
α-Pinene, a Main Component of Pinus Essential Oils, Enhances the Expression of Insulin-Sensitive Glucose Transporter Type 4 in Murine Skeletal Muscle Cells
 ,
,  and
and 
Abstract
:1. Introduction
2. Results
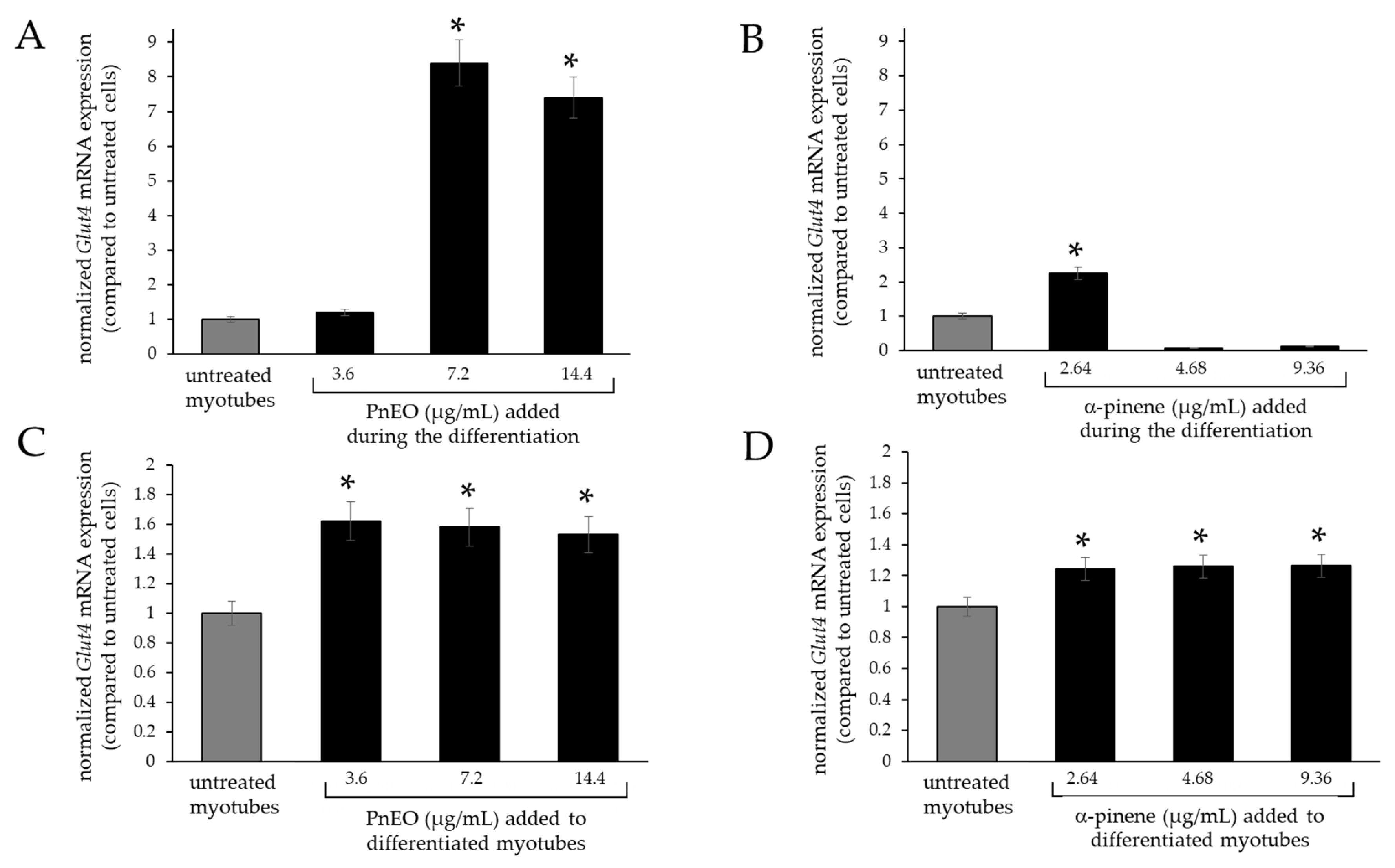
2.1. Analysis of the Effect of Pinus EOs on Glut4 Gene Transcription in Myoblasts
2.2. Effect of the PnEO on Cell Surface Exposure of the GLUT4 Transporter
2.3. Characterization of the Chemical Composition of the PnEO
2.4. Effect of Pinenes on GLUT4 Expression
2.5. Effects of the PnEO and α-Pinene on Glut4 Expression in Myotubes
3. Discussion
4. Materials and Methods
4.1. Materials
4.2. Cell Cultures
4.3. Cell Viability
4.4. RNA Extraction
4.5. Quantitative Real-Time Polymerase Chain Reaction (RT-qPCR)
4.6. GC–MS Analysis
4.7. Hexokinase Activity
4.8. Flow Cytometry
4.9. Statistical Analysis
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Nogueira, C.; Souto, S.B.; Vinha, E.; Braga, D.C.; Carvalho, D. Oral glucose lowering drugs in type 2 diabetic patients with chronic kidney disease. Hormones 2013, 12, 483–494. [Google Scholar] [CrossRef] [PubMed]
- Thorens, B. Glucose transporters in the regulation of intestinal, renal, and liver glucose fluxes. Am. J. Physiol. 1996, 270, G541–G553. [Google Scholar] [CrossRef] [PubMed]
- Thorens, B.; Mueckler, M. Glucose transporters in the 21st Century. Am. J. Physiol. Endocrinol. Metab. 2010, 298, E141–E145. [Google Scholar] [CrossRef] [PubMed]
- Chang, Y.C.; Chan, M.H.; Yang, Y.F.; Li, C.H.; Hsiao, M. Glucose transporter 4: Insulin response mastermind, glycolysis catalyst and treatment direction for cancer progression. Cancer Lett. 2023, 563, 216179. [Google Scholar] [CrossRef] [PubMed]
- Govers, R. Cellular regulation of glucose uptake by glucose transporter GLUT4. Adv. Clin. Chem. 2014, 66, 173–240. [Google Scholar]
- Saltiel, A.R. Insulin signaling in health and disease. J. Clin. Investig. 2021, 131, e142241. [Google Scholar] [CrossRef] [PubMed]
- Richter, E.A.; Hargreaves, M. Exercise, GLUT4, and skeletal muscle glucose uptake. Physiol. Rev. 2013, 93, 993–1017. [Google Scholar] [CrossRef]
- Moraes, P.A.; Yonamine, C.Y.; Pinto Junior, D.C.; Esteves, J.V.; Machado, U.F.; Mori, R.C. Insulin acutely triggers transcription of Slc2a4 gene: Participation of the AT-rich, E-box and NFKB-binding sites. Life Sci. 2014, 114, 36–44. [Google Scholar] [CrossRef]
- Alam, F.; Shafique, Z.; Amjad, S.T.; Bin Asad, M.H.H. Enzymes inhibitors from natural sources with antidiabetic activity: A review. Phytother. Res. 2019, 33, 41–54. [Google Scholar] [CrossRef]
- Carvalho, F.; Lahlou, R.A.; Pires, P.; Salgado, M.; Silva, L.R. Natural Functional Beverages as an Approach to Manage Diabetes. Int. J. Mol. Sci. 2023, 24, 16977. [Google Scholar] [CrossRef]
- Sharma, A.; Chawla, R.; Kaur, J.; Madaan, R. An Overview of Phytotherapy Used in the Management of Type II Diabetes. Curr. Diabetes Rev. 2022, 18, e170621194148. [Google Scholar] [CrossRef] [PubMed]
- Herman, A.; Herman, A.P. Essential oils and their constituents as skin penetration enhancer for transdermal drug delivery: A review. J. Pharm. Pharmacol. 2015, 67, 473–485. [Google Scholar] [CrossRef] [PubMed]
- Sousa, L.D.R.; Viana, N.R.; Coêlho, A.G.; Barbosa, C.O.; Barros, D.S.L.; Martins, M.D.C.C.E.; Ramos, R.M.; Arcanjo, D.D.R. Use of Monoterpenes as Potential Therapeutics in Diabetes Mellitus: A Prospective Review. Adv. Pharmacol. Pharm. Sci. 2023, 2023, 1512974. [Google Scholar] [CrossRef] [PubMed]
- Hoca, M.; Becer, E.; Vatansever, H.S. Carvacrol is potential molecule for diabetes treatment. Arch. Physiol. Biochem. 2023. online ahead of print. [Google Scholar] [CrossRef]
- Bungau, S.G.; Vesa, C.M.; Bustea, C.; Purza, A.L.; Tit, D.M.; Brisc, M.C.; Radu, A.F. Antioxidant and Hypoglycemic Potential of Essential Oils in Diabetes Mellitus and Its Complications. Int. J. Mol. Sci. 2023, 24, 16501. [Google Scholar] [CrossRef] [PubMed]
- Turner, M.C.; Player, D.J.; Martin, N.R.W.; Akam, E.C.; Lewis, M.P. The effect of chronic high insulin exposure upon metabolic and myogenic markers in C2C12 skeletal muscle cells and myotubes. J. Cell. Biochem. 2018, 119, 5686–5695. [Google Scholar] [CrossRef] [PubMed]
- Bakkali, F.; Averbeck, S.; Averbeck, D.; Idaomar, M. Biological effects of essential oils—A review. Food Chem. Toxicol. 2008, 46, 446–475. [Google Scholar] [CrossRef]
- Ritov, V.B.; Kelley, D.E. Hexokinase isozyme distribution in human skeletal muscle. Diabetes 2001, 50, 1253–1262. [Google Scholar] [CrossRef]
- Williams, A.C.; Barry, B.W. Terpenes and the lipid-protein-partitioning theory of skin penetration enhancement. Pharm. Res. 1991, 8, 17–24. [Google Scholar] [CrossRef]
- Zhou, J.Y.; Tang, F.D.; Mao, G.G.; Bian, R.L. Effect of alpha-pinene on nuclear translocation of NF-kappa B in THP-1 cells. Acta Pharmacol. Sin. 2004, 25, 480–484. [Google Scholar]
- Rahmani, H.; Moloudi, M.R.; Hashemi, P.; Hassanzadeh, K.; Izadpanah, E. Alpha-Pinene Alleviates Motor Activity in Animal Model of Huntington’s Disease via Enhancing Antioxidant Capacity. Neurochem. Res. 2023, 48, 1775–1782. [Google Scholar] [CrossRef] [PubMed]
- Darenskaya, M.A.; Kolesnikova, L.I.; Kolesnikov, S.I. Oxidative Stress: Pathogenetic Role in Diabetes Mellitus and Its Complications and Therapeutic Approaches to Correction. Bull. Exp. Biol. Med. 2021, 171, 179–189. [Google Scholar] [CrossRef] [PubMed]
- Yaffe, D.; Saxel, O. Serial passaging and differentiation of myogenic cells isolated from dystrophic mouse muscle. Nature 1977, 270, 725–727. [Google Scholar] [CrossRef] [PubMed]
- Murakami, M.; Ohkuma, M.; Nakamura, M. Molecular mechanism of transforming growth factor-beta-mediated inhibition of growth arrest and differentiation in a myoblast cell line. Dev. Growth Differ. 2008, 50, 121–130. [Google Scholar] [CrossRef]
- Feriotto, G.; Marchetti, N.; Costa, V.; Torricelli, P.; Beninati, S.; Tagliati, F.; Mischiati, C. Selected terpenes from leaves of Ocimum basilicum L. induce hemoglobin accumulation in human K562 cells. Fitoterapia 2018, 127, 173–178. [Google Scholar] [CrossRef]
- Feriotto, G.; Marchetti, N.; Costa, V.; Beninati, S.; Tagliati, F.; Mischiati, C. Chemical Composition of Essential Oils from Thymus vulgaris, Cymbopogon citratus, and Rosmarinus officinalis, and Their Effects on the HIV-1 Tat Protein Function. Chem. Biodivers. 2018, 15, e1700436. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Pinus radiata | Pinus nigra | ||||||
|---|---|---|---|---|---|---|---|
| Peak | Compound | Area (%) | SD (%) | Peak | Compound | Area (%) | SD (%) |
| 1 | α-phellandrene | 0.126 | 0.005 | 1 | santene | 0.164 | 0.006 |
| 2 | α-pinene | 23.416 | 0.890 | 2 | tricyclene | 0.622 | 0.024 |
| 3 | camphene | 0.790 | 0.030 | 3 | α-pinene | 65.308 | 2.482 |
| 4 | β-pinene | 42.635 | 1.620 | 4 | camphene | 5.981 | 0.227 |
| 5 | β-myrcene | 1.900 | 0.072 | 5 | β-pinene | 8.270 | 0.314 |
| 6 | 3-thujene | 0.158 | 0.006 | 6 | limonene | 4.185 | 0.159 |
| 7 | 3-carene | 8.319 | 0.316 | 7 | α-pinene epoxide | 0.949 | 0.036 |
| 8 | limonene | 11.663 | 0.443 | 8 | cis-verbenol | 3.223 | 0.122 |
| 9 | β-trans-ocimene | 0.925 | 0.035 | 9 | α-campholenal | 0.425 | 0.016 |
| 10 | τ-terpinen | 0.526 | 0.020 | 10 | pinocarveol | 0.935 | 0.036 |
| 11 | terpinolen | 2.133 | 0.081 | 11 | cis-verbenol | 2.558 | 0.097 |
| 12 | linalool | 0.358 | 0.014 | 12 | α-terpineol | 0.883 | 0.034 |
| 13 | fenchol | 0.254 | 0.010 | 13 | myrtenol | 0.421 | 0.016 |
| 14 | pinocarveol | 0.333 | 0.013 | 14 | l-verbenone | 0.728 | 0.028 |
| 15 | L-isopulegol | 0.218 | 0.008 | 15 | cis-carveol | 0.502 | 0.019 |
| 16 | 1-terpinen-4-ol | 1.526 | 0.058 | 16 | (-)-bornyl acetate | 3.863 | 0.147 |
| 17 | α-terpineol | 2.893 | 0.110 | 17 | τ-cadinene | 0.638 | 0.024 |
| 18 | (R)-(+)-β-citronellol | 0.988 | 0.038 | 18 | caryophyllene oxide | 0.352 | 0.013 |
| 19 | caryophyllene | 0.191 | 0.007 | ||||
| 20 | cadinene | 0.465 | 0.018 | ||||
| 21 | spathulenol | 0.182 | 0.007 | ||||
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Feriotto, G.; Tagliati, F.; Costa, V.; Monesi, M.; Tabolacci, C.; Beninati, S.; Mischiati, C. α-Pinene, a Main Component of Pinus Essential Oils, Enhances the Expression of Insulin-Sensitive Glucose Transporter Type 4 in Murine Skeletal Muscle Cells. Int. J. Mol. Sci. 2024, 25, 1252. https://doi.org/10.3390/ijms25021252
Feriotto G, Tagliati F, Costa V, Monesi M, Tabolacci C, Beninati S, Mischiati C. α-Pinene, a Main Component of Pinus Essential Oils, Enhances the Expression of Insulin-Sensitive Glucose Transporter Type 4 in Murine Skeletal Muscle Cells. International Journal of Molecular Sciences. 2024; 25(2):1252. https://doi.org/10.3390/ijms25021252
Chicago/Turabian StyleFeriotto, Giordana, Federico Tagliati, Valentina Costa, Marcello Monesi, Claudio Tabolacci, Simone Beninati, and Carlo Mischiati. 2024. "α-Pinene, a Main Component of Pinus Essential Oils, Enhances the Expression of Insulin-Sensitive Glucose Transporter Type 4 in Murine Skeletal Muscle Cells" International Journal of Molecular Sciences 25, no. 2: 1252. https://doi.org/10.3390/ijms25021252