
The Expression Levels of MicroRNAs Differentially Expressed in Sudden Sensorineural Hearing Loss Patients’ Serum Are Unchanged for up to 12 Months after Hearing Loss Onset


Abstract
:1. Introduction
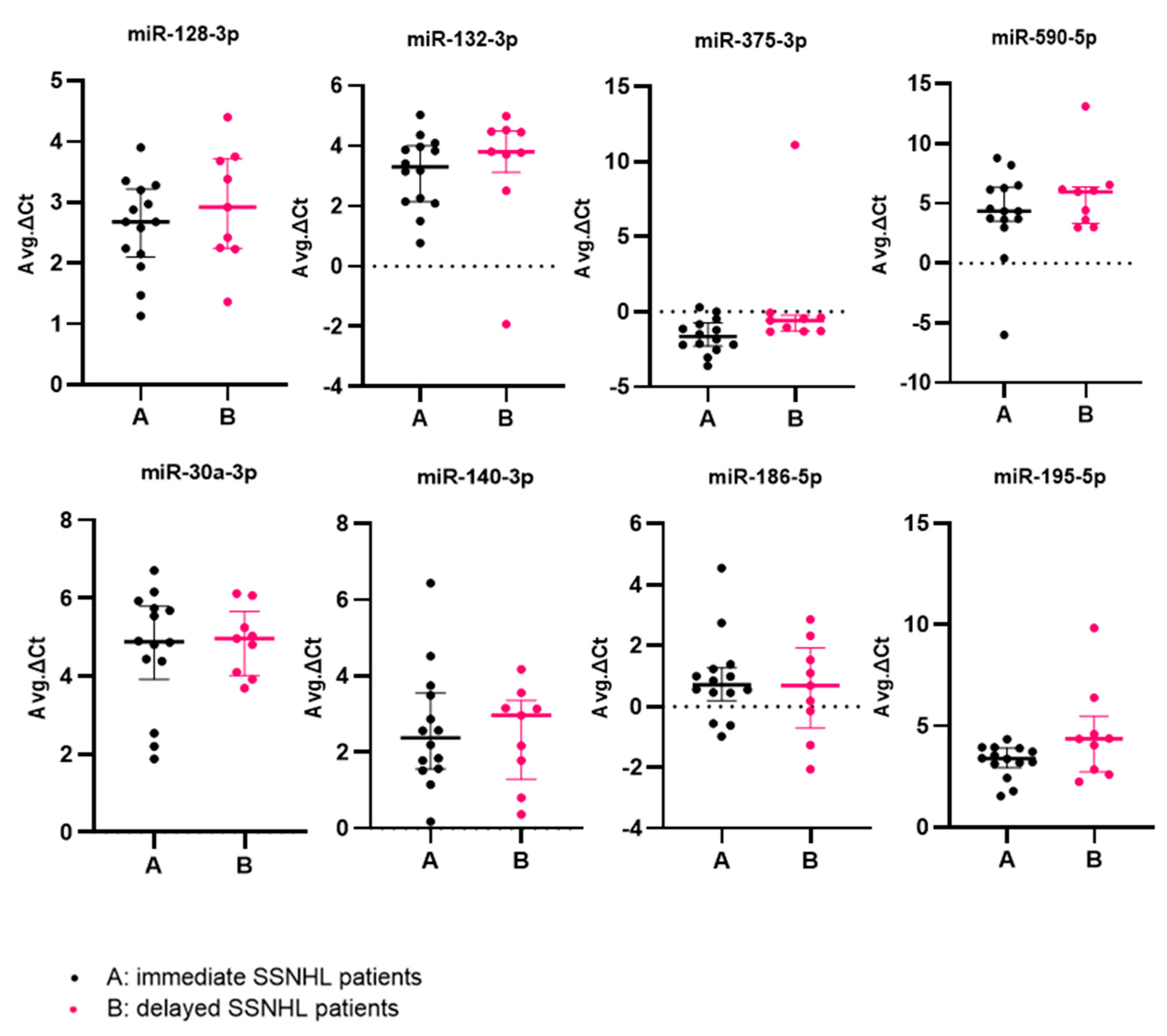
2. Results
3. Discussion
4. Materials and Methods
4.1. Study Populations and Sampling
4.2. Clinical Examination and Patient Recruitment
4.3. Pure Tone Audiometry
4.4. Blood Collection and MicroRNA Extraction
4.5. MicroRNA Reverse Transcription
4.6. MicroRNA Real-Time PCR
4.7. Pure Tone Audiometry
4.8. Statistical Analysis
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Chen, L.; Heikkinen, L.; Wang, C.; Yang, Y.; Sun, H.; Wong, G. Trends in the Development of MiRNA Bioinformatics Tools. Brief. Bioinform. 2019, 20, 1836–1852. [Google Scholar] [CrossRef] [PubMed]
- Mitchell, P.S.; Parkin, R.K.; Kroh, E.M.; Fritz, B.R.; Wyman, S.K.; Pogosova-Agadjanyan, E.L.; Peterson, A.; Noteboom, J.; O’Briant, K.C.; Allen, A.; et al. Circulating MicroRNAs as Stable Blood-Based Markers for Cancer Detection. Proc. Natl. Acad. Sci. USA 2008, 105, 10513–10518. [Google Scholar] [CrossRef] [PubMed]
- Rothenberg, M.; Lu, T. MicroRNA. J Allergy Clin Immunol 2018, 141, 1202–1207. [Google Scholar]
- Ferrante, M.; Conti, G.O. Environment and Neurodegenerative Diseases: An Update on MiRNA Role. Microrna 2017, 6, 157–165. [Google Scholar] [CrossRef] [PubMed]
- Hanson, E.K.; Lubenow, H.; Ballantyne, J. Identification of Forensically Relevant Body Fluids Using a Panel of Differentially Expressed MicroRNAs. Anal. Biochem. 2009, 387, 303–314. [Google Scholar] [CrossRef]
- Wu, L.; Fan, J.; Belasco, J.G. MicroRNAs Direct Rapid Deadenylation of MRNA. Proc. Natl. Acad. Sci. USA 2006, 103, 4034–4039. [Google Scholar] [CrossRef] [PubMed]
- Zhao, Y.; Ransom, J.F.; Li, A.; Vedantham, V.; von Drehle, M.; Muth, A.N.; Tsuchihashi, T.; McManus, M.T.; Schwartz, R.J.; Srivastava, D. Dysregulation of Cardiogenesis, Cardiac Conduction, and Cell Cycle in Mice Lacking MiRNA-1-2. Cell 2007, 129, 303–317. [Google Scholar] [CrossRef] [PubMed]
- Cortez, M.A.; Bueso-Ramos, C.; Ferdin, J.; Lopez-Berestein, G.; Sood, A.K.; Calin, G.A. MicroRNAs in Body Fluids—The Mix of Hormones and Biomarkers. Nat. Rev. Clin. Oncol. 2011, 8, 467–477. [Google Scholar] [CrossRef]
- Etheridge, A.; Lee, I.; Hood, L.; Galas, D.; Wang, K. Extracellular MicroRNA: A New Source of Biomarkers. Mutat. Res. Mol. Mech. Mutagen. 2011, 717, 85–90. [Google Scholar] [CrossRef] [PubMed]
- Kosaka, N.; Iguchi, H.; Yoshioka, Y.; Takeshita, F.; Matsuki, Y.; Ochiya, T. Secretory Mechanisms and Intercellular Transfer of MicroRNAs in Living Cells*♦. J. Biol. Chem. 2010, 285, 17442–17452. [Google Scholar] [CrossRef]
- Vickers, K.; Palmisano, B.; Shoucri, B.; Shamburek, R.; Remaley, A. MicroRNAs Control of HDL Metabolism and Function. Nat. Cell Biol. 2011, 13, 423–433. [Google Scholar] [CrossRef] [PubMed]
- Wang, K.; Zhang, S.; Weber, J.; Baxter, D.; Galas, D.J. Export of MicroRNAs and MicroRNA-Protective Protein by Mammalian Cells. Nucleic Acids Res. 2010, 38, 7248–7259. [Google Scholar] [CrossRef] [PubMed]
- Chen, X.; Ba, Y.; Ma, L.; Cai, X.; Yin, Y.; Wang, K.; Guo, J.; Zhang, Y.; Chen, J.; Guo, X.; et al. Characterization of MicroRNAs in Serum: A Novel Class of Biomarkers for Diagnosis of Cancer and Other Diseases. Cell Res. 2008, 18, 997–1006. [Google Scholar] [CrossRef]
- Gibbings, D.J.; Ciaudo, C.; Erhardt, M.; Voinnet, O. Multivesicular Bodies Associate with Components of MiRNA Effector Complexes and Modulate MiRNA Activity. Nat. Cell Biol. 2009, 11, 1143–1149. [Google Scholar] [CrossRef]
- Valadi, H.; Ekström, K.; Bossios, A.; Sjöstrand, M.; Lee, J.J.; Lötvall, J.O. Exosome-Mediated Transfer of MRNAs and MicroRNAs Is a Novel Mechanism of Genetic Exchange between Cells. Nat. Cell Biol. 2007, 9, 654–659. [Google Scholar] [CrossRef]
- Corsten, M.F.; Dennert, R.; Jochems, S.; Kuznetsova, T.; Devaux, Y.; Hofstra, L.; Wagner, D.R.; Staessen, J.A.; Heymans, S.; Schroen, B. Circulating MicroRNA-208b and MicroRNA-499 Reflect Myocardial Damage in Cardiovascular Disease. Circ. Cardiovasc. Genet. 2010, 3, 499–506. [Google Scholar] [CrossRef] [PubMed]
- Ha, S.M.; Hwang, K.R.; Park, I.H.; Park, S.; Choi, J.S.; Park, D.J.; Park, J.-E.; Lee, S.H.; Lee, H.Y.; Seo, Y.J. Circulating MicroRNAs as Potentially New Diagnostic Biomarkers of Idiopathic Sudden Sensorineural Hearing Loss. Acta Otolaryngol. 2020, 140, 1013–1020. [Google Scholar] [CrossRef]
- Rupaimoole, R.; Slack, F.J. MicroRNA Therapeutics: Towards a New Era for the Management of Cancer and Other Diseases. Nat. Rev. Drug Discov. 2017, 16, 203–222. [Google Scholar] [CrossRef]
- Nunez, D.A.; Wijesinghe, P.; Nabi, S.; Yeh, D.; Garnis, C. MicroRNAs in Sudden Hearing Loss. Laryngoscope 2020, 130, E416–E422. [Google Scholar] [CrossRef]
- Mittal, R.; Liu, G.; Polineni, S.P.; Bencie, N.; Yan, D.; Liu, X.Z. Role of MicroRNAs in Inner Ear Development and Hearing Loss. Gene 2019, 686, 49–55. [Google Scholar] [CrossRef]
- Chen, H.; Wijesinghe, P.; Nunez, D. MicroRNAs in Acquired Sensorineural Hearing Loss. J. Laryngol. Otol. 2019, 133, 650–657. [Google Scholar] [CrossRef]
- Chandrasekhar, S.S.; Tsai Do, B.S.; Schwartz, S.R.; Bontempo, L.J.; Faucett, E.A.; Finestone, S.A.; Hollingsworth, D.B.; Kelley, D.M.; Kmucha, S.T.; Moonis, G.; et al. Clinical Practice Guideline: Sudden Hearing Loss (Update). Otolaryngol. Head Neck Surg. 2019, 161, S1–S45. [Google Scholar] [CrossRef]
- Kuhn, M.; Heman-Ackah, S.E.; Shaikh, J.A.; Roehm, P.C. Sudden Sensorineural Hearing Loss: A Review of Diagnosis, Treatment, and Prognosis. Trends Amplif. 2011, 15, 91–105. [Google Scholar] [CrossRef]
- Alexander, T.H.; Harris, J.P. Incidence of Sudden Sensorineural Hearing Loss. Otol. Neurotol. 2013, 34, 1586–1589. [Google Scholar] [CrossRef]
- Nakashima, T.; Sato, H.; Gyo, K.; Hato, N.; Yoshida, T.; Shimono, M.; Teranishi, M.; Sone, M.; Fukunaga, Y.; Kobashi, G.; et al. Idiopathic Sudden Sensorineural Hearing Loss in Japan. Acta Otolaryngol. 2014, 134, 1158–1163. [Google Scholar] [CrossRef] [PubMed]
- Rhee, T.-M.; Hwang, D.; Lee, J.-S.; Park, J.; Lee, J.M. Addition of Hyperbaric Oxygen Therapy vs Medical Therapy Alone for Idiopathic Sudden Sensorineural Hearing Loss: A Systematic Review and Meta-Analysis. JAMA Otolaryngol. Head Neck Surg. 2018, 144, 1153–1161. [Google Scholar] [CrossRef] [PubMed]
- Li, Q.; Peng, X.; Huang, H.; Li, J.; Wang, F.; Wang, J. RNA Sequencing Uncovers the Key MicroRNAs Potentially Contributing to Sudden Sensorineural Hearing Loss. Medicine 2017, 96, e8837. [Google Scholar] [CrossRef]
- Wangemann, P.; Wonneberger, K. Neurogenic Regulation of Cochlear Blood Flow Occurs along the Basilar Artery, the Anterior Inferior Cerebellar Artery and at Branch Points of the Spiral Modiolar Artery. Hear. Res. 2005, 209, 91–96. [Google Scholar] [CrossRef]
- Jiang, Y.; Chen, Y.; Yao, J.; Tian, Y.; Su, L.; Li, Y. Anatomic Assessment of Petrous Internal Carotid Artery, Facial Nerve, and Cochlea through the Anterior Transpetrosal Approach. J. Craniofac. Surg. 2015, 26, 2180–2183. [Google Scholar] [CrossRef] [PubMed]
- Kurata, N.; Schachern, P.A.; Paparella, M.M.; Cureoglu, S. Histopathologic Evaluation of Vascular Findings in the Cochlea in Patients with Presbycusis. JAMA Otolaryngol. Head Neck Surg. 2016, 142, 173–178. [Google Scholar] [CrossRef]
- Yoshida, T.; Ikemiyagi, Y.; Ikemiyagi, F.; Tamura, Y.; Suzuki, M.; Tsuyusaki, Y. Anterior Inferior Cerebellar Artery Infarction Misdiagnosed as Inner Ear Disease. B-ENT 2016, 12, 143–147. [Google Scholar] [PubMed]
- Hou, Z.-Q.; Wang, Q.-J. A New Disease: Pregnancy-Induced Sudden Sensorineural Hearing Loss? Acta Otolaryngol. 2011, 131, 779–786. [Google Scholar] [CrossRef] [PubMed]
- Niihori, M.; Platto, T.; Igarashi, S.; Hurbon, A.; Dunn, A.M.; Tran, P.; Tran, H.; Mudery, J.A.; Slepian, M.J.; Jacob, A. Zebrafish Swimming Behavior as a Biomarker for Ototoxicity-Induced Hair Cell Damage: A High-Throughput Drug Development Platform Targeting Hearing Loss. Transl. Res. 2015, 166, 440–450. [Google Scholar] [CrossRef]
- Kurtaran, H.; Acar, B.; Ocak, E.; Mirici, E. The Relationship between Senile Hearing Loss and Vestibular Activity. Braz. J. Otorhinolaryngol. 2016, 82, 650–653. [Google Scholar] [CrossRef] [PubMed]
- Attanasio, G.; Cagnoni, L.; Masci, E.; Ciciarello, F.; Diaferia, F.; Bruno, A.; Greco, A.; De Vincentiis, M. Chronic Cerebrospinal Venous Insufficiency as a Cause of Inner Ear Diseases. Acta Otolaryngol. 2017, 137, 460–463. [Google Scholar] [CrossRef] [PubMed]
- Joshua, T.G.; Ayub, A.; Wijesinghe, P.; Nunez, D.A. Hyperbaric Oxygen Therapy for Patients with Sudden Sensorineural Hearing Loss: A Systematic Review and Meta-Analysis. JAMA Otolaryngol. Head Neck Surg. 2022, 148, 5–11. [Google Scholar] [CrossRef] [PubMed]
- Wijesinghe, P.; Nunez, D.A.; Garnis, C. MicroRNA Signature and Cellular Characterization of Undifferentiated and Differentiated House Ear Institute-Organ of Corti 1 (HEI-OC1) Cells. J. Assoc. Res. Otolaryngol. 2022, 23, 467–489. [Google Scholar] [CrossRef]
- Safabakhsh, S.; Wijesinghe, P.; Nunez, M.; Nunez, D.A. The Role of Hypoxia-Associated MiRNAs in Acquired Sensorineural Hearing Loss. Front. Cell. Neurosci. 2022, 16, 361. [Google Scholar] [CrossRef]
- Dugas, J.C.; Cuellar, T.L.; Scholze, A.; Ason, B.; Ibrahim, A.; Emery, B.; Zamanian, J.L.; Foo, L.C.; McManus, M.T.; Barres, B.A. Dicer1 and MiR-219 Are Required for Normal Oligodendrocyte Differentiation and Myelination. Neuron 2010, 65, 597–611. [Google Scholar] [CrossRef]
- Schratt, G.; Tuebing, F.; Nigh, E.; Kane, C.; Sabatini, M.; Kiebler, M. Others Et Hjernespesifikt MikroRNA Regulerer Dendritisk Ryggradutvikling. Nature 2006, 439, 283–289. [Google Scholar] [CrossRef]
- Huang, W.; Feng, Y.; Liang, J.; Yu, H.; Wang, C.; Wang, B.; Wang, M.; Jiang, L.; Meng, W.; Cai, W.; et al. Loss of MicroRNA-128 Promotes Cardiomyocyte Proliferation and Heart Regeneration. Nat. Commun. 2018, 9, 700. [Google Scholar] [CrossRef] [PubMed]
- Motohashi, N.; Alexander, M.S.; Casar, J.C.; Kunkel, L.M. Identification of a Novel MicroRNA That Regulates the Proliferation and Differentiation in Muscle Side Population Cells. Stem Cells Dev. 2012, 21, 3031–3043. [Google Scholar] [CrossRef] [PubMed]
- Jain, R.K. Molecular Regulation of Vessel Maturation. Nat. Med. 2003, 9, 685–693. [Google Scholar] [CrossRef] [PubMed]
- Climent, M.; Quintavalle, M.; Miragoli, M.; Chen, J.; Condorelli, G.; Elia, L. TGFβ Triggers MiR-143/145 Transfer from Smooth Muscle Cells to Endothelial Cells, Thereby Modulating Vessel Stabilization. Circ. Res. 2015, 116, 1753–1764. [Google Scholar] [CrossRef]
- Owens, G.K. Regulation of Differentiation of Vascular Smooth Muscle Cells. Physiol. Rev. 1995, 75, 487–517. [Google Scholar] [CrossRef]
- Farina, F.M.; Hall, I.F.; Serio, S.; Zani, S.; Climent, M.; Salvarani, N.; Carullo, P.; Civilini, E.; Condorelli, G.; Elia, L.; et al. MiR-128-3p Is a Novel Regulator of Vascular Smooth Muscle Cell Phenotypic Switch and Vascular Diseases. Circ. Res. 2020, 126, e120–e135. [Google Scholar] [CrossRef]
- Yin, Y.; Cheng, Z.; Fu, X.; Ji, S. MicroRNA-375-3p Is Implicated in Carotid Artery Stenosis by Promoting the Cell Proliferation and Migration of Vascular Smooth Muscle Cells. BMC Cardiovasc. Disord. 2021, 21, 518. [Google Scholar] [CrossRef]
- Bhinge, A.; Namboori, S.C.; Bithell, A.; Soldati, C.; Buckley, N.J.; Stanton, L.W. MiR-375 Is Essential for Human Spinal Motor Neuron Development and May Be Involved in Motor Neuron Degeneration. Stem Cells 2016, 34, 124–134. [Google Scholar] [CrossRef]
- Wang, C.; Pan, Y.; Cheng, B.; Chen, J.; Bai, B. Identification of Conserved and Novel MicroRNAs in Cerebral Ischemia-Reperfusion Injury of Rat Using Deep Sequencing. J. Mol. Neurosci. 2014, 54, 671–683. [Google Scholar] [CrossRef]
- Wang, Y.; Dong, X.; Li, Z.; Wang, W.; Tian, J.; Chen, J. Downregulated RASD1 and Upregulated MiR-375 Are Involved in Protective Effects of Calycosin on Cerebral Ischemia/Reperfusion Rats. J. Neurol. Sci. 2014, 339, 144–148. [Google Scholar] [CrossRef]
- Wu, Z.; Tan, J.; Lin, L.; Zhang, W.; Yuan, W. MicroRNA-140-3p Protects Hippocampal Neuron against Pyroptosis to Attenuate Sevoflurane Inhalation-Induced Post-Operative Cognitive Dysfunction in Rats via Activation of HTR2A/ERK/Nrf2 Axis by Targeting DNMT1. Cell Death Discov. 2022, 8, 290. [Google Scholar] [CrossRef]
- Hao, L.; Lu, Y.; Ma, Y.; Wei, R.; Jia, Y. MiR-140 Protects against Myocardial Ischemia-Reperfusion Injury by Regulating NF-ΚB Pathway. Eur. Rev. Med. Pharmacol. Sci. 2020, 24, 11266–11272. [Google Scholar]
- Liang, S.; Ren, K.; Li, B.; Li, F.; Liang, Z.; Hu, J.; Xu, B.; Zhang, A. LncRNA SNHG1 Alleviates Hypoxia-Reoxygenation-Induced Vascular Endothelial Cell Injury as a Competing Endogenous RNA through the HIF-1α/VEGF Signal Pathway. Mol. Cell. Biochem. 2020, 465, 1–11. [Google Scholar] [CrossRef]
- Chen, F.; Chu, L.; Li, J.; Shi, Y.; Xu, B.; Gu, J.; Yao, X.; Tian, M.; Yang, X.; Sun, X. Hypoxia Induced Changes in MiRNAs and Their Target MRNAs in Extracellular Vesicles of Esophageal Squamous Cancer Cells. Thorac. Cancer 2020, 11, 570–580. [Google Scholar] [CrossRef]
- Becker, V.; Yuan, X.; Boewe, A.S.; Ampofo, E.; Ebert, E.; Hohneck, J.; Bohle, R.M.; Meese, E.; Zhao, Y.; Menger, M.D.; et al. Hypoxia-Induced Downregulation of MicroRNA-186-5p in Endothelial Cells Promotes Non-Small Cell Lung Cancer Angiogenesis by Upregulating Protein Kinase C Alpha. Mol. Ther.-Nucleic Acids 2023, 31, 421–436. [Google Scholar] [CrossRef] [PubMed]
- Liu, L.; Chen, L.; Wu, T.; Qian, H.; Yang, S. MicroRNA-30a-3p Functions as a Tumor Suppressor in Renal Cell Carcinoma by Targeting WNT2. Am. J. Transl. Res. 2019, 11, 4976–4983. [Google Scholar] [PubMed]
- Liu, C.; Zhou, X.; Long, Q.; Zeng, H.; Sun, Q.; Chen, Y.; Wu, D.; Liu, L. Small Extracellular Vesicles Containing MiR-30a-3p Attenuate the Migration and Invasion of Hepatocellular Carcinoma by Targeting SNAP23 Gene. Oncogene 2021, 40, 233–245. [Google Scholar] [CrossRef]
- Sempere, L.F.; Freemantle, S.; Pitha-Rowe, I.; Moss, E.; Dmitrovsky, E.; Ambros, V. Expression Profiling of Mammalian MicroRNAs Uncovers a Subset of Brain-Expressed MicroRNAs with Possible Roles in Murine and Human Neuronal Differentiation. Genome Biol. 2004, 5, R13. [Google Scholar] [CrossRef] [PubMed]
- Yang, C.-H.; Schrepfer, T.; Schacht, J. Age-Related Hearing Impairment and the Triad of Acquired Hearing Loss. Front. Cell. Neurosci. 2015, 9, 276. [Google Scholar] [CrossRef]
- Leake, P.A.; Hradek, G.T.; Hetherington, A.M.; Stakhovskaya, O. Brain-Derived Neurotrophic Factor Promotes Cochlear Spiral Ganglion Cell Survival and Function in Deafened, Developing Cats. J. Comp. Neurol. 2011, 519, 1526–1545. [Google Scholar] [CrossRef]
- Mullen, L.M.; Pak, K.K.; Chavez, E.; Kondo, K.; Brand, Y.; Ryan, A.F. Ras/P38 and PI3K/Akt but Not Mek/Erk Signaling Mediate BDNF-Induced Neurite Formation on Neonatal Cochlear Spiral Ganglion Explants. Brain Res. 2012, 1430, 25–34. [Google Scholar] [CrossRef]
- Pavlica, S.; Gebhardt, R. Protective Effects of Flavonoids and Two Metabolites against Oxidative Stress in Neuronal PC12 Cells. Life Sci. 2010, 86, 79–86. [Google Scholar] [CrossRef] [PubMed]
- Nelson, P.T.; Wang, W.-X.; Mao, G.; Wilfred, B.R.; Xie, K.; Jennings, M.H.; Gao, Z.; Wang, X. Specific Sequence Determinants of MiR-15/107 MicroRNA Gene Group Targets. Nucleic Acids Res. 2011, 39, 8163–8172. [Google Scholar] [CrossRef]
- Agarwal, V.; Bell, G.; Nam, J.; Bartel, D. Predicting Effective MicroRNA Target Sites in Mammalian MRNAs. eLife 2015, 4, e05005. [Google Scholar] [CrossRef]
- Qazi, T.J.; Lu, J.; Duru, L.; Zhao, J.; Qing, H. Upregulation of Mir-132 Induces Dopaminergic Neuronal Death via Activating SIRT1/P53 Pathway. Neurosci. Lett. 2021, 740, 135465. [Google Scholar] [CrossRef]
- Juźwik, C.A.; Drake, S.S.; Zhang, Y.; Paradis-Isler, N.; Sylvester, A.; Amar-Zifkin, A.; Douglas, C.; Morquette, B.; Moore, C.S.; Fournier, A.E. MicroRNA Dysregulation in Neurodegenerative Diseases: A Systematic Review. Prog. Neurobiol. 2019, 182, 101664. [Google Scholar] [CrossRef] [PubMed]
- Shaked, I.; Meerson, A.; Wolf, Y.; Avni, R.; Greenberg, D.; Gilboa-Geffen, A.; Soreq, H. MicroRNA-132 Potentiates Cholinergic Anti-Inflammatory Signaling by Targeting Acetylcholinesterase. Immunity 2009, 31, 965–973. [Google Scholar] [CrossRef]
- Lagos, D.; Pollara, G.; Henderson, S.; Gratrix, F.; Fabani, M.; Milne, R.S.; Gotch, F.; Boshoff, C. MiR-132 Regulates Antiviral Innate Immunity through Suppression of the P300 Transcriptional Co-Activator. Nat. Cell Biol. 2010, 12, 513–519. [Google Scholar] [CrossRef] [PubMed]
- Germano, J.F.; Sawaged, S.; Saadaeijahromi, H.; Andres, A.M.; Feuer, R.; Gottlieb, R.A.; Sin, J. Coxsackievirus B Infection Induces the Extracellular Release of MiR-590-5p, a Proviral MicroRNA. Virology 2019, 529, 169–176. [Google Scholar] [CrossRef]
- Zhou, Y.; Xia, Z.; Cheng, Z.; Xu, G.; Yang, X.; Liu, S.; Zhu, Y. Inducible MicroRNA-590-5p Inhibits Host Antiviral Response by Targeting the Soluble Interleukin-6 (IL6) Receptor. J. Biol. Chem. 2018, 293, 18168–18179. [Google Scholar] [CrossRef]
- O’Brien, J.; Hayder, H.; Zayed, Y.; Peng, C. Overview of MicroRNA Biogenesis, Mechanisms of Actions, and Circulation. Front. Endocrinol. 2018, 9, 402. [Google Scholar] [CrossRef]
- Butskiy, O.; Ng, D.; Hodgson, M.; Nunez, D.A. Rinne Test: Does the Tuning Fork Position Affect the Sound Amplitude at the Ear? J. Otolaryngol.-Head Neck Surg. 2016, 45, 1–8. [Google Scholar] [CrossRef] [PubMed]
- American Speech-Language-Hearing Association. Guidelines for Manual Pure-Tone Threshold Audiometry; American Speech-Language-Hearing Association: Rockville, MD, USA, 2005. [Google Scholar]
- Committee on Hearing and Equilibrium. Committee on Hearing and Equilibrium Guidelines for the Diagnosis and Evaluation of Therapy in Meniere’s Disease. Otolaryngol. Head Neck Surg. 1995, 113, 181–185. [Google Scholar] [CrossRef] [PubMed]
- Committee on Hearing and Equilibrium. Committee on Hearing and Equilibrium Guidelines for the Evaluation of Results of Treatment of Conductive Hearing Loss. Otolaryngol.-Head Neck Surg. 1995, 113, 186–187. [Google Scholar] [CrossRef] [PubMed]
- Schmittgen, T.D.; Livak, K.J. Analyzing Real-Time PCR Data by the Comparative C(T) Method. Nat. Protoc. 2008, 3, 1101–1108. [Google Scholar] [CrossRef] [PubMed]
- Clark, J.G. Uses and Abuses of Hearing Loss Classification. Asha 1981, 23, 493–500. [Google Scholar] [PubMed]


{kind=link}
| Time of Blood Sampling from Onset of Hearing Loss in Delayed Patients | 3 Months | 6 Months | 9 Months | 12 Months |
|---|---|---|---|---|
| Number of Patients | 2 | 3 | 2 | 2 |
| Demographic Details | Immediate Patients | Delayed Patients | p Value (Statistical Test) |
|---|---|---|---|
| Age in years (mean ± SD) | 58.07 ± 16.95 | 59.56 ± 15.86 | 0.92 (independent samples t-test) |
| Sex (male:female) | 7:7 | 5:4 | 0.36 (Pearson Chi square) |
| Dizziness (with dizziness:without dizziness) | 0:14 | 3:6 | 0.11 (Chi square) |
| Hearing recovery (recovered:not recovered) | 6:8 (42.9%:57.1%) | 5:4 (55.6%:44.4%) | 0.35 (Chi square) |
| Initial median PTA audiometric threshold of the affected ear (dB) | Q1: 40.1 Q3: 82.9 median: 66.9 min: 18.8 max:107.5 | Q1: 58.5 Q3: 91.5 median: 76.3 min: 23.8 max: 105 | 0.33 (Mann–Whitney U-test) |
| Final median PTA audiometric threshold of the affected ear (dB) | Q1: 36.2 Q3: 79.1 median: 63.3 min: 8.8 max: 102.5 | Q1: 14.4 Q3: 82.5 median: 67.5 min: 7.5 max: 105 | 0.92 (Mann–Whitney U-test) |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Abgoon, R.; Wijesinghe, P.; Garnis, C.; Nunez, D.A. The Expression Levels of MicroRNAs Differentially Expressed in Sudden Sensorineural Hearing Loss Patients’ Serum Are Unchanged for up to 12 Months after Hearing Loss Onset. Int. J. Mol. Sci. 2023, 24, 7307. https://doi.org/10.3390/ijms24087307
Abgoon R, Wijesinghe P, Garnis C, Nunez DA. The Expression Levels of MicroRNAs Differentially Expressed in Sudden Sensorineural Hearing Loss Patients’ Serum Are Unchanged for up to 12 Months after Hearing Loss Onset. International Journal of Molecular Sciences. 2023; 24(8):7307. https://doi.org/10.3390/ijms24087307
Chicago/Turabian StyleAbgoon, Reyhaneh, Printha Wijesinghe, Cathie Garnis, and Desmond A. Nunez. 2023. "The Expression Levels of MicroRNAs Differentially Expressed in Sudden Sensorineural Hearing Loss Patients’ Serum Are Unchanged for up to 12 Months after Hearing Loss Onset" International Journal of Molecular Sciences 24, no. 8: 7307. https://doi.org/10.3390/ijms24087307