
LDL Affects the Immunomodulatory Response of Endothelial Cells by Modulation of the Promyelocytic Leukemia Protein (PML) Expression via PKC

Abstract
:1. Introduction
2. Results
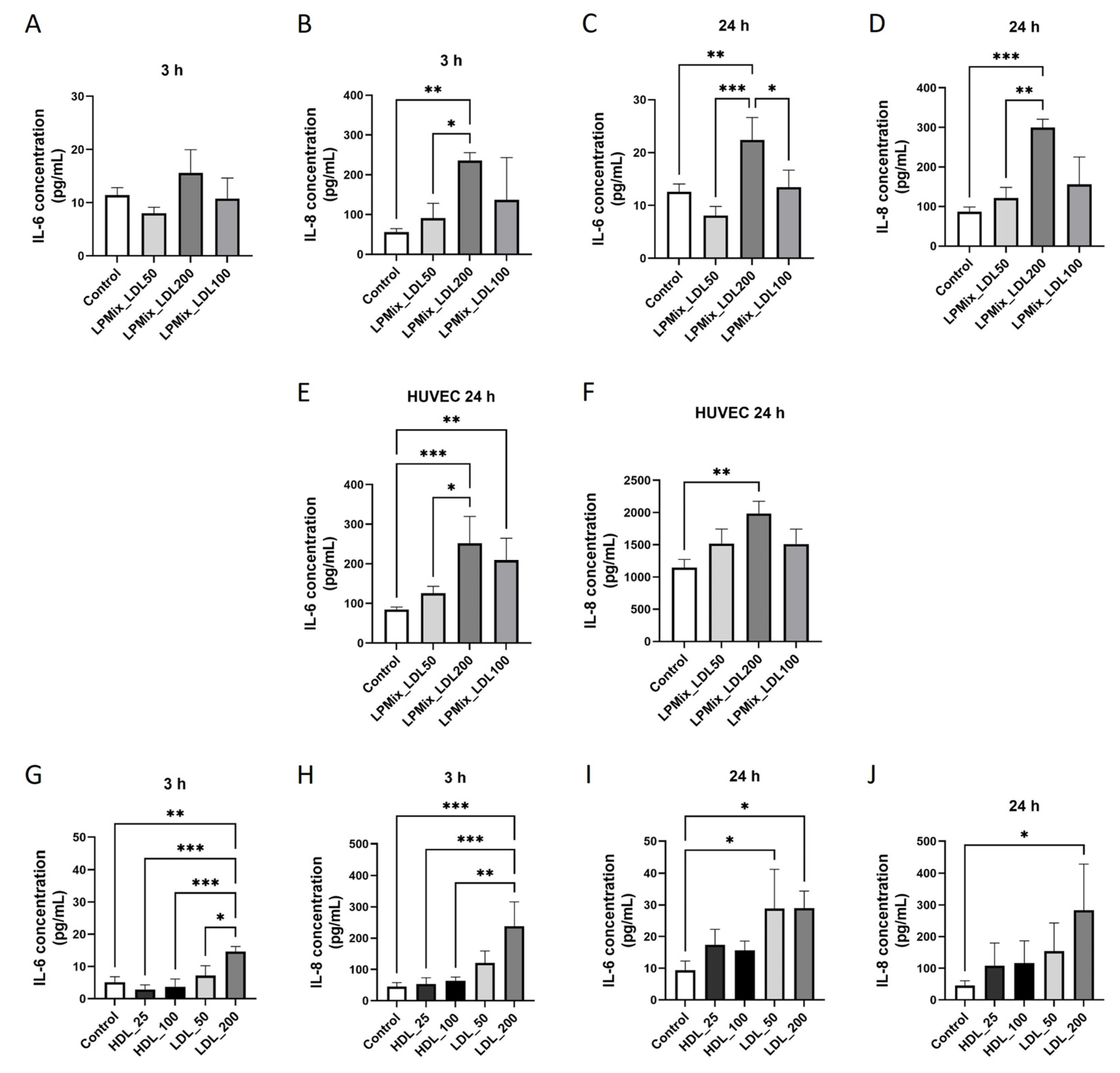
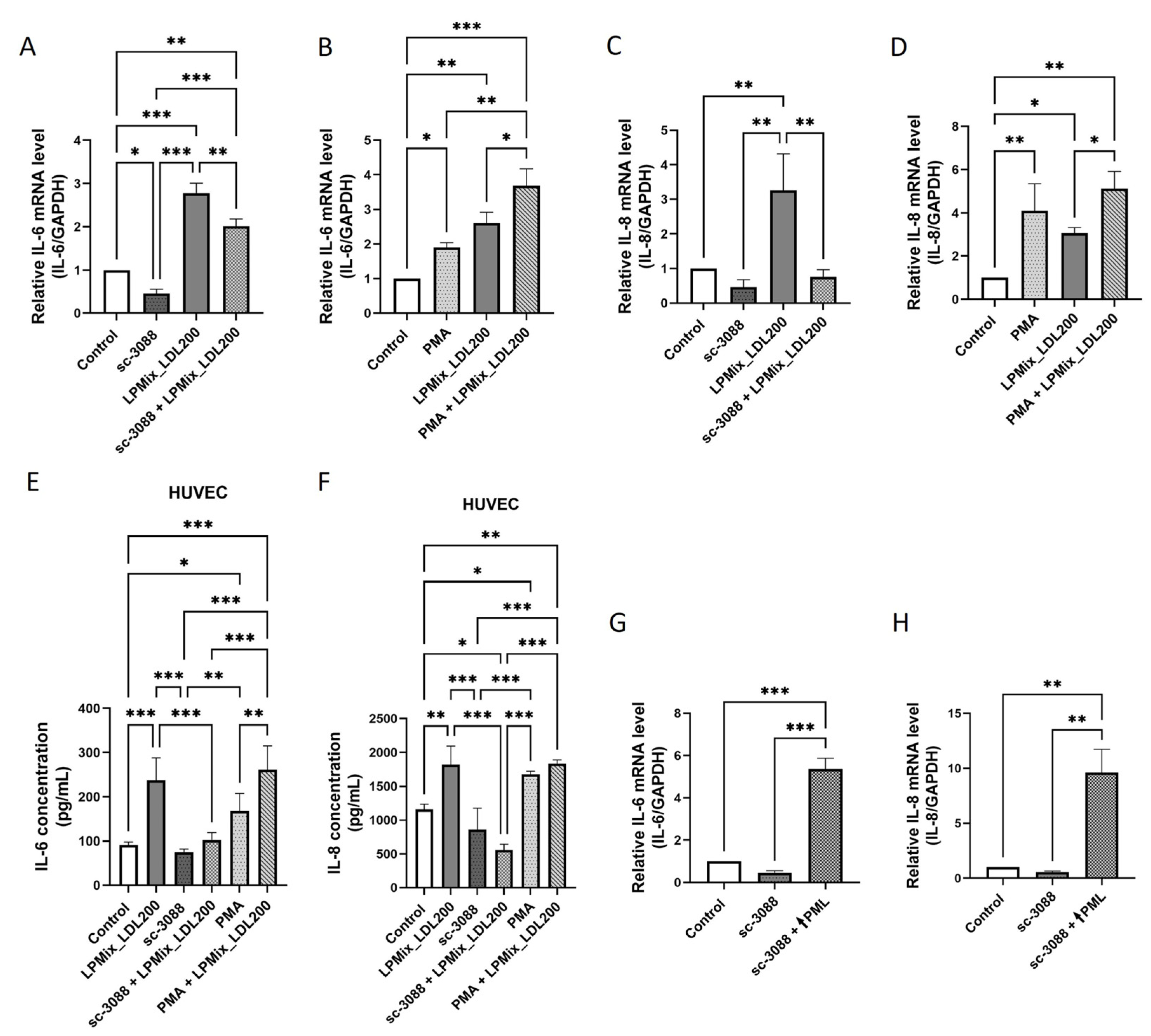
2.1. LDL Induces IL-6 and IL-8 Secretion in Endothelial Cells
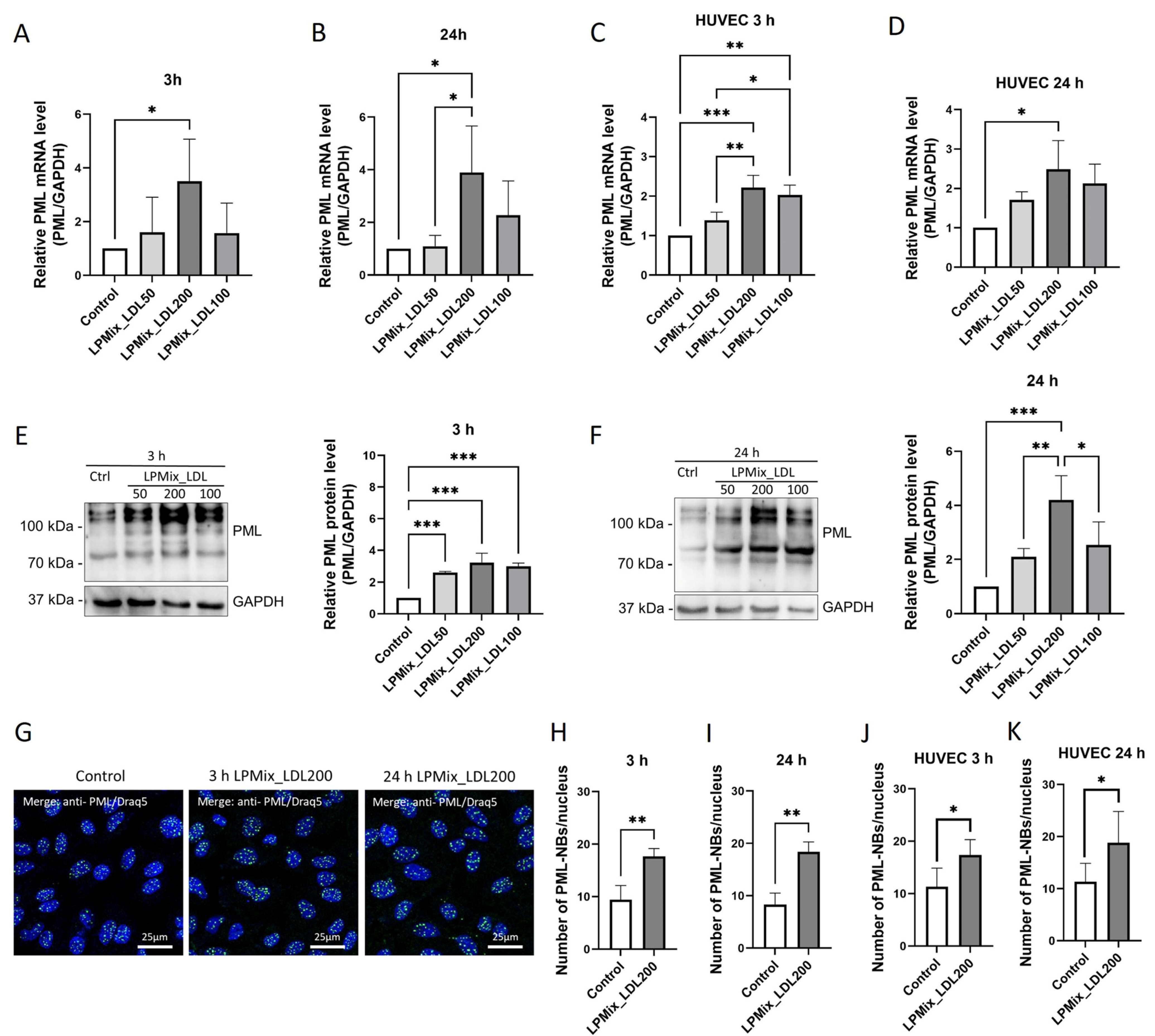
2.2. Stimulation of Endothelial Cells with High Doses of LDL Leads to Higher PML Expression
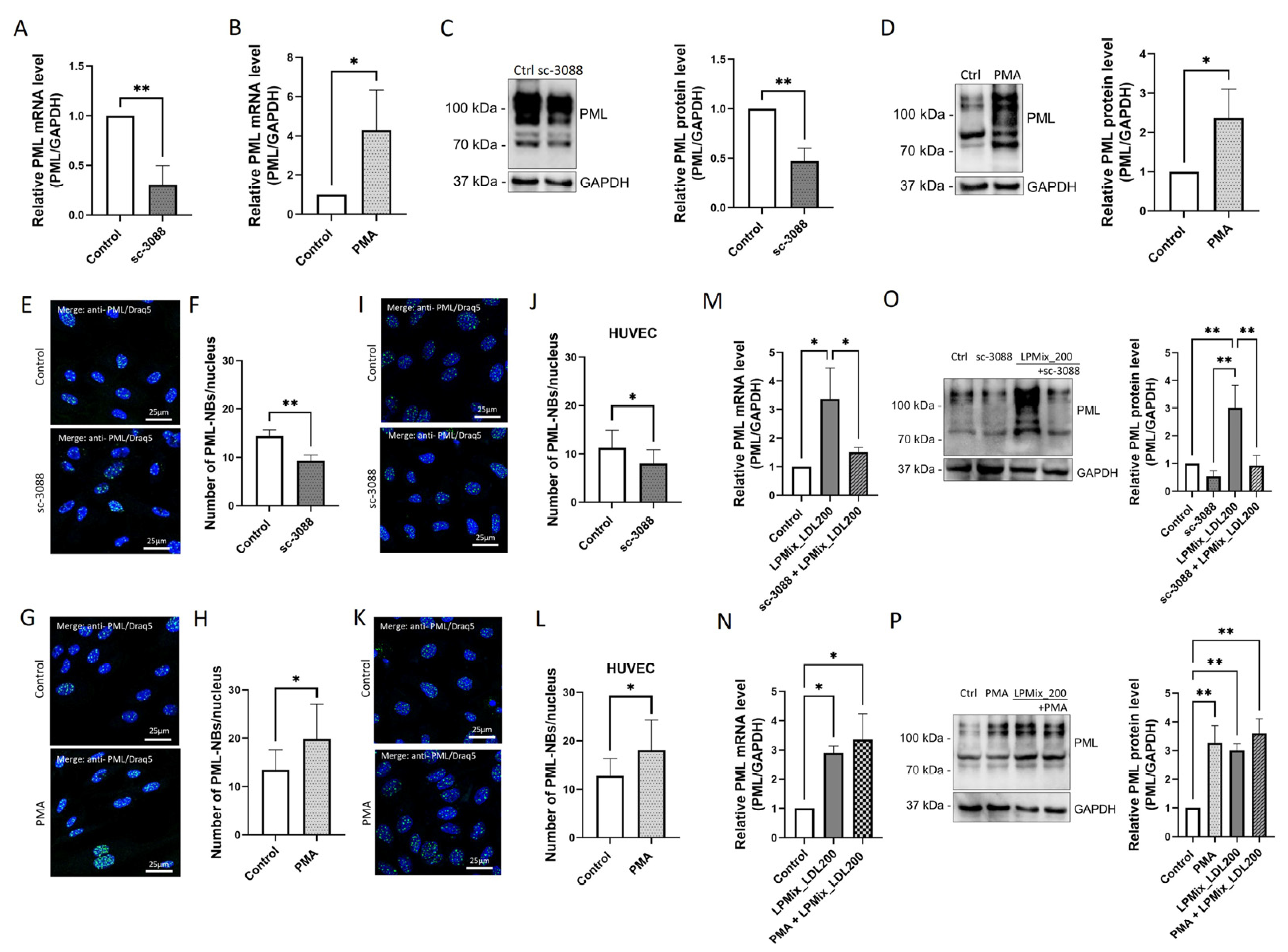
2.3. LDL Increases PML Levels in Endothelial Cells in a Dose-Dependent Manner
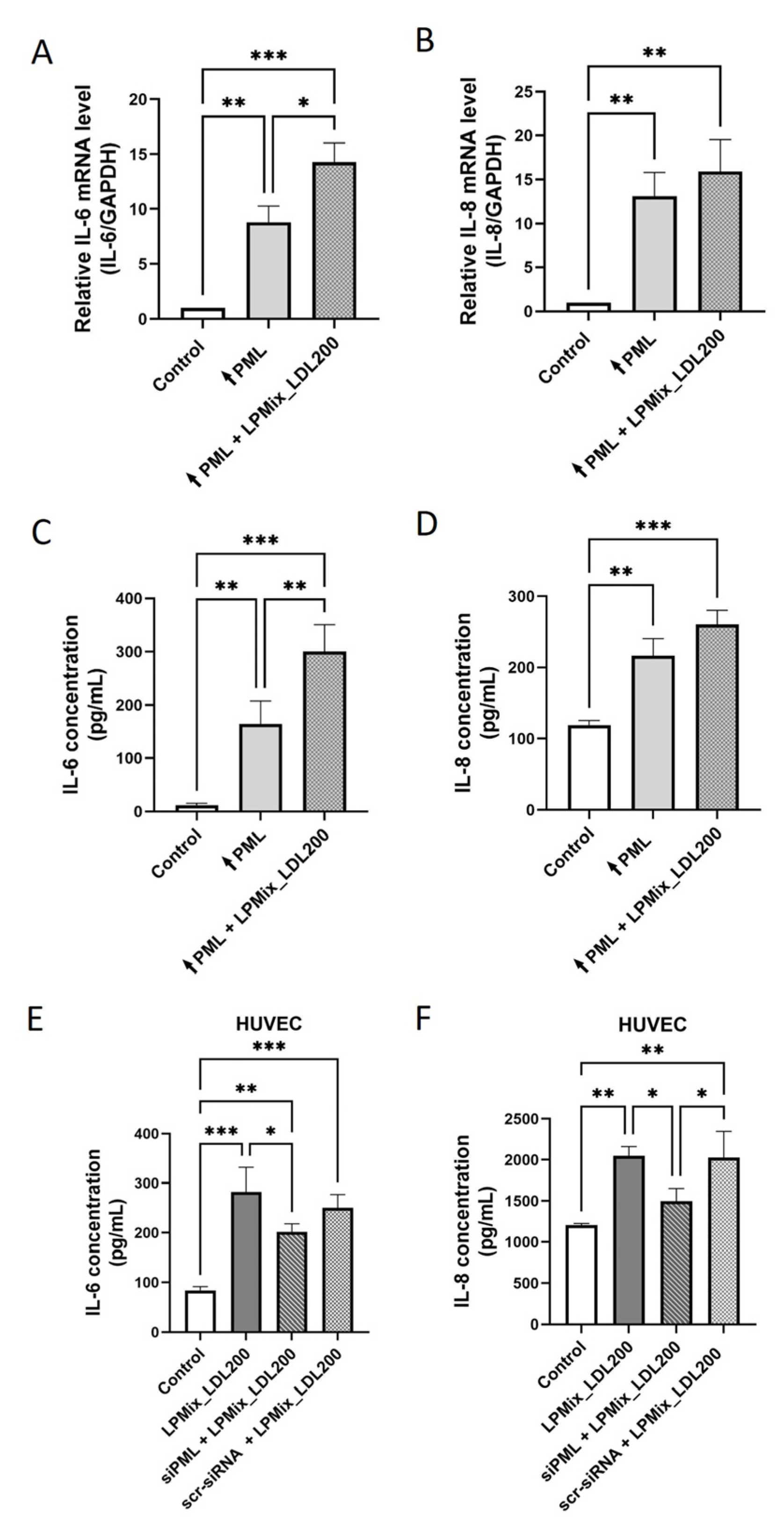
2.4. PML Mediates the LDL-Induced IL-6 and IL-8 Expression in ECs
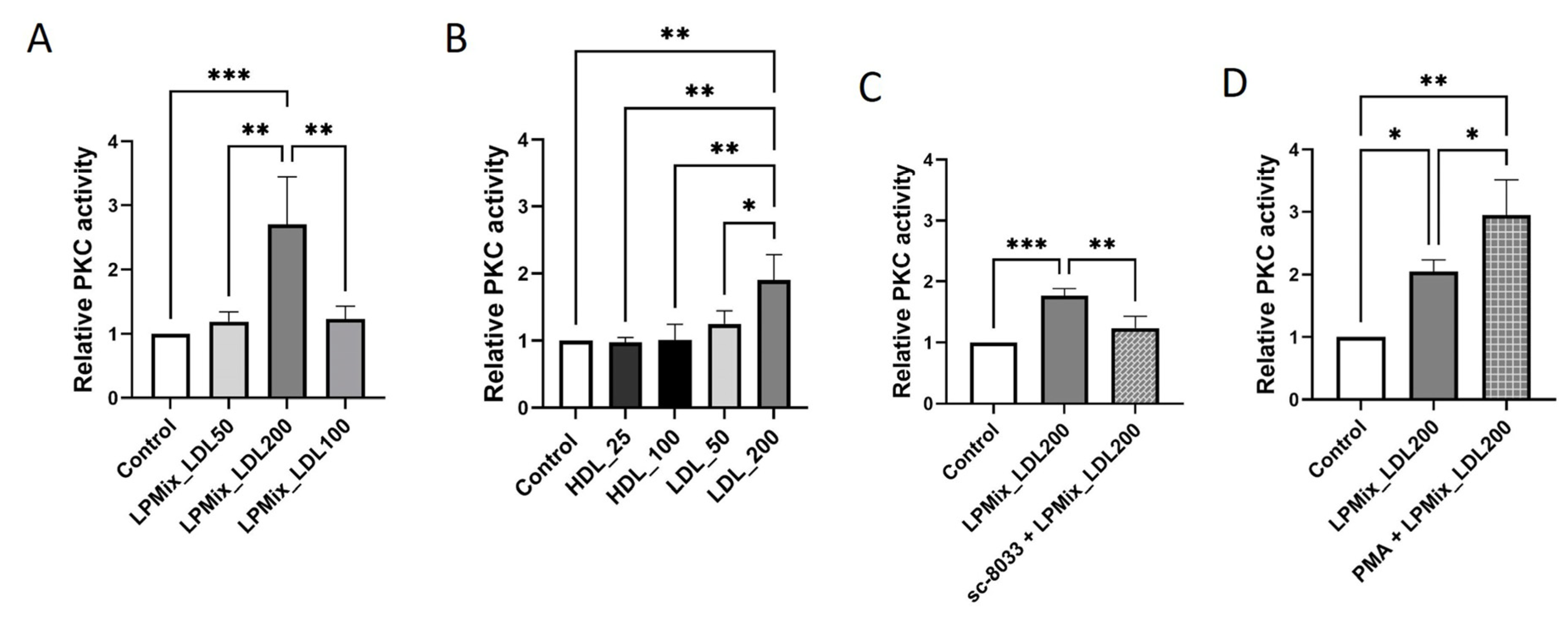
2.5. PKC Activity Is Higher in Endothelial Cells Exposed to Lipoproteins with High LDL Concentrations
2.6. LDL-Induced Upregulation of PML Is Mediated by PKC in Endothelial Cells
2.7. The LDL-Induced Upregulation of IL-6 and IL-8 Expression in Endothelial Cells Is Regulated by a PKC-Mediated Increased Expression of PML
3. Discussion
4. Materials and Methods
4.1. Lipoprotein Preparation
4.2. Cell Culture
4.3. Cell Stimulation Experiments with Lipoproteins
4.4. Modulation of Cellular PKC Activity by sc-3088 and PMA
4.5. Transfection with PML Expression Vector
4.6. Transfection with Small Interfering RNA (siRNA)
4.7. RNA Isolation, Reverse Transcription, and Real-Time PCR
4.8. Immunoblotting
4.9. ELISA Assays and Activity Assays
4.10. Immunofluorescence Analysis
4.11. Statistical Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Feingold, K.R. Introduction to lipids and lipoproteins. In Endotext; Feingold, K.R., Anawalt, B., Blackman, M.R., Boyce, A., Chrousos, G., Corpas, E., de Herder, W.W., Dhatariya, K., Dungan, K., Hofland, J., et al., Eds.; MDText.com, Inc.: South Dartmouth, MA, USA, 2000. [Google Scholar]
- Soppert, J.; Lehrke, M.; Marx, N.; Jankowski, J.; Noels, H. Lipoproteins and lipids in cardiovascular disease: From mechanistic insights to therapeutic targeting. Adv. Drug Deliv. Rev. 2020, 159, 4–33. [Google Scholar] [CrossRef] [PubMed]
- Authors/Task Force Members; ESC Committee for Practice Guidelines (CPG); ESC National Cardiac Societies. ESC/EAS guidelines for the management of dyslipidaemias: Lipid modification to reduce cardiovascular risk. Atherosclerosis 2019, 290, 140–205. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kottke, B.A.; Pineda, A.A.; Case, M.T.; Orsuzar, A.M.; Brzys, K.A. Hypercholesterolemia and atherosclerosis: Present and future therapy including LDL-apheresis. J. Clin. Apher. 1988, 4, 35–46. [Google Scholar] [CrossRef] [PubMed]
- Peng, J.; Luo, F.; Ruan, G.; Peng, R.; Li, X. Hypertriglyceridemia and atherosclerosis. Lipids Health Dis. 2017, 16, 233. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hermida, N.; Balligand, J.L. Low-density lipoprotein-cholesterol-induced endothelial dysfunction and oxidative stress: The role of statins. Antioxid. Redox Signal. 2014, 20, 1216–1237. [Google Scholar] [CrossRef] [PubMed]
- Boren, J.; Chapman, M.J.; Krauss, R.M.; Packard, C.J.; Bentzon, J.F.; Binder, C.J.; Daemen, M.J.; Demer, L.L.; Hegele, R.A.; Nicholls, S.J.; et al. Low-density lipoproteins cause atherosclerotic cardiovascular disease: Pathophysiological, genetic, and therapeutic insights: A consensus statement from the European Atherosclerosis Society Consensus Panel. Eur. Heart J. 2020, 41, 2313–2330. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Millan, J.; Pinto, X.; Munoz, A.; Zuniga, M.; Rubies-Prat, J.; Pallardo, L.F.; Masana, L.; Mangas, A.; Hernandez-Mijares, A.; Gonzalez-Santos, P.; et al. Lipoprotein ratios: Physiological significance and clinical usefulness in cardiovascular prevention. Vasc. Health Risk Manag. 2009, 5, 757–765. [Google Scholar]
- Avraham-Davidi, I.; Grunspan, M.; Yaniv, K. Lipid signaling in the endothelium. Exp. Cell Res. 2013, 319, 1298–1305. [Google Scholar] [CrossRef]
- Mineo, C. Lipoprotein receptor signalling in atherosclerosis. Cardiovasc. Res. 2020, 116, 1254–1274. [Google Scholar] [CrossRef]
- Lubrano, V.; Gabriele, M.; Puntoni, M.R.; Longo, V.; Pucci, L. Relationship among IL-6, LDL cholesterol and lipid peroxidation. Cell. Mol. Biol. Lett. 2015, 20, 310–322. [Google Scholar] [CrossRef]
- Hsu, K.S.; Kao, H.Y. PML: Regulation and multifaceted function beyond tumor suppression. Cell Biosci. 2018, 8, 5. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lunardi, A.; Gaboli, M.; Giorgio, M.; Rivi, R.; Bygrave, A.; Antoniou, M.; Drabek, D.; Dzierzak, E.; Fagioli, M.; Salmena, L.; et al. A role for PML in innate immunity. Genes Cancer 2011, 2, 10–19. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- de Thé, H.; Lavau, C.; Marchio, A.; Chomienne, C.; Degos, L.; Dejean, A. The PML-RAR alpha fusion mRNA generated by the t(15;17) translocation in acute promyelocytic leukemia encodes a functionally altered RAR. Cell 1991, 66, 675–684. [Google Scholar] [CrossRef] [PubMed]
- Terris, B.; Baldin, V.; Dubois, S.; Degott, C.; Flejou, J.F.; Henin, D.; Dejean, A. PML nuclear bodies are general targets for inflammation and cell proliferation. Cancer Res. 1995, 55, 1590–1597. [Google Scholar] [PubMed]
- Karle, W.; Becker, S.; Stenzel, P.; Knosalla, C.; Siegel, G.; Baum, O.; Zakrzewicz, A.; Berkholz, J. Promyelocytic leukemia protein promotes the phenotypic switch of smooth muscle cells in atherosclerotic plaques of human coronary arteries. Clin. Sci. 2021, 135, 887–905. [Google Scholar] [CrossRef] [PubMed]
- Ishov, A.M.; Sotnikov, A.G.; Negorev, D.; Vladimirova, O.V.; Neff, N.; Kamitani, T.; Yeh, E.T.; Strauss, J.F., 3rd; Maul, G.G. PML is critical for ND10 formation and recruits the PML-interacting protein daxx to this nuclear structure when modified by SUMO-1. J. Cell Biol. 1999, 147, 221–234. [Google Scholar] [CrossRef] [Green Version]
- Lallemand-Breitenbach, V.; de Thé, H. PML nuclear bodies. Cold Spring Harb. Perspect. Biol. 2010, 2, a000661. [Google Scholar] [CrossRef]
- Van Damme, E.; Laukens, K.; Dang, T.H.; Van Ostade, X. A manually curated network of the PML nuclear body interactome reveals an important role for PML-NBs in SUMOylation dynamics. Int. J. Biol. Sci. 2010, 6, 51–67. [Google Scholar] [CrossRef]
- Scherer, M.; Stamminger, T. Emerging role of PML nuclear bodies in innate immune signaling. J. Virol. 2016, 90, 5850–5854. [Google Scholar] [CrossRef] [Green Version]
- Allen, S.; Khan, S.; Al-Mohanna, F.; Batten, P.; Yacoub, M. Native low density lipoprotein-induced calcium transients trigger VCAM-1 and E-selectin expression in cultured human vascular endothelial cells. J. Clin. Investig. 1998, 101, 1064–1075. [Google Scholar] [CrossRef] [Green Version]
- Han, K.H.; Tangirala, R.K.; Green, S.R.; Quehenberger, O. Chemokine receptor CCR2 expression and monocyte chemoattractant protein-1-mediated chemotaxis in human monocytes. A regulatory role for plasma LDL. Arterioscler. Thromb. Vasc. Biol. 1998, 18, 1983–1991. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Navab, M.; Imes, S.S.; Hama, S.Y.; Hough, G.P.; Ross, L.A.; Bork, R.W.; Valente, A.J.; Berliner, J.A.; Drinkwater, D.C.; Laks, H.; et al. Monocyte transmigration induced by modification of low density lipoprotein in cocultures of human aortic wall cells is due to induction of monocyte chemotactic protein 1 synthesis and is abolished by high density lipoprotein. J. Clin. Investig. 1991, 88, 2039–2046. [Google Scholar] [CrossRef] [PubMed]
- Zhu, Y.; Lin, J.H.; Liao, H.L.; Friedli, O., Jr.; Verna, L.; Marten, N.W.; Straus, D.S.; Stemerman, M.B. LDL induces transcription factor activator protein-1 in human endothelial cells. Arterioscler. Thromb. Vasc. Biol. 1998, 18, 473–480. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Al-Sharea, A.; Lee, M.K.; Moore, X.L.; Fang, L.; Sviridov, D.; Chin-Dusting, J.; Andrews, K.L.; Murphy, A.J. Native LDL promotes differentiation of human monocytes to macrophages with an inflammatory phenotype. Thromb. Haemost. 2016, 115, 762–772. [Google Scholar] [CrossRef] [PubMed]
- Massy, Z.A.; Kim, Y.; Guijarro, C.; Kasiske, B.L.; Keane, W.F.; O’Donnell, M.P. Low-density lipoprotein-induced expression of interleukin-6, a marker of human mesangial cell inflammation: Effects of oxidation and modulation by lovastatin. Biochem. Biophys. Res. Commun. 2000, 267, 536–540. [Google Scholar] [CrossRef] [PubMed]
- Lo, Y.H.; Huang, Y.W.; Wu, Y.H.; Tsai, C.S.; Lin, Y.C.; Mo, S.T.; Kuo, W.C.; Chuang, Y.T.; Jiang, S.T.; Shih, H.M.; et al. Selective inhibition of the NLRP3 inflammasome by targeting to promyelocytic leukemia protein in mouse and human. Blood 2013, 121, 3185–3194. [Google Scholar] [CrossRef] [Green Version]
- Foey, A.D.; Brennan, F.M. Conventional protein kinase C and atypical protein kinase Czeta differentially regulate macrophage production of tumour necrosis factor-alpha and interleukin-10. Immunology 2004, 112, 44–53. [Google Scholar] [CrossRef]
- Kontny, E.; Ziolkowska, M.; Ryzewska, A.; Maslinski, W. Protein kinase c-dependent pathway is critical for the production of pro-inflammatory cytokines (TNF-alpha, IL-1beta, IL-6). Cytokine 1999, 11, 839–848. [Google Scholar] [CrossRef]
- Zhang, X.; Sessa, W.C.; Fernandez-Hernando, C. Endothelial transcytosis of lipoproteins in atherosclerosis. Front. Cardiovasc. Med. 2018, 5, 130. [Google Scholar] [CrossRef] [Green Version]
- Lefkowitz, R.J. The superfamily of heptahelical receptors. Nat. Cell Biol. 2000, 2, E133–E136. [Google Scholar] [CrossRef]
- Kawata, K.; Kubota, S.; Eguchi, T.; Moritani, N.H.; Shimo, T.; Kondo, S.; Nishida, T.; Minagi, S.; Takigawa, M. Role of the low-density lipoprotein receptor-related protein-1 in regulation of chondrocyte differentiation. J. Cell. Physiol. 2010, 222, 138–148. [Google Scholar] [CrossRef] [PubMed]
- Ren, S.; Shatadal, S.; Shen, G.X. Protein kinase C-beta mediates lipoprotein-induced generation of PAI-1 from vascular endothelial cells. Am. J. Physiol. Endocrinol. Metab. 2000, 278, E656–E662. [Google Scholar] [CrossRef] [PubMed]
- Way, K.J.; Chou, E.; King, G.L. Identification of PKC-isoform-specific biological actions using pharmacological approaches. Trends Pharmacol. Sci. 2000, 21, 181–187. [Google Scholar] [CrossRef]
- Sinclear, C.K.; Maruyama, J.; Nagashima, S.; Arimoto-Matsuzaki, K.; Kuleape, J.A.; Iwasa, H.; Nishina, H.; Hata, Y. Protein kinase Calpha activation switches YAP1 from TEAD-mediated signaling to p73-mediated signaling. Cancer Sci. 2022, 113, 1305–1320. [Google Scholar] [CrossRef]
- Hofmann, J. The potential for isoenzyme-selective modulation of protein kinase C. FASEB J. 1997, 11, 649–669. [Google Scholar] [CrossRef] [PubMed]
- Castagna, M.; Takai, Y.; Kaibuchi, K.; Sano, K.; Kikkawa, U.; Nishizuka, Y. Direct activation of calcium-activated, phospholipid-dependent protein kinase by tumor-promoting phorbol esters. J. Biol. Chem. 1982, 257, 7847–7851. [Google Scholar] [CrossRef]
- Kuhns, D.B.; Young, H.A.; Gallin, E.K.; Gallin, J.I. Ca2+-dependent production and release of IL-8 in human neutrophils. J. Immunol. 1998, 161, 4332–4339. [Google Scholar] [CrossRef] [PubMed]
- Volk, T.; Hensel, M.; Mading, K.; Egerer, K.; Kox, W.J. Intracellular Ca2+ dependence of nitric oxide mediated enhancement of interleukin-8 secretion in human endothelial cells. FEBS Lett. 1997, 415, 169–172. [Google Scholar] [CrossRef] [Green Version]
- Norata, G.D.; Pirillo, A.; Pellegatta, F.; Inoue, H.; Catapano, A.L. Native LDL and oxidized LDL modulate cyclooxygenase-2 expression in HUVECs through a p38-MAPK, NF-kappaB, CRE dependent pathway and affect PGE2 synthesis. Int. J. Mol. Med. 2004, 14, 353–359. [Google Scholar] [CrossRef]
- Trejo, J.; Massamiri, T.; Deng, T.; Dewji, N.N.; Bayney, R.M.; Brown, J.H. A direct role for protein kinase C and the transcription factor Jun/AP-1 in the regulation of the Alzheimer’s beta-amyloid precursor protein gene. J. Biol. Chem. 1994, 269, 21682–21690. [Google Scholar] [CrossRef]
- Diaz-Meco, M.T.; Moscat, J. The atypical PKCs in inflammation: NF-kappaB and beyond. Immunol. Rev. 2012, 246, 154–167. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ahmed, A.; Wan, X.; Mitxitorena, I.; Lindsay, A.J.; Paolo Pandolfi, P.; McCaffrey, M.W.; Keeshan, K.; Chen, Y.H.; Carmody, R.J. Regulation of NF-kappaB by PML and PML-RARalpha. Sci. Rep. 2017, 7, 44539. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wu, W.S.; Xu, Z.X.; Hittelman, W.N.; Salomoni, P.; Pandolfi, P.P.; Chang, K.S. Promyelocytic leukemia protein sensitizes tumor necrosis factor alpha-induced apoptosis by inhibiting the NF-kappaB survival pathway. J. Biol. Chem. 2003, 278, 12294–12304. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dendorfer, U.; Oettgen, P.; Libermann, T.A. Multiple regulatory elements in the interleukin-6 gene mediate induction by prostaglandins, cyclic AMP, and lipopolysaccharide. Mol. Cell. Biol. 1994, 14, 4443–4454. [Google Scholar] [CrossRef]
- Mukaida, N.; Mahe, Y.; Matsushima, K. Cooperative interaction of nuclear factor-kappa B- and cis-regulatory enhancer binding protein-like factor binding elements in activating the interleukin-8 gene by pro-inflammatory cytokines. J. Biol. Chem. 1990, 265, 21128–21133. [Google Scholar] [CrossRef]
- Libermann, T.A.; Baltimore, D. Activation of interleukin-6 gene expression through the NF-kappa B transcription factor. Mol. Cell. Biol. 1990, 10, 2327–2334. [Google Scholar] [CrossRef] [Green Version]
- Pirillo, A.; Norata, G.D.; Catapano, A.L. LOX-1, OxLDL, and atherosclerosis. Mediat. Inflamm. 2013, 2013, 152786. [Google Scholar] [CrossRef] [Green Version]
- Claus, R.; Fyrnys, B.; Deigner, H.P.; Wolf, G. Oxidized low-density lipoprotein stimulates protein kinase C (PKC) and induces expression of PKC-isotypes via prostaglandin-H-synthase in P388D1 macrophage-like cells. Biochemistry 1996, 35, 4911–4922. [Google Scholar] [CrossRef]
- Maziere, C.; Maziere, J.C. Activation of transcription factors and gene expression by oxidized low-density lipoprotein. Free Radic. Biol. Med. 2009, 46, 127–137. [Google Scholar] [CrossRef]
- N’Guessan, P.D.; Riediger, F.; Vardarova, K.; Scharf, S.; Eitel, J.; Opitz, B.; Slevogt, H.; Weichert, W.; Hocke, A.C.; Schmeck, B.; et al. Statins control oxidized LDL-mediated histone modifications and gene expression in cultured human endothelial cells. Arterioscler. Thromb. Vasc. Biol. 2009, 29, 380–386. [Google Scholar] [CrossRef] [Green Version]
- Sohrabi, Y.; Lagache, S.M.M.; Voges, V.C.; Semo, D.; Sonntag, G.; Hanemann, I.; Kahles, F.; Waltenberger, J.; Findeisen, H.M. OxLDL-mediated immunologic memory in endothelial cells. J. Mol. Cell. Cardiol. 2020, 146, 121–132. [Google Scholar] [CrossRef]
- Claise, C.; Edeas, M.; Chalas, J.; Cockx, A.; Abella, A.; Capel, L.; Lindenbaum, A. Oxidized low-density lipoprotein induces the production of interleukin-8 by endothelial cells. FEBS Lett. 1996, 398, 223–227. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ryoo, S.W.; Kim, D.U.; Won, M.; Chung, K.S.; Jang, Y.J.; Oh, G.T.; Park, S.K.; Maeng, P.J.; Yoo, H.S.; Hoe, K.L. Native LDL induces interleukin-8 expression via H2O2, p38 Kinase, and activator protein-1 in human aortic smooth muscle cells. Cardiovasc. Res. 2004, 62, 185–193. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Brasier, A.R. The nuclear factor-kappaB-interleukin-6 signalling pathway mediating vascular inflammation. Cardiovasc. Res. 2010, 86, 211–218. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kempe, S.; Kestler, H.; Lasar, A.; Wirth, T. NF-kappaB controls the global pro-inflammatory response in endothelial cells: Evidence for the regulation of a pro-atherogenic program. Nucleic Acids Res. 2005, 33, 5308–5319. [Google Scholar] [CrossRef]
- Yu, Y.; De Waele, C.; Chadee, K. Calcium-dependent interleukin-8 gene expression in T84 human colonic epithelial cells. Inflamm. Res. 2001, 50, 220–226. [Google Scholar] [CrossRef] [PubMed]
- Ko, Y.; Totzke, G.; Seewald, S.; Schmitz, U.; Schiermeyer, B.; Meyer zu Brickwedde, M.K.; Vetter, H.; Sachinidis, A. Native low-density lipoprotein (LDL) induces the expression of the early growth response gene-1 in human umbilical arterial endothelial cells. Eur. J. Cell Biol. 1995, 68, 306–312. [Google Scholar]
- Schaefer, H.I.; Hold, K.M.; Egas-Kenniphaas, J.M.; van der Laarse, A. Intracellular calcium signalling after binding of low-density lipoprotein to confluent and nonconfluent cultures of an endothelial cell line, EA.hy 926. Cell Calcium 1993, 14, 507–516. [Google Scholar] [CrossRef]
- Mudau, M.; Genis, A.; Lochner, A.; Strijdom, H. Endothelial dysfunction: The early predictor of atherosclerosis. Cardiovasc. J. Afr. 2012, 23, 222–231. [Google Scholar] [CrossRef]
- Chisari, F.V.; Curtiss, L.K.; Jensen, F.C. Physiologic concentrations of normal human plasma lipoproteins inhibit the immortalization of peripheral B lymphocytes by the Epstein-Barr virus. J. Clin. Investig. 1981, 68, 329–336. [Google Scholar] [CrossRef]
- Leonhardt, W.; Pietzsch, J.; Julius, U.; Hanefeld, M. Recovery of cholesterol and triacylglycerol in very-fast ultracentrifugation of human lipoproteins in a large range of concentrations. Eur. J. Clin. Chem. Clin. Biochem. 1994, 32, 929–933. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Becker, S.; Steinemann, G.; Karle, W.; Roos, K.; Liem, C.H.; Muralikumar, S.; Volkamer, A.; Munz, B.; Zakrzewicz, A.; Berkholz, J. Stability of Smyd1 in endothelial cells is controlled by PML-dependent SUMOylation upon cytokine stimulation. Biochem. J. 2021, 478, 217–234. [Google Scholar] [CrossRef] [PubMed]
- Shamloul, A.; Steinemann, G.; Roos, K.; Liem, C.H.; Bernd, J.; Braun, T.; Zakrzewicz, A.; Berkholz, J. The Methyltransferase Smyd1 Mediates LPS-Triggered Up-Regulation of IL-6 in Endothelial Cells. Cells 2021, 10, 3515. [Google Scholar] [CrossRef] [PubMed]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Template | Forward Primer | Reverse Primer | Product Size | Annealing Temperature |
|---|---|---|---|---|
| GAPDH | 5′-ATG ACC TTG CCC ACA GCC TT-3′ | 5′-AAC TGC TTA GCA CCC CTG GC-3′ | 200 bp | 60 °C |
| IL-6 | 5′-TGC CAG CCT GCT GAC GAA G-3′ | 5′-AGC TGC GCA GAA TGA GAT GAG-3′ | 77 bp | 58 °C |
| IL-8 | 5′-ATG ACT TCC AAG CTG GCC GTG GC-3′ | 5′-TCT CAG CCC TCT TCA AAA ACT TCT C-3′ | 292 bp | 58 °C |
| PML | 5′-CCG CAA GAC CAA CAA CAT CTT-3′ | 5′-CAG CGG CTT GGA ACA TCC T-3′ | 91 bp | 58 °C |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Roos, K.; Berkholz, J. LDL Affects the Immunomodulatory Response of Endothelial Cells by Modulation of the Promyelocytic Leukemia Protein (PML) Expression via PKC. Int. J. Mol. Sci. 2023, 24, 7306. https://doi.org/10.3390/ijms24087306
Roos K, Berkholz J. LDL Affects the Immunomodulatory Response of Endothelial Cells by Modulation of the Promyelocytic Leukemia Protein (PML) Expression via PKC. International Journal of Molecular Sciences. 2023; 24(8):7306. https://doi.org/10.3390/ijms24087306
Chicago/Turabian StyleRoos, Kerrin, and Janine Berkholz. 2023. "LDL Affects the Immunomodulatory Response of Endothelial Cells by Modulation of the Promyelocytic Leukemia Protein (PML) Expression via PKC" International Journal of Molecular Sciences 24, no. 8: 7306. https://doi.org/10.3390/ijms24087306