
Aluminum Supplementation Mediates the Changes in Tea Plant Growth and Metabolism in Response to Calcium Stress
 and
and {kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results and Discussion
2.1. Effects of Calcium and Aluminum Treatments on the Root Growth of Tea Plants
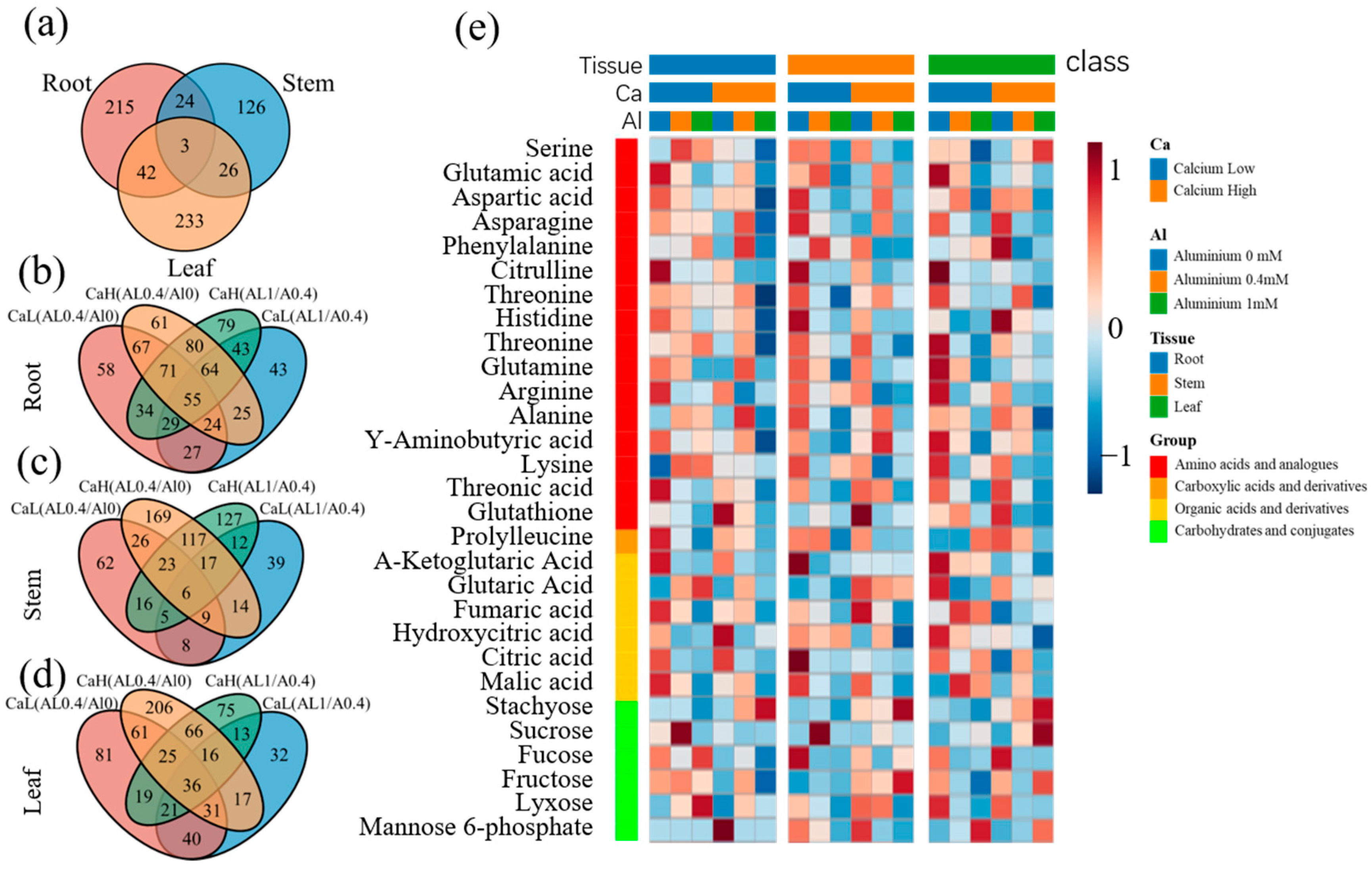
2.2. Differential Metabolites Identified with UPLC-Q-TOF/MS
2.3. Gene Expression Analysis
3. Materials and Methods
3.1. Plant Materials
3.2. Metabolomics Analyses
3.3. Quantitative Real-Time Polymerase Chain Reaction (qRT-PCR) Analysis
3.4. Statistical Analysis
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Lee, J.A. The Calcicole—Calcifuge Problem Revisited. In Advances in Botanical Research; Callow, J.A., Ed.; Academic Press: Cambridge, MA, USA, 1998; Volume 29, pp. 1–30. [Google Scholar]
- Su, H.; Li, L.; Ma, H.; Lyu, D.; Sun, J. Calcium Alleviates Temperature Stress by Regulating Nitrogen and Respiratory Metabolism in Malus Baccata Roots. IJAB 2016, 18, 286–292. [Google Scholar] [CrossRef]
- He, L.; Li, B.; Lu, X.; Yuan, L.; Yang, Y.; Yuan, Y.; Du, J.; Guo, S. The Effect of Exogenous Calcium on Mitochondria, Respiratory Metabolism Enzymes and Ion Transport in Cucumber Roots under Hypoxia. Sci. Rep. 2015, 5, 11391. [Google Scholar] [CrossRef] [PubMed]
- Giel, P.; Bojarczuk, K. Effects of High Concentrations of Calcium Salts in the Substrate and Its pH on the Growth of Selected Rhododendron Cultivars. Acta Soc. Bot. Pol. 2011, 80, 105–114. [Google Scholar] [CrossRef]
- Jones, J.; Massicotte, H.; Fredeen, A. Calcium and pH Co-Restrict Abundance of Drosera Rotundifolia (Droseraceae) in a Central BC Sphagnum Bog. Botany 2015, 94, 139–146. [Google Scholar] [CrossRef]
- Nunez, G.H.; Arzola, C.I.; Turner, A.; Leventini, D.; Doron, M. Root Tip Oxidative Stress Is Part of the Blueberry Response to High pH Nutrient Solutions. In Proceedings of the ASHS 2019 Annual Conference, Las Vegas, NV, USA, 21–25 July 2019. [Google Scholar]
- Ni, K.; Liao, W.; Yi, X.; Niu, S.; Ma, L.; Zhang, Q.F.; Liu, M.Y.; Ruan, J.Y. Fertilization status and reduction potential in tea gardens of China. J. Plant Nutr. Fertil. 2019, 25, 421–432. [Google Scholar]
- Li, C.-L.; Xu, J.; Xu, H.-M.; Liu, J.; Zhang, L.-X.; Wang, Z.-K. Analysis of Growth Resistance Mechanisms and Causes in Tea Plants (Camellia sinensis) in High-pH Regions of Northern China. Front. Nutr. 2023, 10, 1131380. [Google Scholar] [CrossRef] [PubMed]
- Ruan, J.; Gerendás, J.; Härdter, R.; Sattelmacher, B. Effect of Root Zone pH and Form and Concentration of Nitrogen on Accumulation of Quality-Related Components in Green Tea. J. Sci. Food Agric. 2007, 87, 1505–1516. [Google Scholar] [CrossRef]
- Xu, Q.; Wang, Y.; Ding, Z.; Song, L.; Li, Y.; Ma, D.; Wang, Y.; Shen, J.; Jia, S.; Sun, H.; et al. Aluminum Induced Metabolic Responses in Two Tea Cultivars. Plant Physiol. Biochem. 2016, 101, 162–172. [Google Scholar] [CrossRef]
- White, P.J.; Broadley, M.R. Calcium in Plants. Ann. Bot. 2003, 92, 487–511. [Google Scholar] [CrossRef]
- Zohlen, A.; Tyler, G. Immobilization of Tissue Iron on Calcareous Soil: Differences between Calcicole and Calcifuge Plants. Oikos 2000, 89, 95–106. [Google Scholar] [CrossRef]
- Tyler, G. A New Approach to Understanding the Calcifuge Habit of Plants. Ann. Bot. 1994, 73, 327–330. [Google Scholar] [CrossRef]
- Luo, Y.P. (Ed.) Cultivation of Tea Plants, 5th ed.; China Agricultural Publishing House: Beijing, China, 2015. (In Chinese) [Google Scholar]
- Jiazhi, S.; Xinfu, Z.; Jianhui, H. Dynamic Effects of Excessive Calcium on Biochemical Components in Young Shoots and Root Growth of Tea. Hubei Nongye Kexue 2014, 53, 4108–4111. [Google Scholar]
- Li, C.; Chen, Y. Effects of External Calcium on the Cell Ultrastructure of Tea Plants under Aluminium and Fluoride Stress. Fluoride 2018, 51, 243–251. [Google Scholar]
- Ruan, J.; Ma, L.; Shi, Y.; Han, W. The Impact of pH and Calcium on the Uptake of Fluoride by Tea Plants (Camellia sinensis L.). Ann. Bot. 2004, 93, 97–105. [Google Scholar] [CrossRef] [PubMed]
- Shu, W.; Zhang, Z.; Lan, C.; Wong, M. Fluoride and Aluminium Concentrations of Tea Plants and Tea Products from Sichuan Province, PR China. Chemosphere 2003, 52, 1475–1482. [Google Scholar] [CrossRef]
- Morita, A.; Horie, H.; Fujii, Y.; Takatsu, S.; Watanabe, N.; Yagi, A.; Yokota, H. Chemical Forms of Aluminum in Xylem Sap of Tea Plants (Camellia sinensis L.). Phytochemistry 2004, 65, 2775–2780. [Google Scholar] [CrossRef]
- Liu, Y.; Tao, J.; Cao, J.; Zeng, Y.; Li, X.; Ma, J.; Huang, Z.; Jiang, M.; Sun, L. The Beneficial Effects of Aluminum on the Plant Growth in Camellia Japonica. J. Soil Sci. Plant Nutr. 2020, 20, 1799–1809. [Google Scholar] [CrossRef]
- Mukhopadyay, M.; Bantawa, P.; Das, A.; Sarkar, B.; Bera, B.; Ghosh, P.; Mondal, T.K. Changes of Growth, Photosynthesis and Alteration of Leaf Antioxidative Defence System of Tea [Camellia sinensis (L.) O. Kuntze] Seedlings under Aluminum Stress. Biometals 2012, 25, 1141–1154. [Google Scholar] [CrossRef]
- Hajiboland, R.; Bahrami Rad, S.; Barceló, J.; Poschenrieder, C. Mechanisms of Aluminum-Induced Growth Stimulation in Tea (Camellia sinensis). J. Plant Nutr. Soil Sci. 2013, 176, 616–625. [Google Scholar] [CrossRef]
- Morita, A.; Yanagisawa, O.; Takatsu, S.; Maeda, S.; Hiradate, S. Mechanism for the Detoxification of Aluminum in Roots of Tea Plant (Camellia sinensis (L.) Kuntze). Phytochemistry 2008, 69, 147–153. [Google Scholar] [CrossRef]
- Gassmann, W.; Schroeder, J.I. Inward-Rectifying K+ Channels in Root Hairs of Wheat (A Mechanism for Aluminum-Sensitive Low-Affinity K+ Uptake and Membrane Potential Control). Plant Physiol. 1994, 105, 1399–1408. [Google Scholar] [CrossRef] [PubMed]
- Rengel, Z. Role of Calcium in Aluminium Toxicity. New Phytol. 1992, 121, 499–513. [Google Scholar] [CrossRef]
- Sivasubramaniam, S.; Talibudeen, O. Effect of Aluminium on Growth of Tea (Camellia sinensis) and Its Uptake of Potassium and Phosphorus. J. Sci. Food Agric. 1971, 22, 325–329. [Google Scholar] [CrossRef]
- Fung, K.F.; Wong, M.H. Effects of Soil pH on the Uptake of Al, F and Other Elements by Tea Plants. J. Sci. Food Agric. 2002, 82, 146–152. [Google Scholar] [CrossRef]
- Demarty, M.; Morvan, C.; Thellier, M. Calcium and the Cell Wall. Plant Cell Environ. 1984, 7, 441–448. [Google Scholar] [CrossRef]
- Pandey, S.; Tiwari, S.B.; Upadhyaya, K.C.; Sopory, S.K. Calcium Signaling: Linking Environmental Signals to Cellular Functions. Crit. Rev. Plant Sci. 2000, 19, 291–318. [Google Scholar] [CrossRef]
- Thor, K. Calcium—Nutrient and Messenger. Front. Plant Sci. 2019, 10, 440. [Google Scholar] [CrossRef]
- Jain, M.; Nagar, P.; Goel, P.; Singh, A.K.; Kumari, S.; Mustafiz, A. Second Messengers: Central Regulators in Plant Abiotic Stress Response. In Abiotic Stress-Mediated Sensing and Signaling in Plants: An Omics Perspective; Zargar, S.M., Zargar, M.Y., Eds.; Springer: Singapore, 2018; pp. 47–94. [Google Scholar]
- Sachs, J. A Text-Book of Botany: Morphological and Physiological; Bennett, A.W., Ed.; Cambridge Library Collection—Botany and Horticulture; Cambridge University Press: Cambridge, MA, USA, 2011. [Google Scholar]
- Liao, H.; Wan, H.; Shaff, J.; Wang, X.; Yan, X.; Kochian, L.V. Phosphorus and Aluminum Interactions in Soybean in Relation to Aluminum Tolerance. Exudation of Specific Organic Acids from Different Regions of the Intact Root System. Plant Physiol. 2006, 141, 674–684. [Google Scholar] [CrossRef]
- Iqbal, S.; Hussain, S.; Quayyaum, M.A.; Ashraf, M.; Saifullah, S. The Response of Maize Physiology under Salinity Stress and Its Coping Strategies. In Plant Stress Physiology; IntechOpen: London, UK, 2020. [Google Scholar]
- Meriño-Gergichevich, C.; Alberdi, M.; Ivanov, A.G.; Reyes-Diaz, M. Al3+-Ca2+ Interaction in Plants Growing in Acid Soils: Al-Phytotoxicity Response to Calcareous Amendments. J. Soil Sci. Plant Nutr. 2010, 10, 217–243. [Google Scholar]
- Zhang, Q.; Hu, J.; Liu, M.; Shi, Y.; De Vos, R.C.; Ruan, J. Stimulated Biosynthesis of Delphinidin-Related Anthocyanins in Tea Shoots Reducing the Quality of Green Tea in Summer. J. Sci. Food Agric. 2020, 100, 1505–1514. [Google Scholar] [CrossRef]
- Liang, L.L.; Song, Y.; Qian, W.; Ruan, J.; Ding, Z.T.; Zhang, Q.F.; Hu, J.H. Metabolomics Analysis Reveals the Responses of Tea Plants to Excessive Calcium. J. Sci. Food Agric. 2021, 101, 5678–5687. [Google Scholar] [CrossRef] [PubMed]








Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zhang, H.; Song, Y.; Fan, Z.; Ruan, J.; Hu, J.; Zhang, Q. Aluminum Supplementation Mediates the Changes in Tea Plant Growth and Metabolism in Response to Calcium Stress. Int. J. Mol. Sci. 2024, 25, 530. https://doi.org/10.3390/ijms25010530
Zhang H, Song Y, Fan Z, Ruan J, Hu J, Zhang Q. Aluminum Supplementation Mediates the Changes in Tea Plant Growth and Metabolism in Response to Calcium Stress. International Journal of Molecular Sciences. 2024; 25(1):530. https://doi.org/10.3390/ijms25010530
Chicago/Turabian StyleZhang, Hua, Yakang Song, Zhenglei Fan, Jianyun Ruan, Jianhui Hu, and Qunfeng Zhang. 2024. "Aluminum Supplementation Mediates the Changes in Tea Plant Growth and Metabolism in Response to Calcium Stress" International Journal of Molecular Sciences 25, no. 1: 530. https://doi.org/10.3390/ijms25010530