
In Vitro Approaches to Determine the Potential Carcinogenic Risk of Environmental Pollutants
 , , and
, , and
Abstract
:1. Introduction
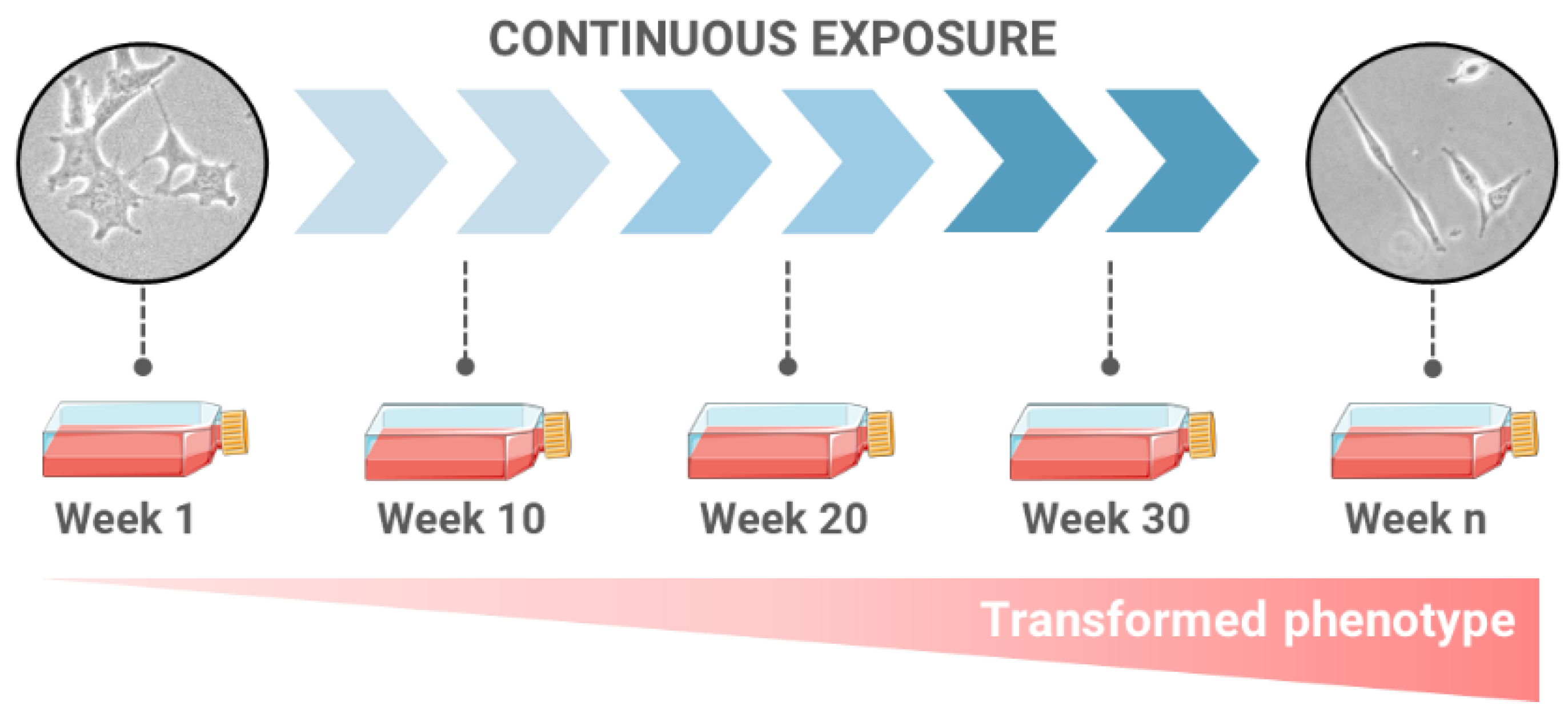
2. Long-Term Exposures as a Novel Approach for Carcinogenesis Evaluation
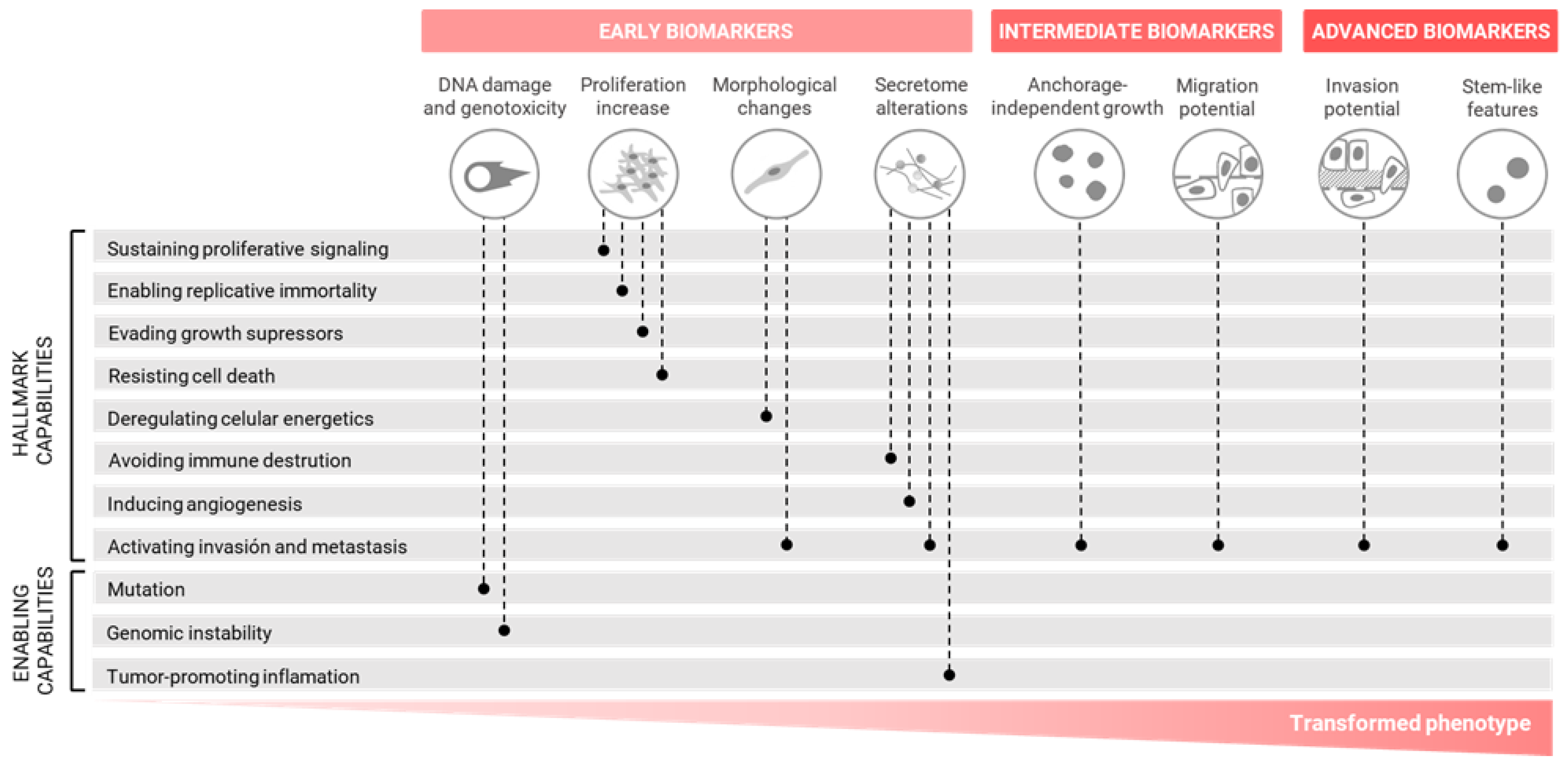
3. Alternative Cell Transformation Assays Based on the Hallmarks of Carcinogenesis
3.1. In Vitro Cell Transformation Biomarkers and Methods
3.1.1. Early Biomarkers
DNA Damage and Genotoxicity
Uncontrolled Proliferation
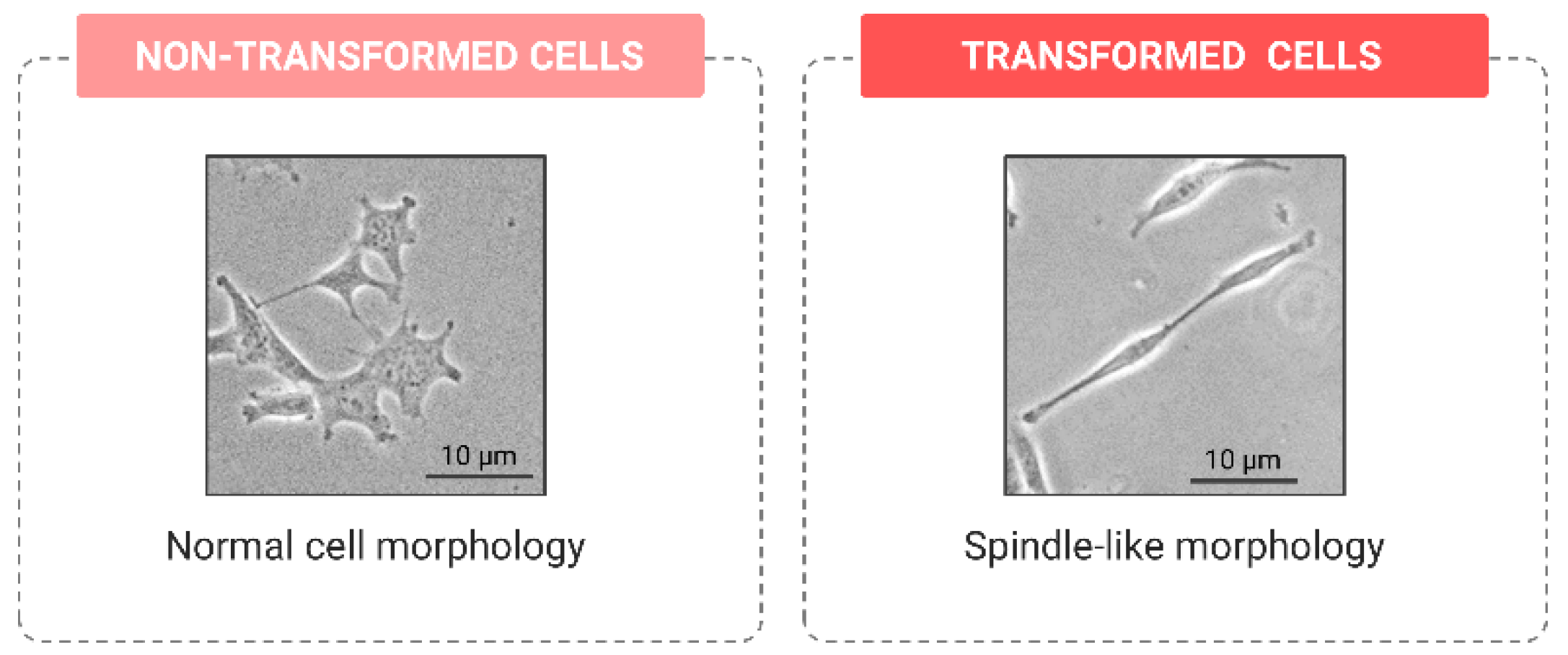
Morphological Changes
Secretome Alterations
3.1.2. Intermediate Biomarkers
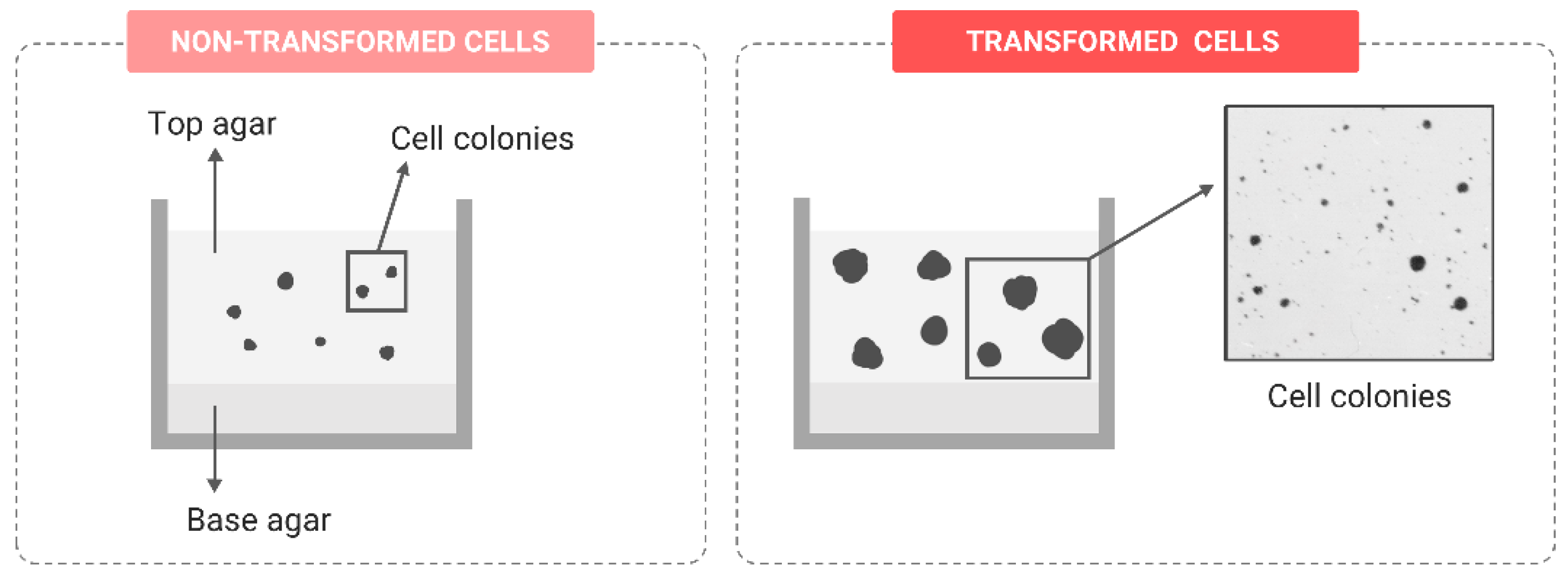
Anchorage-Independent Growth
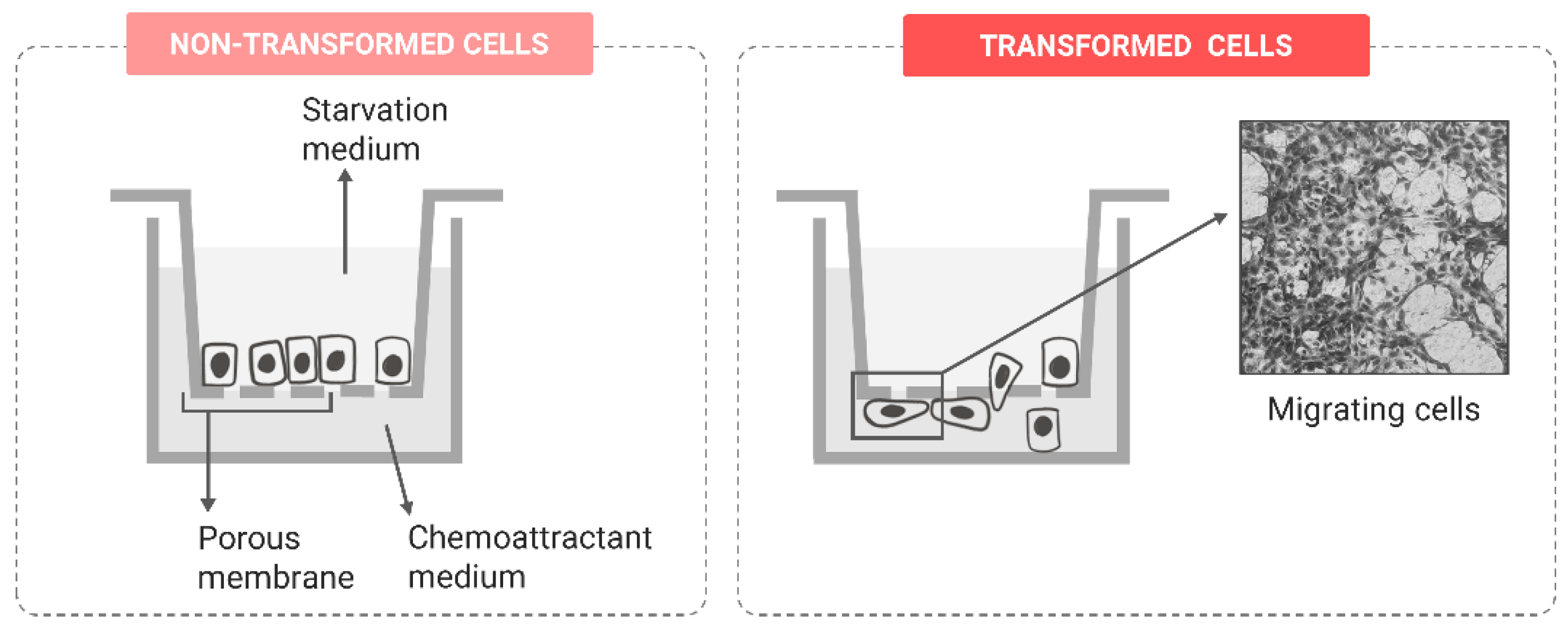
Migration Potential
3.1.3. Advanced Biomarkers
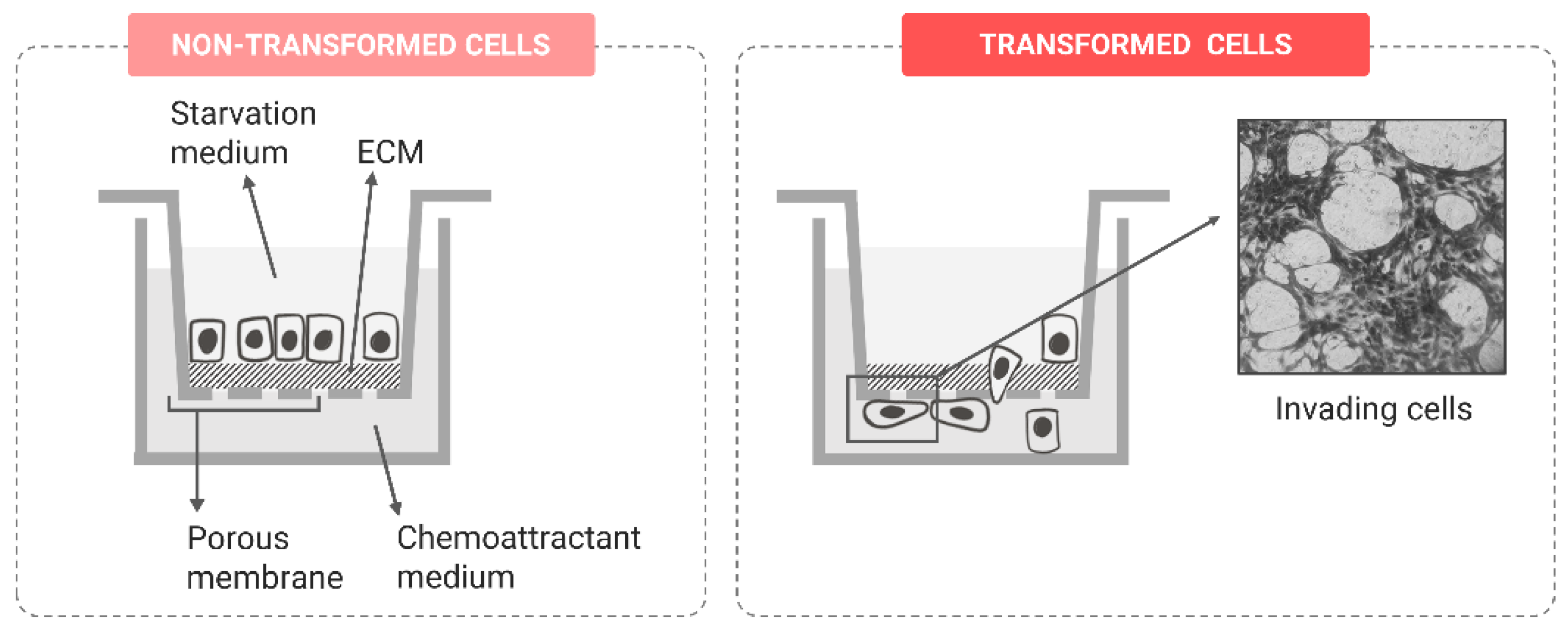
Invasion Potential
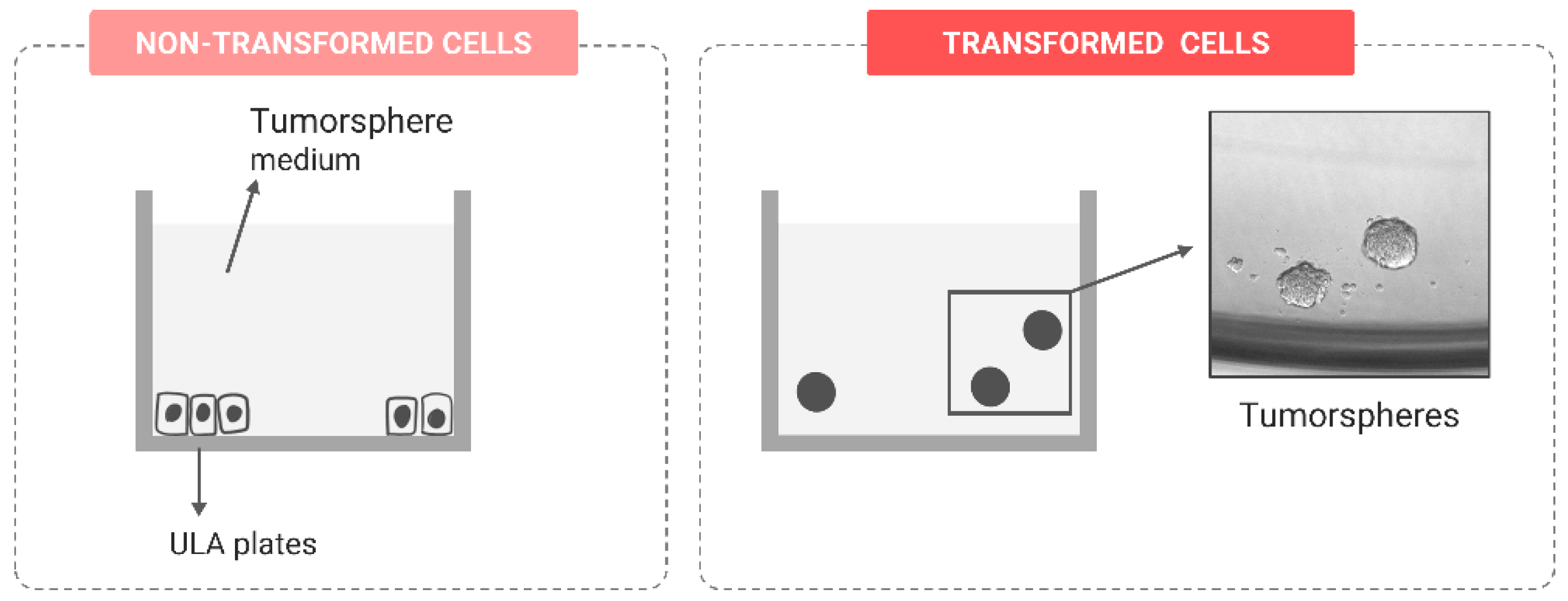
Stem-like Features
4. Representative Studies Using the Proposed Approach to Evaluate In Vitro Carcinogenesis
5. Conclusions and Further Steps
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- EPA/600/R-14/004Next Generation Risk Assessment: Incorporation of Recent Advances in Molecular, Computational, and Systems Biology; EPA: Washington, DC, USA, 2014. Available online: https://cfpub.epa.gov/si/si_public_record_report.cfm?Lab=NCEA&dirEntryId=286690 (accessed on 5 December 2022).
- Rusche, B. The 3Rs and animal welfare—Conflict or the way forward? Altern. Anim. Exp. 2003, 20 (Suppl. S1), 63–76. [Google Scholar]
- Bailey, J.; Thew, M.; Balls, M. Predicting human drug toxicity and safety via animal tests: Can any one species predict drug toxicity in any other, and do monkeys help? Altern. Lab. Anim. 2015, 43, 393–403. [Google Scholar] [CrossRef] [Green Version]
- Andersen, M.E.; Meek, M.E.; Boorman, G.A.; Brusick, D.J.; Cohen, S.M.; Dragan, Y.P.; Frederick, C.B.; Goodman, J.I.; Hard, G.C.; O'Flaherty, E.J.; et al. Lessons learned in applying the U.S. EPA proposed cancer guidelines to specific compounds. Toxicol. Sci. 2000, 53(2), 159–172. [Google Scholar] [CrossRef] [Green Version]
- Boberg, J.; Dybdahl, M.; Petersen, A.; Hass, U.; Svingen, T.; Vinggaard, A.M. A pragmatic approach for human risk assessment of chemical mixtures. Curr. Opin. Toxicol. 2019, 15, 127–152. [Google Scholar] [CrossRef] [Green Version]
- Wambaugh, J.F.; Bare, J.C.; Carignan, C.C.; Dionisio, K.L.; Dodson, R.E.; Jolliet, O.; Liu, X.; Meyer, D.E.; Newton, S.R.; Phillips, K.A.; et al. New approach methodologies for exposure science. Curr. Opin. Toxicol. 2019, 15, 76–92. [Google Scholar] [CrossRef]
- European Chemicals Agency (ECHA). New Approach Methodologies in Regulatory Science: Proceedings of a Scientific Workshop; European Chemicals Agency: Helsinki, Finland, 2016; Available online: https://data.europa.eu/doi/10.2823/543644 (accessed on 5 December 2022).
- Pham, L.L.; Sheffield, T.; Pradeep, P.; Brown, J.; Haggard, D.E.; Wambaugh, J.; Judson, R.; Friedman, K.P. Estimating uncertainty in the context of new approach methodologies for potential use in chemical safety evaluation. Curr. Opin. Toxicol. 2019, 15, 40–47. [Google Scholar] [CrossRef]
- Graepel, R.; Ter Braak, B.; Escher, S.E.; Fisher, C.; Gardner, I.; Kamp, H.; Kroese, D.; Leist, M.; Moné, M.J.; Pastor, M.; et al. Paradigm shift in safety assessment using new approach methods: The EU-ToxRisk strategy. Curr. Opin. Toxicol. 2019, 15, 33–39. [Google Scholar] [CrossRef]
- Kripke, M.; Brody, J.G.; Hawk, E.; Hernandez, A.B.; Hoppin, P.J.; Jacobs, M.M.; Rudel, R.A.; Rebbeck, T.R. Rethinking environmental carcinogenesis. Cancer Epidemiol. Biomark. Prev. 2020, 29, 1870–1875. [Google Scholar] [CrossRef]
- Kumar, P.; Nagarajan, A.; Uchil, P.D. Analysis of cell viability by the alamarBlue assay. Cold Spring Harb. Protoc. 2018, 2018, pdb-prot095489. [Google Scholar] [CrossRef]
- OECD. Test No. 451: Carcinogenicity Studies, OECD Guidelines for the Testing of Chemicals; Section 4; OECD Publishing: Paris, France, 2018. [Google Scholar] [CrossRef]
- Annys, E.; Billington, R.; Clayton, R.; Bremm, K.D.; Graziano, M.; McKelvie, J.; Ragan, I.; Schwarz, M.; van der Laan, J.W.; Wood, C.; et al. Advancing the 3Rs in regulatory toxicology—Carcinogenicity testing: Scope for harmonisation and advancing the 3Rs in regulated sectors of the European Union. Regul. Toxicol. Pharmacol. 2014, 69, 234–242. [Google Scholar] [CrossRef]
- Organization for Economic Co-operation and Development (OECD). Guidance Document on the In Vitro Bhas 42 Cell Transformation Assay; Series on Testing & Assessment No. 231; OECD Environment Directorate: Paris, France; Environment, Health and Safety Division: Paris, France, 2016. [Google Scholar]
- Steinberg, P. In vitro-in vivo carcinogenicity. Adv. Biochem. Eng. Biotechnol. 2017, 157, 81–96. [Google Scholar] [CrossRef] [PubMed]
- Ohmori, K.; Kamei, A.; Watanabe, Y.; Abe, K. Gene expression over time during cell transformation due to non-genotoxic carcinogen treatment of Bhas 42 cells. Int. J. Mol. Sci. 2022, 23, 3216. [Google Scholar] [CrossRef] [PubMed]
- Sasaki, K.; Bohnenberger, S.; Hayashi, K.; Kunkelmann, T.; Muramatsu, D.; Phrakonkham, P.; Poth, A.; Sakai, A.; Salovaara, S.; Tanaka, N.; et al. Recommended protocol for the BALB/c 3T3 cell transformation assay. Mutat. Res. 2012, 744, 30–35. [Google Scholar] [CrossRef] [PubMed]
- Sasaki, K.; Huk, A.; El Yamani, N.; Tanaka, N.; Dusinska, M. Bhas 42 Cell transformation assay for genotoxic and non-genotoxic carcinogens. In Genotoxicity and DNA Repair: A Practical Approach, Methods in Pharmacology and Toxicology; Sierra, M., Gaivão, I., Eds.; Springer Science + Business Media: New York, NY, USA, 2014. [Google Scholar] [CrossRef]
- Maire, M.A.; Pant, K.; Phrakonkham, P.; Poth, A.; Schwind, K.R.; Rast, C.; Bruce, S.W.; Sly, J.E.; Bohnenberger, S.; Kunkelmann, T.; et al. Recommended protocol for the Syrian hamster embryo (SHE) cell transformation assay. Mutat. Res. 2012, 744, 76–81. [Google Scholar] [CrossRef] [PubMed]
- Vasseur, P.; Lasne, C. OECD detailed review paper (DRP) number 31 on “Cell Transformation Assays for Detection of Chemical Carcinogens”: Main results and conclusions. Mutat. Res. 2012, 744, 8–11. [Google Scholar] [CrossRef] [PubMed]
- Creton, S.; Aardema, M.J.; Carmichael, P.L.; Harvey, J.S.; Martin, F.L.; Newbold, R.F.; O’Donovan, M.R.; Pant, K.; Poth, A.; Sakai, A.; et al. Cell transformation assays for prediction of carcinogenic potential: State of the science and future research needs. Mutagenesis 2012, 27, 93–101. [Google Scholar] [CrossRef] [Green Version]
- Knight, A. Systematic reviews of animal experiments demonstrate poor human clinical and toxicological utility. Altern. Lab. Anim. 2007, 35, 641–659. [Google Scholar] [CrossRef] [Green Version]
- Knight, A. Reviewing existing knowledge prior to conducting animal studies. Altern. Lab. Anim. 2008, 36, 709–712. [Google Scholar] [CrossRef] [Green Version]
- Corvi, R.; Aardema, M.J.; Gribaldo, L.; Hayashi, M.; Hoffmann, S.; Schechtman, L.; Vanparys, P. ECVAM prevalidation study on in vitro cell transformation assays: General outline and conclusions of the study. Mutat. Res. 2012, 744, 12–19. [Google Scholar] [CrossRef] [PubMed]
- Mascolo, M.G.; Perdichizzi, S.; Vaccari, M.; Rotondo, F.; Zanzi, C.; Grilli, S.; Paparella, M.; Jacobs, M.N.; Colacci, A. The transformics assay: First steps for the development of an integrated approach to investigate the malignant cell transformation in vitro. Carcinogenesis 2018, 39, 955–967. [Google Scholar] [CrossRef] [Green Version]
- Tokar, E.J.; Diwan, B.A.; Waalkes, M.P. Arsenic exposure transforms human epithelial stem/progenitor cells into a cancer stem-like phenotype. Environ. Health Perspect. 2010, 118, 108–115. [Google Scholar] [CrossRef] [Green Version]
- Wang, L.; Luanpitpong, S.; Castranova, V.; Tse, W.; Lu, Y.; Pongrakhananon, V.; Rojanasakul, Y. Carbon nanotubes induce malignant transformation and tumorigenesis of human lung epithelial cells. Nano Lett. 2011, 11, 2796–2803. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mandriota, S.J.; Tenan, M.; Nicolle, A.; Jankowska, J.D.; Ferrari, P.; Tille, J.C.; Durin, M.A.; Green, C.M.; Tabruyn, S.; Moralli, D.; et al. Genomic instability is an early event in aluminium-induced tumorigenesis. Int. J. Mol. Sci. 2020, 21, 9332. [Google Scholar] [CrossRef] [PubMed]
- Chen, K.Y.; Tseng, C.H.; Feng, P.H.; Sun, W.L.; Ho, S.C.; Lin, C.W.; Van Hiep, N.; Luo, C.S.; Tseng, Y.H.; Chen, T.T.; et al. 3-Nitrobenzanthrone promotes malignant transformation in human lung epithelial cells through the epiregulin-signaling pathway. Cell Biol. Toxicol. 2021, 38, 865–887. [Google Scholar] [CrossRef]
- Breheny, D.; Oke, O.; Faux, S.P. The use of in vitro systems to assess cancer mechanisms and the carcinogenic potential of chemicals. Altern. Lab. Anim. 2011, 39, 233–255. [Google Scholar] [CrossRef]
- Sierra, M.I.; Rubio, L.; Bayón, G.F.; Cobo, I.; Menedez, P.; Morales, P.; Mangas, C.; Urdinguio, R.G.; López, V.; Valdés, A.; et al. DNA methylation changes in human lung epithelial cells exposed to multi-walled carbon nanotubes. Nanotoxicology 2017, 11, 857–870. [Google Scholar] [CrossRef] [PubMed]
- Barguilla, I.; Bach, J.; Peremartí, J.; Marcos, R.; Hernández, A. FRA1 is essential for the maintenance of the oncogenic phenotype induced by in vitro long-term arsenic exposure. Metallomics 2020, 12, 2161–2173. [Google Scholar] [CrossRef] [PubMed]
- Barguilla, I.; Peremartí, J.; Bach, J.; Marcos, R.; Hernández, A. Role of As3mt and Mth1 in the genotoxic and carcinogenic effects induced by long-term exposures to arsenic in MEF cells. Toxicol. Appl. Pharmacol. 2020, 409, 115303. [Google Scholar] [CrossRef] [PubMed]
- Hanahan, D.; Weinberg, R.A. The hallmarks of cancer. Cell 2000, 100, 57–70. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hanahan, D.; Weinberg, R.A. Hallmarks of cancer: The next generation. Cell 2011, 144, 646–674. [Google Scholar] [CrossRef] [Green Version]
- Hanahan, D. Hallmarks of cancer: New dimensions. Cancer Discov. 2022, 12, 31–46. [Google Scholar] [CrossRef]
- Bailey, M.H.; Tokheim, C.; Porta-Pardo, E.; Sengupta, S.; Bertrand, D.; Weerasinghe, A.; Colaprico, A.; Wendl, M.C.; Kim, J.; Reardon, B.; et al. Comprehensive characterization of cancer driver genes and mutations. Cell 2018, 173, 371–385.e18. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sonugür, F.G.; Akbulut, H. The role of tumor microenvironment in genomic instability of malignant tumors. Front. Genet. 2019, 10, 1063. [Google Scholar] [CrossRef]
- Phillips, D.H.; Arlt, V.M. Genotoxicity: Damage to DNA and its consequences. Exp. Suppl. 2009, 99, 87–110. [Google Scholar] [CrossRef] [Green Version]
- OECD. Test No. 489: In Vivo Mammalian Alkaline Comet Assay, OECD Guidelines for the Testing of Chemicals; Section 4; OECD Publishing: Paris, France, 2016. [Google Scholar] [CrossRef]
- OECD. Test No. 487: In Vitro Mammalian Cell Micronucleus Test, OECD Guidelines for the Testing of Chemicals; Section 4; OECD Publishing: Paris, France, 2016. [Google Scholar] [CrossRef]
- Olive, P.L.; Banáth, J.P. The comet assay: A method to measure DNA damage in individual cells. Nat. Protoc. 2006, 1, 23–29. [Google Scholar] [CrossRef] [PubMed]
- Collins, A.R. Measuring oxidative damage to DNA and its repair with the comet assay. Biochim. Biophys. Acta 2014, 1840, 794–800. [Google Scholar] [CrossRef] [PubMed]
- Azqueta, A.; Gutzkow, K.B.; Priestley, C.C.; Meier, S.; Walker, J.S.; Brunborg, G.; Collins, A.R. A comparative performance test of standard, medium- and high-throughput comet assays. Toxicol. Vitr. 2013, 27, 768–773. [Google Scholar] [CrossRef] [PubMed]
- Sykora, P.; Witt, K.L.; Revanna, P.; Smith-Roe, S.L.; Dismukes, J.; Lloyd, D.G.; Engelward, B.P.; Sobol, R.W. Next generation high throughput DNA damage detection platform for genotoxic compound screening. Sci. Rep. 2018, 8, 2771. [Google Scholar] [CrossRef] [Green Version]
- Fenech, M. Cytokinesis-block micronucleus cytome assay. Nat. Protoc. 2007, 2, 1084–1104. [Google Scholar] [CrossRef] [Green Version]
- Avlasevich, S.; Bryce, S.; De Boeck, M.; Elhajouji, A.; Van Goethem, F.; Lynch, A.; Nicolette, J.; Shi, J.; Dertinger, S. Flow cytometric analysis of micronuclei in mammalian cell cultures: Past, present and future. Mutagenesis 2011, 26, 147–152. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rodrigues, M.A.; Probst, C.E.; Zayats, A.; Davidson, B.; Riedel, M.; Li, Y.; Venkatachalam, V. The in vitro micronucleus assay using imaging flow cytometry and deep learning. NPJ Syst. Biol. Appl. 2021, 7, 20. [Google Scholar] [CrossRef]
- Evan, G.I.; Vousden, K.H. Proliferation, cell cycle and apoptosis in cancer. Nature 2001, 411, 342–348. [Google Scholar] [CrossRef] [PubMed]
- Darwiche, N. Epigenetic mechanisms and the hallmarks of cancer: An intimate affair. Am. J. Cancer Res. 2020, 10, 1954–1978. [Google Scholar] [PubMed]
- Mohammad, R.M.; Muqbil, I.; Lowe, L.; Yedjou, C.; Hsu, H.Y.; Lin, L.T.; Siegelin, M.D.; Fimognari, C.; Kumar, N.B.; Dou, Q.P.; et al. Broad targeting of resistance to apoptosis in cancer. Semin. Cancer Biol. 2015, 27, S78–S103. [Google Scholar] [CrossRef]
- Adan, A.; Kiraz, Y.; Baran, Y. Cell proliferation and cytotoxicity assays. Curr. Pharm. Biotechnol. 2016, 17, 1213–1221. [Google Scholar] [CrossRef]
- Präbst, K.; Engelhardt, H.; Ringgeler, S.; Hübner, H. Basic colorimetric proliferation assays: MTT, WST, and Resazurin. In Cell Viability Assays: Methods in Molecular Biology; Gilbert, D.F., Friedich, O., Eds.; Humana Press: New York, NY, USA; Volume 1601. [CrossRef]
- ATCC. Animal Cell Culture Guide. 2021. Available online: https://www.atcc.org/resources/culture-guides/animal-cell-culture-guide#Cell (accessed on 5 December 2022).
- Berridge, M.V.; Herst, P.M.; Tan, A.S. Tetrazolium dyes as tools in cell biology: New insights into their cellular reduction. Biotechnol. Annu. Rev. 2005, 11, 127–152. [Google Scholar] [CrossRef]
- Kumar, V.; Boobis, A.R.; Moretto, A. Test and risk assessment strategies for combined exposure to multiple chemicals. Food Chem. Toxicol. 2020, 144, 111607. [Google Scholar] [CrossRef]
- Vichai, V.; Kirtikara, K. Sulforhodamine B colorimetric assay for cytotoxicity screening. Nat. Protoc. 2006, 1, 1112–1116. [Google Scholar] [CrossRef]
- Ormerod, M.G. Analysis of cell proliferation using the bromodeoxyuridine/Hoechst-ethidium bromide method. In Basic Cell Culture Protocols: Methods in Molecular Biology; Pollard, J.W., Walker, J.M., Eds.; Humana Press: Totowa, NJ, USA, 1997; Volume 75, pp. 357–365. [Google Scholar] [CrossRef]
- Salic, A.; Mitchison, T.J. A chemical method for fast and sensitive detection of DNA synthesis in vivo. Proc. Natl. Acad. Sci. USA 2008, 105, 2415–2420. [Google Scholar] [CrossRef] [Green Version]
- Lanigan, T.M.; Rasmussen, S.M.; Weber, D.P.; Athukorala, K.S.; Campbell, P.L.; Fox, D.A.; Ruth, J.H. Real time visualization of cancer cell death, survival and proliferation using fluorochrome-transfected cells in an IncuCyte® imaging system. J. Biol. Methods 2020, 7, e133. [Google Scholar] [CrossRef] [PubMed]
- Keskinoğlu, C.; Aydın, A. Ultrasound based noninvasive real-time cell proliferation process monitoring. J. Acoust. Soc. Am. 2021, 149, 3345–3351. [Google Scholar] [CrossRef]
- Gonzalez, D.M.; Medici, D. Signaling mechanisms of the epithelial-mesenchymal transition. Sci. Signal. 2014, 7, re8. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yang, J.; Weinberg, R.A. Epithelial-mesenchymal transition: At the crossroads of development and tumor metastasis. Dev. Cell 2008, 14, 818–829. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sahai, E.; Astsaturov, I.; Cukierman, E.; De Nardo, D.G.; Egeblad, M.; Evans, R.M.; Fearon, D.; Greten, F.R.; Hingorani, S.R.; Hunter, T.; et al. A framework for advancing our understanding of cancer-associated fibroblasts. Nat. Rev. Cancer 2020, 20, 174–186. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ren, Z.X.; Yu, H.B.; Li, J.S.; Shen, J.L.; Du, W.S. Suitable parameter choice on quantitative morphology of A549 cell in epithelial-mesenchymal transition. Biosci. Rep. 2015, 35, e00202. [Google Scholar] [CrossRef]
- Leggett, S.E.; Sim, J.Y.; Rubins, J.E.; Neronha, Z.J.; Williams, E.K.; Wong, I.Y. Morphological single cell profiling of the epithelial-mesenchymal transition. Integr. Biol. 2016, 8, 1133–1144. [Google Scholar] [CrossRef] [Green Version]
- Alizadeh, E.; Castle, J.; Quirk, A.; Taylor, C.D.L.; Xu, W.; Prasad, A. Cellular morphological features are predictive markers of cancer cell state. Comput. Biol. Med. 2020, 126, 104044. [Google Scholar] [CrossRef]
- Arismendi-Morillo, G. Electron microscopy morphology of the mitochondrial network in human cancer. Int. J. Biochem. Cell Biol. 2009, 41, 2062–2068. [Google Scholar] [CrossRef]
- Badrinath, N.; Yoo, S.Y. Mitochondria in cancer: In the aspects of tumorigenesis and targeted therapy. Carcinogenesis 2018, 39, 1419–1430. [Google Scholar] [CrossRef]
- Faitg, J.; Davey, T.; Turnbull, D.M.; White, K.; Vincent, A.E. Mitochondrial morphology and function: Two for the price of one! J. Microsc. 2020, 278, 89–106. [Google Scholar] [CrossRef] [Green Version]
- Liu, H.; Wang, N.; Zhang, Z.; Wang, H.; Du, J.; Tang, J. Effects of tumor necrosis factor-α on morphology and mechanical properties of HCT116 human colon cancer cells investigated by atomic force microscopy. Scanning 2017, 2017, 2027079. [Google Scholar] [CrossRef] [Green Version]
- Lam, J.; Katti, P.; Biete, M.; Mungai, M.; AshShareef, S.; Neikirk, K.; Garza Lopez, E.; Vue, Z.; Christensen, T.A.; Beasley, H.K.; et al. A universal approach to analyzing transmission electron microscopy with ImageJ. Cells 2021, 10, 2177. [Google Scholar] [CrossRef]
- Anderson, N.M.; Simon, M.C. The tumor microenvironment. Curr. Biol. 2020, 30, R921–R925. [Google Scholar] [CrossRef]
- Bussard, K.M.; Mutkus, L.; Stumpf, K.; Gomez-Manzano, C.; Marini, F.C. Tumor-associated stromal cells as key contributors to the tumor microenvironment. Breast Cancer Res. 2016, 18, 84. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ritchie, S.; Reed, D.A.; Pereira, B.A.; Timpson, P. The cancer cell secretome drives cooperative manipulation of the tumour microenvironment to accelerate tumourigenesis. Fac. Rev. 2021, 10, 4. [Google Scholar] [CrossRef]
- Guyot, B.; Lefort, S.; Voeltzel, T.; Pécheur, E.I.; Maguer-Satta, V. Altered BMP2/4 signaling in stem cells and their niche: Different cancers but similar mechanisms, the example of myeloid leukemia and breast cancer. Front. Cell Dev. Biol. 2022, 9, 787989. [Google Scholar] [CrossRef] [PubMed]
- Habanjar, O.; Diab-Assaf, M.; Caldefie-Chezet, F.; Delort, L. 3D cell culture systems: Tumor application, advantages, and disadvantages. Int. J. Mol. Sci. 2021, 22, 12200. [Google Scholar] [CrossRef] [PubMed]
- Borowicz, S.; Van Scoyk, M.; Avasarala, S.; Karuppusamy Rathinam, M.K.; Tauler, J.; Bikkavilli, R.K.; Winn, R.A. The soft agar colony formation assay. J. Vis. Exp. 2014, 92, e51998. [Google Scholar] [CrossRef] [Green Version]
- Bach, J.; Peremartí, J.; Annangi, B.; Marcos, R.; Hernández, A. Oxidative DNA damage enhances the carcinogenic potential of in vitro chronic arsenic exposures. Arch. Toxicol. 2016, 90, 1893–1905. [Google Scholar] [CrossRef]
- Landskron, G.; De la Fuente, M.; Thuwajit, P.; Thuwajit, C.; Hermoso, M.A. Chronic inflammation and cytokines in the tumor microenvironment. J. Immunol. Res. 2014, 2014, 149185. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zamarron, B.F.; Chen, W. Dual roles of immune cells and their factors in cancer development and progression. Int. J. Biol. Sci. 2011, 7, 651–658. [Google Scholar] [CrossRef] [PubMed]
- Najafi, M.; Farhood, B.; Mortezaee, K. Extracellular matrix (ECM) stiffness and degradation as cancer drivers. J. Cell. Biochem. 2019, 120, 2782–2790. [Google Scholar] [CrossRef]
- Cox, T.R. The matrix in cancer. Nat. Rev. Cancer 2021, 21, 217–238. [Google Scholar] [CrossRef]
- Tajhya, R.B.; Patel, R.S.; Beeton, C. Detection of matrix metalloproteinases by zymography. Methods Mol. Biol. 2017, 1579, 231–244. [Google Scholar] [CrossRef] [Green Version]
- Inanc, S.; Keles, D.; Oktay, G. An improved collagen zymography approach for evaluating the collagenases MMP-1, MMP-8, and MMP-13. Biotechniques 2017, 63, 174–180. [Google Scholar] [CrossRef] [Green Version]
- Janiszewska, M.; Primi, M.C.; Izard, T. Cell adhesion in cancer: Beyond the migration of single cells. J. Biol. Chem. 2020, 295, 2495–2505. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fang, M.; Yuan, J.; Peng, C.; Li, Y. Collagen as a double-edged sword in tumor progression. Tumour Biol. 2014, 35, 2871–2882. [Google Scholar] [CrossRef] [Green Version]
- Du, F.; Zhao, X.; Fan, D. Soft agar colony formation assay as a hallmark of carcinogenesis. Bio-Protocol 2017, 7, e2351. [Google Scholar] [CrossRef]
- Huang, Y.; Wang, Q.; Tang, Y.; Liu, Z.; Sun, G.; Lu, Z.; Chen, Y. Identification and validation of a cigarette smoke-related five-gene signature as a prognostic biomarker in kidney renal clear cell carcinoma. Sci. Rep. 2022, 12, 2189. [Google Scholar] [CrossRef]
- Michailidi, C.; Hayashi, M.; Datta, S.; Sen, T.; Zenner, K.; Oladeru, O.; Brait, M.; Izumchenko, E.; Baras, A.; VandenBussche, C.; et al. Involvement of epigenetics and EMT-related miRNA in arsenic-induced neoplastic transformation and their potential clinical use. Cancer Prev. Res. 2015, 8, 208–221. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Barguilla, I.; Barszczewska, G.; Annangi, B.; Domenech, J.; Velázquez, A.; Marcos, R.; Hernández, A. MTH1 is involved in the toxic and carcinogenic long-term effects induced by zinc oxide and cobalt nanoparticles. Arch. Toxicol. 2020, 94, 1073–1984. [Google Scholar] [CrossRef]
- Martin, T.A.; Ye, L.; Sanders, A.J.; Lane, J.; Jiang, W.G. Cancer invasion and metastasis: Molecular and cellular perspective. In Madame Curie Bioscience Database; Landes Bioscience: Austin, TX, USA, 2013. Available online: https://www.ncbi.nlm.nih.gov/books/NBK164700 (accessed on 5 December 2022).
- Friedl, P.; Wolf, K. Tumour-cell invasion and migration: Diversity and escape mechanisms. Nat. Rev. Cancer 2003, 3, 362–374. [Google Scholar] [CrossRef] [PubMed]
- Kramer, N.; Walzl, A.; Unger, C.; Rosner, M.; Krupitza, G.; Hengstschläger, M.; Dolznig, H. In vitro cell migration and invasion assays. Mutat. Res. 2013, 752, 10–24. [Google Scholar] [CrossRef]
- Pijuan, J.; Barceló, C.; Moreno, D.F.; Maiques, O.; Sisó, P.; Marti, R.M.; Macià, A.; Panosa, A. In vitro cell migration, invasion, and adhesion assays: From cell imaging to data analysis. Front. Cell Dev. Biol. 2019, 7, 107. [Google Scholar] [CrossRef] [Green Version]
- Liang, C.C.; Park, A.Y.; Guan, J.L. In vitro scratch assay: A convenient and inexpensive method for analysis of cell migration in vitro. Nat. Protoc. 2007, 2, 329–333. [Google Scholar] [CrossRef] [Green Version]
- Justus, C.R.; Leffler, N.; Ruiz-Echevarria, M.; Yang, L.V. In vitro cell migration and invasion assays. J. Vis. Exp. 2014, 88, e51046. [Google Scholar] [CrossRef] [Green Version]
- Liarte, S.; Bernabé-García, Á.; Armero-Barranco, D.; Nicolás, F.J. Microscopy based methods for the assessment of epithelial cell migration during in vitro wound healing. J. Vis. Exp. 2018, 131, e56799. [Google Scholar] [CrossRef]
- Cappiello, F.; Casciaro, B.; Mangoni, M.L. A novel in vitro wound healing assay to evaluate cell migration. J. Vis. Exp. 2018, 133, e56825. [Google Scholar] [CrossRef]
- Vang Mouritzen, M.; Jenssen, H. Optimized scratch assay for in vitro testing of cell migration with an automated optical camera. J. Vis. Exp. 2018, 138, e57691. [Google Scholar] [CrossRef] [Green Version]
- Guy, J.B.; Espenel, S.; Vallard, A.; Battiston-Montagne, P.; Wozny, A.S.; Ardail, D.; Alphonse, G.; Rancoule, C.; Rodriguez-Lafrasse, C.; Magne, N. Evaluation of the cell invasion and migration process: A comparison of the video microscope-based scratch wound assay and the Boyden chamber assay. J. Vis. Exp. 2017, 129, e56337. [Google Scholar] [CrossRef]
- Mudduluru, G.; Large, N.; Park, T. Impedance-based real-time measurement of cancer cell migration and invasion. J. Vis. Exp. 2020, 158, e60997. [Google Scholar] [CrossRef]
- Gau, D.M.; Roy, P. Single cell migration assay using human breast cancer MDA-MB-231 cell line. Bio Protoc. 2020, 10, e3586. [Google Scholar] [CrossRef] [PubMed]
- Falasca, M.; Raimondi, C.; Maffucci, T. Boyden chamber. Wells, C., Parsons, M., Eds.; In Cell Migration. Methods in Molecular Biology; Humana Press: Totowa, NJ, USA, 2011; Volume 769, pp. 87–95. [Google Scholar] [CrossRef]
- Marshall, J. Transwell invasion assays. In Cell Migration. Methods in Molecular Biology; Wells, C., Parsons, M., Eds.; Humana Press: Totowa, NJ, USA, 2011; Volume 769, pp. 97–110. [Google Scholar] [CrossRef]
- Vinci, M.; Box, C.; Eccles, S.A. Three-dimensional (3D) tumor spheroid invasion assay. J. Vis. Exp. 2015, 99, e52686. [Google Scholar] [CrossRef] [Green Version]
- Naakka, E.; Tuomainen, K.; Wistrand, H.; Palkama, M.; Suleymanova, I.; Al-Samadi, A.; Salo, T. Fully human tumor-based matrix in three-dimensional spheroid invasion assay. J. Vis. Exp. 2019, 147, e59567. [Google Scholar] [CrossRef] [Green Version]
- Conti, S.; Kato, T.; Park, D.; Sahai, E.; Trepat, X.; Labernadie, A. CAFs and cancer cells co-migration in 3D spheroid invasion assay. In The Epithelial-to Mesenchymal Transition: Methods in Molecular Biology; Campbell, K., Theveneau, E., Eds.; Humana: New York, NY, USA, 2021; Volume 2179, pp. 242–256. [Google Scholar] [CrossRef]
- López de Andrés, J.; Griñán-Lisón, C.; Jiménez, G.; Marchal, J.A. Cancer stem cell secretome in the tumor microenvironment: A key point for an effective personalized cancer treatment. J. Hematol. Oncol. 2020, 13, 136. [Google Scholar] [CrossRef]
- Tanabe, A.; Sahara, H. The metabolic heterogeneity and flexibility of cancer stem cells. Cancers 2020, 12, 2780. [Google Scholar] [CrossRef] [PubMed]
- Johnson, S.; Chen, H.; Lo, P.K. In vitro tumorsphere formation assays. Bio-Protocol 2013, 3, e325. [Google Scholar] [CrossRef] [Green Version]
- Balla, M.M.S.; Yadav, H.D.; Pandey, B.N. Tumorsphere assay provides a better in vitro method for cancer stem-like cells enrichment in A549 lung adenocarcinoma cells. Tissue Cell 2019, 60, 21–24. [Google Scholar] [CrossRef]
- Lee, C.H.; Yu, C.C.; Wang, B.Y.; Chang, W.W. Tumorsphere as an effective in vitro platform for screening anti-cancer stem cell drugs. Oncotarget 2016, 7, 1215–1226. [Google Scholar] [CrossRef] [Green Version]
- Barguilla, I.; Domenech, J.; Ballesteros, S.; Rubio, L.; Marcos, R.; Hernández, A. Long-term exposure to nanoplastics alters molecular and functional traits related to the carcinogenic process. J. Hazard Mater. 2022, 438, 129470. [Google Scholar] [CrossRef] [PubMed]
- Carpenter, R.L.; Jiang, Y.; Jing, Y.; He, J.; Rojanasakul, Y.; Liu, L.Z.; Jiang, B.H. Arsenite induces cell transformation by reactive oxygen species, AKT, ERK1/2, and p70S6K1. Biochem. Biophys. Res. Commun. 2011, 414, 533–538. [Google Scholar] [CrossRef]
- Weinmuellner, R.; Kryeziu, K.; Zbiral, B.; Tav, K.; Schoenhacker-Alte, B.; Groza, D.; Wimmer, L.; Schosserer, M.; Nagelreiter, F.; Rösinger, S.; et al. Long-term exposure of immortalized keratinocytes to arsenic induces EMT, impairs differentiation in organotypic skin models and mimics aspects of human skin derangements. Arch. Toxicol. 2018, 92, 181–194. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ghosh, A.; Mukherjee, S.; Roy, M. Black tea extract prevents inorganic arsenic induced uncontrolled proliferation, epithelial to mesenchymal transition and induction of metastatic properties in HaCaT keratinocytes - an in vitro study. Toxicol. Vitr. 2022, 85, 105478. [Google Scholar] [CrossRef] [PubMed]
- Sappino, A.P.; Buser, R.; Lesne, L.; Gimelli, S.; Béna, F.; Belin, D.; Mandriota, S.J. Aluminium chloride promotes anchorage-independent growth in human mammary epithelial cells. J. Appl. Toxicol. 2012, 32, 233–243. [Google Scholar] [CrossRef] [Green Version]
- Zou, E.; Matsumura, F. Long-term exposure to beta-hexachlorocyclohexane (beta-HCH) promotes transformation and invasiveness of MCF-7 human breast cancer cells. Biochem. Pharmacol. 2003, 66, 831–840. [Google Scholar] [CrossRef]
- Fararjeh, A.S.; Tu, S.H.; Chen, L.C.; Cheng, T.C.; Liu, Y.R.; Chang, H.L.; Chang, H.W.; Huang, C.C.; Wang, H.R.; Hwang-Verslues, W.W.; et al. Long-term exposure to extremely low-dose of nicotine and 4-(methylnitrosamino)-1-(3-pyridyl)-1-butanone (NNK) induce non-malignant breast epithelial cell transformation through activation of the a9-nicotinic acetylcholine receptor-mediated signaling pathway. Environ. Toxicol. 2019, 34, 73–82. [Google Scholar] [CrossRef] [Green Version]
- Lin, H.D.; Tseng, Y.K.; Yuh, C.H.; Chen, S.C. Low concentrations of 4-ABP promote liver carcinogenesis in human liver cells and a zebrafish model. J. Hazard. Mater. 2022, 423 Pt A, 126954. [Google Scholar] [CrossRef]
- Zhang, Y.; Huang, L.; Zhao, Y.; Hu, T. Musk xylene induces malignant transformation of human liver cell line L02 via repressing the TGF-β signaling pathway. Chemosphere 2017, 168, 1506–1514. [Google Scholar] [CrossRef]
- Clément, F.; Xu, X.; Donini, C.F.; Clément, A.; Omarjee, S.; Delay, E.; Treilleux, I.; Fervers, B.; Le Romancer, M.; Cohen, P.A.; et al. Long-term exposure to bisphenol A or benzo(a)pyrene alters the fate of human mammary epithelial stem cells in response to BMP2 and BMP4, by pre-activating BMP signaling. Cell Death Differ. 2017, 24, 155–166. [Google Scholar] [CrossRef]
- Chapellier, M.; Bachelard-Cascales, E.; Schmidt, X.; Clément, F.; Treilleux, I.; Delay, E.; Jammot, A.; Ménétrier-Caux, C.; Pochon, G.; Besançon, R.; et al. Disequilibrium of BMP2 levels in the breast stem cell niche launches epithelial transformation by overamplifying BMPR1B cell response. Stem Cell Rep. 2015, 4, 239–254. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jung, N.; Maguer-Satta, V.; Guyot, B. Early steps of mammary stem cell transformation by exogenous signals; effects of bisphenol endocrine disrupting chemicals and bone morphogenetic proteins. Cancers 2019, 11, 1351. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Annangi, B.; Bach, J.; Vales, G.; Rubio, L.; Marcos, R.; Hernández, A. Long-term exposures to low doses of cobalt nanoparticles induce cell transformation enhanced by oxidative damage. Nanotoxicology 2015, 9, 138–147. [Google Scholar] [CrossRef]
- Annangi, B.; Rubio, L.; Alaraby, M.; Bach, J.; Marcos, R.; Hernández, A. Acute and long-term in vitro effects of zinc oxide nanoparticles. Arch. Toxicol. 2016, 90, 2201–2213. [Google Scholar] [CrossRef] [PubMed]
- Meng, J.; Zhou, X.; Yang, J.; Qu, X.; Cui, S. Exposure to low dose ZnO nanoparticles induces hyperproliferation and malignant transformation through activating the CXCR2/NF-κB/STAT3/ERK and AKT pathways in colonic mucosal cells. Environ. Pollut. 2020, 263, 114578. [Google Scholar] [CrossRef]
- Rubio, L.; Bach, J.; Marcos, R.; Hernández, A. Synergistic role of nanoceria on the ability of tobacco smoke to induce carcinogenic hallmarks in lung epithelial cells. Nanomedicine 2017, 12, 2623–2635. [Google Scholar] [CrossRef]
- Ballesteros, S.; Barguilla, I.; Marcos, R.; Hernández, A. Nanoceria, alone or combined with cigarette-smoke condensate, induce transforming and epigenetic cancer-like features. Nanomedicine 2021, 16, 293–305. [Google Scholar] [CrossRef]
- Gliga, A.R.; Di Bucchianico, S.; Åkerlund, E.; Karlsson, H.L. Transcriptome profiling and toxicity following long-term, low dose exposure of human lung cells to Ni and NiO nanoparticles-comparison with NiCl2. Nanomaterials 2020, 10, 649. [Google Scholar] [CrossRef] [Green Version]
- Mo, Y.; Zhang, Y.; Zhang, Y.; Yuan, J.; Mo, L.; Zhang, Q. Nickel nanoparticle-induced cell transformation: Involvement of DNA damage and DNA repair defect through HIF-1α/miR-210/Rad52 pathway. J. Nanobiotechnol. 2021, 19, 370. [Google Scholar] [CrossRef]
- Kornberg, T.G.; Stueckle, T.A.; Coyle, J.; Derk, R.; Demokritou, P.; Rojanasakul, Y.; Rojanasakul, L.W. Iron oxide nanoparticle-induced neoplastic-like cell transformation in vitro is reduced with a protective amorphous silica coating. Chem. Res. Toxicol. 2019, 32, 2382–2397. [Google Scholar] [CrossRef]
- Domenech, J.; de Britto, M.; Velázquez, A.; Pastor, S.; Hernández, A.; Marcos, R.; Cortés, C. Long-term effects of polystyrene nanoplastics in human intestinal Caco-2 cells. Biomolecules 2021, 11, 1442. [Google Scholar] [CrossRef] [PubMed]
- Gopinath, P.M.; Twayana, K.S.; Ravanan, P.; John Thomas Mukherjee, A.; Jenkins, D.F.; Chandrasekaran, N. Prospects on the nano-plastic particles internalization and induction of cellular response in human keratinocytes. Part. Fibre. Toxicol. 2021, 18, 35. [Google Scholar] [CrossRef] [PubMed]
- Visalli, G.; Facciolà, A.; Pruiti Ciarello, M.; De Marco, G.; Maisano, M.; Di Pietro, A. Acute and sub-chronic effects of microplastics (3 and 10 µm) on the human intestinal cells HT-29. Int. J. Environ. Res. Public Health 2021, 18, 5833. [Google Scholar] [CrossRef]
- Barguilla, I.; Domenech, J.; Rubio, L.; Marcos, R.; Hernández, A. Nanoplastics and arsenic co-exposures exacerbate oncogenic biomarkers under an in vitro long-term exposure scenario. Int. J. Mol. Sci. 2022, 23, 2958. [Google Scholar] [CrossRef] [PubMed]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Biomarkers of In Vitro Transformation | Assays | Protocols | |
|---|---|---|---|
| Early biomarkers | DNA damage and genotoxicity | Comet assay | [42,43,44,45] |
| Micronucleus assay | [46,47,48] | ||
| Uncontrolled proliferation | Population-doubling time | [54] | |
| Metabolic assays | [55,56,57] | ||
| DNA synthesis and quantification assays | [58,59] | ||
| Real-time monitoring of cell proliferation | [60,61] | ||
| Morphological changes | Microscopic imaging | [65,66,67] | |
| Ultrastructural imaging (TEM and SEM) | [71,72] | ||
| Secretome alterations | Indirect soft-agar assay | [78,79] | |
| Cytokine arrays | [114] | ||
| Zymogram | [84] | ||
| Intermediate biomarkers | Anchorage-independent growth | Soft-agar assay | [78,88] |
| Migration potential | Wound-healing assay | [96,97,98,99,100,101] | |
| Boyden chamber assay | [97] | ||
| Impedance-based real-time migration assay | [102] | ||
| Single-cell tracking | [103] | ||
| Advanced biomarkers | Invasion potential | Boyden chamber assay | [97,104,105] |
| Impedance-based real-time migration assay | [102] | ||
| Single-cell tracking | [103] | ||
| Spheroid invasion assay | [106,107,108] | ||
| Stem-like feature acquisition | Tumorsphere formation assay | [111,112] | |
| Reference | Agent | Cell Line | Time of Exposure | Concentration | Early Biomarkers | Intermediate Biomarkers | Advanced Biomarkers | Transformed Phenotype | |||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| DNA Damage and Genotoxicity | Uncontrolled Proliferation | Morphological Changes | Secretome Alterations | Anchorage-Independent Growth | Migration Potential | Invasion Potential | Sem-Like Features Acquisition | ||||||
| [126] | CoNPs | DNA damage-sensitive MEFs | 12 weeks (3 months) | 0.05 μg/mL | + | + | + | Yes | |||||
| 0.1 μg/mL | + | + | + | Yes | |||||||||
| [127] | ZnONPs | DNA damage-sensitive MEFs | 12 weeks (3 months) | 1 μg/mL | - | - | - | - | No | ||||
| ZnCl2 | - | - | - | - | No | ||||||||
| [79] | As | MEFs | 40 weeks (9 months) | 0.5–2 μM | - | - | + | + | Mild | ||||
| DNA damage-sensitive MEFs | + | + | + | + | Yes | ||||||||
| [130] | CeO2NPs | BEAS-2B | 6 weeks (1.5 months) | 2.5 μg/mL CeO2 5 μg/mL CSC | + | + | Yes | ||||||
| CSC | + | + | Yes | ||||||||||
| CeO2NPs + CSC | + | + | Yes | ||||||||||
| [137] | PSNPLs | Prone-to-transformation progress MEFs | 12 weeks (3 months) | 25 μg/mL PSNPLs 2 μM As | + | - | + | - | - | - | - | Mild | |
| As | + | - | + | - | - | - | - | Mild | |||||
| PSNPLs + As | + | - | + | + | + | + | - | Enhanced | |||||
| [114] | PSNPLs | Prone-to-transformation progress MEFs | 24 weeks (6 months) | 25 μg/mL | + | + | + | + | Yes | ||||
| [115] | As | BEAS-2B | 26 weeks (6.5 months) | 0.25 μM | + | + | Yes | ||||||
| 1 μM | + | + | Yes | ||||||||||
| 5 μM | + | + | Yes | ||||||||||
| [123] | BPA | MCF-10A | 8.5 weeks (2 months) | 10−10 M BPA 10−10 M B(a)P | +* | + | Enhanced | ||||||
| B(a)P | +* | + | Enhanced | ||||||||||
| [120] | Nic | HBL-100 | 9 weeks (2 months) | 1 mM Nic 100 femtM NNK | - | + | + | - | + | Yes | |||
| NKK | - | + | + | + | + | Yes | |||||||
| Nic + NKK | + | + | + | + | + | Enhanced | |||||||
| [117] | As | HaCaT | 34 weeks (8 months) | 100 nM As 1 μM BTE | + | + | + | + | + | Yes | |||
| As + BTE | - | - | - | - | - | Prevented | |||||||
| [131] | NiNPs | BEAS-2B | 6 weeks (1.5 months) | 0.5 μg/mL | + | - | + * | - | + * | Mild | |||
| NiONPs | + | - | + * | - | + * | Mild | |||||||
| NiCl2 | + | - | + * | - | + * | Mild | |||||||
| [133] | Fe2O3NPs | BEAS-2B | 28 weeks (6.5 months) | 2.88 μg/mL | + | + | + | Yes | |||||
| SiO2- Fe2O3NPs | - | - | - | No | |||||||||
| GMA-MS | + | + | + | Yes | |||||||||
| [127] | 4-ABP | L-02 | 8 weeks (2 months) | 10 nM | + | + | + | Yes | |||||
| HepG2 | + | + | + | Yes | |||||||||
| [126] | AlCl3 | NMuMG epithelial cells | 16 weeks (4 months) | 100 μM | - | + | + | Yes | |||||
| [128] | ZnONPs | IMCE | 30 passages | 1 μg/mL | + | + | + | + | Yes | ||||
| [132] | NiNPs | BEAS-2B | 21 weeks (5 months) | 0.25 μg/mL | + | + | Yes | ||||||
| 0.5 μg/mL | + | + | Yes | ||||||||||
| [129] | CeO2NPs | BEAS-2B | 6 weeks (1.5 months) | 2.5 μg/mL CeO2 5 μg/mL CSC | - | - | - | - | - | No | |||
| CSC | - | + | + ** | + | + | Yes | |||||||
| CeO2NPs + CSC | + | + | + | + | + | Yes | |||||||
| [118] | AlCl3 | MCF-10A | 9 weeks (2 months) | 10–300 μM | + | + | + | + | Yes | ||||
| [27] | SWCNT | BEAS-2B | 24 weeks (6 months) | 0.02 μg/cm2 | + | + | + | + | + | Yes | |||
| [116] | ATO | Immortalized human keratinocytes | 24 weeks (6 months) | 0.05 μM | + | - | + | Mild | |||||
| 0.1 μM | + | + | + | Mild | |||||||||
| 0.25 μM | + | + | + | Mild | |||||||||
| [122] | Musk xylene | L-02 | 20 passages | 10 μg/L | + | + | + | + | + | Yes | |||
| 100 μg/L | + | + | + | + | + | Yes | |||||||
| 1000 μg/L | + | + | + | + | + | Yes | |||||||
| [119] | E2 | MCF-7 | 56 weeks (13 months) | 1 nM E2 | + | + | + | Yes | |||||
| β-HCH | 100 nM B-HCH | + | + | + | Yes | ||||||||
| 1 μM B-HCH | + | + | + | Yes | |||||||||
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Barguilla, I.; Maguer-Satta, V.; Guyot, B.; Pastor, S.; Marcos, R.; Hernández, A. In Vitro Approaches to Determine the Potential Carcinogenic Risk of Environmental Pollutants. Int. J. Mol. Sci. 2023, 24, 7851. https://doi.org/10.3390/ijms24097851
Barguilla I, Maguer-Satta V, Guyot B, Pastor S, Marcos R, Hernández A. In Vitro Approaches to Determine the Potential Carcinogenic Risk of Environmental Pollutants. International Journal of Molecular Sciences. 2023; 24(9):7851. https://doi.org/10.3390/ijms24097851
Chicago/Turabian StyleBarguilla, Irene, Veronique Maguer-Satta, Boris Guyot, Susana Pastor, Ricard Marcos, and Alba Hernández. 2023. "In Vitro Approaches to Determine the Potential Carcinogenic Risk of Environmental Pollutants" International Journal of Molecular Sciences 24, no. 9: 7851. https://doi.org/10.3390/ijms24097851