
Human Embryonic Stem-Cell-Derived Exosomes Repress NLRP3 Inflammasome to Alleviate Pyroptosis in Nucleus Pulposus Cells by Transmitting miR-302c

Abstract
:1. Introduction
2. Results
2.1. NLRP3 Inflammasome Was Excessively Activated in Nucleus Pulposus (NP) Tissues with the Grades of IVDD
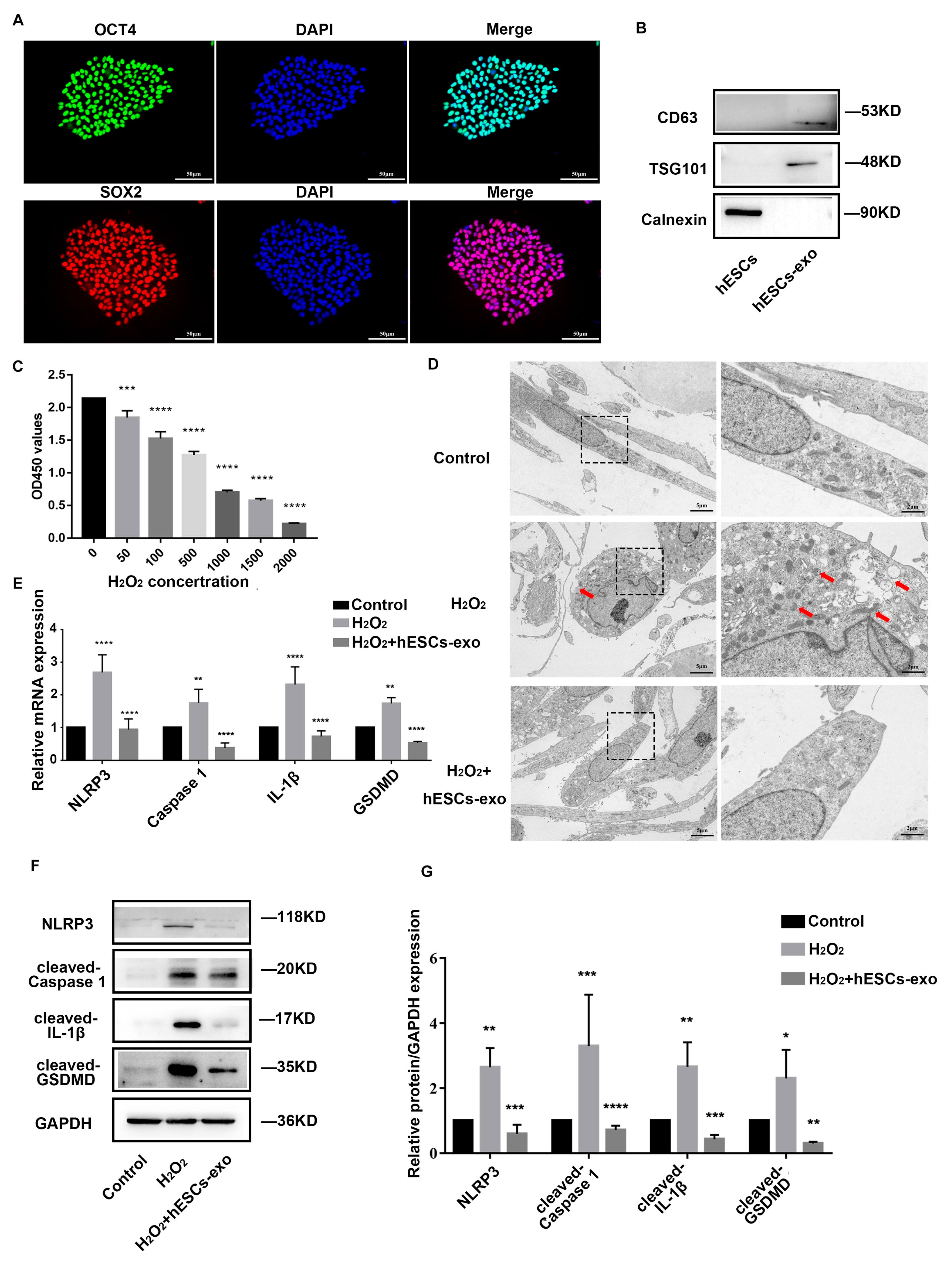
2.2. hESCs-Exo Attenuate Pyroptosis of NPCs Treated with H2O2
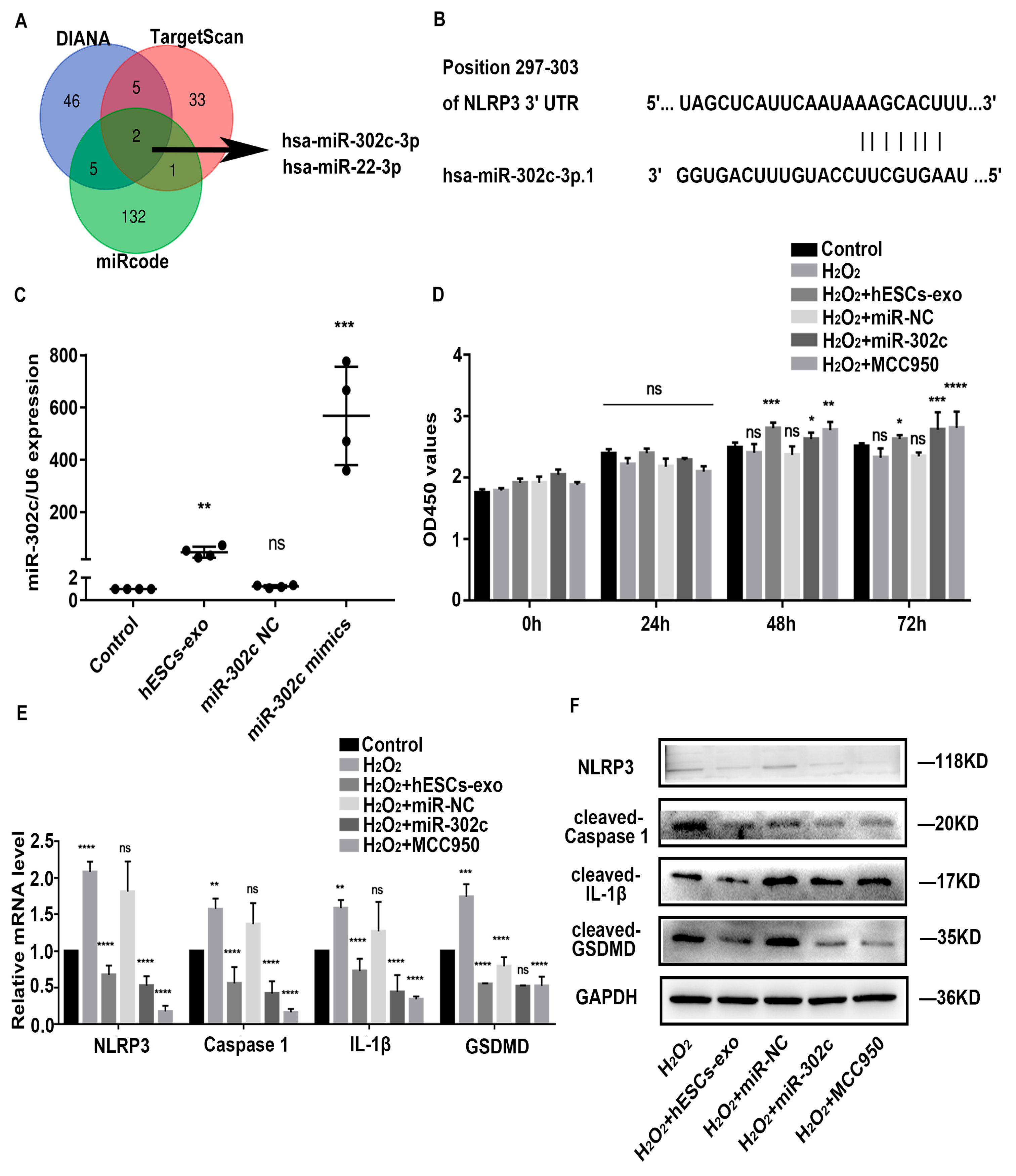
2.3. hESCs-Exo-Derived miR-302c Targets NLRP3 to Alleviate Pyroptosis of NPCs
2.4. hESCs-Exo-Derived miR302c Could Retard IVDD in a Rat Model
3. Discussion
4. Materials and Methods
4.1. Nucleus Pulposus Cells (NPCs) Isolation and Culture
4.2. Human Embryonic Stem Cells (hESCs) Culture
4.3. Human Embryonic Stem Cell-Derived Exosomes (hESCs-Exo) Isolation, Purification, and Characterization
4.4. hESCs-Exo Treatment of NPCs Induced by H2O2 Pyroptosis
4.5. Cell Transfection
4.6. Transmission Electron Microscopy (TEM)
4.7. Quantitative Real-Time Polymerase Chain Reaction (qRT-PCR)
4.8. Western Blot (WB)
4.9. Cell-Counting Kit-8 (CCK-8) Assay
4.10. Immunofluorescence (IF) Analysis
4.11. Rat IVDD Model
4.12. Histological Valuation
4.13. Statistical Analysis
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Kandel, R.; Roberts, S.; Urban, J.P. Tissue engineering and the intervertebral disc: The challenges. Eur. Spine J. 2008, 17 (Suppl. 4), 480–491. [Google Scholar] [CrossRef] [PubMed]
- Sun, K.; Zhu, J.; Yan, C.; Li, F.; Kong, F.; Sun, J.; Sun, X.; Shi, J.; Wang, Y. CGRP Regulates Nucleus Pulposus Cell Apoptosis and Inflammation via the MAPK/NF-kappaB Signaling Pathways during Intervertebral Disc Degeneration. Oxid. Med. Cell. Longev. 2021, 2021, 2958584. [Google Scholar] [CrossRef] [PubMed]
- Buckley, C.T.; Hoyland, J.A.; Fujii, K.; Pandit, A.; Iatridis, J.C.; Grad, S. Critical aspects and challenges for intervertebral disc repair and regeneration-Harnessing advances in tissue engineering. JOR Spine 2018, 1, e1029. [Google Scholar] [CrossRef] [PubMed]
- Lyu, F.J.; Cheung, K.M.; Zheng, Z.; Wang, H.; Sakai, D.; Leung, V.Y. IVD progenitor cells: A new horizon for understanding disc homeostasis and repair. Nat. Rev. Rheumatol. 2019, 15, 102–112. [Google Scholar] [CrossRef]
- Feng, C.; Yang, M.; Lan, M.; Liu, C.; Zhang, Y.; Huang, B.; Liu, H.; Zhou, Y. ROS: Crucial Intermediators in the Pathogenesis of Intervertebral Disc Degeneration. Oxid. Med. Cell. Longev. 2017, 2017, 5601593. [Google Scholar] [CrossRef]
- Molinos, M.; Almeida, C.R.; Caldeira, J.; Cunha, C.; Goncalves, R.M.; Barbosa, M.A. Inflammation in intervertebral disc degeneration and regeneration. J. R. Soc. Interface 2015, 12, 20141191. [Google Scholar] [CrossRef]
- Ma, K.; Chen, S.; Li, Z.; Deng, X.; Huang, D.; Xiong, L.; Shao, Z. Mechanisms of endogenous repair failure during intervertebral disc degeneration. Osteoarthr. Cartil. 2019, 27, 41–48. [Google Scholar] [CrossRef]
- Chen, Z.H.; Jin, S.H.; Wang, M.Y.; Jin, X.L.; Lv, C.; Deng, Y.F.; Wang, J.L. Enhanced NLRP3, caspase-1, and IL- 1beta levels in degenerate human intervertebral disc and their association with the grades of disc degeneration. Anat. Rec. 2015, 298, 720–726. [Google Scholar] [CrossRef]
- Penolazzi, L.; Bergamin, L.S.; Lambertini, E.; Poma, V.V.; Sarti, A.C.; De Bonis, P.; Di Virgilio, F.; Piva, R. The P2X7 purinergic receptor in intervertebral disc degeneration. J. Cell. Physiol. 2022, 237, 1418–1428. [Google Scholar] [CrossRef]
- Xia, C.; Zeng, Z.; Fang, B.; Tao, M.; Gu, C.; Zheng, L.; Wang, Y.; Shi, Y.; Fang, C.; Mei, S.; et al. Mesenchymal stem cell-derived exosomes ameliorate intervertebral disc degeneration via anti-oxidant and anti-inflammatory effects. Free Radic. Biol. Med. 2019, 143, 1–15. [Google Scholar] [CrossRef]
- Paik, S.; Kim, J.K.; Silwal, P.; Sasakawa, C.; Jo, E.K. An update on the regulatory mechanisms of NLRP3 inflammasome activation. Cell. Mol. Immunol. 2021, 18, 1141–1160. [Google Scholar] [CrossRef]
- Zhang, J.; Zhang, J.; Zhang, Y.; Liu, W.; Ni, W.; Huang, X.; Yuan, J.; Zhao, B.; Xiao, H.; Xue, F. Mesenchymal stem cells-derived exosomes ameliorate intervertebral disc degeneration through inhibiting pyroptosis. J. Cell. Mol. Med. 2020, 24, 11742–11754. [Google Scholar] [CrossRef]
- Bai, Z.; Liu, W.; He, D.; Wang, Y.; Yi, W.; Luo, C.; Shen, J.; Hu, Z. Protective effects of autophagy and NFE2L2 on reactive oxygen species-induced pyroptosis of human nucleus pulposus cells. Aging 2020, 12, 7534. [Google Scholar] [CrossRef]
- Chen, F.; Jiang, G.; Liu, H.; Li, Z.; Pei, Y.; Wang, H.; Pan, H.; Cui, H.; Long, J.; Wang, J.; et al. Melatonin alleviates intervertebral disc degeneration by disrupting the IL-1beta/NF-kappaB-NLRP3 inflammasome positive feedback loop. Bone Res. 2020, 8, 10. [Google Scholar] [CrossRef]
- Tang, P.; Gu, J.M.; Xie, Z.A.; Gu, Y.; Jie, Z.W.; Huang, K.M.; Wang, J.Y.; Fan, S.W.; Jiang, X.S.; Hu, Z.J. Honokiol alleviates the degeneration of intervertebral disc via suppressing the activation of TXNIP-NLRP3 inflammasome signal pathway. Free Radic. Biol. Med. 2018, 120, 368–379. [Google Scholar] [CrossRef]
- Chen, J.; Bian, M.; Pan, L.; Yang, H. alpha-Mangostin protects lipopolysaccharide-stimulated nucleus pulposus cells against NLRP3 inflammasome-mediated apoptosis via the NF-kappaB pathway. J. Appl. Toxicol. 2022, 42, 1467–1476. [Google Scholar] [CrossRef]
- Zhao, K.; An, R.; Xiang, Q.; Li, G.; Wang, K.; Song, Y.; Liao, Z.; Li, S.; Hua, W.; Feng, X.; et al. Acid-sensing ion channels regulate nucleus pulposus cell inflammation and pyroptosis via the NLRP3 inflammasome in intervertebral disc degeneration. Cell Prolif. 2021, 54, e12941. [Google Scholar] [CrossRef]
- Gerecht-Nir, S.; Itskovitz-Eldor, J. Human embryonic stem cells: A potential source for cellular therapy. Am. J. Transplant. 2004, 4 (Suppl. 6), 51–57. [Google Scholar] [CrossRef]
- He, Q.; Li, J.; Bettiol, E.; Jaconi, M.E. Embryonic stem cells: New possible therapy for degenerative diseases that affect elderly people. J. Gerontol. A Biol. Sci. Med. Sci. 2003, 58, 279–287. [Google Scholar] [CrossRef]
- Zhang, Y.; Hu, Y.; Wang, W.; Guo, Z.; Yang, F.; Cai, X.; Xiong, L. Current Progress in the Endogenous Repair of Intervertebral Disk Degeneration Based on Progenitor Cells. Front. Bioeng. Biotechnol. 2020, 8, 629088. [Google Scholar] [CrossRef]
- Diaz-Hernandez, M.E.; Khan, N.M.; Trochez, C.M.; Yoon, T.; Maye, P.; Presciutti, S.M.; Gibson, G.; Drissi, H. Derivation of notochordal cells from human embryonic stem cells reveals unique regulatory networks by single cell-transcriptomics. J. Cell. Physiol. 2020, 235, 5241–5255. [Google Scholar] [CrossRef] [PubMed]
- Hoben, G.M.; Willard, V.P.; Athanasiou, K.A. Fibrochondrogenesis of hESCs: Growth factor combinations and cocultures. Stem Cells Dev. 2009, 18, 283–292. [Google Scholar] [CrossRef] [PubMed]
- Sheikh, H.; Zakharian, K.; De La Torre, R.P.; Facek, C.; Vasquez, A.; Chaudhry, G.R.; Svinarich, D.; Perez-Cruet, M.J. In vivo intervertebral disc regeneration using stem cell-derived chondroprogenitors. J. Neurosurg. Spine 2009, 10, 265–272. [Google Scholar] [CrossRef] [PubMed]
- Khan, M.; Nickoloff, E.; Abramova, T.; Johnson, J.; Verma, S.K.; Krishnamurthy, P.; Mackie, A.R.; Vaughan, E.; Garikipati, V.N.; Benedict, C.; et al. Embryonic stem cell-derived exosomes promote endogenous repair mechanisms and enhance cardiac function following myocardial infarction. Circ. Res. 2015, 117, 52–64. [Google Scholar] [CrossRef] [PubMed]
- Chen, B.; Sun, Y.; Zhang, J.; Zhu, Q.; Yang, Y.; Niu, X.; Deng, Z.; Li, Q.; Wang, Y. Human embryonic stem cell-derived exosomes promote pressure ulcer healing in aged mice by rejuvenating senescent endothelial cells. Stem Cell Res. Ther. 2019, 10, 142. [Google Scholar] [CrossRef]
- Ke, Y.; Fan, X.; Hao, R.; Dong, L.; Xue, M.; Tan, L.; Yang, C.; Li, X.; Ren, X. Correction to: Human embryonic stem cell-derived extracellular vesicles alleviate retinal degeneration by upregulating Oct4 to promote retinal Muller cell retrodifferentiation via HSP90. Stem Cell Res. Ther. 2021, 12, 134. [Google Scholar] [CrossRef]
- Lee, W.H.; Chen, W.Y.; Shao, N.Y.; Xiao, D.; Qin, X.; Baker, N.; Bae, H.R.; Wei, T.T.; Wang, Y.; Shukla, P.; et al. Comparison of Non-Coding RNAs in Exosomes and Functional Efficacy of Human Embryonic Stem Cell- versus Induced Pluripotent Stem Cell-Derived Cardiomyocytes. Stem Cells 2017, 35, 2138–2149. [Google Scholar] [CrossRef]
- Singla, D.K.; Johnson, T.A.; Tavakoli Dargani, Z. Exosome Treatment Enhances Anti-Inflammatory M2 Macrophages and Reduces Inflammation-Induced Pyroptosis in Doxorubicin-Induced Cardiomyopathy. Cells 2019, 8, 1224. [Google Scholar] [CrossRef]
- Tavakoli Dargani, Z.; Singla, D.K. Embryonic stem cell-derived exosomes inhibit doxorubicin-induced TLR4-NLRP3-mediated cell death-pyroptosis. Am. J. Physiol. Heart. Circ. Physiol. 2019, 317, H460–H471. [Google Scholar] [CrossRef]
- Junqueira, L.C.; Bignolas, G.; Brentani, R.R. Picrosirius staining plus polarization microscopy, a specific method for collagen detection in tissue sections. Histochem. J. 1979, 11, 447–455. [Google Scholar] [CrossRef]
- Kubaski, F.; Osago, H.; Mason, R.W.; Yamaguchi, S.; Kobayashi, H.; Tsuchiya, M.; Orii, T.; Tomatsu, S. Glycosaminoglycans detection methods: Applications of mass spectrometry. Mol. Genet. Metab. 2017, 120, 67–77. [Google Scholar] [CrossRef]
- Suh, M.R.; Lee, Y.; Kim, J.Y.; Kim, S.K.; Moon, S.H.; Lee, J.Y.; Cha, K.Y.; Chung, H.M.; Yoon, H.S.; Moon, S.Y.; et al. Human embryonic stem cells express a unique set of microRNAs. Dev. Biol. 2004, 270, 488–498. [Google Scholar] [CrossRef]
- Ren, J.; Jin, P.; Wang, E.; Marincola, F.M.; Stroncek, D.F. MicroRNA and gene expression patterns in the differentiation of human embryonic stem cells. J. Transl. Med. 2009, 7, 20. [Google Scholar] [CrossRef]
- Sakai, D.; Nakamura, Y.; Nakai, T.; Mishima, T.; Kato, S.; Grad, S.; Alini, M.; Risbud, M.V.; Chan, D.; Cheah, K.S.; et al. Exhaustion of nucleus pulposus progenitor cells with ageing and degeneration of the intervertebral disc. Nat. Commun. 2012, 3, 1264. [Google Scholar] [CrossRef]
- Silva, M.J.; Holguin, N. Aging aggravates intervertebral disc degeneration by regulating transcription factors toward chondrogenesis. FASEB J. 2020, 34, 1970–1982. [Google Scholar] [CrossRef]
- Chen, S.; Wu, X.; Lai, Y.; Chen, D.; Bai, X.; Liu, S.; Wu, Y.; Chen, M.; Lai, Y.; Cao, H.; et al. Kindlin-2 inhibits Nlrp3 inflammasome activation in nucleus pulposus to maintain homeostasis of the intervertebral disc. Bone Res. 2022, 10, 5. [Google Scholar] [CrossRef]
- Liao, Z.; Li, S.; Liu, R.; Feng, X.; Shi, Y.; Wang, K.; Li, S.; Zhang, Y.; Wu, X.; Yang, C. Autophagic Degradation of Gasdermin D Protects against Nucleus Pulposus Cell Pyroptosis and Retards Intervertebral Disc Degeneration In Vivo. Oxid. Med. Cell. Longev. 2021, 2021, 5584447. [Google Scholar] [CrossRef]
- He, R.; Wang, Z.; Cui, M.; Liu, S.; Wu, W.; Chen, M.; Wu, Y.; Qu, Y.; Lin, H.; Chen, S.; et al. HIF1A Alleviates compression-induced apoptosis of nucleus pulposus derived stem cells via upregulating autophagy. Autophagy 2021, 17, 3338–3360. [Google Scholar] [CrossRef]
- Wu, J.; Chen, Y.; Liao, Z.; Liu, H.; Zhang, S.; Zhong, D.; Qiu, X.; Chen, T.; Su, D.; Ke, X.; et al. Self-amplifying loop of NF-kappaB and periostin initiated by PIEZO1 accelerates mechano-induced senescence of nucleus pulposus cells and intervertebral disc degeneration. Mol. Ther. 2022, 30, 3241–3256. [Google Scholar] [CrossRef]
- Chao-Yang, G.; Peng, C.; Hai-Hong, Z. Roles of NLRP3 inflammasome in intervertebral disc degeneration. Osteoarthr. Cartil. 2021, 29, 793–801. [Google Scholar] [CrossRef]
- Elliott, E.I.; Sutterwala, F.S. Initiation and perpetuation of NLRP3 inflammasome activation and assembly. Immunol. Rev. 2015, 265, 35–52. [Google Scholar] [CrossRef] [PubMed]
- Rashidi, M.; Wicks, I.P.; Vince, J.E. Inflammasomes and Cell Death: Common Pathways in Microparticle Diseases. Trends Mol. Med. 2020, 26, 1003–1020. [Google Scholar] [CrossRef] [PubMed]
- Kennon, J.C.; Awad, M.E.; Chutkan, N.; DeVine, J.; Fulzele, S. Current insights on use of growth factors as therapy for Intervertebral Disc Degeneration. Biomol. Concepts 2018, 9, 43–52. [Google Scholar] [CrossRef] [PubMed]
- Krut, Z.; Pelled, G.; Gazit, D.; Gazit, Z. Stem Cells and Exosomes: New Therapies for Intervertebral Disc Degeneration. Cells 2021, 10, 2241. [Google Scholar] [CrossRef] [PubMed]
- Tessier, S.; Risbud, M.V. Understanding embryonic development for cell-based therapies of intervertebral disc degeneration: Toward an effort to treat disc degeneration subphenotypes. Dev. Dyn. 2021, 250, 302–317. [Google Scholar] [CrossRef]
- Wang, Y.; Yu, D.; Liu, Z.; Zhou, F.; Dai, J.; Wu, B.; Zhou, J.; Heng, B.C.; Zou, X.H.; Ouyang, H.; et al. Exosomes from embryonic mesenchymal stem cells alleviate osteoarthritis through balancing synthesis and degradation of cartilage extracellular matrix. Stem Cell Res. Ther. 2017, 8, 189. [Google Scholar] [CrossRef]
- Rosa, A.; Brivanlou, A.H. A regulatory circuitry comprised of miR-302 and the transcription factors OCT4 and NR2F2 regulates human embryonic stem cell differentiation. EMBO J. 2011, 30, 237–248. [Google Scholar] [CrossRef]
- Subramanyam, D.; Lamouille, S.; Judson, R.L.; Liu, J.Y.; Bucay, N.; Derynck, R.; Blelloch, R. Multiple targets of miR-302 and miR-372 promote reprogramming of human fibroblasts to induced pluripotent stem cells. Nat. Biotechnol. 2011, 29, 443–448. [Google Scholar] [CrossRef]
- Vidigal, J.A.; Ventura, A. Embryonic stem cell miRNAs and their roles in development and disease. Semin. Cancer Biol. 2012, 22, 428–436. [Google Scholar] [CrossRef]
- Chen, L.; Heikkinen, L.; Knott, K.E.; Liang, Y.C.; Wong, G. Evolutionary conservation and function of the human embryonic stem cell specific miR-302/367 cluster. Comp. Biochem. Physiol. D-Genom. Proteom. 2015, 16, 83–98. [Google Scholar] [CrossRef]
- Sun, Y.; Zhang, W.; Li, X. Induced pluripotent stem cell-derived mesenchymal stem cells deliver exogenous miR-105-5p via small extracellular vesicles to rejuvenate senescent nucleus pulposus cells and attenuate intervertebral disc degeneration. Stem Cell Res. Ther. 2021, 12, 286. [Google Scholar] [CrossRef]
- Issy, A.C.; Castania, V.; Castania, M.; Salmon, C.E.; Nogueira-Barbosa, M.H.; Bel, E.D.; Defino, H.L. Experimental model of intervertebral disc degeneration by needle puncture in Wistar rats. Braz. J. Med. Biol. Res. 2013, 46, 235–244. [Google Scholar] [CrossRef]
- Han, B.; Zhu, K.; Li, F.C.; Xiao, Y.X.; Feng, J.; Shi, Z.L.; Lin, M.; Wang, J.; Chen, Q.X. A simple disc degeneration model induced by percutaneous needle puncture in the rat tail. Spine 2008, 33, 1925–1934. [Google Scholar] [CrossRef]
- Liao, Z.; Luo, R.; Li, G.; Song, Y.; Zhan, S.; Zhao, K.; Hua, W.; Zhang, Y.; Wu, X.; Yang, C. Exosomes from mesenchymal stem cells modulate endoplasmic reticulum stress to protect against nucleus pulposus cell death and ameliorate intervertebral disc degeneration in vivo. Theranostics 2019, 9, 4084–4100. [Google Scholar] [CrossRef]
- Yang, S.H.; Hu, M.H.; Wu, C.C.; Chen, C.W.; Sun, Y.H.; Yang, K.C. CD24 expression indicates healthier phenotype and less tendency of cellular senescence in human nucleus pulposus cells. Artif. Cells Nanomed. Biotechnol. 2019, 47, 3021–3028. [Google Scholar] [CrossRef]
- Zhu, Q.; Ling, X.; Yang, Y.; Zhang, J.; Li, Q.; Niu, X.; Hu, G.; Chen, B.; Li, H.; Wang, Y.; et al. Embryonic Stem Cells-Derived Exosomes Endowed with Targeting Properties as Chemotherapeutics Delivery Vehicles for Glioblastoma Therapy. Adv. Sci. 2019, 6, 1801899. [Google Scholar] [CrossRef]
- Ji, M.L.; Jiang, H.; Zhang, X.J.; Shi, P.L.; Li, C.; Wu, H.; Wu, X.T.; Wang, Y.T.; Wang, C.; Lu, J. Preclinical development of a microRNA-based therapy for intervertebral disc degeneration. Nat. Commun. 2018, 9, 5051. [Google Scholar] [CrossRef]
- Chen, J.J.; Huang, J.F.; Du, W.X.; Tong, P.J. Expression and significance of MMP3 in synovium of knee joint at different stage in osteoarthritis patients. Asian Pac. J. Trop. Med. 2014, 7, 297–300. [Google Scholar] [CrossRef]
- Qin, R.; Dai, S.; Zhang, X.; Liu, H.; Zhou, B.; Zhou, P.; Hu, C. Danshen Attenuates Intervertebral Disc Degeneration via Antioxidation in SD Rats. Oxid. Med. Cell. Longev. 2020, 2020, 6660429. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Case No. | Age (Years) | Gender | Diagnosis | Disc Level | Pfirrmann Grading |
|---|---|---|---|---|---|
| 1 | 45 | M | Lumbar disc herniation | L4/5 | II |
| 2 | 52 | F | Lumbar disc herniation | L5/S1 | IV |
| 3 | 53 | M | Lumbar disc herniation | L4/5 | IV |
| 4 | 31 | M | Lumbar disc herniation | L4/5 | II |
| 5 | 66 | F | Lumbar disc herniation | L4/5 | III |
| 6 | 42 | F | Lumbar disc herniation | L4/5 | IV |
| 7 | 47 | F | Lumbar disc herniation | L4/5 | III |
| 8 | 24 | M | Lumbar disc herniation | L5/S1 | II |
| 9 | 43 | M | Lumbar disc herniation | L4/5 | III |
| 10 | 49 | M | Lumbar disc herniation | L4/5 | IV |
| 11 | 25 | M | Lumbar disc herniation | L3/4 | II |
| 12 | 40 | F | Lumbar disc herniation | L4/5 | IV |
| 13 | 59 | F | Lumbar disc herniation | L3/4 | III |
| 14 | 30 | M | Lumbar disc herniation | L4/5 | III |
| 15 | 67 | F | Lumbar disc herniation | L4/5 | IV |
| 16 | 52 | M | Lumbar disc herniation | L4/5 | III |
| 17 | 54 | F | Lumbar disc herniation | L4/5 | III |
| 18 | 72 | M | Lumbar disc herniation | L3/4 | III |
| 19 | 69 | F | Lumbar disc herniation | L4/5 | IV |
| 20 | 26 | M | Lumbar disc herniation | L5/S1 | II |
| Target Gene | Forward | Reverse |
|---|---|---|
| GAPDH | GGGAGCCAAAAGGGTCAT | GAGTCCTTCCACGATACCAA |
| NLRP3 | CAACCTCACGTCACACTGCT | TTTCAGACAACCCCAGGTTC |
| Caspase 1 | TTTCCGCAAGGTTCGATTTTCA | GGCATCTGCGCTCTACCATC |
| GSDMD | GACCCTAACACCTGGCAGAC | CACCTCAGTCACCACGTACAC |
| IL-1β | TTCGACACATGGGATAACGAGG | TTTTTGCTGTGAGTCCCGGAG |
| hsa-miR-302c mimics | UAAGUGCUUCCAUGUUUCAGUGG | AUUCACGAAGGUACAAAGUCACC |
| hsa-miR-302c mimics NC | UUUGUACUACACAAAAGUACUG | AAACAUGAUGUGUUUUCAUGAC |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Yu, Y.; Li, W.; Xian, T.; Tu, M.; Wu, H.; Zhang, J. Human Embryonic Stem-Cell-Derived Exosomes Repress NLRP3 Inflammasome to Alleviate Pyroptosis in Nucleus Pulposus Cells by Transmitting miR-302c. Int. J. Mol. Sci. 2023, 24, 7664. https://doi.org/10.3390/ijms24087664
Yu Y, Li W, Xian T, Tu M, Wu H, Zhang J. Human Embryonic Stem-Cell-Derived Exosomes Repress NLRP3 Inflammasome to Alleviate Pyroptosis in Nucleus Pulposus Cells by Transmitting miR-302c. International Journal of Molecular Sciences. 2023; 24(8):7664. https://doi.org/10.3390/ijms24087664
Chicago/Turabian StyleYu, Yawen, Wenting Li, Tinghui Xian, Mei Tu, Hao Wu, and Jiaqing Zhang. 2023. "Human Embryonic Stem-Cell-Derived Exosomes Repress NLRP3 Inflammasome to Alleviate Pyroptosis in Nucleus Pulposus Cells by Transmitting miR-302c" International Journal of Molecular Sciences 24, no. 8: 7664. https://doi.org/10.3390/ijms24087664