
The Identification, Characterization, and Functional Analysis of the Sugar Transporter Gene Family of the Rice False Smut Pathogen, Villosiclava virens

Abstract
:1. Introduction
2. Results
2.1. Identification of the STP Family in V. virens
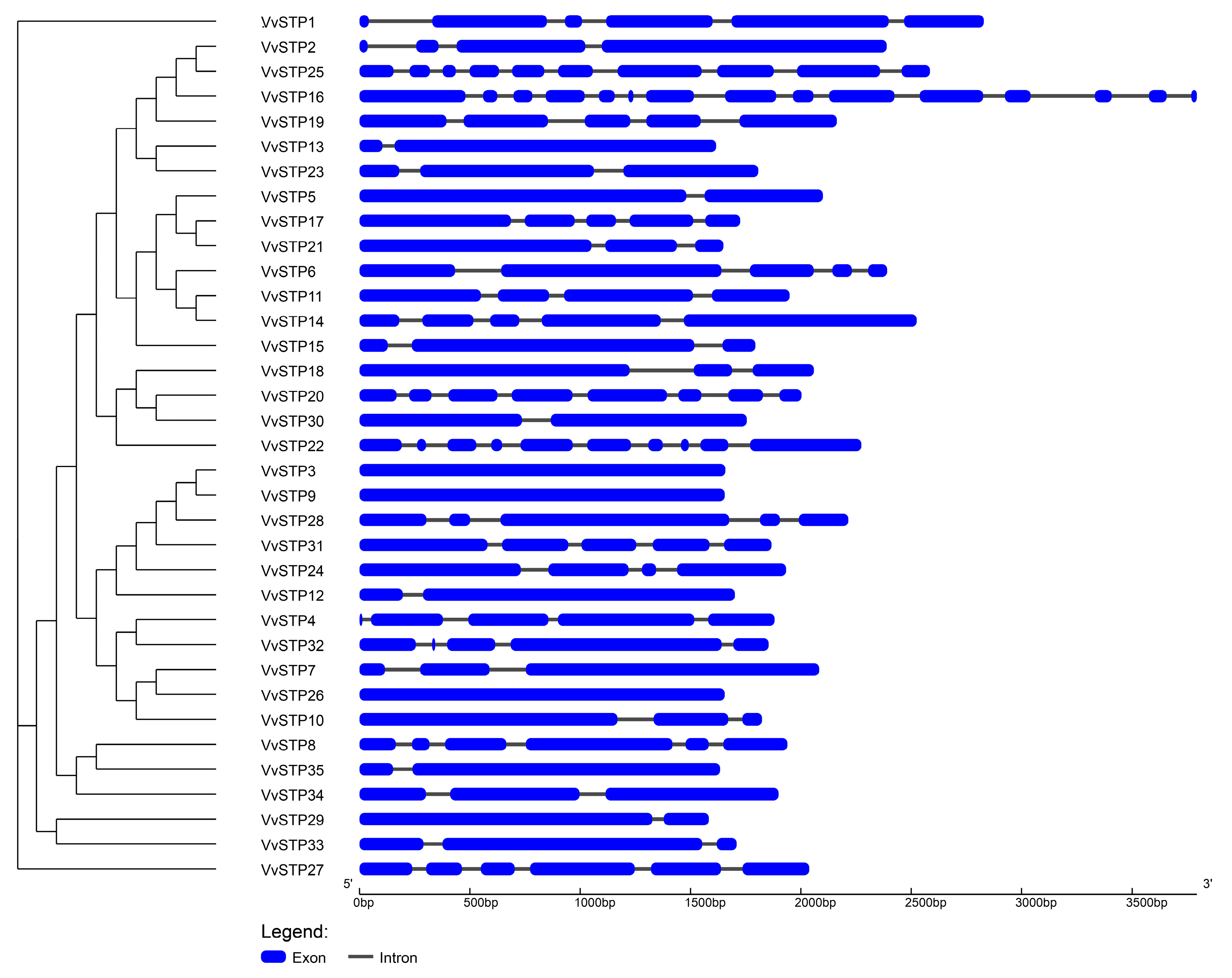
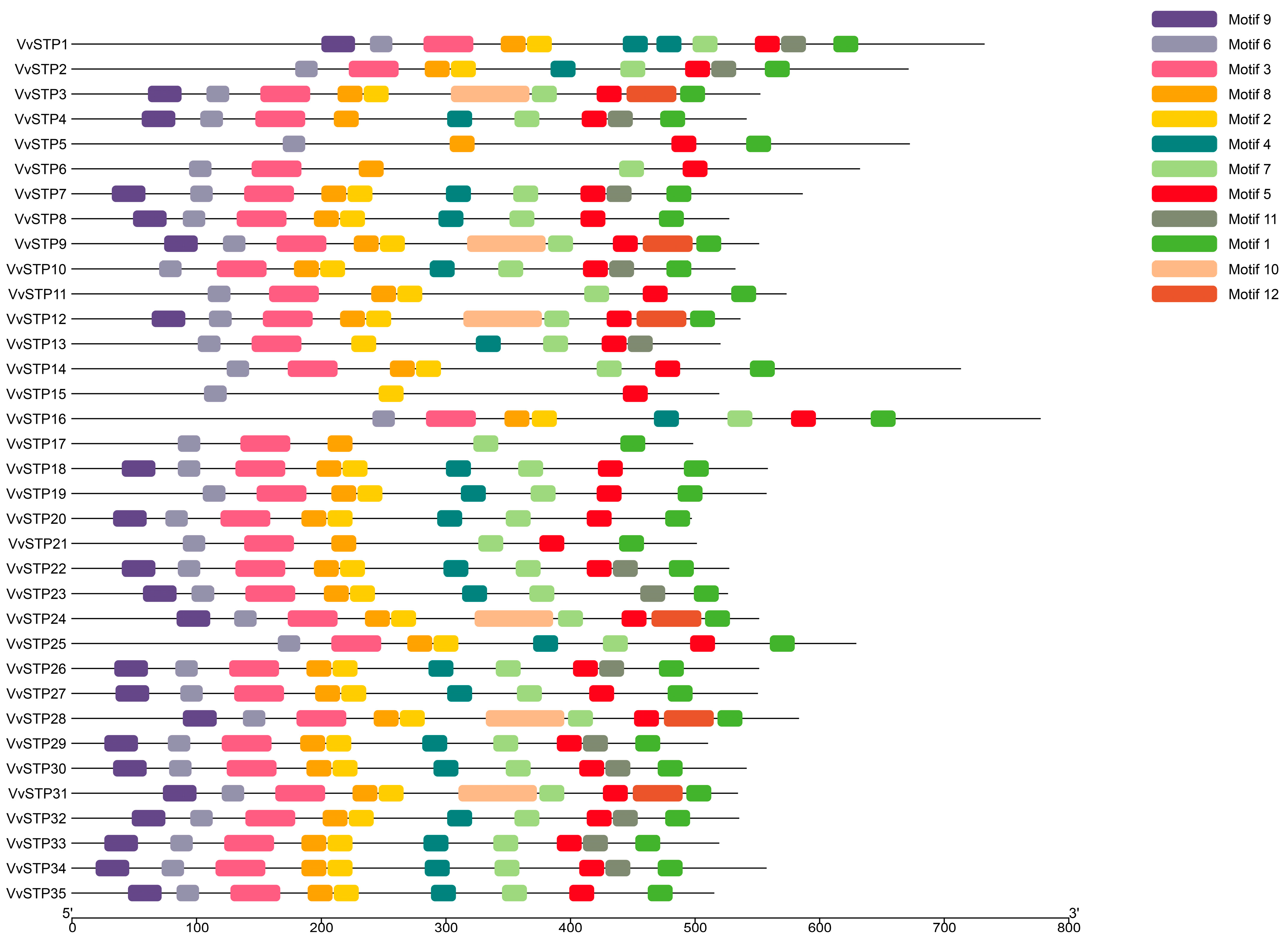
2.2. Phylogenetic Relationship, Gene Structure, Conserved Motifs of STP Genes of V. virens
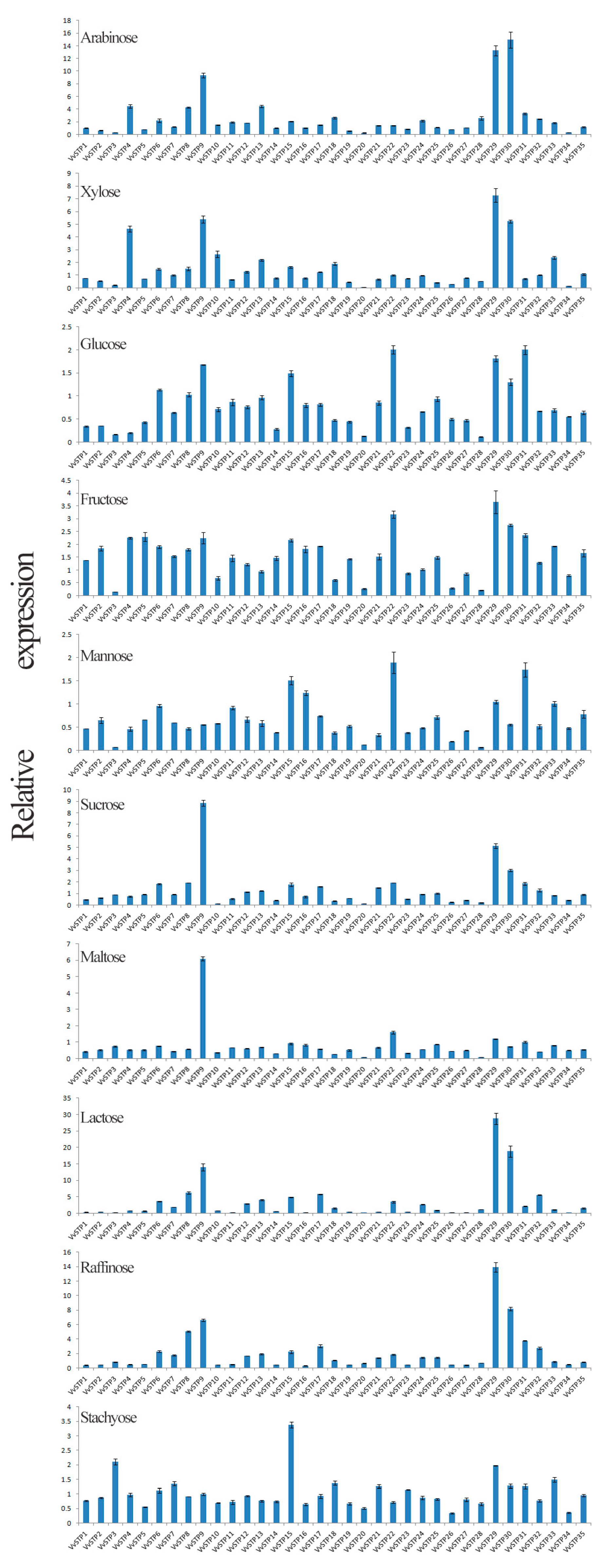
2.3. Expression Pattern of VvSTPs on Different Sole Carbon Sources
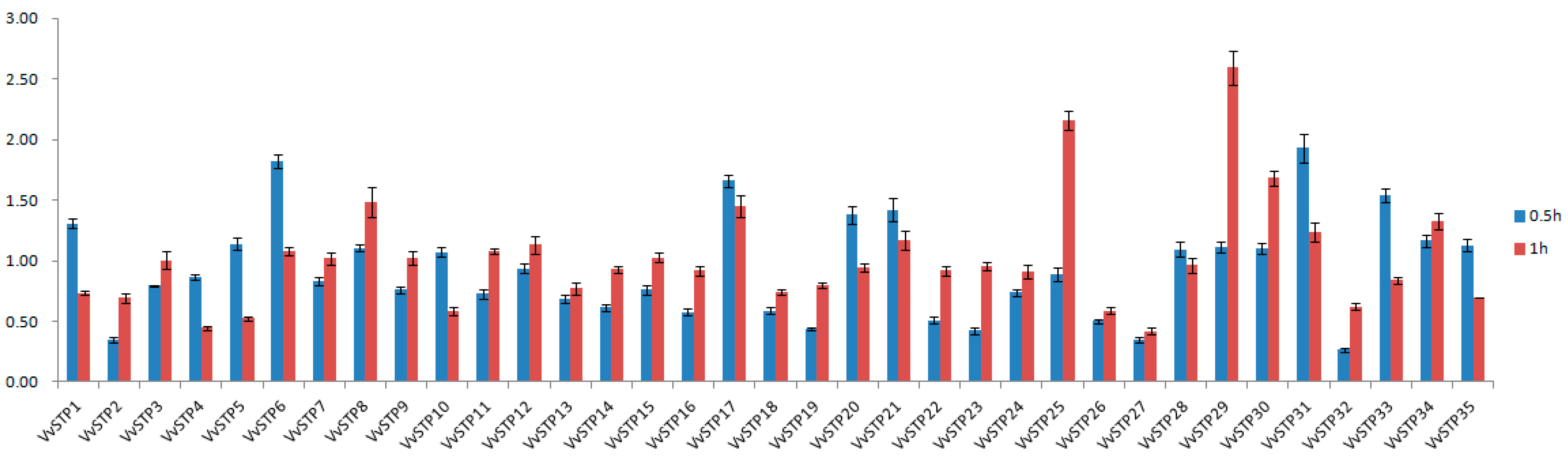
2.4. Expression Response of VvSTPs to H2O2 Treatment
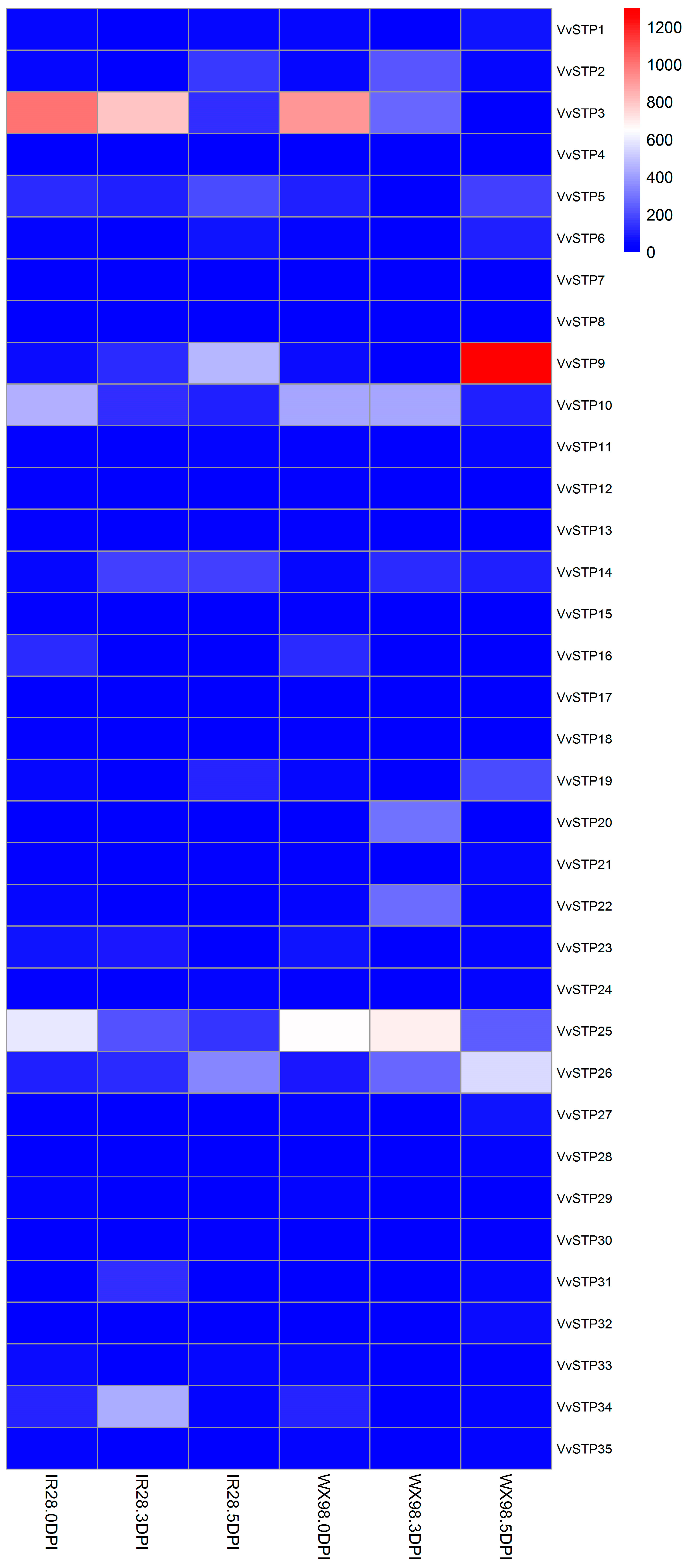
2.5. Expression Pattern of VvSTPs after Inoculation on Resistant and Susceptible Varieties
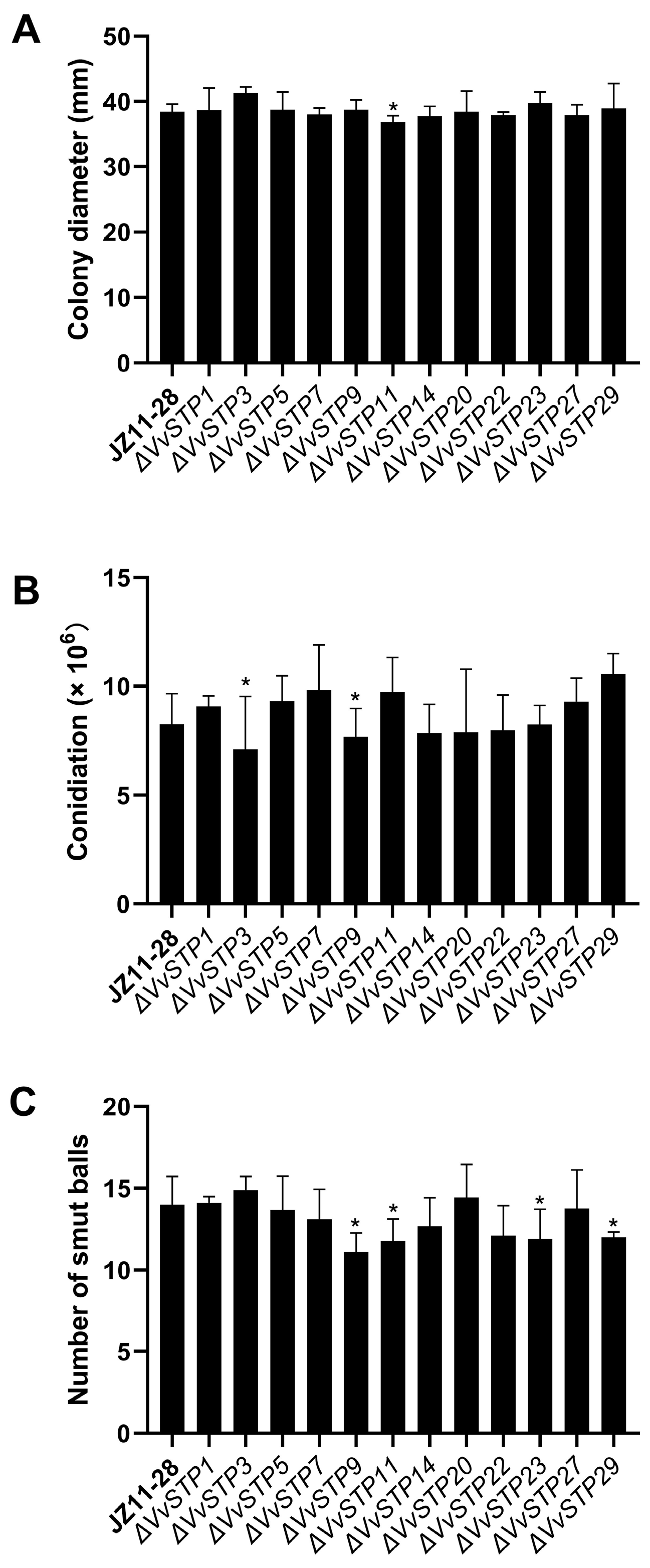
2.6. Phenotype of V. virens Conferred by Deletion of VvSTP Genes
3. Discussion
4. Material and Methods
4.1. Identification and Sequence Analysis of Sugar Transporter Genes of V. virens
4.2. Gene Structure and Conserved Motifs
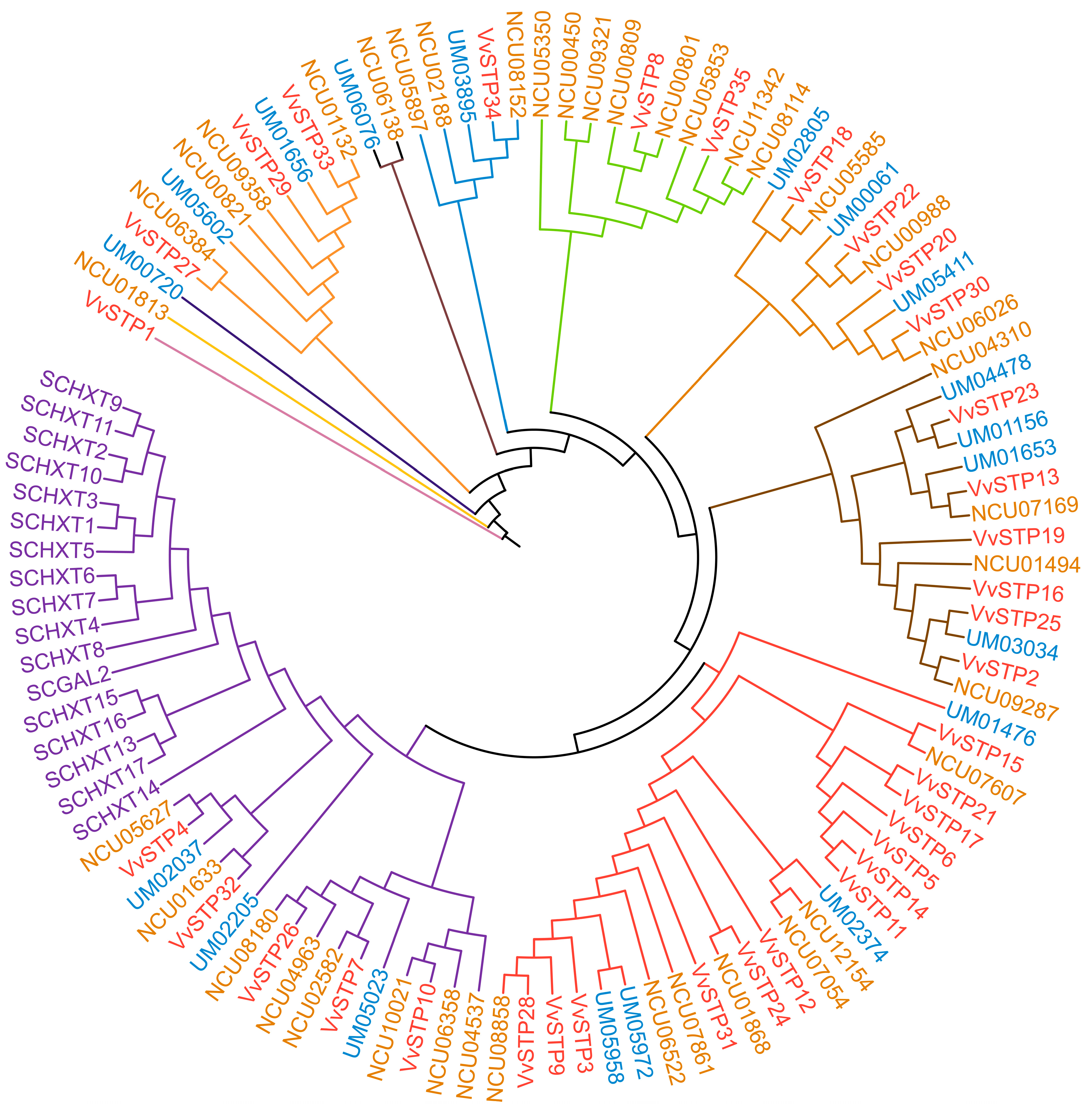
4.3. Gene Duplication Analysis and Phylogenetic Analysis of VvSTPs
4.4. Strain and Variety, Inoculation Experiments, and RNA-Seq
4.5. Quantitative Real-Time PCR Analysis
4.6. Generation of VvSTP Gene Deletion Mutants and Phenotype Assays
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Salvi, P.; Agarrwal, R.; Kajal; Gandass, N.; Manna, M.; Kaur, H.; Deshmukh, R. Sugar transporters and their molecular tradeoffs during abiotic stress responses in plants. Physiol. Plant. 2022, 174, e13652. [Google Scholar] [CrossRef]
- Williams, L.E.; Lemoine, R.; Sauer, N. Sugar transporters in higher plants—A diversity of roles and complex regulation. Trends Plant Sci. 2000, 5, 283–290. [Google Scholar] [CrossRef] [PubMed]
- Sun, C.; Wang, Y.; Yang, X.; Tang, L.; Wan, C.; Liu, J.; Chen, C.; Zhang, H.; He, C.; Liu, C. MATE transporter GFD1 cooperates with sugar transporters, mediates carbohydrate partitioning and controls grain-filling duration, grain size and number in rice. Plant Biotechnol. J. 2023, 21, 621–634. [Google Scholar] [CrossRef] [PubMed]
- Boorer, K.J.; Loo, D.F.; Frommer, W.B.; Wright, E.M. Transport mechanism of the cloned potato H+/sucrose cotransporter StSUT1. J. Biol. Chem. 1996, 271, 25139–25144. [Google Scholar] [CrossRef] [PubMed]
- Lemoine, R. Sucrose transporters in plants: Update on function and structure. Biochim. Biophys. Acta (BBA)-Biomembr. 2000, 1465, 246–262. [Google Scholar] [CrossRef]
- Khanna, K.; Ohri, P.; Bhardwaj, R. Decoding sugar regulation and homeostasis in plants: Cracking functional roles under stresses. J. Plant Growth Regul. 2023, 42, 4797–4817. [Google Scholar] [CrossRef]
- Wahl, R.; Wippel, K.; Goos, S.; Kämper, J.; Sauer, N. A novel high-affinity sucrose transporter is required for virulence of the plant pathogen Ustilago maydis. PLoS Biol. 2010, 8, e1000303. [Google Scholar] [CrossRef]
- Scholes, J.; Lee, P.; Horton, P.; Lewis, D. Invertase: Understanding changes in the photosynthetic and carbohydrate metabolism of barley leaves infected with powdery mildew. New Phytol. 1994, 126, 213–222. [Google Scholar] [CrossRef]
- TANG, X.; Rolfe, S.; Scholes, J. The effect of Albugo candida (white blister rust) on the photosynthetic and carbohydrate metabolism of leaves of Arabidopsis thaliana. Plant Cell Environ. 1996, 19, 967–975. [Google Scholar] [CrossRef]
- Chen, D.; Kamran, M.; Chen, S.; Xing, J.; Qu, Z.; Liu, C.; Ren, Z.; Cai, X.; Chen, X.L.; Xu, J. Two nucleotide sugar transporters are important for cell wall integrity and full virulence of Magnaporthe oryzae. Mol. Plant Pathol. 2023, 24, 374–390. [Google Scholar] [CrossRef]
- Chen, L.; Chen, B.; Zhu, Q.-H.; Zhang, X.; Sun, T.; Liu, F.; Yang, Y.; Sun, J.; Li, Y. Identification of sugar transporter genes and their roles in the pathogenicity of Verticillium dahliae on cotton. Front. Plant Sci. 2023, 14, 1123523. [Google Scholar] [CrossRef] [PubMed]
- Voegele, R.T.; Struck, C.; Hahn, M.; Mendgen, K. The role of haustoria in sugar supply during infection of broad bean by the rust fungus Uromyces fabae. Proc. Natl. Acad. Sci. USA 2001, 98, 8133–8138. [Google Scholar] [CrossRef] [PubMed]
- Schüßler, A.; Martin, H.; Cohen, D.; Fitz, M.; Wipf, D. Characterization of a carbohydrate transporter from symbiotic glomeromycotan fungi. Nature 2006, 444, 933–936. [Google Scholar] [CrossRef] [PubMed]
- Polidori, E.; Ceccaroli, P.; Saltarelli, R.; Guescini, M.; Menotta, M.; Agostini, D.; Palma, F.; Stocchi, V. Hexose uptake in the plant symbiotic ascomycete Tuber borchii Vittadini: Biochemical features and expression pattern of the transporter TBHXT1. Fungal Genet. Biol. 2007, 44, 187–198. [Google Scholar] [CrossRef] [PubMed]
- Herbers, K.; Meuwly, P.; Métraux, J.-P.; Sonnewald, U. Salicylic acid-independent induction of pathogenesis-related protein transcripts by sugars is dependent on leaf developmental stage. Febs Lett. 1996, 397, 239–244. [Google Scholar] [CrossRef] [PubMed]
- Ehness, R.; Ecker, M.; Godt, D.E.; Roitsch, T. Glucose and stress independently regulate source and sink metabolism and defense mechanisms via signal transduction pathways involving protein phosphorylation. Plant Cell 1997, 9, 1825–1841. [Google Scholar] [CrossRef] [PubMed]
- Kocal, N.; Sonnewald, U.; Sonnewald, S. Cell wall-bound invertase limits sucrose export and is involved in symptom development and inhibition of photosynthesis during compatible interaction between tomato and Xanthomonas campestris pv vesicatoria. Plant Physiol. 2008, 148, 1523–1536. [Google Scholar] [CrossRef]
- Schaarschmidt, S.; Kopka, J.; Ludwig-Müller, J.; Hause, B. Regulation of arbuscular mycorrhization by apoplastic invertases: Enhanced invertase activity in the leaf apoplast affects the symbiotic interaction. Plant J. 2007, 51, 390–405. [Google Scholar] [CrossRef]
- Wu, J.-N. An Illustrated Chinese Materia Medica; Oxford University Press: Oxford, UK, 2005. [Google Scholar]
- Khanal, S.; Gaire, S.P.; Zhou, X.-G. Kernel Smut and False smut: The old-emerging diseases of rice—A review. Phytopathology 2023, 113, 931–944. [Google Scholar] [CrossRef]
- Chen, X.; Hai, D.; Tang, J.; Liu, H.; Huang, J.; Luo, C.; Hsiang, T.; Zheng, L. UvCom1 is an important regulator required for development and infection in the rice false smut fungus Ustilaginoidea virens. Phytopathology 2020, 110, 483–493. [Google Scholar] [CrossRef]
- Sun, Q.; Qian, Z.; Liu, H.; Zhang, Y.; Kong, R.; Cheng, S.; Man, J.; Zheng, L.; Huang, J.; Su, G. Occurrence and translocation of ustiloxins in rice false smut-occurred paddy fields, Hubei, China. Environ. Pollut. 2022, 307, 119460. [Google Scholar] [CrossRef] [PubMed]
- Cheng, S.; Liu, H.; Sun, Q.; Kong, R.; Letcher, R.J.; Liu, C. Occurrence of the fungus mycotoxin, ustiloxin A, in surface waters of paddy fields in Enshi, Hubei, China, and toxicity in Tetrahymena thermophila. Environ. Pollut. 2019, 251, 901–909. [Google Scholar] [CrossRef] [PubMed]
- Brooks, S.A.; Anders, M.M.; Yeater, K.M. Effect of cultural management practices on the severity of false smut and kernel smut of rice. Plant Dis. 2009, 93, 1202–1208. [Google Scholar] [CrossRef] [PubMed]
- Lu, D.-H.; Yang, X.-Q.; Mao, J.-H.; Ye, H.-L.; Wang, P.; Chen, Y.-P.; He, Z.-Q.; Chen, F. Characterising the pathogenicity diversity of ustiiaginoidea virens in hybrid rice in China. J. Plant Pathol. 2009, 91, 443–451. [Google Scholar]
- Zhou, Y.; Xu, J.; Xie, X.; Gao, X.; Wang, J.; Li, Z. Effects of maturity group of cultivar, pathogen amount and sowing date on the severity of rice false smut in the field. Acta Phytophylacica Sin. 2010, 37, 97–103. [Google Scholar]
- Fan, J.; Yang, J.; Wang, Y.Q.; Li, G.B.; Li, Y.; Huang, F.; Wang, W.M. Current understanding on Villosiclava virens, a unique flower-infecting fungus causing rice false smut disease. Mol. Plant Pathol. 2016, 17, 1321–1330. [Google Scholar] [CrossRef]
- Fan, J.; Liu, J.; Gong, Z.Y.; Xu, P.Z.; Hu, X.H.; Wu, J.L.; Li, G.B.; Yang, J.; Wang, Y.Q.; Zhou, Y.F. The false smut pathogen Ustilaginoidea virens requires rice stamens for false smut ball formation. Environ. Microbiol. 2020, 22, 646–659. [Google Scholar] [CrossRef]
- Song, J.H.; Wei, W.; Lv, B.; Lin, Y.; Yin, W.X.; Peng, Y.L.; Schnabel, G.; Huang, J.B.; Jiang, D.H.; Luo, C.X. Rice false smut fungus hijacks the rice nutrients supply by blocking and mimicking the fertilization of rice ovary. Environ. Microbiol. 2016, 18, 3840–3849. [Google Scholar] [CrossRef]
- Kühn, C.; Barker, L.; Bürkle, L.; Frommer, W.-B. Update on sucrose transport in higher plants. J. Exp. Bot. 1999, 50, 935–953. [Google Scholar] [CrossRef]
- van Bel, A.J. Interaction between sieve element and companion cell and the consequences for photoassimilate distribution. Two structural hardware frames with associated physiological software packages in dicotyledons? J. Exp. Bot. 1996, 47, 1129–1140. [Google Scholar] [CrossRef]
- Zhang, Y.; Zhang, K.; Fang, A.; Han, Y.; Yang, J.; Xue, M.; Bao, J.; Hu, D.; Zhou, B.; Sun, X. Specific adaptation of Ustilaginoidea virens in occupying host florets revealed by comparative and functional genomics. Nat. Commun. 2014, 5, 3849. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.-Q.; Li, G.-B.; Gong, Z.-Y.; Li, Y.; Huang, F.; Fan, J.; Wang, W.-M. Stachyose is a preferential carbon source utilized by the rice false smut pathogen, Villosiclava virens. Physiol. Mol. Plant Pathol. 2016, 96, 69–76. [Google Scholar] [CrossRef]
- Saitoh, H.; Hirabuchi, A.; Fujisawa, S.; Mitsuoka, C.; Terauchi, R.; Takano, Y. MoST1 encoding a hexose transporter-like protein is involved in both conidiation and mycelial melanization of Magnaporthe oryzae. FEMS Microbiol. Lett. 2014, 352, 104–113. [Google Scholar] [CrossRef] [PubMed]
- Kumar, A.; Ghosh, S.; Bhatt, D.N.; Narula, A.; Datta, A. M agnaporthe oryzae aminosugar metabolism is essential for successful host colonization. Environ. Microbiol. 2016, 18, 1063–1077. [Google Scholar] [CrossRef] [PubMed]
- Gao, Y.; Xiong, X.; Wang, H.; Wang, J.; Bi, Y.; Yan, Y.; Cao, Z.; Li, D.; Song, F. Ero1-Pdi1 module-catalysed dimerization of a nucleotide sugar transporter, FonNst2, regulates virulence of Fusarium oxysporum on watermelon. Environ. Microbiol. 2022, 24, 1200–1220. [Google Scholar] [CrossRef]
- Bluhm, B.; Kim, H.; Butchko, R.; Woloshuk, C. Involvement of ZFR1 of Fusarium verticillioides in kernel colonization and the regulation of FST1, a putative sugar transporter gene required for fumonisin biosynthesis on maize kernels. Mol. Plant Pathol. 2008, 9, 203–211. [Google Scholar] [CrossRef]
- Liu, X.; Matsumoto, H.; Lv, T.; Zhan, C.; Fang, H.; Pan, Q.; Xu, H.; Fan, X.; Chu, T.; Chen, S. Phyllosphere microbiome induces host metabolic defence against rice false-smut disease. Nat. Microbiol. 2023, 8, 1419–1433. [Google Scholar] [CrossRef]
- Chen, X.; Pei, Z.; Liu, H.; Huang, J.; Chen, X.; Luo, C.; Hsiang, T.; Zheng, L. Host-induced gene silencing of fungal-specific genes of Ustilaginoidea virens confers effective resistance to rice false smut. Plant Biotechnol. J. 2022, 20, 253. [Google Scholar] [CrossRef]
- Mantle, P.; Nisbet, L. Differentiation of Claviceps purpurea in axenic culture. Microbiology 1976, 93, 321–334. [Google Scholar] [CrossRef]
- Chen, L.-Q.; Hou, B.-H.; Lalonde, S.; Takanaga, H.; Hartung, M.L.; Qu, X.-Q.; Guo, W.-J.; Kim, J.-G.; Underwood, W.; Chaudhuri, B. Sugar transporters for intercellular exchange and nutrition of pathogens. Nature 2010, 468, 527–532. [Google Scholar] [CrossRef]
- Büttner, M. The monosaccharide transporter(-like) gene family in Arabidopsis. FEBS Lett. 2007, 581, 2318–2324. [Google Scholar] [CrossRef] [PubMed]
- Chiou, T.-J.; Bush, D.R. Molecular cloning, immunochemical localization to the vacuole, and expression in transgenic yeast and tobacco of a putative sugar transporter from sugar beet. Plant Physiol. 1996, 110, 511–520. [Google Scholar] [CrossRef] [PubMed]
- Qiao, K.; Lv, J.; Chen, L.; Wang, Y.; Ma, L.; Wang, J.; Wang, Z.; Wang, L.; Ma, Q.; Fan, S. GhSTP18, a member of sugar transport proteins family, negatively regulates salt stress in cotton. Physiol. Plant. 2023, 175, e13982. [Google Scholar] [CrossRef] [PubMed]
- Rose, A.B. Introns as gene regulators: A brick on the accelerator. Front. Genet. 2019, 9, 672. [Google Scholar] [CrossRef] [PubMed]
- Mattick, J.S. Introns: Evolution and function. Curr. Opin. Genet. Dev. 1994, 4, 823–831. [Google Scholar] [CrossRef]
- Julius, B.T.; Leach, K.A.; Tran, T.M.; Mertz, R.A.; Braun, D.M. Sugar transporters in plants: New insights and discoveries. Plant Cell Physiol. 2017, 58, 1442–1460. [Google Scholar] [CrossRef]
- Afoufa-Bastien, D.; Medici, A.; Jeauffre, J.; Coutos-Thévenot, P.; Lemoine, R.; Atanassova, R.; Laloi, M. The Vitis vinifera sugar transporter gene family: Phylogenetic overview and macroarray expression profiling. BMC Plant Biol. 2010, 10, 245. [Google Scholar] [CrossRef] [PubMed]
- Li, J.M.; Zheng, D.M.; Li, L.T.; Qiao, X.; Wei, S.W.; Bai, B.; Zhang, S.L.; Wu, J. Genome-wide function, evolutionary characterization and expression analysis of sugar transporter family genes in pear (Pyrus bretschneideri Rehd). Plant Cell Physiol. 2015, 56, 1721–1737. [Google Scholar] [CrossRef]
- Liu, Q.; Dang, H.; Chen, Z.; Wu, J.; Chen, Y.; Chen, S.; Luo, L. Genome-wide identification, expression, and functional analysis of the sugar transporter gene family in cassava (Manihot esculenta). Int. J. Mol. Sci. 2018, 19, 987. [Google Scholar] [CrossRef]
- Li, J.; Lin, L.; Li, H.; Tian, C.; Ma, Y. Transcriptional comparison of the filamentous fungus Neurospora crassa growing on three major monosaccharides D-glucose, D-xylose and L-arabinose. Biotechnol. Biofuels 2014, 7, 31. [Google Scholar] [CrossRef]
- Gao, J.; Wang, B.; Han, X.; Tian, C. Genome-wide screening of predicted sugar transporters in Neurospora crassa and the application in hexose fermentation by Saccharomyces cerevisiae. Chin. J. Biotechnol. 2017, 33, 79–89. [Google Scholar]
- Wittek, A.; Dreyer, I.; Al-Rasheid, K.A.; Sauer, N.; Hedrich, R.; Geiger, D. The fungal UmSrt1 and maize ZmSUT1 sucrose transporters battle for plant sugar resources. J. Integr. Plant Biol. 2017, 59, 422–435. [Google Scholar] [CrossRef] [PubMed]
- Yin, W.; Cui, P.; Wei, W.; Lin, Y.; Luo, C. Genome-wide identification and analysis of the basic leucine zipper (bZIP) transcription factor gene family in Ustilaginoidea virens. Genome 2017, 60, 1051–1059. [Google Scholar] [CrossRef] [PubMed]
- Finn, R.D.; Clements, J.; Eddy, S.R. HMMER web server: Interactive sequence similarity searching. Nucleic Acids Res. 2011, 39, W29–W37. [Google Scholar] [CrossRef] [PubMed]
- Finn, R.D.; Coggill, P.; Eberhardt, R.Y.; Eddy, S.R.; Mistry, J.; Mitchell, A.L.; Potter, S.C.; Punta, M.; Qureshi, M.; Sangrador-Vegas, A. The Pfam protein families database: Towards a more sustainable future. Nucleic Acids Res. 2016, 44, D279–D285. [Google Scholar] [CrossRef]
- Marchler-Bauer, A.; Bo, Y.; Han, L.; He, J.; Lanczycki, C.J.; Lu, S.; Chitsaz, F.; Derbyshire, M.K.; Geer, R.C.; Gonzales, N.R. CDD/SPARCLE: Functional classification of proteins via subfamily domain architectures. Nucleic Acids Res. 2017, 45, D200–D203. [Google Scholar] [CrossRef]
- Bjellqvist, B.; Hughes, G.J.; Pasquali, C.; Paquet, N.; Ravier, F.; Sanchez, J.C.; Frutiger, S.; Hochstrasser, D. The focusing positions of polypeptides in immobilized pH gradients can be predicted from their amino acid sequences. Electrophoresis 1993, 14, 1023–1031. [Google Scholar] [CrossRef] [PubMed]
- Hallgren, J.; Tsirigos, K.D.; Pedersen, M.D.; Almagro Armenteros, J.J.; Marcatili, P.; Nielsen, H.; Krogh, A.; Winther, O. DeepTMHMM predicts alpha and beta transmembrane proteins using deep neural networks. BioRxiv 2022. [Google Scholar] [CrossRef]
- Horton, P.; Park, K.-J.; Obayashi, T.; Fujita, N.; Harada, H.; Adams-Collier, C.; Nakai, K. WoLF PSORT: Protein localization predictor. Nucleic Acids Res. 2007, 35, W585–W587. [Google Scholar] [CrossRef]
- Hu, B.; Jin, J.; Guo, A.-Y.; Zhang, H.; Luo, J.; Gao, G. GSDS 2.0: An upgraded gene feature visualization server. Bioinformatics 2015, 31, 1296–1297. [Google Scholar] [CrossRef]
- Bailey, T.L.; Boden, M.; Buske, F.A.; Frith, M.; Grant, C.E.; Clementi, L.; Ren, J.; Li, W.W.; Noble, W.S. MEME SUITE: Tools for motif discovery and searching. Nucleic Acids Res. 2009, 37, W202–W208. [Google Scholar] [CrossRef] [PubMed]
- Chen, C.; Wu, Y.; Li, J.; Wang, X.; Zeng, Z.; Xu, J.; Liu, Y.; Feng, J.; Chen, H.; He, Y.; et al. TBtools-II: A “one for all, all for one” bioinformatics platform for biological big-data mining. Mol. Plant 2023, 16, 1733–1742. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.; Tang, H.; DeBarry, J.D.; Tan, X.; Li, J.; Wang, X.; Lee, T.-h.; Jin, H.; Marler, B.; Guo, H. MCScanX: A toolkit for detection and evolutionary analysis of gene synteny and collinearity. Nucleic Acids Res. 2012, 40, e49. [Google Scholar] [CrossRef] [PubMed]
- Kruckeberg, A.L. The hexose transporter family of Saccharomyces cerevisiae. Arch. Microbiol. 1996, 166, 283–292. [Google Scholar] [CrossRef] [PubMed]
- Edgar, R.C. MUSCLE: Multiple sequence alignment with improved accuracy and speed. In Proceedings of the 2004 IEEE Computational Systems Bioinformatics Conference, Stanford, CA, USA, 16–19 August 2004; CSB 2004. pp. 728–729. [Google Scholar]
- Capella-Gutiérrez, S.; Silla-Martínez, J.M.; Gabaldón, T. trimAl: A tool for automated alignment trimming in large-scale phylogenetic analyses. Bioinformatics 2009, 25, 1972–1973. [Google Scholar] [CrossRef]
- Minh, B.Q.; Schmidt, H.A.; Chernomor, O.; Schrempf, D.; Woodhams, M.D.; Von Haeseler, A.; Lanfear, R. IQ-TREE 2: New models and efficient methods for phylogenetic inference in the genomic era. Mol. Biol. Evol. 2020, 37, 1530–1534. [Google Scholar] [CrossRef]
- Kalyaanamoorthy, S.; Minh, B.Q.; Wong, T.K.; Von Haeseler, A.; Jermiin, L.S. ModelFinder: Fast model selection for accurate phylogenetic estimates. Nat. Methods 2017, 14, 587–589. [Google Scholar] [CrossRef]
- Letunic, I.; Bork, P. Interactive tree of life (iTOL) v3: An online tool for the display and annotation of phylogenetic and other trees. Nucleic Acids Res. 2016, 44, W242–W245. [Google Scholar] [CrossRef]
- Untergasser, A.; Cutcutache, I.; Koressaar, T.; Ye, J.; Faircloth, B.C.; Remm, M.; Rozen, S.G. Primer3—New capabilities and interfaces. Nucleic Acids Res. 2012, 40, e115. [Google Scholar] [CrossRef]
- Livak, K.J.; Schmittgen, T.D. Analysis of relative gene expression data using real-time quantitative PCR and the 2−ΔΔCT method. Methods 2001, 25, 402–408. [Google Scholar] [CrossRef]
- Zhao, K.; Liu, L.; Huang, S. Genome-Wide Identification and Functional Analysis of the bZIP Transcription Factor Family in Rice Bakanae Disease Pathogen, Fusarium fujikuroi. Int. J. Mol. Sci. 2022, 23, 6658. [Google Scholar] [CrossRef] [PubMed]
- Yu, J.-H.; Hamari, Z.; Han, K.-H.; Seo, J.-A.; Reyes-Domínguez, Y.; Scazzocchio, C. Double-joint PCR: A PCR-based molecular tool for gene manipulations in filamentous fungi. Fungal Genet. Biol. 2004, 41, 973–981. [Google Scholar] [CrossRef] [PubMed]
- Xie, S.; Wang, Y.; Wei, W.; Li, C.; Liu, Y.; Qu, J.; Meng, Q.; Lin, Y.; Yin, W.; Yang, Y. The Bax inhibitor UvBI-1, a negative regulator of mycelial growth and conidiation, mediates stress response and is critical for pathogenicity of the rice false smut fungus Ustilaginoidea virens. Curr. Genet. 2019, 65, 1185–1197. [Google Scholar] [CrossRef] [PubMed]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Gene ID | Access Number | Chromosome | Gene Size | AA Length | PI | MW (kDa) | TMD | Subcellular Localization |
|---|---|---|---|---|---|---|---|---|
| VvSTP1 | XP_043001649 | 7 | 2199 | 732 | 7.27 | 80.93 | 12 | Plasma membrane |
| VvSTP2 | XP_043001702 | 7 | 2016 | 671 | 7.56 | 74.92 | 12 | Plasma membrane |
| VvSTP3 | XP_042999314 | 4 | 1659 | 552 | 8.95 | 61.50 | 12 | Plasma membrane |
| VvSTP4 | GAO15224 | 3 | 1882 | 541 | 9.02 | 59.64 | 12 | Plasma membrane |
| VvSTP5 | XP_042994690 | 1 | 2019 | 672 | 6.87 | 75.55 | 12 | Plasma membrane |
| VvSTP6 | XP_042994673 | 1 | 1899 | 632 | 8.76 | 70.19 | 12 | Plasma membrane |
| VvSTP7 | XP_042994564 | 1 | 1761 | 586 | 7.01 | 62.84 | 12 | Plasma membrane |
| VvSTP8 | XP_042996797 | 3 | 1584 | 527 | 9.22 | 57.39 | 12 | Plasma membrane |
| VvSTP9 | XP_042998161 | 4 | 1656 | 551 | 6.98 | 60.17 | 12 | Plasma membrane |
| VvSTP10 | XP_042994317 | 1 | 1599 | 532 | 6.9 | 58.27 | 12 | Plasma membrane |
| VvSTP11 | XP_042995010 | 1 | 1722 | 573 | 8.17 | 61.47 | 12 | Plasma membrane |
| VvSTP12 | XP_042995051 | 1 | 1611 | 536 | 9.04 | 57.44 | 12 | Plasma membrane |
| VvSTP13 | XP_043001089 | 6 | 1563 | 520 | 9.21 | 54.58 | 12 | Plasma membrane |
| VvSTP14 | XP_042998431 | 1 | 2142 | 713 | 9.09 | 77.91 | 12 | Plasma membrane |
| VvSTP15 | XP_042993682 | 1 | 1560 | 519 | 6.44 | 56.69 | 12 | Plasma membrane |
| VvSTP16 | XP_042994804 | 1 | 2334 | 777 | 9.34 | 86.56 | 12 | Plasma membrane |
| VvSTP17 | XP_042999475 | 5 | 1497 | 498 | 6.84 | 54.12 | 12 | Plasma membrane |
| VvSTP18 | XP_042993975 | 1 | 1677 | 558 | 9.14 | 60.71 | 12 | Plasma membrane |
| VvSTP19 | XP_042994240 | 1 | 1674 | 557 | 5.69 | 60.75 | 12 | Plasma membrane |
| VvSTP20 | XP_043001123 | 7 | 1494 | 497 | 8.92 | 54.59 | 12 | Plasma membrane |
| VvSTP21 | XP_042995111 | 1 | 1506 | 501 | 8.31 | 53.81 | 12 | Plasma membrane |
| VvSTP22 | XP_043000182 | 5 | 1584 | 527 | 8.6 | 57.75 | 12 | Plasma membrane |
| VvSTP23 | XP_043000657 | 6 | 1581 | 526 | 6.97 | 55.69 | 12 | Plasma membrane |
| VvSTP24 | XP_042998299 | 4 | 1656 | 551 | 5.13 | 60.48 | 12 | Plasma membrane |
| VvSTP25 | XP_042993458 | 1 | 1890 | 629 | 8.26 | 70.23 | 12 | Plasma membrane |
| VvSTP26 | XP_042998199 | 4 | 1656 | 551 | 9 | 59.96 | 12 | Plasma membrane |
| VvSTP27 | XP_042993861 | 1 | 1653 | 550 | 8.55 | 60.95 | 12 | Plasma membrane |
| VvSTP28 | XP_042993876 | 1 | 1752 | 583 | 5.64 | 64.89 | 12 | Plasma membrane |
| VvSTP29 | XP_043001720 | 7 | 1533 | 510 | 9.18 | 54.75 | 12 | Plasma membrane |
| VvSTP30 | XP_043000610 | 6 | 1626 | 541 | 9.26 | 59.40 | 12 | Plasma membrane |
| VvSTP31 | XP_042995206 | 2 | 1605 | 534 | 7.07 | 59.58 | 12 | Plasma membrane |
| VvSTP32 | XP_042995203 | 2 | 1608 | 535 | 8.55 | 58.13 | 12 | Plasma membrane |
| VvSTP33 | XP_042999770 | 5 | 1560 | 519 | 6.27 | 56.75 | 12 | Plasma membrane |
| VvSTP34 | XP_042998602 | 4 | 1674 | 557 | 7.97 | 61.46 | 12 | Plasma membrane |
| VvSTP35 | XP_042999744 | 5 | 1548 | 515 | 8.69 | 57.26 | 12 | Plasma membrane |
| Species | Number |
|---|---|
| Villosiclava virens | 35 |
| Aspergillus fumigatus | 89 |
| Claviceps purpurea | 33 |
| Cordyceps militaris | 57 |
| Fusarium graminearum | 116 |
| Metarhizium acridum | 89 |
| Metarhizium anisopliae | 62 |
| Metarhizium robertsii | 59 |
| Sclerotinia sclerotiorum | 61 |
| Magnaporthe oryzae | 122 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Qin, H.; Yin, W.; Luo, C.; Liu, L. The Identification, Characterization, and Functional Analysis of the Sugar Transporter Gene Family of the Rice False Smut Pathogen, Villosiclava virens. Int. J. Mol. Sci. 2024, 25, 600. https://doi.org/10.3390/ijms25010600
Qin H, Yin W, Luo C, Liu L. The Identification, Characterization, and Functional Analysis of the Sugar Transporter Gene Family of the Rice False Smut Pathogen, Villosiclava virens. International Journal of Molecular Sciences. 2024; 25(1):600. https://doi.org/10.3390/ijms25010600
Chicago/Turabian StyleQin, Huimin, Weixiao Yin, Chaoxi Luo, and Lianmeng Liu. 2024. "The Identification, Characterization, and Functional Analysis of the Sugar Transporter Gene Family of the Rice False Smut Pathogen, Villosiclava virens" International Journal of Molecular Sciences 25, no. 1: 600. https://doi.org/10.3390/ijms25010600