
Teleost Eye Is the Portal of IHNV Entry and Contributes to a Robust Mucosal Immune Response
Abstract
:1. Introduction
2. Results
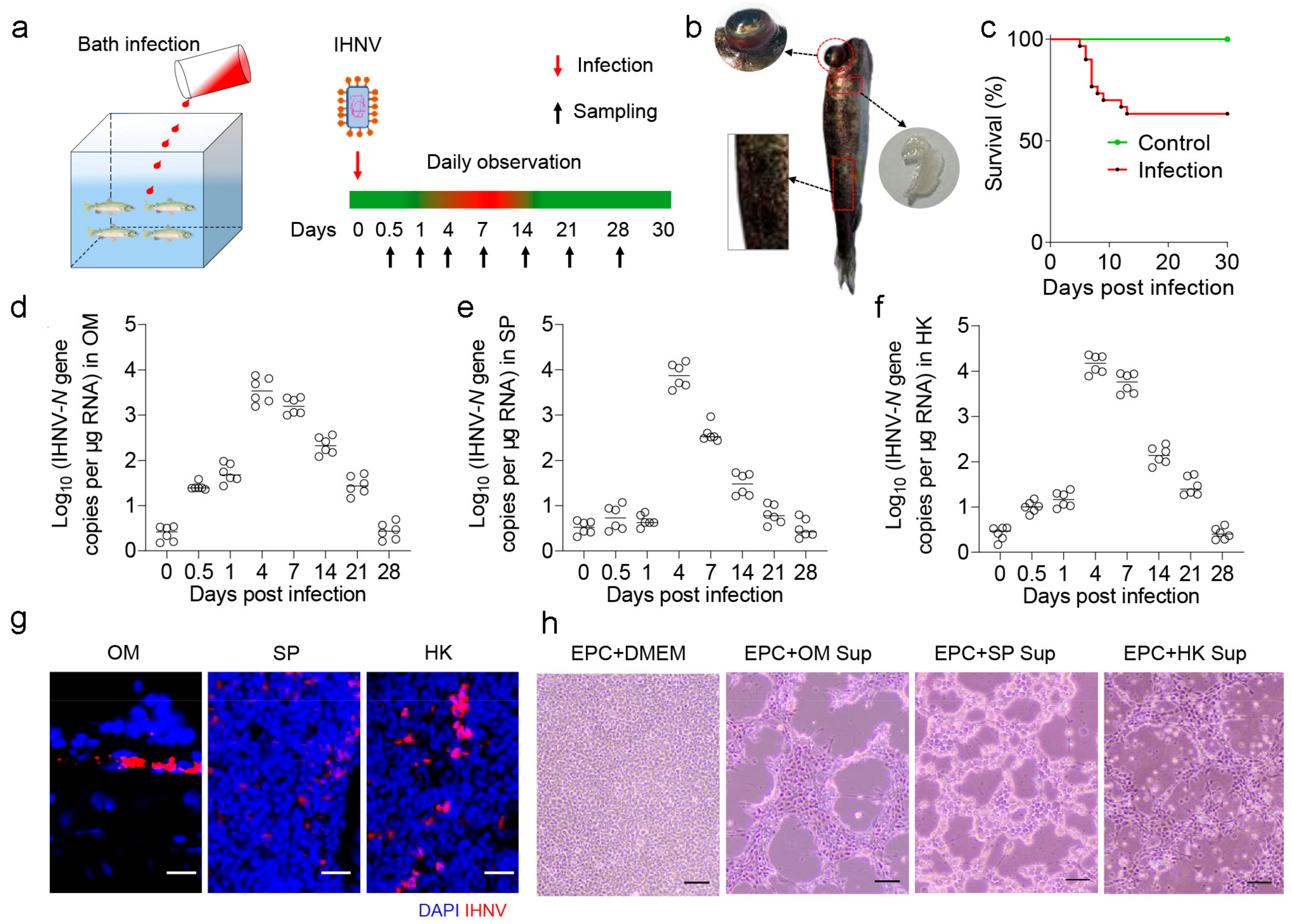
2.1. Establishment of IHNV Virus Infection Model in Rainbow Trout
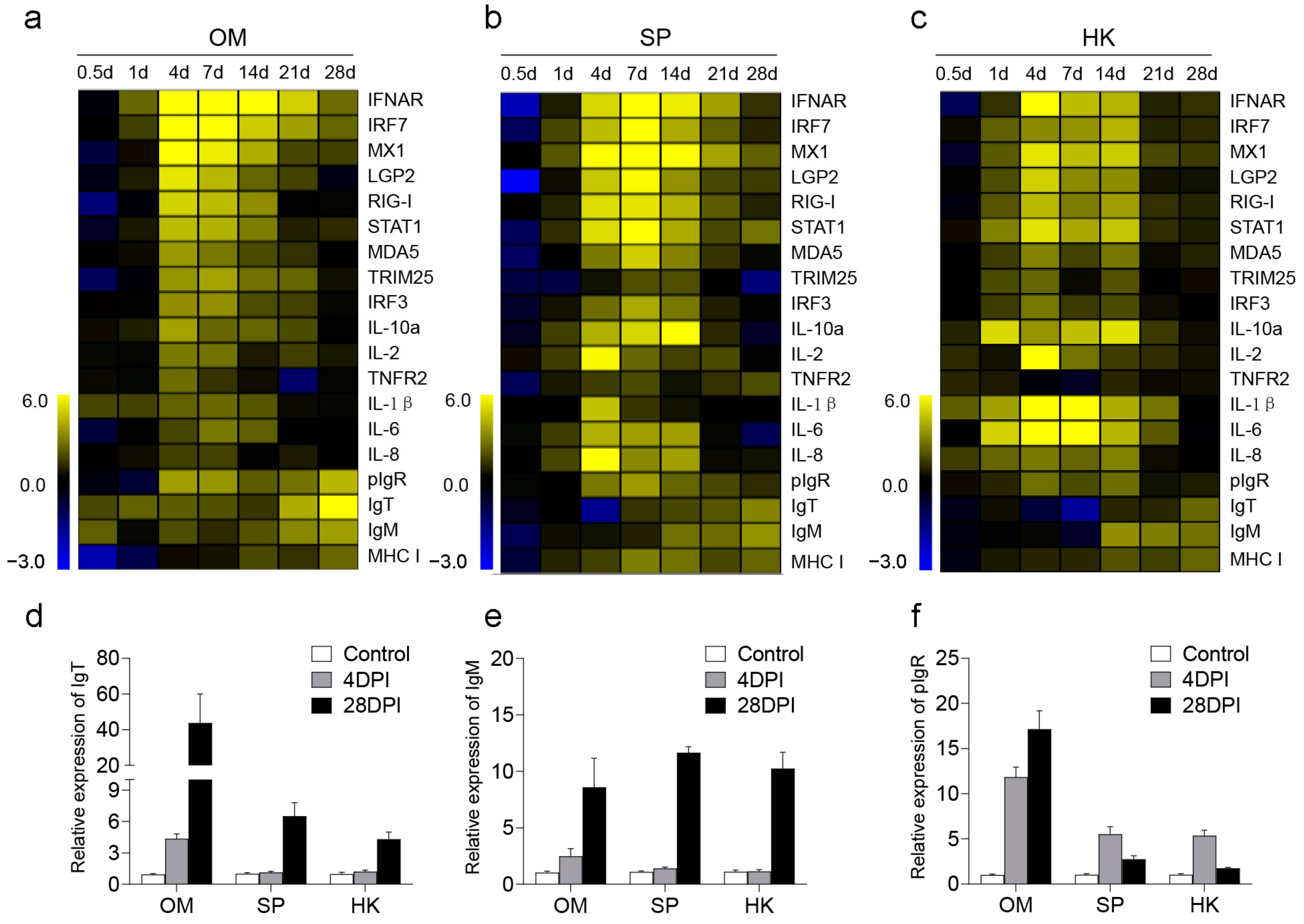
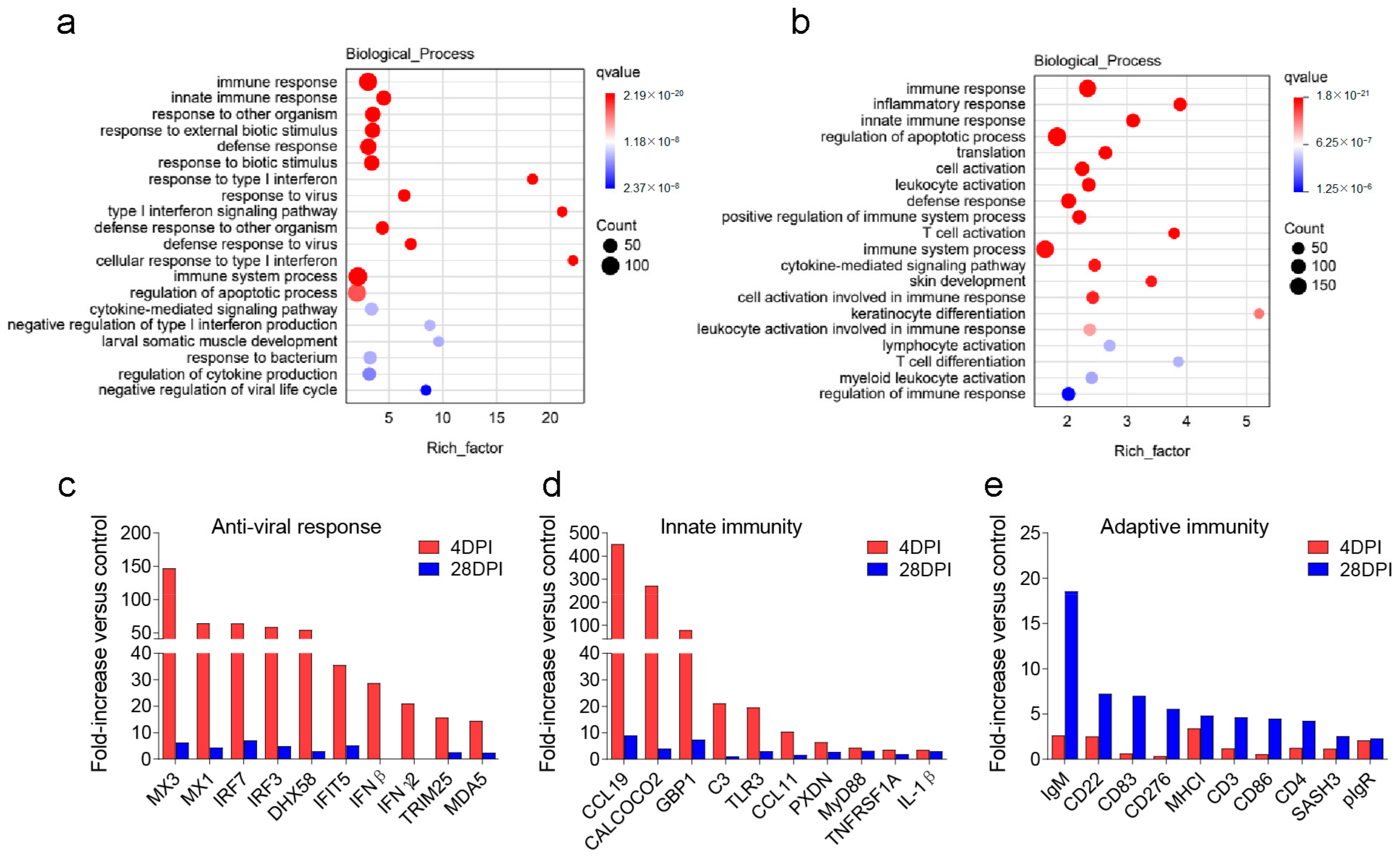
2.2. Innate and Adaptive Immunity Genes Were Induced in OM, SP, and HK after Viral Infection
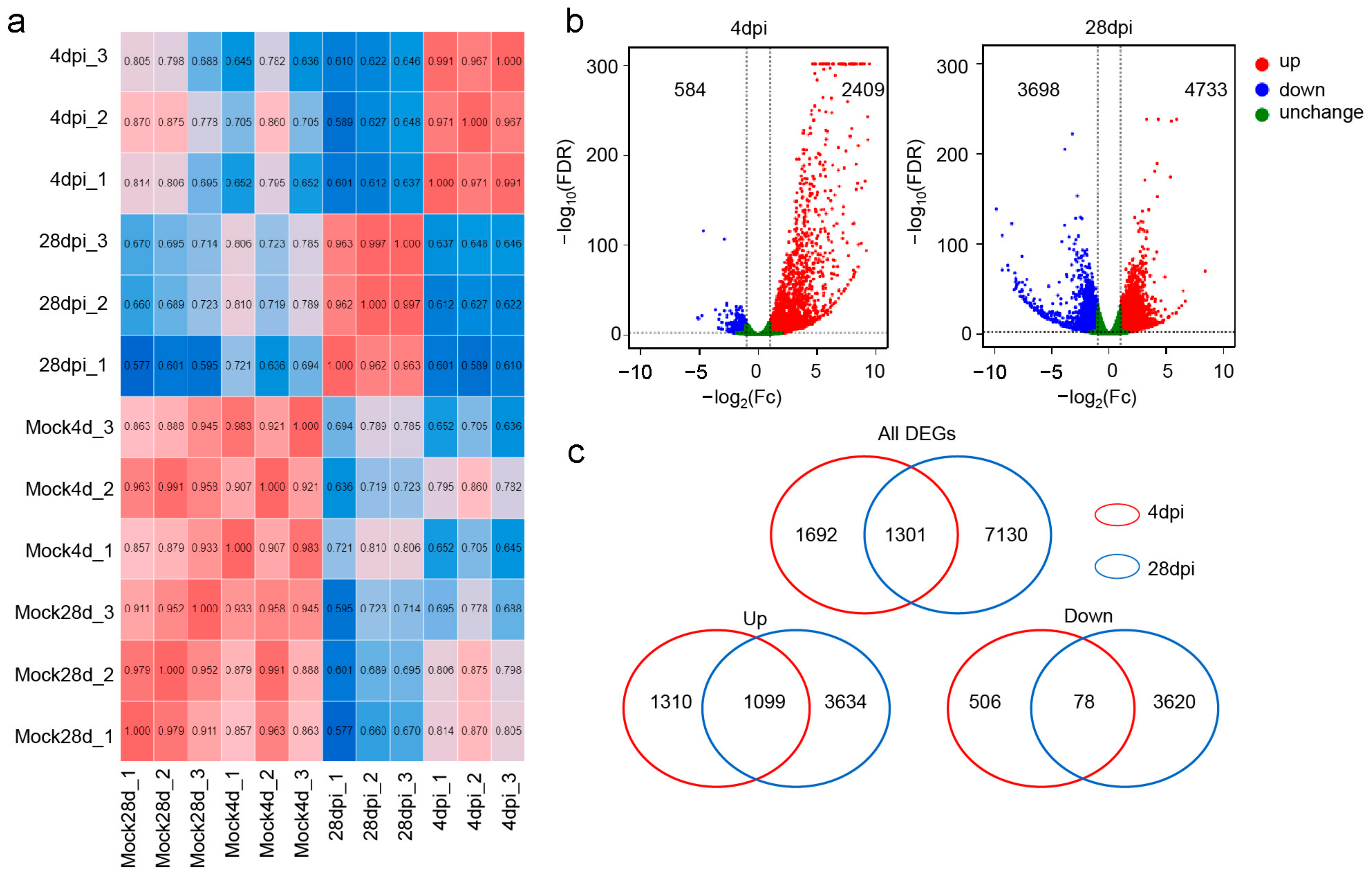
2.3. Dentification of the Transcriptome in the OM after IHNV Infection
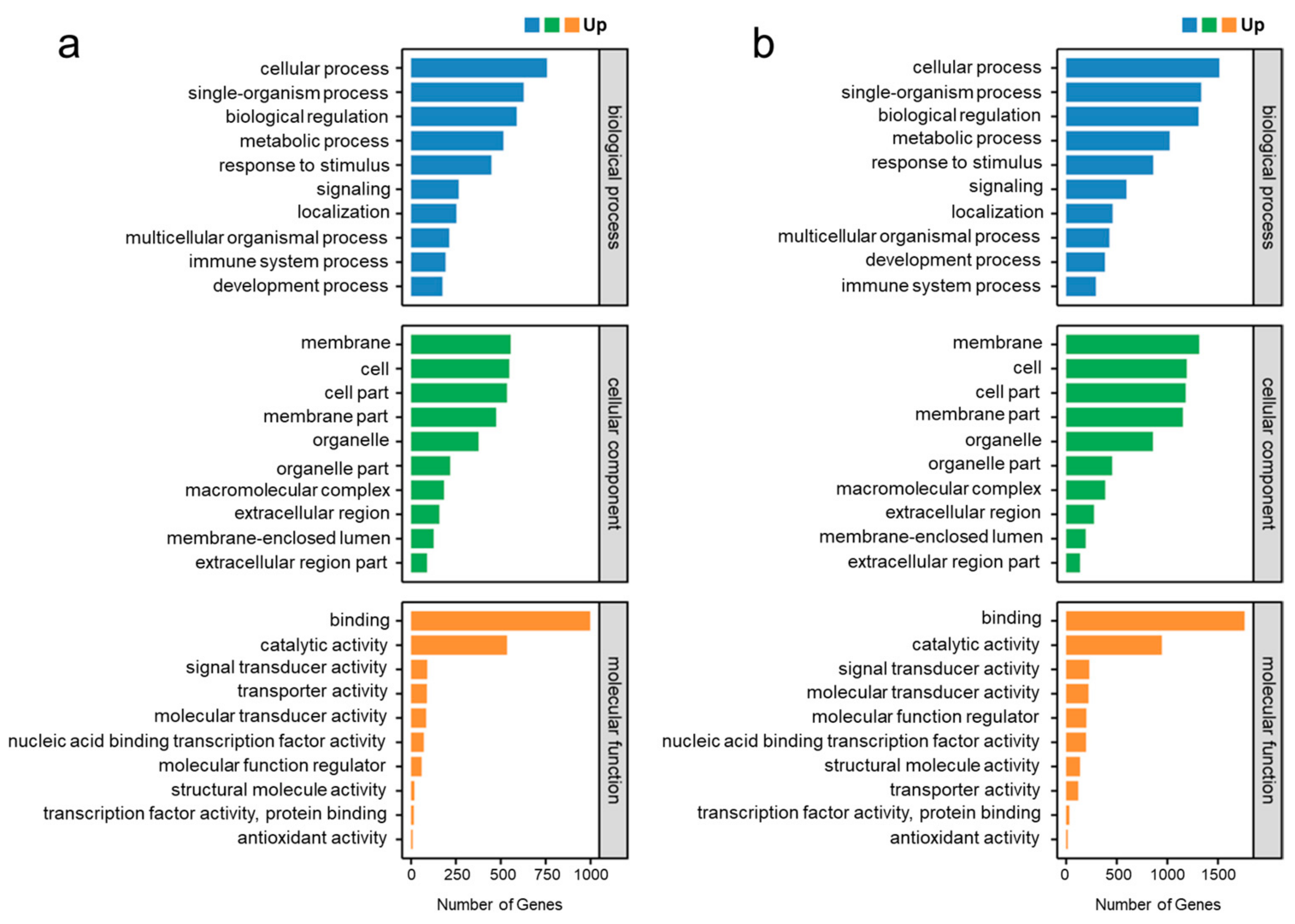
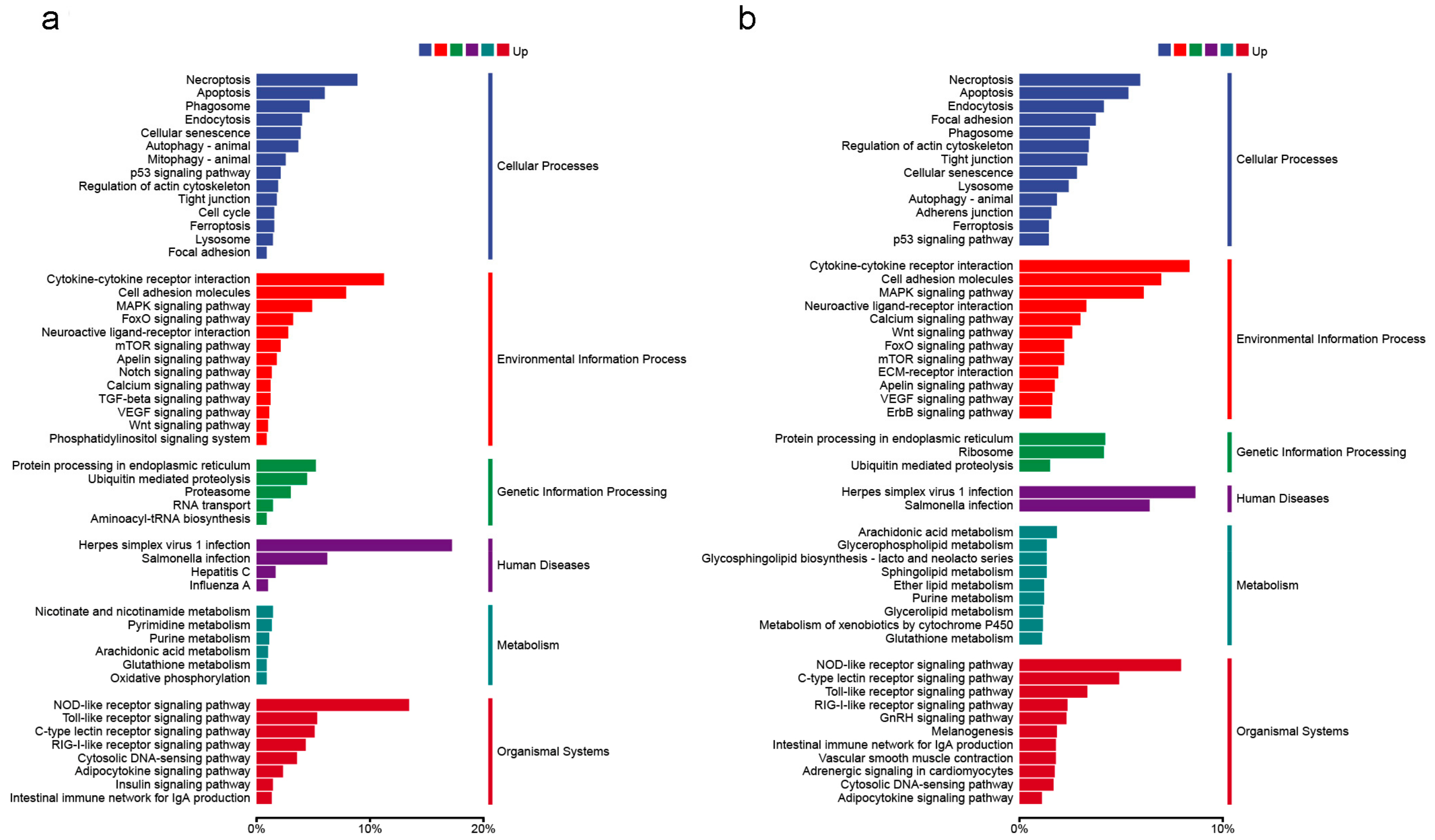
2.4. Functional Annotation and Classification of Genes
2.5. Classification and Analysis of DEGs
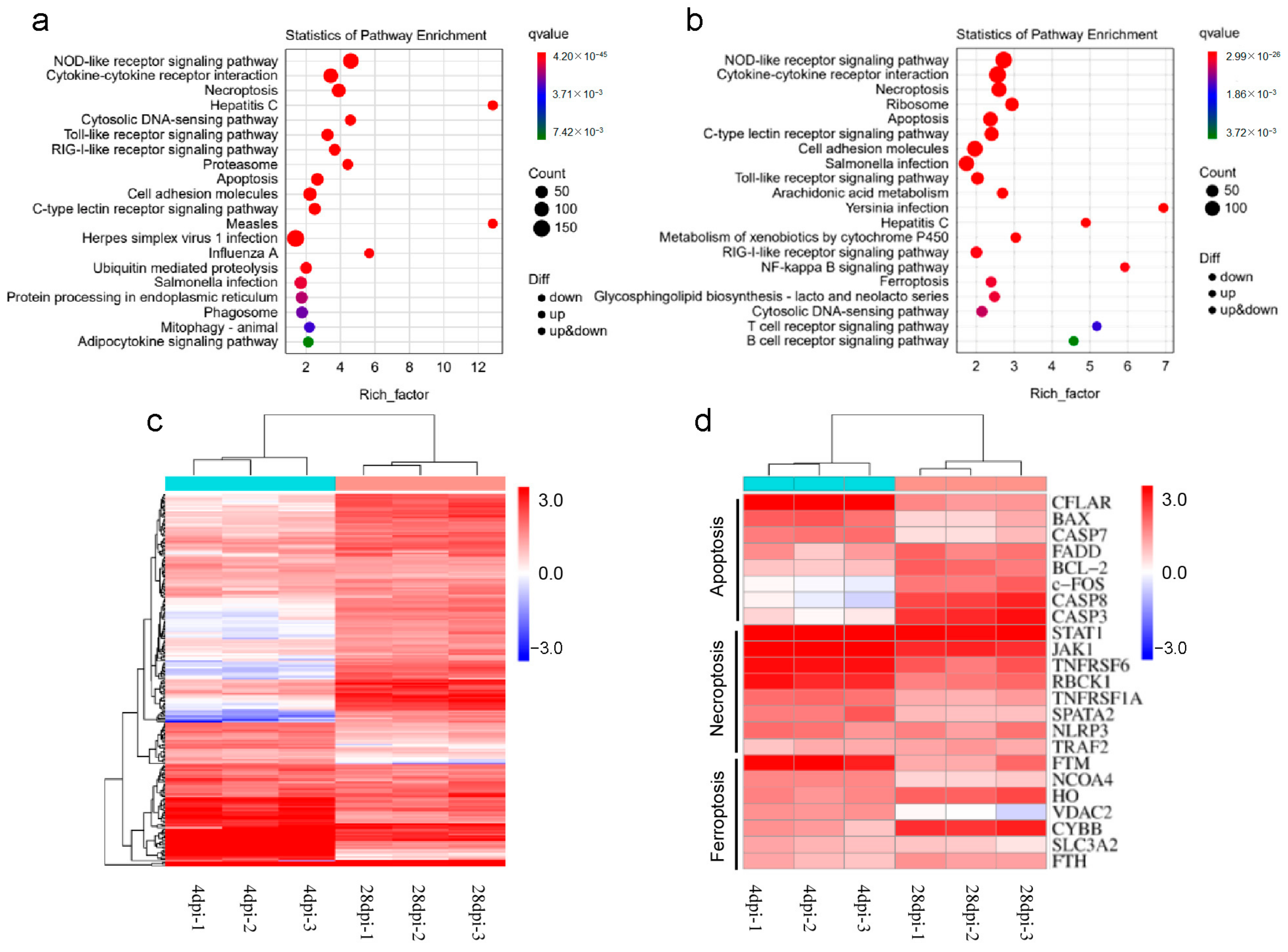
2.6. Enrichment and Analysis of Pathways after Infection
2.7. Enrichment and Analysis of Pathways after Infection
3. Discussion
4. Materials and Methods
4.1. Fish Maintenance
4.2. Infection of Fish with IHNV and Sample Collection
4.3. Standard Curve for IHNV
4.4. RNA Isolation and qPCR Analysis
4.5. Immunofluorescence Studies
4.6. Plaque Assay of IHNV
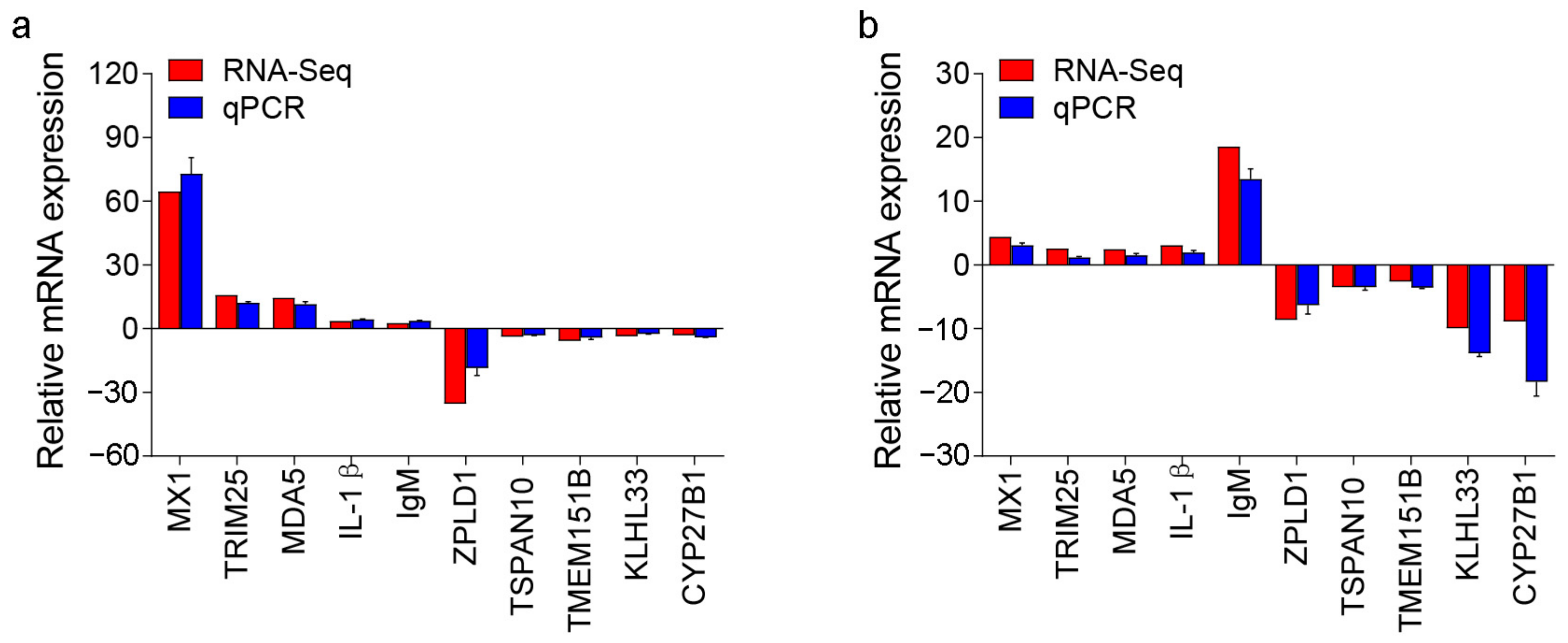
4.7. RNA-Seq Library Construction, Sequencing, and Data Analyses
4.8. Statistical Analysis
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Bergman, J. Did eyes evolve by Darwinian mechanisms. J. Creat. 2008, 22, 67–74. [Google Scholar]
- Lamb, T.D.; Collin, S.P.; Pugh, E.N., Jr. Evolution of the vertebrate eye: Opsins, photoreceptors, retina and eye cup. Nat. Rev. Neurosci. 2007, 8, 960–976. [Google Scholar] [CrossRef] [PubMed]
- Armstrong, R.A.; Cubbidge, R.P. The Eye and Vision: An Overview. In Handbook of Nutrition, Diet and the Eye, 2nd ed.; Preedy, V.R., Watson, R.R., Eds.; Academic Press: Cambridge, MA, USA, 2014; pp. 3–9. [Google Scholar]
- Williams, D. Ophthalmology of cartilaginous fish: Skates, rays, and sharks. In Wild and Exotic Animal Ophthalmology; Montiani-Ferreira, F., Moore, B.A., Ben-Shlomo, G., Eds.; Springer: Cham, Switzerland, 2022; pp. 47–59. [Google Scholar]
- Chang, C.H.; Chiao, C.C.; Yan, H.Y. The structure and possible functions of the milkfish Chanos chanos adipose eyelid. J. Fish Biol. 2009, 75, 87–99. [Google Scholar] [CrossRef] [PubMed]
- O’Sullivan, N.L.; Montgomery, P.C. Ocular Mucosal Immunity. In Mucosal Immunology; Academic Press: Cambridge, MA, USA, 2015; pp. 1873–1897. [Google Scholar]
- Deepthi, K.G.; Prabagaran, S.R. Ocular bacterial infections: Pathogenesis and diagnosis. Microb. Pathog. 2020, 145, 104206. [Google Scholar] [CrossRef] [PubMed]
- Borkowski, P.K. New trends in ocular toxoplasmosis—The review. Prz. Epidemiol. 2001, 55, 483–493. [Google Scholar]
- Mun, J.J.; Tam, C.; Kowbel, D.; Hawgood, S.; Barnett, M.J.; Evans, D.J.; Fleiszig, S.M. Clearance of Pseudomonas aeruginosa from a healthy ocular surface involves surfactant protein D and is compromised by bacterial elastase in a murine null-infection model. Infect. Immun. 2009, 77, 2392–2398. [Google Scholar] [CrossRef] [PubMed]
- Stern, M.E.; Schaumburg, C.S.; Dana, R.; Calonge, M.; Niederkorn, J.Y.; Pflugfelder, S.C. Autoimmunity at the ocular surface: Pathogenesis and regulation. Mucosal Immunol. 2010, 3, 425–442. [Google Scholar] [CrossRef]
- Muto, T.; Imaizumi, S.; Kamoi, K. Viral Conjunctivitis. Viruses 2023, 15, 676. [Google Scholar] [CrossRef]
- Epling, J. Bacterial conjunctivitis. BMJ Clin. Evid. 2012, 2012, 0704. [Google Scholar]
- Satpathy, G.; Behera, H.S.; Ahmed, N.H. Chlamydial eye infections: Current perspectives. Indian J. Ophthalmol. 2017, 65, 97–102. [Google Scholar] [CrossRef]
- Armstrong, L.; Collin, J.; Mostafa, I.; Queen, R.; Figueiredo, F.C.; Lako, M. In the eye of the storm: SARS-CoV-2 infection and replication at the ocular surface? Stem Cells Transl. Med. 2021, 10, 976–986. [Google Scholar] [CrossRef] [PubMed]
- Kumaratunga, V.; Adams, V.J.; Donaldson, D.; Pont, R.T.; Stidworthy, M.F. Ocular pathology in aquarium fish with a focus on the Syngnathidae and Apogonidae families. J. Comp. Pathol. 2023, 200, 1–11. [Google Scholar] [CrossRef] [PubMed]
- Elkhalil, L.; Majd, Z.; Bakir, R.; Perez-Mendez, O.; Castro, G.; Poulain, P.; Lacroix, B.; Duhal, N.; Fruchart, J.C.; Luc, G. Fish-eye disease: Structural and in vivo metabolic abnormalities of high-density lipoproteins. Metab. Clin. Exp. 1997, 46, 474–483. [Google Scholar] [CrossRef] [PubMed]
- Kanai, M.; Koh, S.; Masuda, D.; Koseki, M.; Nishida, K. Clinical features and visual function in a patient with Fish-eye disease: Quantitative measurements and optical coherence tomography. Am. J. Ophthalmol. Case Rep. 2018, 10, 137–141. [Google Scholar] [CrossRef] [PubMed]
- Chentoufi, A.A.; BenMohamed, L. Mucosal Herpes Immunity and Immunopathology to Ocular and Genital Herpes Simplex Virus Infections. Clin. Dev. Immunol. 2012, 2012, 149135. [Google Scholar] [CrossRef]
- de Paiva, C.S.; St Leger, A.J.; Caspi, R.R. Mucosal immunology of the ocular surface. Mucosal Immunol. 2022, 15, 1143–1157. [Google Scholar] [CrossRef] [PubMed]
- Krunkosky, M.; García, M.; Beltran, G.; Williams, S.M.; Hurley, D.J.; Gogal, R.M., Jr. Ocular exposure to infectious laryngotracheitis virus alters leukocyte subsets in the head-associated lymphoid tissues and trachea of 6-week-old White Leghorn chickens. Avian Pathol. 2020, 49, 404–417. [Google Scholar] [CrossRef]
- Toro, H.; Suarez, D.L.; Tang, D.C.; van Ginkel, F.W.; Breedlovea, C. Avian influenza mucosal vaccination in chickens with replication-defective recombinant adenovirus vaccine. Avian Dis. 2011, 55, 43–47. [Google Scholar] [CrossRef]
- Eggestøl, H.Ø.; Lunde, H.S.; Rønneseth, A.; Fredman, D.; Petersen, K.; Mishra, C.K.; Furmanek, T.; Colquhoun, D.J.; Wergeland, H.I.; Haugland, G.T. Transcriptome-wide mapping of signaling pathways and early immune responses in lumpfish leukocytes upon in vitro bacterial exposure. Sci. Rep. 2018, 8, 5261. [Google Scholar] [CrossRef]
- Koppang, E.O.; Bjerkås, E.; Bjerkås, I.; Sveier, H.; Hordvik, I. Vaccination induces major histocompatibility complex class II expression in the Atlantic salmon eye. Scand. J. Immunol. 2003, 58, 9–14. [Google Scholar] [CrossRef]
- Yong, C.Y.; Ong, H.K.; Tang, H.C.; Yeap, S.K.; Omar, A.R.; Ho, K.L.; Tan, W.S. Infectious hematopoietic necrosis virus: Advances in diagnosis and vaccine development. PeerJ 2019, 7, e7151. [Google Scholar] [CrossRef] [PubMed]
- Dong, S.; Ding, L.G.; Cao, J.F.; Liu, X.; Xu, H.Y.; Meng, K.F.; Yu, Y.Y.; Wang, Q.; Xu, Z. Viral-infected change of the digestive tract microbiota associated with mucosal immunity in teleost fish. Front. Immunol. 2019, 10, 2878. [Google Scholar] [CrossRef] [PubMed]
- Huang, Z.; Zhan, M.; Cheng, G.; Lin, R.; Zhai, X.; Zheng, H.; Wang, Q.; Yu, Y.; Xu, Z. IHNV infection induces strong mucosal immunity and changes of microbiota in trout intestine. Viruses 2022, 14, 1838. [Google Scholar] [CrossRef]
- Kong, W.G.; Yu, Y.Y.; Dong, S.; Huang, Z.Y.; Ding, L.G.; Cao, J.F.; Dong, F.; Zhang, X.T.; Liu, X.; Xu, H.Y.; et al. Pharyngeal immunity in early vertebrates provides functional and evolutionary insight into mucosal homeostasis. J. Immunol. 2019, 203, 3054–3067. [Google Scholar] [CrossRef] [PubMed]
- Schwab, I. The evolution of eyes: Major steps. The Keeler lecture 2017: Centenary of Keeler Ltd. Eye 2018, 32, 302–313. [Google Scholar] [CrossRef] [PubMed]
- Musilova, Z.; Salzburger, W.; Cortesi, F. The Visual Opsin Gene Repertoires of Teleost Fishes: Evolution, Ecology, and Function. Annu. Rev. Cell Dev. Biol. 2021, 37, 441–468. [Google Scholar] [CrossRef] [PubMed]
- Ueta, M. Innate immunity of the ocular surface and ocular surface inflammatory disorders. Cornea 2008, 27, S31–S40. [Google Scholar] [CrossRef]
- Popov, A. Mucus-penetrating particles and the role of ocular mucus as a barrier to micro- and nanosuspensions. J. Ocul. Pharmacol. Ther. 2020, 36, 366–375. [Google Scholar] [CrossRef]
- Ahmadivand, S.; Soltani, M.; Mardani, K.; Shokrpoor, S.; Hassanzadeh, R.; Ahmadpoor, M.; Rahmati-Holasoo, H.; Meshkini, S. Infectious hematopoietic necrosis virus (IHNV) outbreak in farmed rainbow trout in Iran: Viral isolation, pathological findings, molecular confirmation, and genetic analysis. Virus Res. 2017, 229, 17–23. [Google Scholar] [CrossRef]
- Shao, Y.Z.; Zhao, J.Z.; Ren, G.M.; Lu, T.Y.; Xu, L.M. Infectious hematopoietic necrosis virus promoted infectious pancreatic necrosis virus multiplication during co-infection in rainbow trout (Oncorhynchus mykiss). Aquaculture 2022, 561, 738649. [Google Scholar] [CrossRef]
- Yu, Y.Y.; Xu, Y.S.; Chen, X.Y.; Xu, Z. Molecular cloning and expression analysis IgM, IgD and IgT heavy chain genes in Misgurnus anguillicaudates. Fish Shellfish Immunol. 2016, 53, 115. [Google Scholar] [CrossRef]
- Zhang, Y.A.; Salinas, I.; Li, J.; Parra, D.; Bjork, S.; Xu, Z.; LaPatra, S.E.; Bartholomew, J.; Sunyer, J.O. IgT, a primitive immunoglobulin class specialized in mucosal immunity. Nat. Immunol. 2010, 11, 827–835. [Google Scholar] [CrossRef] [PubMed]
- Sutherland, D.B.; Fagarasan, S. IgA synthesis: A form of functional immune adaptation extending beyond gut. Curr. Opin. Immunol. 2012, 24, 261–268. [Google Scholar] [CrossRef] [PubMed]
- Ueta, M.; Kinoshita, S. Innate immunity of the ocular surface. Brain Res. Bull. 2010, 81, 219–228. [Google Scholar] [CrossRef] [PubMed]
- Perry, A.K.; Chen, G.; Zheng, D.; Tang, H.; Cheng, G. The host type I interferon response to viral and bacterial infections. Cell Res. 2005, 15, 407–422. [Google Scholar] [CrossRef]
- Kim, Y.K.; Shin, J.S.; Nahm, M.H. NOD-like receptors in infection, immunity, and diseases. Yonsei Med. J. 2016, 57, 5–14. [Google Scholar] [CrossRef]
- Jin, J.; Zhou, T.J.; Ren, G.L.; Cai, L.; Meng, X.M. Novel insights into NOD-like receptors in renal diseases. Acta Pharmacol. Sin. 2022, 43, 2789–2806. [Google Scholar] [CrossRef]
- Shaw, P.J.; Lamkanfi, M.; Kanneganti, T.D. NOD-like receptor (NLR) signaling beyond the inflammasome. Eur. J. Immunol. 2010, 40, 624–627. [Google Scholar] [CrossRef]
- Akira, S. Toll-like receptor signaling. J. Biol. Chem. 2003, 278, 38105–38108. [Google Scholar] [CrossRef]
- Thompson, A.J.; Locarnini, S.A. Toll-like receptors, RIG-I-like RNA helicases and the antiviral innate immune response. Immunol. Cell Biol. 2007, 85, 435–445. [Google Scholar] [CrossRef]
- Yoneyama, M.; Kikuchi, M.; Natsukawa, T.; Shinobu, N.; Imaizumi, T.; Miyagishi, M.; Taira, K.; Akira, S.; Fujita, T. The RNA helicase RIG-I has an essential function in double-stranded RNA-induced innate antiviral responses. Nat. Immunol. 2004, 5, 730–737. [Google Scholar] [CrossRef] [PubMed]
- Kaileh, M.; Sen, R. NF-κB function in B lymphocytes. Immunol. Rev. 2012, 246, 254–271. [Google Scholar] [CrossRef] [PubMed]
- He, M.X.; He, Y.W. CFLAR/c-FLIPL: A star in the autophagy, apoptosis and necroptosis alliance. Autophagy 2013, 9, 791–793. [Google Scholar] [CrossRef] [PubMed]
- Hu, W.H.; Johnson, H.; Shu, H.B. Activation of NF-κB by FADD, Casper, and caspase-8. J. Biol. Chem. 2000, 275, 10838–10844. [Google Scholar] [CrossRef] [PubMed]
- Rawlings, J.S.; Rosler, K.M.; Harrison, D.A. The JAK/STAT signaling pathway. J. Cell Sci. 2004, 117, 1281–1283. [Google Scholar] [CrossRef] [PubMed]
- Liu, C.; Li, Z.; Xi, H. Bioinformatics analysis and in vivo validation of ferroptosis-related genes in ischemic stroke. Front. Pharmacol. 2022, 13, 940260. [Google Scholar] [CrossRef] [PubMed]
- Kong, W.; Yang, P.; Ding, G.; Cheng, G.; Xu, Z. Elucidating the dynamic immune responses within the ocular mucosa of rainbow trout (Oncorhynchus mykiss) after infection with Flavobacterium columnare. Front. Immunol. 2023, 14, 1288223. [Google Scholar] [CrossRef]
- Dobin, A.; Davis, C.A.; Schlesinger, F.; Drenkow, J.; Zaleski, C.; Jha, S.; Batut, P.; Chaisson, M.; Gingeras, T.R. STAR: Ultrafast universal RNA-seq aligner. Bioinformatics 2013, 29, 15–21. [Google Scholar] [CrossRef]
- Liao, Y.; Smyth, G.K.; Shi, W. featureCounts: An efficient general purpose program for assigning sequence reads to genomic features. Bioinformatics 2014, 30, 923–930. [Google Scholar] [CrossRef]
- Robinson, M.D.; McCarthy, D.J.; Smyth, G.K. edgeR: A Bioconductor package for differential expression analysis of digital gene expression data. Bioinformatics 2010, 26, 139–140. [Google Scholar] [CrossRef]
- Wu, J.; Mao, X.; Cai, T.; Luo, J.; Wei, L. KOBAS server: A web-based platform for automated annotation and pathway identification. Nucleic Acids Res. 2006, 34, W720–W724. [Google Scholar] [CrossRef] [PubMed]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Gene | Forward Primer (5′-3′) | Reverse Primer (5′-3′) |
|---|---|---|
| EF1α | CAACGATATCCGTCGTGGCA | ACAGCGAAACGACCAAGAGG |
| MDA5 | CAGTGGAGATGACGATGGG | ACTTGGCGTTCTTGTGCTT |
| LGP2 | AGTTTGGCACGCAGGAGTA | CAAGCAGGAAGAAGTCGGT |
| IRF7 | GATGCCTCAAAATGAAATGG | TCAGGGAACTTCTCACCAAA |
| IFNAR | CAGAGCCTCAGGAAGAACT | CAAGGGGTAGAAGAGCATA |
| RIG-I | CAGAGGTACTACAGGAAATGG | TTACTGGTCTTCAAGCAATG |
| STAT1 | CTCATCCCCTGGACCAAGTT | TTATTGTAGCCCTCCACCCA |
| TRIM25 | AAAGATTCACCCCAAAACC | AAGGCAGGGGAATCATAGT |
| IRF3 | GGAGACCACTCTGTTTGGAAG | CGGCATCGTTCTTGTTGTC |
| IL-10a | CGAAGGGATTCTACACCACTTG | GAGATTTAAAGTCGTTGTTGTTCTG |
| IL-2 | CGAACGGGACGAGAAATGG | TGATAAAGAGCTGCTGTGAATG |
| TNFR2 | GGAAAGGAGGCCATTAAATGTCTGC | CAGATGTCAGAGGGCTGGTGGA |
| pIgR | GATGACCCAGAATCACCCCC | GTCAGACACCCACATTCCCC |
| ZPLD1 | AAAGCCACAAACCTG | TTGACTCTTCCCATTC |
| TMEM151B | TGCGGTCAAATCGTC | ATGGCAGAATCAAGTCA |
| KLHL33 | AGGTATTCCAGGGTCT | GAACAAGGGTACTCACA |
| TSPAN10 | AACCTCCTCTTCACGG | CCACAAACACCAGCAT |
| CYP28B1 | AGGCTGTGGTCAAGG | GTGGCAGAGGGTGAT |
| IgM | AAGAAAGCCTACAAGAGGGAGA | CGTCAACAAGCCAAGCCACTA |
| IgT | CAGACAACAGCACCTCACCTA | GAGTCAATAAGAAGACACAACGA |
| IL-6 | GCTCGGGACAGACGGTGAAA | GAAACTCCTCCACAAACTGTTG |
| IL-8 | GGCCCTCCTGACCATTACTGA | TCCAGACAAATCTCCTGACCG |
| IL-1β | ACCGAGTTCAAGGACAAGGA | CATTCATCAGGACCCAGCAC |
| MHC-I | AGGCTGGAGTCACAGAGGG | CGTGGCAGGTCACTGGAGA |
| MX1 | GATGCTGCACCTCAAGTCCTACTA | CGGATCACCATGGGAATCTGA |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Wang, X.; Ding, G.; Yang, P.; Cheng, G.; Kong, W.; Xu, Z. Teleost Eye Is the Portal of IHNV Entry and Contributes to a Robust Mucosal Immune Response. Int. J. Mol. Sci. 2024, 25, 160. https://doi.org/10.3390/ijms25010160
Wang X, Ding G, Yang P, Cheng G, Kong W, Xu Z. Teleost Eye Is the Portal of IHNV Entry and Contributes to a Robust Mucosal Immune Response. International Journal of Molecular Sciences. 2024; 25(1):160. https://doi.org/10.3390/ijms25010160
Chicago/Turabian StyleWang, Xinyou, Guangyi Ding, Peng Yang, Gaofeng Cheng, Weiguang Kong, and Zhen Xu. 2024. "Teleost Eye Is the Portal of IHNV Entry and Contributes to a Robust Mucosal Immune Response" International Journal of Molecular Sciences 25, no. 1: 160. https://doi.org/10.3390/ijms25010160