google Scholar] [CrossRef]
- Zhong, X.; Sun, Y.; Lu, Y.; Xu, L. Immunomodulatory role of estrogen in ischemic stroke: Neuroinflammation and effect of sex. Front. Immunol. 2023, 14, 1164258. [Google Scholar] [CrossRef]
- Mackern-Oberti, J.P.; Jara, E.L.; Riedel, C.A.; Kalergis, A.M. Hormonal Modulation of Dendritic Cells Differentiation, Maturation and Function: Implications for the Initiation and Progress of Systemic Autoimmunity. Arch. Immunol. Ther. Exp. 2017, 65, 123–136. [Google Scholar] [CrossRef]
- Kodama, L.; Gan, L. Do Microglial Sex Differences Contribute to Sex Differences in Neurodegenerative Diseases? Trends Mol. Med. 2019, 25, 741–749. [Google Scholar] [CrossRef]
- Hong, W.; Wang, B.; Zhu, Y.; Wu, J.; Qiu, L.; Ling, S.; Zhou, Z.; Dai, Y.; Zhong, Z.; Zheng, Y. Female germline stem cells: Aging and anti-aging. J. Ovarian Res. 2022, 15, 79. [Google Scholar] [CrossRef]
- Barth, C.; de Lange, A.-M.G. Towards an understanding of women’s brain aging: The immunology of pregnancy and menopause. Front. Neuroendocrinol. 2020, 58, 100850. [Google Scholar] [CrossRef] [PubMed]
- Bustamante-Barrientos, F.A.; Méndez-Ruette, M.; Ortloff, A.; Luz-Crawford, P.; Rivera, F.J.; Figueroa, C.D.; Molina, L.; Bátiz, L.F. The Impact of Estrogen and Estrogen-Like Molecules in Neurogenesis and Neurodegeneration: Beneficial or Harmful? Front. Cell Neurosci. 2021, 15, 636176. [Google Scholar] [CrossRef] [PubMed]
- Cheng, Y.-J.; Lin, C.-H.; Lane, H.-Y. From Menopause to Neurodegeneration—Molecular Basis and Potential Therapy. Int. J. Mol. Sci. 2021, 22, 8654. [Google Scholar] [CrossRef] [PubMed]
- Kase, Y.; Shimazaki, T.; Okano, H. Current understanding of adult neurogenesis in the mammalian brain: How does adult neurogenesis decrease with age? Inflamm. Regener 2020, 40, 10. [Google Scholar] [CrossRef] [PubMed]
- Yun, M.H. Changes in Regenerative Capacity through Lifespan. Int. J. Mol. Sci. 2015, 16, 25392–25432. [Google Scholar] [CrossRef]
- Javanmehr, N.; Saleki, K.; Alijanizadeh, P.; Rezaei, N. Microglia dynamics in aging-related neurobehavioral and neuroinflammatory diseases. J. Neuroinflammation 2022, 19, 273. [Google Scholar] [CrossRef]
- Nicaise, A.M.; Willis, C.M.; Crocker, S.J.; Pluchino, S. Stem Cells of the Aging Brain. Front. Aging Neurosci. 2020, 12, 247. [Google Scholar] [CrossRef]
- Yuan, T.F.; Dong, Y.; Zhang, L.; Qi, J.; Yao, C.; Wang, Y.; Chai, R.; Liu, Y.; So, K.-F. Neuromodulation-Based Stem Cell Therapy in Brain Repair: Recent Advances and Future Perspectives. Neurosci. Bull. 2021, 37, 735–745. [Google Scholar] [CrossRef] [PubMed]
- Adugna, D.G.; Aragie, H.; Kibret, A.A.; Belay, D.G. Therapeutic Application of Stem Cells in the Repair of Traumatic Brain Injury. Stem Cells Cloning 2022, 15, 53–61. [Google Scholar] [CrossRef] [PubMed]
- Singh, M.; Pandey, P.K.; Bhasin, A.; Padma, M.V.; Mohanty, S. Application of Stem Cells in Stroke: A Multifactorial Approach. Front. Neurosci. 2020, 14, 473. [Google Scholar] [CrossRef]
- Genc, B.; Bozan, H.R.; Genc, S.; Genc, K. Stem Cell Therapy for Multiple Sclerosis. Adv. Exp. Med. Biol. 2019, 1084, 145–174. [Google Scholar] [CrossRef]
- Shang, Z.; Wang, M.; Zhang, B.; Wang, X.; Wanyan, P. Clinical translation of stem cell therapy for spinal cord injury still premature: Results from a single-arm meta-analysis based on 62 clinical trials. BMC Med. 2022, 20, 284. [Google Scholar] [CrossRef]
- Han, F.; Ebrahimi-Barough, S.; Abolghasemi, R.; Ai, J.; Liu, Y. Cell-Based Therapy for Spinal Muscular Atrophy. Adv. Exp. Med. Biol. 2020, 1266, 117–125. [Google Scholar] [CrossRef]
- Marsh, S.E.; Blurton-Jones, M. Neural stem cell therapy for neurodegenerative disorders: The role of neurotrophic support. Neurochem. Int. 2017, 106, 94–100. [Google Scholar] [CrossRef]
- Wang, G.; Wang, W. Advanced Cell Therapies for Glioblastoma. Front. Immunol. 2022, 13, 904133. [Google Scholar] [CrossRef]
- Wang, F.; Sun, Z.; Peng, D.; Gianchandani, S.; Le, W.; Boltze, J.; Li, S. Cell-therapy for Parkinson’s disease: A systematic review and meta-analysis. J. Transl. Med. 2023, 21, 601. [Google Scholar] [CrossRef] [PubMed]
- Qin, C.; Wang, K.; Zhang, L.; Bai, L. Stem cell therapy for Alzheimer’s disease: An overview of experimental models and reality. Animal Model. Exp. Med. 2022, 5, 15–26. [Google Scholar] [CrossRef] [PubMed]
- Najafi, S.; Najafi, P.; Kaffash Farkhad, N.; Hosseini Torshizi, G.; Assaran Darban, R.; Boroumand, A.R.; Sahab-Negah, S.; Khodadoust, M.A.; Tavakol-Afshari, J. Mesenchymal stem cell therapy in amyotrophic lateral sclerosis (ALS) patients: A comprehensive review of disease information and future perspectives. Iran. J. Basic. Med. Sci. 2023, 26, 872–881. [Google Scholar] [CrossRef]
- Wang, K.; Li, Y.; Ren, C.; Wang, Y.; He, W.; Jiang, Y. Extracellular Vesicles as Innovative Treatment Strategy for Amyotrophic Lateral Sclerosis. Front. Cell Dev. Biol. 2021, 11, 754630. [Google Scholar] [CrossRef] [PubMed]
- Mo, Z.; Zeng, Z.; Liu, Y.; Zeng, L.; Fang, J.; Ma, Y. Activation of Wnt/Beta-Catenin Signaling Pathway as a Promising Therapeutic Candidate for Cerebral Ischemia/Reperfusion Injury. Front. Pharmacol. 2022, 13, 914537. [Google Scholar] [CrossRef] [PubMed]
- Ojeda-Hernández, D.D.; Hernández-Sapiéns, M.A.; Reza-Zaldívar, E.E.; Canales-Aguirre, A.; Matías-Guiu, J.A.; Matías-Guiu, J.; Mateos-Díaz, J.C.; Gómez-Pinedo, U.; Sancho-Bielsa, F. Exosomes and Biomaterials: In Search of a New Therapeutic Strategy for Multiple Sclerosis. Life 2022, 12, 1417. [Google Scholar] [CrossRef] [PubMed]
- Krienke, C.; Kolb, L.; Diken, E.; Streuber, M.; Kirchhoff, S.; Bukur, T.; Akilli-Öztürk.; Kranz, L.M.; Berger, H.; Petschenka, J.; et al. A noninflammatory mRNA vaccine for treatment of experimental autoimmune encephalomyelitis. Science 2021, 371, 145–153. [Google Scholar] [CrossRef]
- Maric, D.M.; Vojvodic, D.; Maric, D.L.; Velikic, G.; Radomir, M.; Sokolovac, I.; Stefik, D.; Ivkovic, N.; Susnjevic, S.; Puletic, M.; et al. Cytokine Dynamics in Autism: Analysis of BMAC Therapy Outcomes. Int. J. Mol. Sci. 2023, 24, 15080. [Google Scholar] [CrossRef]
- Maroso, M. A quest into the human brain. Science 2023, 382, 166–167. [Google Scholar] [CrossRef]
- Møllgård, K.; Beinlich, F.R.M.; Kusk, P.; Miyakoshi, L.M.; Delle, C.; Plá, V.; Hauglund, N.L.; Esmail, T.; Rasmussen, M.K.; Gomolka, R.S.; et al. A mesothelium divides the subarachnoid space into functional compartments. Science 2023, 379, 84–88. [Google Scholar] [CrossRef]
- Kataoka, M.; Gyngell, C.; Savulescu, J.; Sawai, T. The Ethics of Human Brain Organoid Transplantation in Animals. Neuroethics 2023, 16, 27. [Google Scholar] [CrossRef]
- Li, S.; Lei, Z.; Sun, T. The role of microRNAs in neurodegenerative diseases: A review. Cell Biol. Toxicol. 2023, 39, 53–83. [Google Scholar] [CrossRef]
- Qian, X.; Song, H.; Ming, G.-L. Brain organoids: Advances, applications and challenges. Development 2019, 146, dev166074. [Google Scholar] [CrossRef]
- Kanton, S.; Boyle, M.J.; He, Z.; Santel, M.; Weigert, A.; Sanchís-Calleja, F.; Guijarro, P.; Sidow, L.; Fleck, J.S.; Han, D.; et al. Organoid single-cell genomic atlas uncovers human-specific features of brain development. Nature 2019, 574, 418–422. [Google Scholar] [CrossRef] [PubMed]
- García-Delgado, A.B.; Campos-Cuerva, R.; Rosell-Valle, C.; Martin-López, M.; Casado, C.; Ferrari, D.; Márquez-Rivas, J.; Sánchez-Pernaute, R.; Fernández-Muñoz, B. Brain Organoids to Evaluate Cellular Therapies. Animals 2022, 12, 3150. [Google Scholar] [CrossRef] [PubMed]
- Revah, O.; Gore, F.; Kelley, K.W.; Andersen, J.; Sakai, N.; Chen, X.; Li, M.-Y.; Birey, F.; Yang, X.; Saw, N.L.; et al. Maturation and circuit integration of transplanted human cortical organoids. Nature 2022, 610, 319–326. [Google Scholar] [CrossRef] [PubMed]
- Seiler, S.T.; Mantalas, G.L.; Selberg, J.; Cordero, S.; Torres-Montoya, S.; Baudin, P.V.; Ly, V.T.; Amend, F.; Tran, L.; Hoffman, R.N.; et al. Modular automated microfluidic cell culture platform reduces glycolytic stress in cerebral cortex organoids. Sci. Rep. 2022, 12, 20173. [Google Scholar] [CrossRef] [PubMed]
- de Hoyos-Vega, J.M.; Yu, X.; Gonzalez-Suarez, A.M.; Chen, S.; Mercado-Perez, A.; Krueger, E.; Hernandez, J.; Fedyshyn, Y.; Druliner, B.R.; Linden, D.R.; et al. Modeling gut neuro-epithelial connections in a novel microfluidic device. Microsyst. Nanoeng. 2023, 9, 144. [Google Scholar] [CrossRef]
- Angelopoulos, I.; Gakis, G.; Birmpas, K.; Kyrousi, C.; Habeos, E.E.; Kaplani, K.; Lygerou, Z.; Habeos, I.; Taraviras, S. Metabolic regulation of the neural stem cell fate: Unraveling new connections, establishing new concepts. Front. Neurosci. 2022, 16, 1009125. [Google Scholar] [CrossRef]
- Kalebic, N.; Gilardi, C.; Albert, M.; Namba, T.; Long, K.R.; Kostic, M.; Lange, B.; Huttner, W.B. Human-specific ARHGAP11B induces hallmarks of neocortical expansion in developing ferret neocortex. eLife 2018, 7, e41241. [Google Scholar] [CrossRef]
- Taelman, J.; Diaz, M.; Guiu, J. Human Intestinal Organoids: Promise and Challenge. Front. Cell Dev. Biol. 2022, 10, 854740. [Google Scholar] [CrossRef] [PubMed]
- Tsuruta, S.; Uchida, H.; Akutsu, H. Intestinal Organoids Generated from Human Pluripotent Stem Cells. JMA 2019, 3, 9–19. [Google Scholar] [CrossRef]
- Harrison, S.P.; Baumgarten, S.F.; Verma, R.; Lunov, O.; Dejneka, A.; Sullivan, G.J. Liver Organoids: Recent Developments, Limitations and Potential. Front. Med. 2021, 8, 574047. [Google Scholar] [CrossRef] [PubMed]
- Naderi-Meshkin, H.; Cornelius, V.A.; Eleftheriadou, M.; Potel, K.N.; Setyaningsih, W.A.W.; Margariti, A. Vascular organoids: Unveiling advantages, applications, challenges, and disease modelling strategies. Stem Cell Res. Ther. 2023, 14, 292. [Google Scholar] [CrossRef] [PubMed]
- Brown, J.A.; Pensabene, V.; Markov, D.A.; Allwardt, V.; Neely, M.D.; Shi, M.; Britt, C.M.; Hoilett, O.S.; Yang, Q.; Brewer, B.M.; et al. Recreating blood-brain barrier physiology and structure on chip: A novel neurovascular microfluidic bioreactor. Biomicrofluidics 2015, 9, 054124. [Google Scholar] [CrossRef] [PubMed]
- Choe, A.; Ha, S.K.; Choi, I.; Choi, N.; Sung, J.H. Microfluidic Gut-liver chip for reproducing the first pass metabolism. Biomed. Microdevices 2017, 19, 4. [Google Scholar] [CrossRef] [PubMed]
- Setty, Y. In-silico models of stem cell and developmental systems. Theor. Biol. Med. Model. 2014, 11, 1. [Google Scholar] [CrossRef] [PubMed]
- Tuladhar, A.; Moore, J.A.; Ismail, Z.; Forkert, N.D. Modeling Neurodegeneration in silico With Deep Learning. Front. Neuroinform. 2021, 15, 748370. [Google Scholar] [CrossRef]
- Martínez-García, M.; Hernández-Lemus, E. Data Integration Challenges for Machine Learning in Precision Medicine. Front. Med. 2022, 8, 784455. [Google Scholar] [CrossRef]
- Chander, B.S.; Chakravarthy, V.S. A Computational Model of Neuro-Glio-Vascular Loop Interactions. PLoS ONE 2012, 7, e48802. [Google Scholar] [CrossRef]
- Bozhko, D.V.; Galumov, G.K.; Polovian, A.I.; Kolchanova, S.M.; Myrov, V.O.; Stelmakh, V.A.; Schiöth, H.B. BCNNM: A Framework for in silico Neural Tissue Development Modeling. Front. Comput. Neurosci. 2021, 14, 588224. [Google Scholar] [CrossRef]
- Ruiz-Clavijo, L.; Martín-Suárez, S. The differential response to neuronal hyperexcitation and neuroinflammation of the hippocampal neurogenic niche. Front. Neurosci. 2023, 17, 1186256. [Google Scholar] [CrossRef]
- Xuan, Y.; Li, Z.; Ghatak, S.; Sen, C.K. Tissue Nanotransfection in Regenerative Medicine. In MicroRNA in Regenerative Medicine, 2nd ed.; Elsevier: Amsterdam, The Netherlands, 2023; pp. 1051–1074. [Google Scholar]
- Huang, H.; Sanberg, P.R.; Moviglia, G.A.; Sharma, A.; Chen, L.; Chen, D. Clinical results of neurorestorative cell therapies and therapeutic indications according to cellular bio-proprieties. Regen. Ther. 2023, 23, 52–59. [Google Scholar] [CrossRef] [PubMed]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Approach | Conventional Strategies | Niche-Targeted Strategies |
|---|---|---|
| Goal | Manage symptoms and slow disease progression by targeting downstream effects | Stimulate neurogenesis and neural regeneration by targeting endogenous stem cells and niche factors [1] |
| Methods | Pharmacological agents, vitamins, lifestyle modifications | Growth factors, cytokines, gene and cell therapy, small molecules, biomaterials [1] |
| Targets | Neurotransmitter deficits, protein aggregation, inflammation, oxidative stress | Endogenous neural stem cells, support cells (astrocytes, endothelial cells), signaling factors (Wnt, BMP, Notch), vasculature, extracellular matrix (ECM) [9] |
| Mechanisms | Acetylcholinesterase inhibition (donepezil), NMDA receptor antagonism (memantine), dopamine replacement (levodopa), monoamine oxidase inhibition (selegiline), reactive oxygen species scavenging (vitamin E) | Niche pathway modulation, ECM modifications, anti-inflammatories, signaling molecule administration [9] |
| Outcomes | Modest symptom improvement, some modification of disease progression | Increased neurogenesis, integration of new neurons, possible reversal of cognitive/motor decline [9] |
| Limitations | Do not restore lost neurons or neural connections, benefits not sustained | Integrating new neurons, ensuring survival, unclear long-term benefits [10] |
| Function | Description | Examples |
|---|---|---|
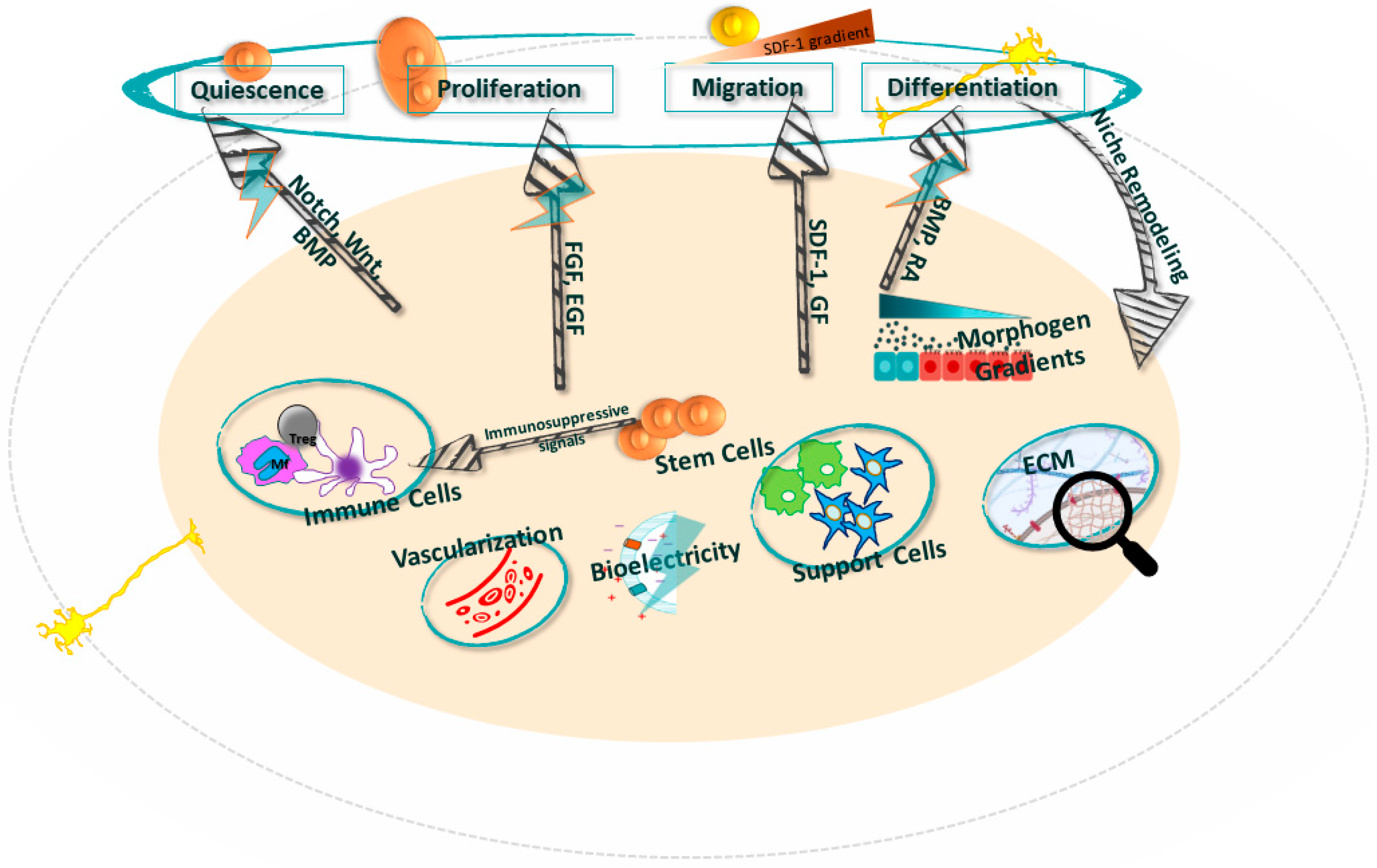
| Quiescence | The niche maintains stem cells in a nondividing quiescent state through factors like hypoxia, cell–cell contact, adhesive interactions, and signaling molecules. This preserves stemness and prevents premature exhaustion of the stem cell pool [11,12]. |
|
| Activation | The niche can rapidly activate quiescent stem cells in response to injury/stress signals, often mediated by inflammatory cytokines, growth factors, chemokines, ECM remodeling, and release of retention factors that trigger proliferation and mobilization [13,14]. |
|
| Proliferation | The niche provides spatial and biochemical cues to ensure stem cells undergo symmetric, self-renewing cell divisions to maintain or expand the stem cell pool. This is regulated through signaling pathways like Wnt, Notch, Hedgehog, BMP [15,16,17,18,19,20,21]. |
|
| Differentiation | The niche provides biochemical and biophysical cues to direct stem cell differentiation into specific lineages, often via morphogen gradients, ECM properties, epigenetic changes, and bioelectric signals [22]. |
|
| Homing | The niche produces chemotactic factors like SDF-1 to guide migration and homing of circulating stem cells back to their niche residence after injury or transplantation [23]. |
|
| Engraftment | The niche provides adhesion molecules, ECM components, and growth factors to enable proper anchoring, survival, and integration of transplanted stem cells [12]. |
|
| Immunomodulation | The niche shields stem cells from immune attack and modulates inflammatory signals through physical barriers, immunosuppressive molecules, and lack of MHC 1 on stem cells to foster a conducive environment [12]. |
|
| Apoptosis | The niche tightly regulates stem cell numbers by inducing programmed cell death via death receptor signaling, cytokine withdrawal, or loss of survival signals to clear excess or dysfunctional stem cells [12]. |
|
| Condition | Impact Factor | Specific Impact on Disease | Implications for Stem Cell Niche Restoration and Regeneration |
|---|---|---|---|
| ALS [182] | Inflammation, loss of neurons and reactive glia | Alters signaling, reduces stemness. | Anti-inflammatory therapies, cell replacement strategies. |
| Traumatic Brain Injury [215] | Damaged neurons and glia | Impairs stemness and signaling. | Anti-inflammatory therapies, NSC transplantation, modulation of post-injury niche factors. |
| Stroke [216] | Disrupted niche vasculature, BBB integrity, epigenetic alterations | Reduces oxygen/nutrient supply, alters gene regulation. | Vascularization, transplantation of neural progenitors, epigenetic therapies. |
| MS [217] | Demyelination, inflammation | Disrupts signaling, alters cell communication. | GF, remyelinating agents, anti-inflammatory treatments. |
| Spinal Cord Injury [218] | Loss of niche factors and signals below the injury site reduces neurogenesis | Disconnects signaling, deprives of nutrients. | Delivering niche factors, molecules, and stem cell transplants may encourage regrowth and remyelination. |
| SMA [219] | Loss of neurons and glia, aging | Reduces support cells and signals, affects stem cell function with age. | NSC transplants, gene therapy, rejuvenation strategies. |
| LBD [220] | Toxic molecules—alpha-synuclein aggregation | Interferes with signaling pathways, disrupts neurogenic niches impairing dopamine neuron generation. | Molecule-clearing treatments with cell therapy to restore niches. |
| GBM [221] | Tumor growth, epigenetic alterations | Distorts niches, redirects stem cells to tumor growth, alters gene regulation. | Target tumor microenvironments to restrict tumorigenic niche signals, epigenetic modification therapies. |
| Parkinson’s [222] | Loss of neurons and glia, aging | Removes support cells and signals, aging affects stemness. | Cell replacement strategies, anti-aging therapies, GDNF, or other regenerative factors may encourage neurogenesis. |
| Alzheimer’s [223] | Toxic molecules, aging | Distorts niche signaling and neurogenesis. | Clearing toxic molecules, anti-inflammatory, rejuvenation and anti-aging strategies. |
| Disease | Therapy | Relevant Outcomes | Limitations | Connection with Stem Cell Niche | Speculations on Interaction/Modulation of Niches |
|---|---|---|---|---|---|
| ALS [224,225] | Stem Cell Administration (MSC) | Slowed disease progression | Immune response to transplanted cells, limited long-term efficacy, variability in humans | Enhanced niche support | Transplanted stem cells may secrete factors that enhance the local niche environment, promoting neuronal survival and reducing neuroinflammation. This interaction can provide neuroprotection and support for damaged neurons. |
| Exosomes | Neuroprotection | Purification | Modulation of niche signaling | Exosomes released by stem cells may contain bioactive molecules that can modulate niche signaling pathways, promoting neuroprotection and potentially reducing neuroinflammation. | |
| Traumatic Brain Injury [215] | Stem Cell Administration | Improved cognitive function | Cell survival and integration | Niche-derived factors | Transplanted stem cells can interact with the local niche, receiving signals that promote their survival, differentiation, and integration into the injured tissue. |
| mRNA | Enhanced neurorepair | Limited delivery to target site | Niche regulation of mRNA signaling | Stem-cell-derived mRNA therapies may be designed to respond to niche-specific signals, ensuring that they are activated in the appropriate microenvironment for neurorepair. | |
| MSC Exosomes | Improved neurological function, reduced inflammation in models | Nontargeted delivery, unclear mechanisms | Niche-derived exosomal factors | Exosomal bioactive molecules which can promote neuronal regeneration | |
| Stroke [216,226] | Stem Cell Administration | Functional recovery | Immune rejection | Niche-mediated immune modulation | Transplanted stem cells can modulate the local immune response within the niche, reducing inflammation and promoting tissue repair. |
| Modulating Wnt Signaling Pathways | Reduced neuroinflammation, neuroprotection, angiogenesis in ischemia models | Risk of tumorigenesis | Niche-related signaling pathways | Manipulating stem cell signaling pathways can indirectly affect the niche by altering the production of niche-related factors and reducing neuroinflammation. | |
| Multiple Sclerosis [217,227,228] | Stem Cell Administration | Disease stabilization | Immune-related complications | Niche–immune interactions | Transplanted stem cells can interact with the immune cells in the niche, potentially modulating autoimmune responses and stabilizing the disease state. |
| MSC-derived Exosomes | Neuroprotection | Standardization of exosome isolation, limited data | Niche-derived exosomal factors | Exosomes from stem cells may carry factors that can interact with and modulate the immune and glial cells within the niche, providing neuroprotection. | |
| mRNA | Enhanced axonal regeneration | Delivery challenges | Niche modulation of mRNA signaling | mRNA therapies can be designed to respond to niche-specific cues, facilitating axonal regeneration and remyelination. | |
| Spinal Cord Injury [218] | Stem Cell Administration | Motor function improvement | Limited axonal regeneration | Niche support for axonal growth | Stem cells may interact with the niche to promote axonal regeneration by secreting factors that create a conducive microenvironment for axon growth and remyelination. |
| SMA [219] | Stem Cell Administration | Prolonged survival | Limited availability of suitable donors | Niche-mediated survival signals | Transplanted stem cells can receive signals from the niche that promote their survival and function, potentially prolonging patient survival. |
| LBD [220] | Stem Cell Administration | Improved cognitive function | Immune-related complications | Niche support for neural function | Transplanted stem cells may interact with the neural niche to enhance cognitive function through neuroprotection and synaptic support. |
| GBM [221] | Stem Cell Administration | Tumor suppression | Risk of promoting tumor growth | Niche influence on tumor microenvironment | Stem cells may interact with the tumor microenvironment, modulating it to suppress tumor growth and invasion. |
| Parkinson’s [222] | Stem Cell Administration | Improved motor function | Variable clinical responses | Niche support for dopamine production | Transplanted stem cells may enhance dopamine production within the niche, improving motor function in Parkinson’s disease patients. |
| Alzheimer’s [223] | Stem Cell Administration | Cognitive improvement | Limited engraftment in the brain | Niche support for cognitive function | Transplanted stem cells may enhance cognitive function by modulating the local niche environment and supporting neuronal health. |
| Exosomes | Neuroprotection | Standardization of exosome isolation | Niche-derived exosomal factors | Exosomes released by stem cells may contain factors that interact with the niche to provide neuroprotection and support cognitive function. |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Velikic, G.; Maric, D.M.; Maric, D.L.; Supic, G.; Puletic, M.; Dulic, O.; Vojvodic, D. Harnessing the Stem Cell Niche in Regenerative Medicine: Innovative Avenue to Combat Neurodegenerative Diseases. Int. J. Mol. Sci. 2024, 25, 993. https://doi.org/10.3390/ijms25020993
Velikic G, Maric DM, Maric DL, Supic G, Puletic M, Dulic O, Vojvodic D. Harnessing the Stem Cell Niche in Regenerative Medicine: Innovative Avenue to Combat Neurodegenerative Diseases. International Journal of Molecular Sciences. 2024; 25(2):993. https://doi.org/10.3390/ijms25020993
Chicago/Turabian StyleVelikic, Gordana, Dusan M. Maric, Dusica L. Maric, Gordana Supic, Miljan Puletic, Oliver Dulic, and Danilo Vojvodic. 2024. "Harnessing the Stem Cell Niche in Regenerative Medicine: Innovative Avenue to Combat Neurodegenerative Diseases" International Journal of Molecular Sciences 25, no. 2: 993. https://doi.org/10.3390/ijms25020993