
Kinetic and Regulatory Properties of Yarrowia lipolytica Aconitate Hydratase as a Model-Indicator of Cell Redox State under pH Stress
 , , and
, , and
Abstract
:1. Introduction
2. Results
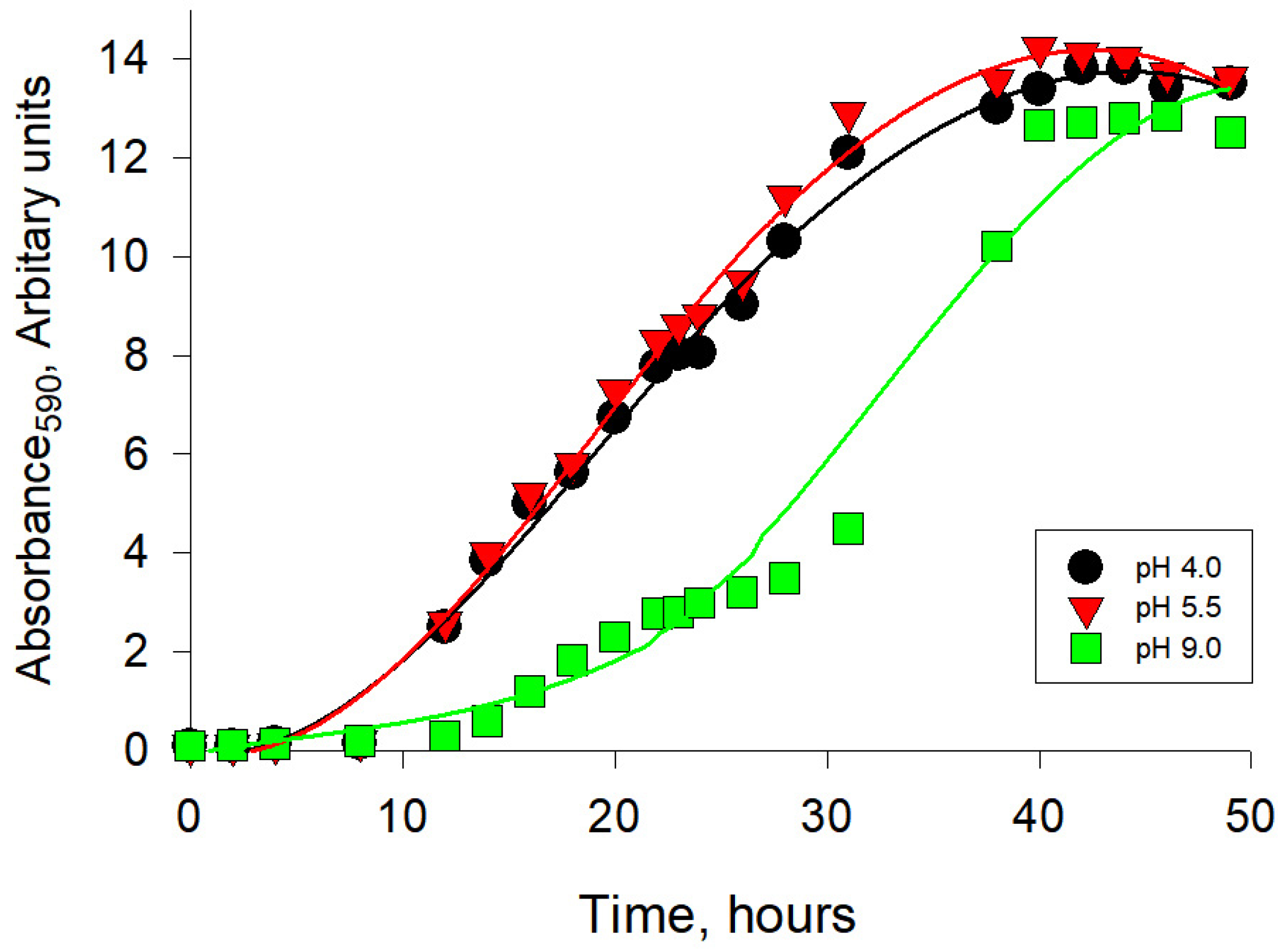
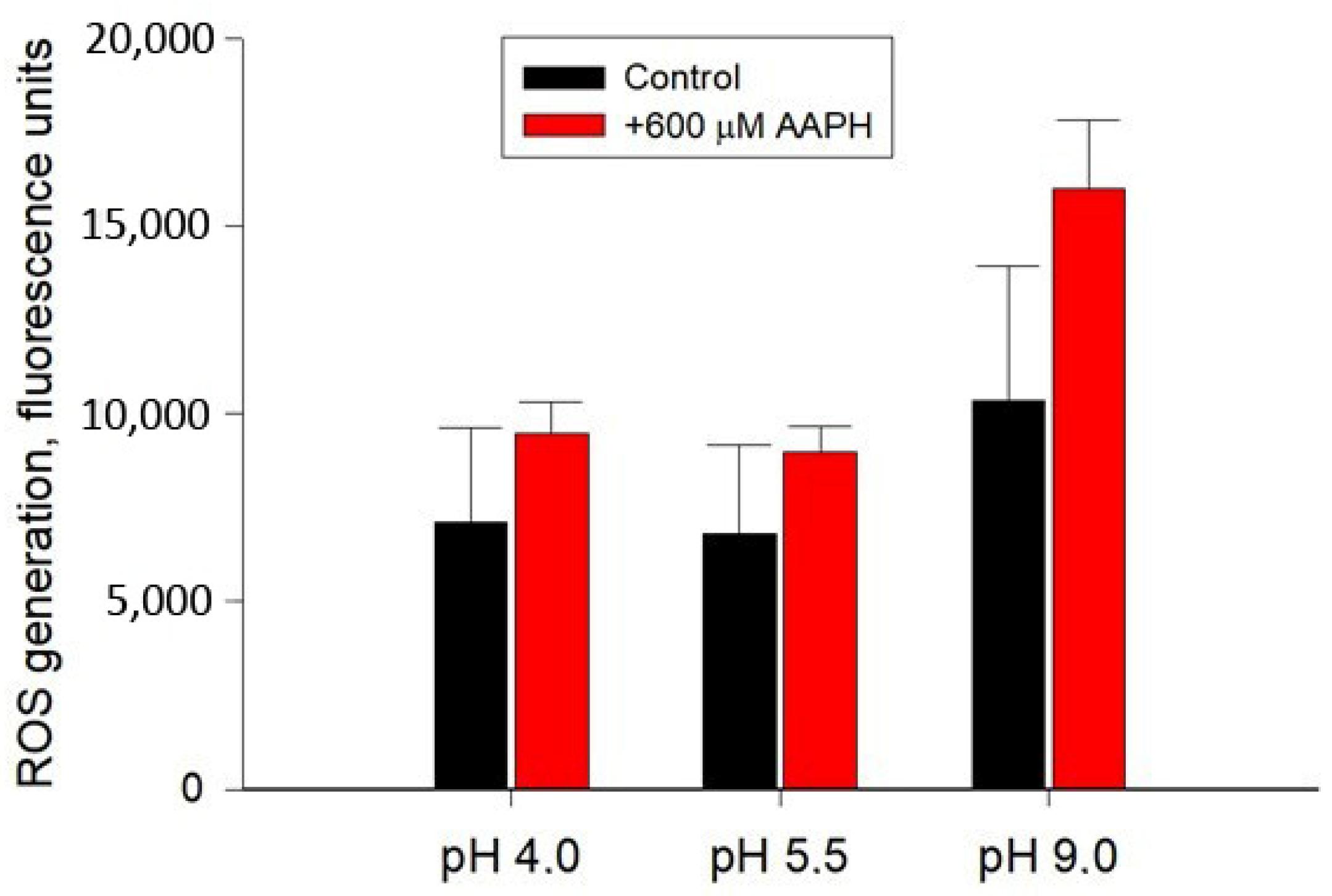
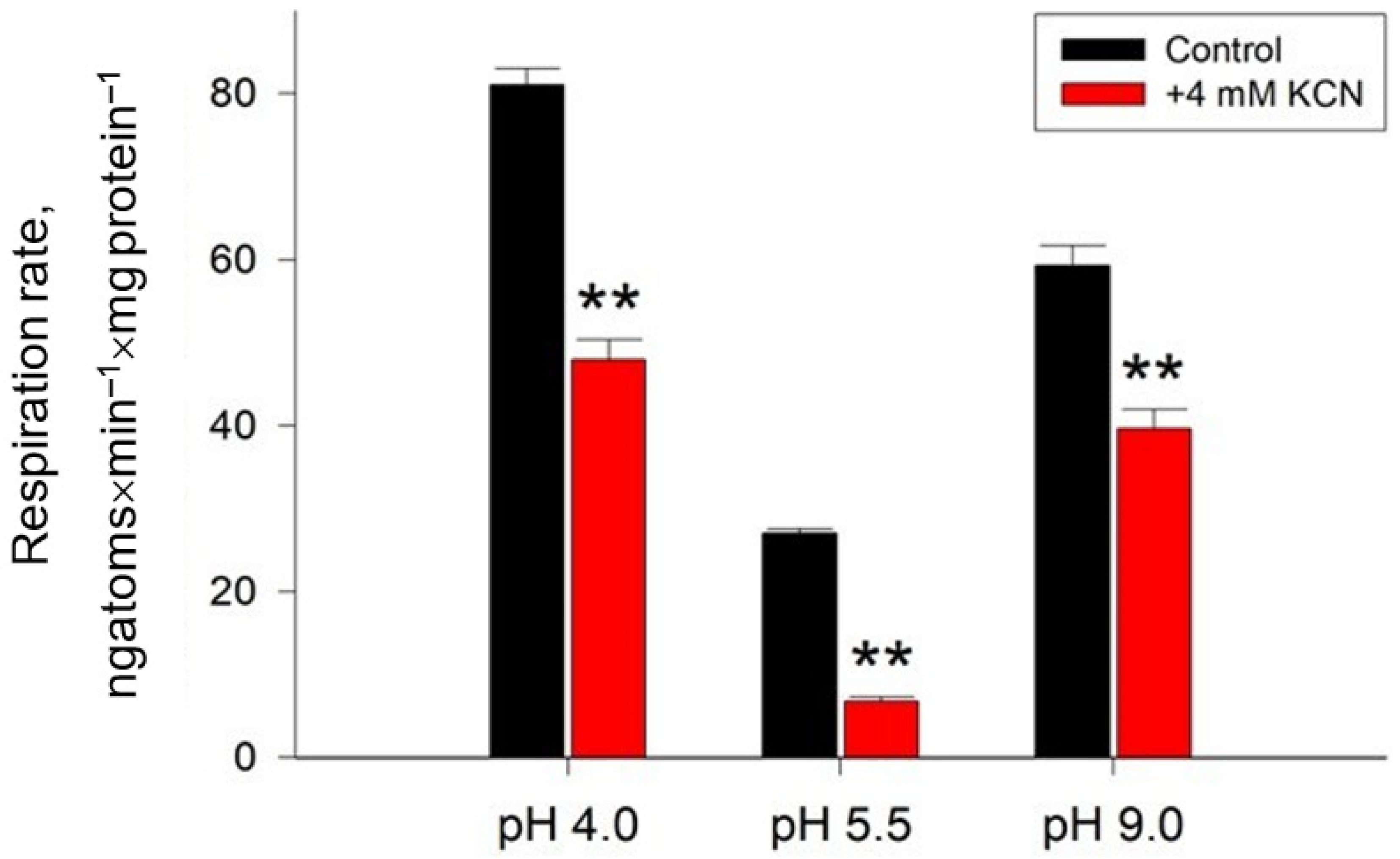
2.1. Growth, ROS Generation, and Respiratory Activity of the Y. lipolytica Yeast under Different Ambient pH
2.2. Isolation and Purification of Aconitate Hydratase from the Y. lipolytica Yeast Grown at Different pH
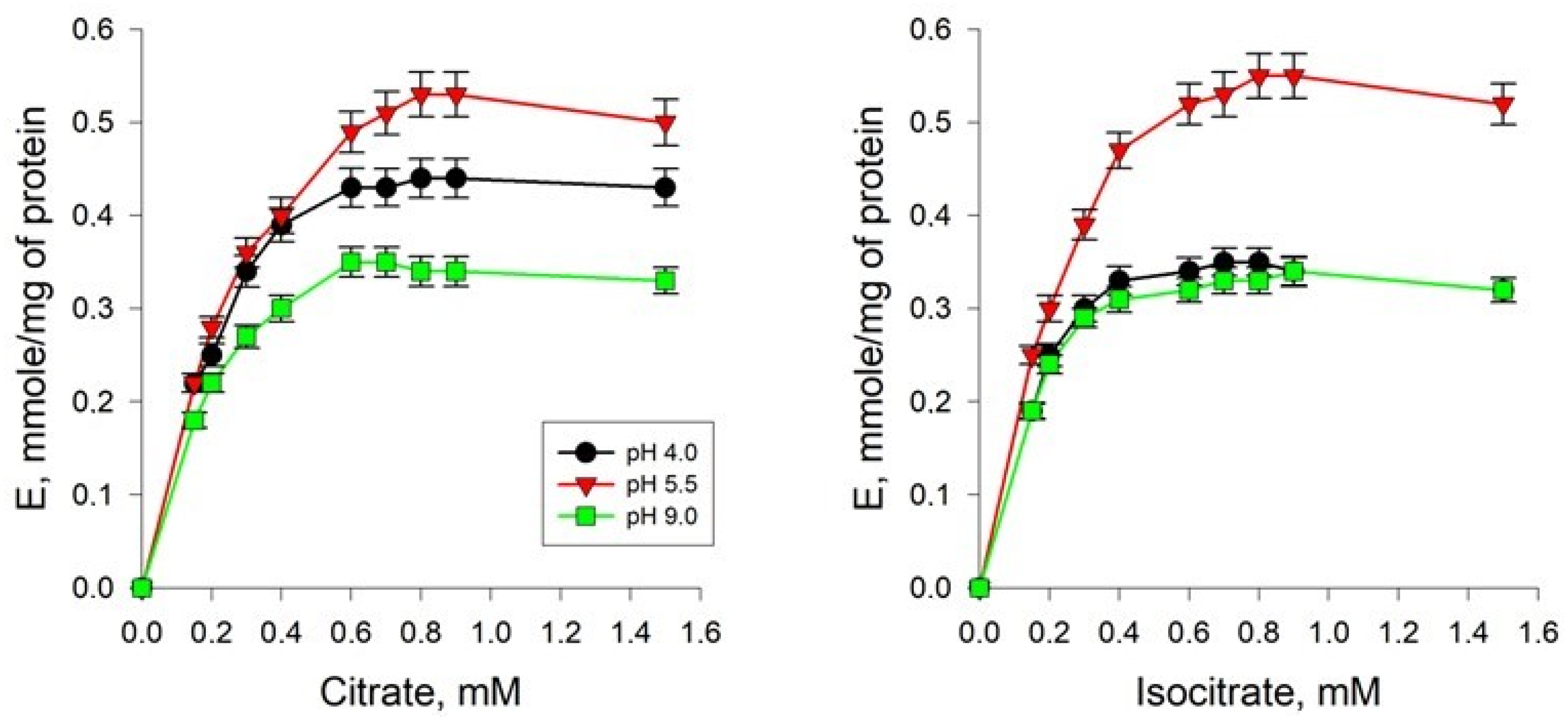
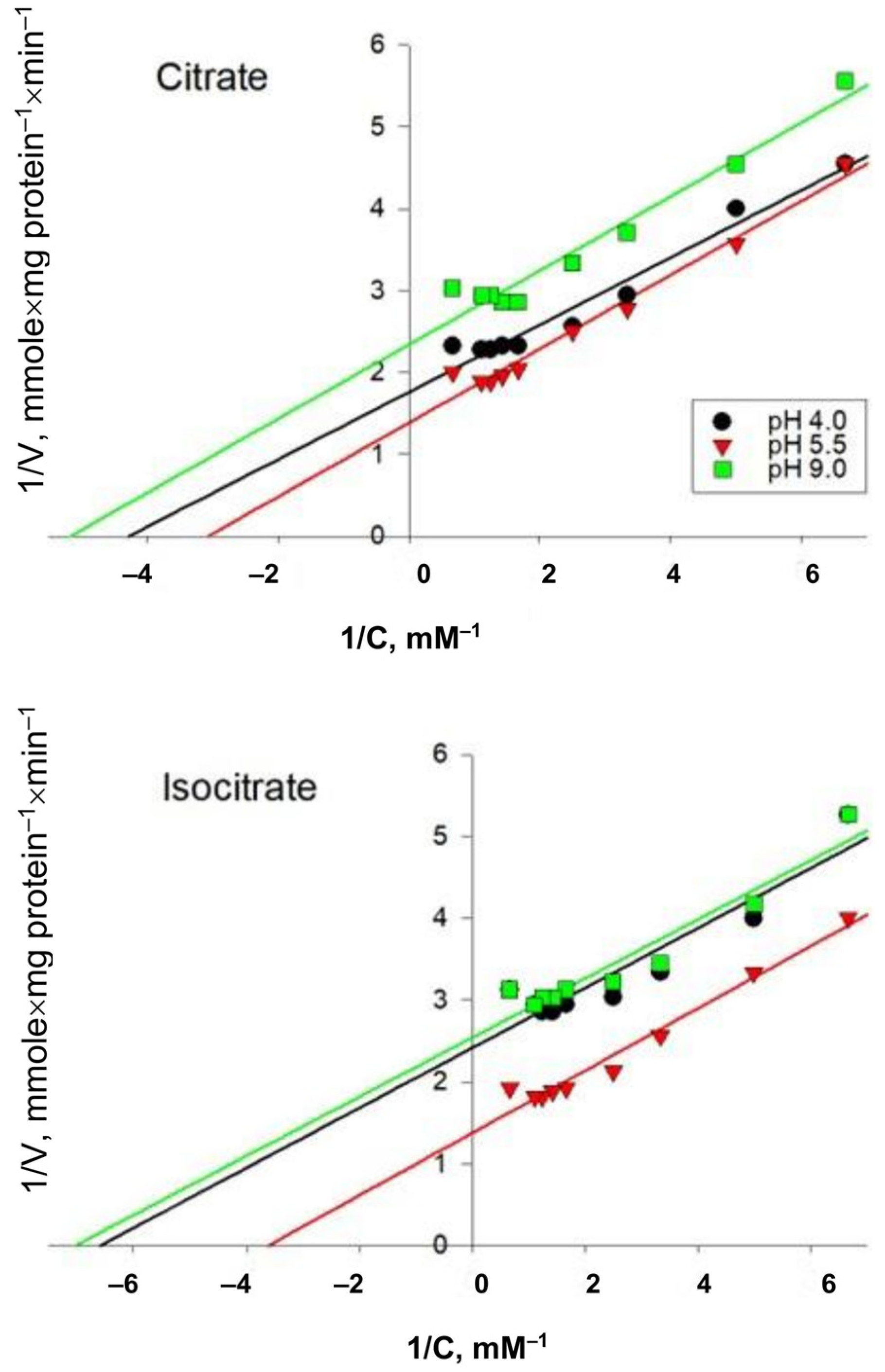
2.3. Study of the Kinetics of the Reaction Catalyzed by AH from Y. lipolytica Cells Grown on Media with Different pH Values
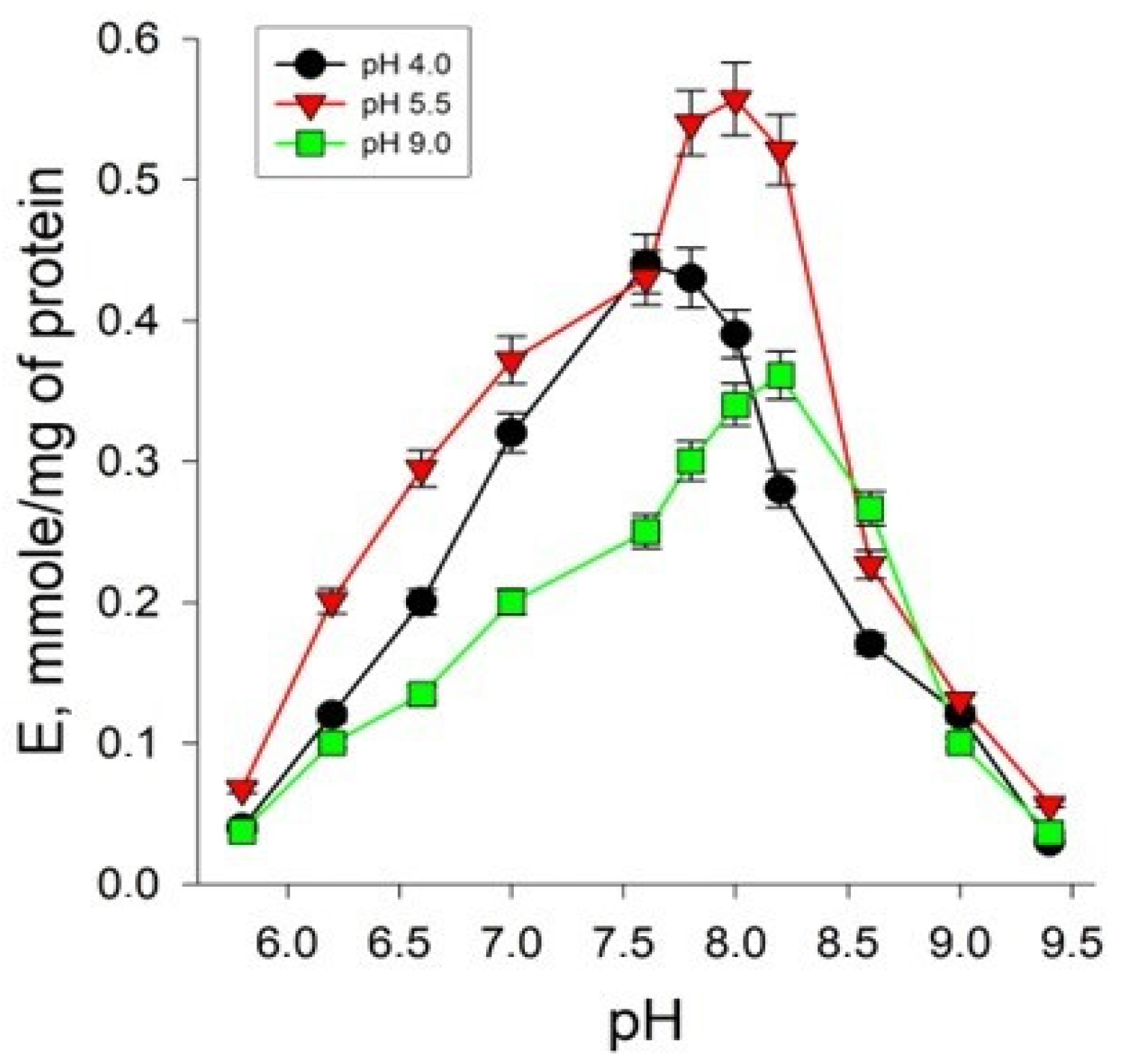
2.4. Study of the Influence of the Concentration of Hydrogen Ions on the Rate of the Enzymatic Reaction Catalyzed by AH Isolated from Yeast Cells under Different Cultivation Conditions
2.5. Regulatory Properties of AH from Y. lipolytica Cells Grown on Media with Different pH Values
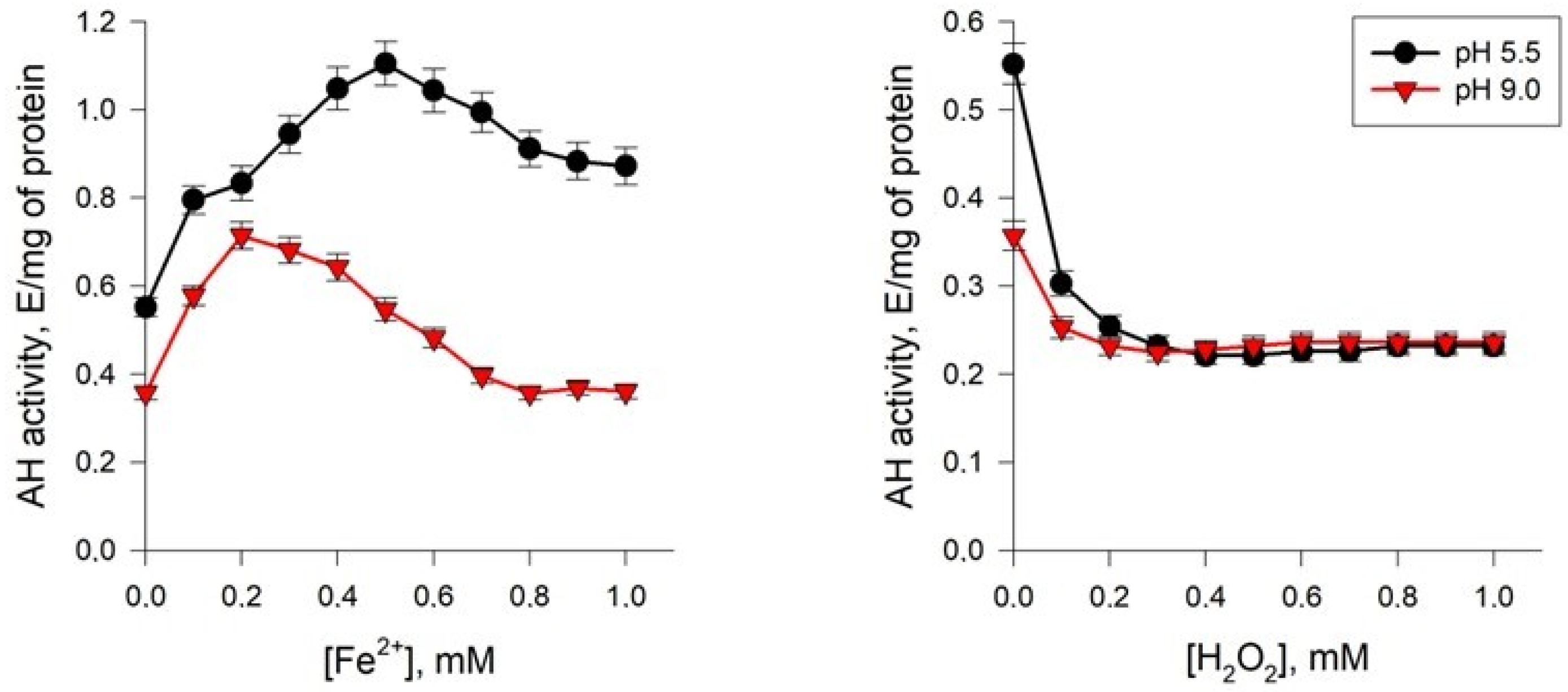
2.5.1. Effect of Fe2+ Ions
2.5.2. Effect of H2O2
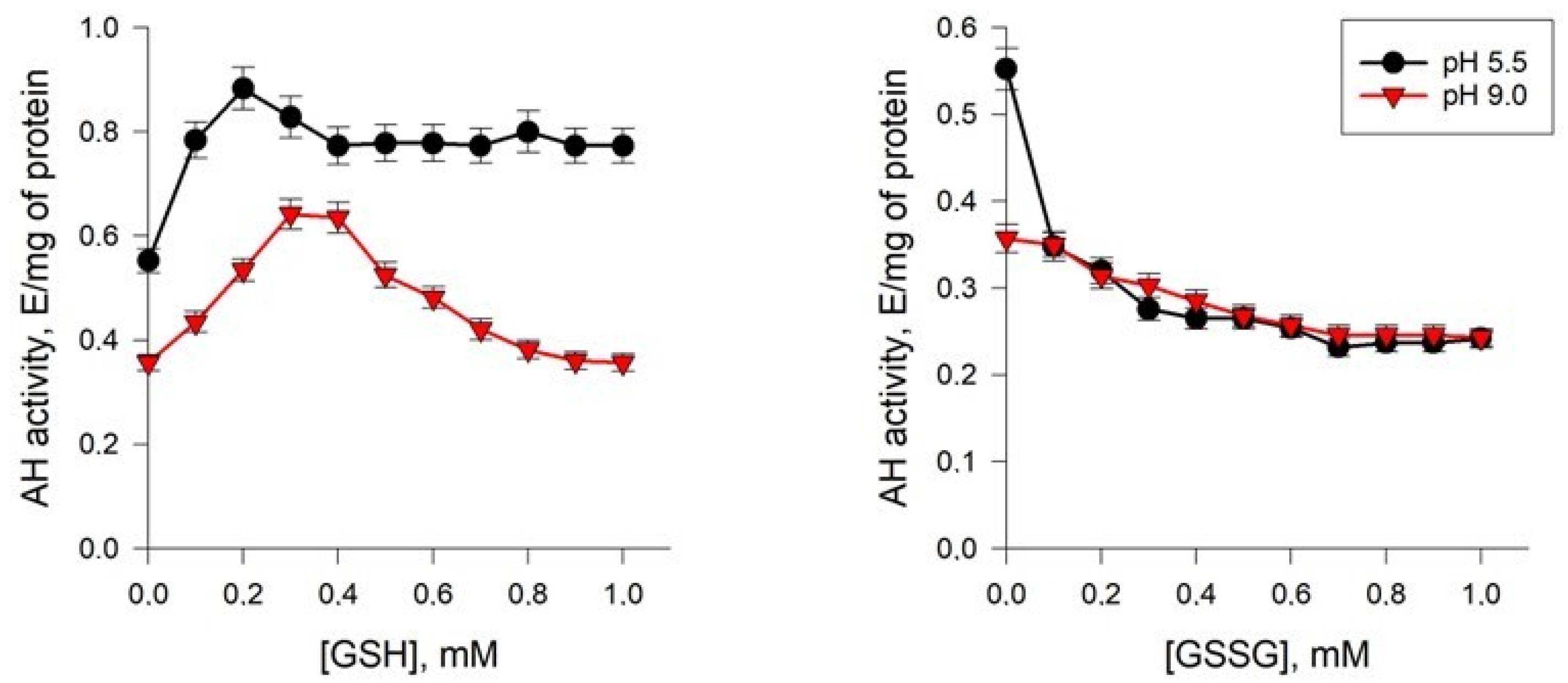
2.5.3. Effects of Glutathione
3. Discussion
4. Conclusions
5. Materials and Methods
5.1. Yeast Strains and Growth Conditions
5.2. Detection of ROS
5.3. Cell Respiration
5.4. Preparation of Cellular Homogenate
5.5. Purification of AH from Y. lipolytica Cells
5.5.1. Gel Filtration on Sephadex G-25
5.5.2. Ion Exchange Chromatography on DEAE-Cellulose
5.6. Measurement of AH Activity
5.7. Determining the Amount of Protein
5.8. Statistical Processing of Experimental Data
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Castro, L.; Tórtora, V.; Mansilla, S.; Radi, R. Aconitases: Non-redox Iron–Sulfur Proteins Sensitive to Reactive Species. Acc. Chem. Res. 2019, 52, 2609–2619. [Google Scholar] [CrossRef] [PubMed]
- Hirling, H.; Henderson, B.R.; Kühn, L.C. Mutational analysis of the [4Fe-4S]-cluster converting iron regulatory factor from its RNA-binding form to cytoplasmic aconitase. EMBO J. 1994, 13, 453–461. [Google Scholar] [CrossRef]
- Zhu, T.; Xiao, Z.; Yuan, H.; Tian, H.; Chen, T.; Chen, Q.; Chen, M.; Yang, J.; Zhou, Q.; Guo, W.; et al. ACO1 and IREB2 downregulation confer poor prognosis and correlate with autophagy-related ferroptosis and immune infiltration in KIRC. Front. Oncol. 2022, 12, 929838. [Google Scholar] [CrossRef] [PubMed]
- Brown, N.M.; Kennedy, M.C.; Antholine, W.E.; Eisenstein, R.S.; Walden, W.E. Detection of a [3Fe-4S] cluster intermediate of cytosolic aconitase in yeast expressing iron regulatory protein 1. Insights into the mechanism of fe-s cluster cycling. J. Biol. Chem. 2002, 277, 7246–7254. [Google Scholar] [CrossRef] [PubMed]
- Lushchak, O.V.; Piroddi, M.; Galli, F.; Lushchak, V.I. Aconitase post-translational modification as a key in linkage between Krebs cycle, iron homeostasis, redox signaling, and metabolism of reactive oxygen species. Redox Rep. 2014, 19, 8–15. [Google Scholar] [CrossRef] [PubMed]
- Tong, W.-H.; Rouault, T.A. Metabolic regulation of citrate and iron by aconitases: Role of iron–sulfur cluster biogenesis. Biometals 2007, 20, 549–564. [Google Scholar] [CrossRef]
- Muckenthaler, M.U.; Galy, B.; Hentze, M.W. Systemic iron homeostasis and the iron-responsive element/iron-regulatory protein (IRE/IRP) regulatory network. Annu. Rev. Nutr. 2008, 28, 197–213. [Google Scholar] [CrossRef] [PubMed]
- You, X.; Tian, J.; Zhang, H.; Guo, Y.; Yang, J.; Zhu, C.; Song, M.; Wang, P.; Liu, Z.; Cancilla, J.; et al. Loss of mitochondrial aconitase promotes colorectal cancer progression via SCD1-mediated lipid remodeling. Mol. Metab. 2021, 48, 101203. [Google Scholar] [CrossRef]
- Kim, S.L.; Shin, S.; Yang, S.J. Iron Homeostasis and Energy Metabolism in Obesity. Clin. Nutr. Res. 2022, 11, 316–330. [Google Scholar] [CrossRef]
- Gardner, P.R.; Nguyen, D.D.; White, C.W. Aconitase is a sensitive and critical target of oxygen poisoning in cultured mammalian cells and rat lungs. Proc. Nat. Acad. Sci. USA 1994, 91, 12248–12252. [Google Scholar] [CrossRef] [PubMed]
- Gardner, P.R.; Raineri, I.; Epstein, L.B.; White, C.W. Superoxide radical and iron modulated aconitase activity in mammalian cells. J. Biol. Chem. 1995, 270, 11399–13405. [Google Scholar] [CrossRef] [PubMed]
- Murakami, K.; Yoshino, M. Inactivation of aconitase in yeast exposed to oxidative stress. Biochem. Mol. Biol. Int. 1997, 41, 481–486. [Google Scholar] [CrossRef] [PubMed]
- Jung, M.; Mertens, C.; Tomat, E.; Brüne, B. Iron as a central player and promising target in cancer progression. Int. J. Mol. Sci. 2019, 20, 273. [Google Scholar] [CrossRef] [PubMed]
- Phillips, J.; Kinikini, D.; Yu, Y.; Guo, B.; Leibold, E. Differential regulation of IRP1 and IRP2 by nitric oxide in rat hepatoma cells. Blood 1996, 87, 2983–2992. [Google Scholar] [CrossRef]
- Chen, G.; Fillebeen, C.; Wang, J.; Pantopoulos, K. Overexpression of iron regulatory protein 1 suppresses growth of tumor xenografts. Carcinogenesis 2007, 28, 785–791. [Google Scholar] [CrossRef]
- Gonzalez-Sanchez, L.; Cobos-Fernandez, M.A.; Lopez-Nieva, P.; Villa-Morales, M.C.; Stamatakis, K.; Cuezva, J.M.; Marin-Rubio, J.L.; Vazquez-Dominguez, I.; Gonzalez-Vasconcellos, I.; Salido, E.; et al. Exploiting the passenger ACO1-deficiency arising from 9p21 deletions to kill T-cell lymphoblastic neoplasia cells. Carcinogenesis 2020, 41, 1113–1122. [Google Scholar] [CrossRef]
- Liu, Z.; Yang, Z.; Xiong, L.; Li, D.; Zou, Q.; Yuan, Y. ACO2 and ANPEP as novel prognostic markers for gallbladder squamous cell/adenosquamous carcinomas and adenocarcinomas. Int. J. Clin. Oncol. 2020, 25, 1346–1355. [Google Scholar] [CrossRef] [PubMed]
- Gibson, S.; Azamian, M.S.; Lalani, S.R.; Yen, K.G.; Sutton, V.R.; Scott, D.A. Recessive ACO2 variants as a cause of isolated ophthalmologic phenotypes. Am. J. Med. Genet. A 2020, 182, 1960–1966. [Google Scholar] [CrossRef] [PubMed]
- Zvyagilskaya, R.A.; Parkhomenko, O.A.; Gordeeva, A.V.; Deryabina, Y.I.; Persson, B.L. Bioenergetics of Yarrowia lipolytica cells grown at alkaline conditions. Biosci. Rep. 2004, 24, 117–125. [Google Scholar] [CrossRef]
- Epova, E.Y.; Balovneva, M.V.; Isakova, E.P.; Kudykina, Y.K.; Zylkova, M.V.; Deryabina, Y.I.; Shevelev, A.B. Expression system for Yarrowia lipolytica based on a promoter of the mitochondrial potential-dependent porin VDAC gene. Biotechnol. Bioprocess Eng. 2016, 21, 408–413. [Google Scholar] [CrossRef]
- Liu, H.-H.; Ji, X.-J.; Huang, H. Biotechnological applications of Yarrowia lipolytica: Past, present and future. Biotechnol. Adv. 2015, 33, 1522–1546. [Google Scholar] [CrossRef] [PubMed]
- Praphailong, W.; Van Gestel, M.; Fleet, G.H.; Heard, G.M. Evaluation of the Biolog system for the identification of food and beverage yeasts. Lett. Appl. Microbiol. 1997, 24, 455–459. [Google Scholar] [CrossRef] [PubMed]
- Farooq, M.A.; Pracheil, T.M.; Dong, Z.; Xiao, F.; Liu, Z. Mitochondrial DNA instability in cells lacking aconitase correlates with iron citrate toxicity. Oxid. Med. Cell. Longev. 2013, 2013, 493536. [Google Scholar] [CrossRef]
- Chen, X.J.; Wang, X.; Butow, R.A. Yeast aconitase binds and provides metabolically coupled protection to mitochondrial DNA. Proc. Natl. Acad. Sci. USA 2007, 104, 13738–13743. [Google Scholar] [CrossRef]
- Cho, S.-Y.; Jung, S.-J.; Kim, K.-D.; Roe, J.-H. Non-mitochondrial aconitase regulates the expression of iron-uptake genes by controlling the RNA turnover process in fission yeast. J. Microbiol. 2021, 59, 1075–1082. [Google Scholar] [CrossRef] [PubMed]
- Isakova, E.P.; Matushkina, I.N.; Popova, T.N.; Dergacheva, D.I.; Gessler, N.N.; Klein, O.I.; Semenikhina, A.V.; Deryabina, Y.I.; La Porta, N.; Saris, N.-E.L. Metabolic Remodeling during Long-Lasting Cultivation of the Endomyces magnusii Yeast on Oxidative and Fermentative Substrates. Microorganisms 2020, 8, 91. [Google Scholar] [CrossRef] [PubMed]
- da Silva, S.M.; Batista-Nascimento, L.; Gaspar-Cordeiro, A.; Vernis, L.; Pimentel, C.; Rodrigues-Pousada, C. Transcriptional regulation of Fe S biogenesis genes: A possible shield against arsenate toxicity activated by Yap1. Biochim. Biophys. Acta Gen. Subj. 2018, 1862, 2152–2161. [Google Scholar] [CrossRef]
- Mounkoro, P.; Michel, T.; Blandin, S.; Golinelli-Cohen, M.-P.; Davioud-Charvet, E.; Meunier, B. Investigating the mode of action of the redox-active antimalarial drug plasmodione using the yeast model. Free. Radic. Biol. Med. 2019, 141, 269–278. [Google Scholar] [CrossRef] [PubMed]
- Murakami, K.; Yoshino, M. Generation of reactive oxygen species by hydroxypyridone compound/iron complexes. Redox Rep. 2020, 25, 59–63. [Google Scholar] [CrossRef] [PubMed]
- Murakami, K.; Yoshino, M. Prooxidant activity of aminophenol compounds: Copper-dependent generation of reactive oxygen species. Biometals 2022, 35, 329–334. [Google Scholar] [CrossRef] [PubMed]
- Sekova, V.Y.; Dergacheva, D.I.; Isakova, E.P.; Gessler, N.N.; Tereshina, V.M.; Deryabina, Y.I. Soluble Sugar and Lipid Readjustments in the Yarrowia lipolytica Yeast at Various Temperatures and pH. Metabolites 2019, 9, 307. [Google Scholar] [CrossRef] [PubMed]
- Grum-Grzhimaylo, O.A.; Debets, A.J.; Bilanenko, E.N. The diversity of microfungi in peatlands originated from the White Sea. Mycologia 2016, 108, 233–254. [Google Scholar] [CrossRef] [PubMed]
- Vishwakarma, A.; Dalal, A.; Tetali, S.D.; Kirti, P.B.; Padmasree, K. Genetic engineering of AtAOX1a in Saccharomyces cerevisiae prevents oxidative damage and maintains redox homeostasis. FEBS Open Bio 2016, 6, 135–146. [Google Scholar] [CrossRef]
- Malecki, M.; Kamrad, S.; Ralser, M.; Bähler, J. Mitochondrial respiration is required to provide amino acids during fermentative proliferation of fission yeast. EMBO Rep. 2020, 21, e50845. [Google Scholar] [CrossRef]
- Medentsev, A.G.; Arinbasarova, A.Y.; Akimenko, V.K. Regulation and physiological role of cyanide-resistant oxidases in fungi and plants. Biochemistry 1999, 64, 1230–1243. [Google Scholar] [PubMed]
- Magnan, C.; Yu, J.; Chang, I.; Jahn, E.; Kanomata, Y.; Wu, J.; Zeller, M.; Oakes, M.; Baldi, P.; Sandmeyer, S. Sequence assembly of Yarrowia lipolytica strain W29/CLIB89 shows transposable element diversity. PLoS ONE 2016, 11, e0162363. [Google Scholar] [CrossRef] [PubMed]
- Medentsev, A.G.; Arinbasarova, A.Y.; Golovchenko, N.P.; Akimenko, V.K. Involvement of the alternative oxidase in respiration of Yarrowia lipolytica mitochondria is controlled by the activity of the cytochrome pathway. FEMS Yeast Res. 2002, 2, 519–524. [Google Scholar] [PubMed]
- Matasova, L.V.; Popova, T.N. Aconitate hydratase of mammals under oxidative stress. Biochemistry 2008, 73, 957–964. [Google Scholar] [CrossRef]
- Radi, R. Oxygen radicals, nitric oxide, and peroxynitrite: Redox pathways in molecular medicine. Proc. Natl. Acad. Sci. USA 2018, 115, 5839–5848. [Google Scholar] [CrossRef]
- Vasquez-Vivar, J.; Kalyanaraman, B.; Kennedy, M.C. Mitochondrial aconitase is a source of hydroxyl radical. An electron spin resonance investigation. J. Biol. Chem. 2000, 275, 14064–14069. [Google Scholar] [CrossRef]
- Baumgart, M.; Bott, M. Biochemical characterisation of aconitase from Corynebacterium glutamicum. J. Biotechnol. 2011, 154, 163–170. [Google Scholar] [CrossRef] [PubMed]
- Kennedy, S.C.; Rauner, R.; Gawron, O. On pig heart aconitase. Biochem. Biophys. Res. Commun. 1972, 47, 740–745. [Google Scholar] [CrossRef] [PubMed]
- Brouquisse, R.; James, F.; Raymond, P.; Pradet, A. Study of glucose starvation in excised maize root tips. Plant Physiol. 1991, 96, 619–626. [Google Scholar] [CrossRef] [PubMed]
- Brouquisse, R.; Nishimura, M.; Gaillard, J.; Douce, R. Characterization of a cytosolic aconitase in higher plant cells. Plant Physiol. 1987, 84, 1402–1407. [Google Scholar] [CrossRef]
- Stiben, J.; So, M.; Kaguni, L.S. Participation of iron-sulfur clusters in mitochondrial metabolism: The diversity of functions of this cofactor. Biochemistry 2016, 81, 1332–1348. [Google Scholar] [CrossRef]
- Mumby, S.; Koizumi, M.; Taniguchi, N.; Gutteridge, J.M. Reactive iron species in biological fluids activate the iron–sulphur cluster of aconitase. Biochim. Biophys. Acta 1998, 1380, 102–108. [Google Scholar] [CrossRef]
- Geng, P.; Zhang, L.; Shi, G.Y. Omics analysis of acetic acid tolerance in Saccharomyces cerevisiae. World J. Microbiol. Biotechnol. 2017, 33, 94. [Google Scholar] [CrossRef] [PubMed]
- Martínez-Pastor, M.T.; Perea-García, A.; Puig, S. Mechanisms of iron sensing and regulation in the yeast Saccharomyces cerevisiae. World J. Microbiol. Biotechnol. 2017, 33, 75. [Google Scholar] [CrossRef]
- Devaux, F.; Thiébaut, A. The regulation of iron homeostasis in the fungal human pathogen Candida glabrata. Microbiology 2019, 165, 1041–1060. [Google Scholar] [CrossRef] [PubMed]
- Gerwien, F.; Skrahina, V.; Kasper, L.; Hube, B.; Brunke, S. Metals in fungal virulence. FEMS Microbiol. Rev. 2018, 42, fux050. [Google Scholar] [CrossRef]
- Ueta, R.; Fujiwara, N.; Iwai, K.; Yamaguchi-Iwai, Y. Iron-induced dissociation of the Aft1p transcriptional regulator from target gene promoters is an initial event in iron-dependent gene suppression. Mol. Cell. Biol. 2012, 32, 4998–5008. [Google Scholar] [CrossRef] [PubMed]
- Penalva, M.A.; Lucena-Agell, D.; Arst, H.N., Jr. Liaison alcaline: Pals entice non-endosomal ESCRTs to the plasma membrane for pH signaling. Curr. Opin. Microbiol. 2014, 22, 49–59. [Google Scholar] [CrossRef] [PubMed]
- Markina-Iñarrairaegui, A.; Spielvogel, A.; Etxebeste, O.; Ugalde, U.; Espeso, E.A. Tolerance to alkaline ambient pH in Aspergillus nidulans depends on the activity of ENA proteins. Sci. Rep. 2020, 10, 14325. [Google Scholar] [CrossRef]
- Sies, H. Oxidative eustress: On constant alert for redox homeostasis. Redox Biol. 2021, 41, 101867. [Google Scholar] [CrossRef] [PubMed]
- Andreeshcheva, E.M.; Popova, T.N.; Artyukhov, V.G.; Matasova, L.V. Free radical oxidation and catalytic activity of aconitate hydratase in rat liver under normal conditions and during toxic hepatitis. Bull. Exp. Biol. Med. 2004, 137, 352–354. [Google Scholar] [CrossRef]
- Kennedy, M.C.; Spoto, G.; Emptage, M.H.; Beinert, H. The active site sulfhydryl of aconitase is not required for catalytic activity. J. Biol. Chem. 1988, 263, 8190–8193. [Google Scholar] [CrossRef]
- Plank, D.W.; Howard, J.B. Identification of the reactive sulfhydryl and sequences of cysteinyl-tryptic peptides from beef heart aconitase. J. Biol. Chem. 1988, 263, 8184–8189. [Google Scholar] [CrossRef] [PubMed]
- Popova, T.N.; de Carvalho, M.P. Citrate and isocitrate in plant metabolism. Biochim. Biophys. Acta 1998, 1364, 307–325. [Google Scholar] [CrossRef] [PubMed]
- Li, X.; Wang, P.; Ge, Y.; Wang, W.; Abbas, A.; Zhu, G. NADP+-specific isocitrate dehydrogenase from oleaginous yeast Yarrowia lipolytica CLIB122: Biochemical characterization and coenzyme sites evaluation. Appl. Biochem. Biotechnol. 2013, 171, 403–416. [Google Scholar] [CrossRef]
- Nguyen, T.T.; Kitajima, S.; Izawa, S.J. Importance of glucose-6-phosphate dehydrogenase (G6PDH) for vanillin tolerance in Saccharomyces cerevisiae. Biosci Bioeng. 2014, 118, 263–269. [Google Scholar] [CrossRef]
- Brown, S.M.; Upadhya, R.; Shoemaker, J.D.; Lodge, J.K. Isocitrate dehydrogenase is important for nitrosative stress resistance in Cryptococcus neoformans, but oxidative stress resistance is not dependent on glucose-6-phosphate dehydrogenase. Eukaryot. Cell 2010, 9, 971–980. [Google Scholar] [CrossRef] [PubMed]
- Dhatariya, K.K.; Glaser, N.S.; Codner, E.; Umpierrez, G.E. Diabetic ketoacidosis. Nat. Rev. Dis. Prim. 2020, 6, 40. [Google Scholar] [CrossRef]
- Zanza, C.; Facelli, V.; Romenskaya, T.; Bottinelli, M.; Caputo, G.; Piccioni, A.; Franceschi, F.; Saviano, A.; Ojetti, V.; Savioli, G.; et al. Lactic Acidosis Related to Pharmacotherapy and Human Diseases. Pharmaceuticals 2022, 15, 1496. [Google Scholar] [CrossRef]
- Emmett, M. Metabolic Alkalosis: A Brief Pathophysiologic Review. Clin. J. Am. Soc. Nephrol. 2020, 15, 1848–1856. [Google Scholar] [CrossRef] [PubMed]
- Kim, R.S.; LaBella, F.S. Comparison of analytical methods for monitoring autoxidation profiles of authentic lipids. J. Lipid Res. 1987, 28, 1110–1117. [Google Scholar] [CrossRef] [PubMed]
- Deryabina, Y.; Isakova, E.; Antipov, A.; Saris, N.-E.L. The inhibitors of antioxidant cell enzymes induce permeability transition in yeast mitochondria. J. Bioenerg. Biomembr. 2013, 45, 491–504. [Google Scholar] [CrossRef] [PubMed]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Stage of Purification | Group | Total Activity, Units | The Protein Amount, mg | Specific Activity, E/mg Protein | Yield, % | Degree of Purification |
|---|---|---|---|---|---|---|
| Homogenate | 1 (pH 4.0) | 2.25 ± 0.101 | 250 ± 11.5 | 0.009 ± 0.0003 | 100 | 1 |
| 2 (pH 5.5) | 2.79 ± 0.112 a,b | 233 ± 10.8 a | 0.012 ± 0.0003 a | 100 | 1 | |
| 3 (pH 9.0) | 1.94 ± 0.096 b | 277 ± 12.3 b | 0.007 ± 0.0002 b | 100 | 1 | |
| Chromatography on sephadex G-25 | 1 (pH 4.0) | 1.77 ± 0.082 | 183 ± 9.2 | 0.015 ± 0.0006 | 79 | 1.7 |
| 2 (pH 5.5) | 2.29 ± 0.111 a,c | 111 ± 5.2 a | 0.018 ± 0.0007 a | 82 | 1.5 | |
| 3 (pH 9.0) | 1.43 ± 0.061 | 182 ± 8.5 | 0.012 ± 0.0006 | 74 | 1.7 | |
| Chromatography on DEAE cellulose | 1 (pH 4.0) | 0.765 ± 0.034 c | 1.77 ± 0.08 c | 0.432 ± 0.0212 c | 34 | 48 |
| 2 (pH 5.5) | 0.865 ± 0.042 d | 1.57 ± 0.07 | 0.552 ± 0.0271 | 31 | 46 | |
| 3 (pH 9.0) | 0.357 ± 0.015 d | 1.58 ± 0.07 | 0.357 ± 0.0158 d | 29 | 51 |
| Growth Conditions | Citrate | Isocitrate | pH-Optimum | ||
|---|---|---|---|---|---|
| Vmax, mmols/mg of Protein- | Km, mM | Vmax, mmols/mg of Protein- | Km, mM | ||
| pH 4.0 | 0.570 ± 0.042 a | 0.230 ± 0.009 b,c | 0.415 ± 0.017 a | 0.150 ± 0.008 c | 7.6 |
| pH 5.5 | 0.720 ± 0.031 c | 0.320 ± 0.011 | 0.726 ± 0.037 c | 0.270 ± 0.015 b,c | 8.0 |
| pH 9.0 | 0.428 ± 0.022 a | 0.190 ± 0.007 b | 0.394 ± 0.016 a | 0.140 ± 0.006 c | 8.2 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Rakhmanova, T.I.; Sekova, V.Y.; Gessler, N.N.; Isakova, E.P.; Deryabina, Y.I.; Popova, T.N.; Shurubor, Y.I.; Krasnikov, B.F. Kinetic and Regulatory Properties of Yarrowia lipolytica Aconitate Hydratase as a Model-Indicator of Cell Redox State under pH Stress. Int. J. Mol. Sci. 2023, 24, 7670. https://doi.org/10.3390/ijms24087670
Rakhmanova TI, Sekova VY, Gessler NN, Isakova EP, Deryabina YI, Popova TN, Shurubor YI, Krasnikov BF. Kinetic and Regulatory Properties of Yarrowia lipolytica Aconitate Hydratase as a Model-Indicator of Cell Redox State under pH Stress. International Journal of Molecular Sciences. 2023; 24(8):7670. https://doi.org/10.3390/ijms24087670
Chicago/Turabian StyleRakhmanova, Tatyana I., Varvara Yu. Sekova, Natalya N. Gessler, Elena P. Isakova, Yulia I. Deryabina, Tatyana N. Popova, Yevgeniya I. Shurubor, and Boris F. Krasnikov. 2023. "Kinetic and Regulatory Properties of Yarrowia lipolytica Aconitate Hydratase as a Model-Indicator of Cell Redox State under pH Stress" International Journal of Molecular Sciences 24, no. 8: 7670. https://doi.org/10.3390/ijms24087670