
The Role of Mast Cells in the Induction and Maintenance of Inflammation in Selected Skin Diseases
 , , ,
, , ,
Abstract
:1. Introduction
2. Mast Cells and Their Role in the Skin
3. Mast Cell Activation Syndrome—Why It Is an Important Disease for Dermatologists?
- typical clinical symptoms arising from recurrent acute systemic MC activation (resembling recurrent anaphylaxis) have been documented;
- MC-derived mediators increase substantially in the serum (tryptase) or urine (histamine or prostaglandin-D2 metabolites) over the individual’s baseline (standard test: documented increase in serum tryptase levels following the 120% + 2 ng/mL formula);
- -
- primary (clonal = monoclonal)—characterized by the detection of KIT D816V mutation, MCs aberrantly display CD25 in most cases with confirmed mastocytosis (CM or SM), with only two minor SM criteria;
- -
- secondary (reactive)—characterized by an IgE-mediated allergy, another hypersensitivity reaction, or another immunologic disease that can induce the activation of MCs;
- -
- idiopathic—the criteria to diagnose MCAS are met, but no related reactive disease, no IgE-dependent allergy, no hereditary alpha tryptasemia, and no neoplastic MC are found [29].
4. Role of Mast Cells in Autoimmune Diseases
- they stimulate T lymphocytes through the expression of MHC class II, ensuring direct co-stimulation through the expression of receptors on the surface of the cell;
- they influence the increased proliferation and activation of T lymphocytes through various produced cytokines;
- they influence the maturing, migration, and functions of dendric cells and may have an indirect impact on the function of T cells;
- they reverse the Treg suppression of T effector cells and reduce the susceptibility of T effector cells to the Treg suppression;
- they recruit neutrophiles [60].
5. Role of Mast Cells in Psoriasis
5.1. Inflammatory Response to Injury—The Koebner Phenomenon
5.2. Response to Stress—Neuroinflammation in Psoriasis
6. Role of Mast Cells in Atopic Dermatitis
- -
- preformed chemical and protein mediators, such as histamine, serotonin, heparin and chondroitin sulphates, proteases, acid hydrolases, cathepsin, etc.;
- -
- lipid mediators, such as prostaglandins, leukotrienes, and the platelet-activating factor (PAF);
- -
- preformed and/or newly synthesized growth factors, cytokines, and chemokines, such as TNF-α, transforming growth factor beta (TGF- β), macrophage inflammatory protein-1a (MIP-1a/CCL3), monocyte chemoattractant protein-1 (MCP-1/CCL2), VEGF, IFN-α/β/γ, GM—CSF, IL-1a/b, IL-2, IL-3, IL-4, IL-5, IL-6, IL-8, IL- 9, IL-10, IL-11, IL-12, IL-13, IL-15, IL-16, IL-18, and IL-25 [77].
7. Role of Mast Cells in Kounis Syndrome in the Progression of Urticaria
- type
- I—patients with normal coronary arteries without factors predisposing them for coronary disease, in whom an acute allergic reaction causes a spasm of the coronary artery with normal cardiac enzymes and troponins, or a coronary artery vasospasm leading to acute myocardial infarction with increased cardiac enzymes and troponins (symptom of endothelium dysfunction or microvascular angina);
- type
- type
8. Role of Mast Cells in Ehlers–Danlos Syndrome (EDS)
9. Role of Mast Cells in Exanthemas in the Course of Hypersensitivity Reaction to Drugs
9.1. Anaphilactoidal Reaction (Pseudoallergy)
9.2. Red Man Syndrome
10. Role of Mast Cells in Allergic Contact Dermatitis
11. Role of Mast Cells in Rosacea
12. Cutaneous Mastocytosis
12.1. Clinical Presentation of Mastocytosis
12.2. Mast Cell Mediator-Related Symptoms in Patients with Mastocytosis
13. Discussion
- -
- “cancerous MCs” associated with clonal proliferation (mastocytosis group) [122];
- -
- -
14. Conclusions
Funding
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Doulatov, S.; Notta, F.; Laurenti, E.; Dick, J.E. Hematopoiesis: A Human Perspective. Cell Stem Cell 2012, 10, 120–136. [Google Scholar] [CrossRef] [Green Version]
- Dahlin, J.S.; Malinovschi, A.; Öhrvik, H.; Sandelin, M.; Janson, C.; Alving, K.; Hallgren, J. Lin− CD34hi CD117int/Hi FcεRI+ Cells in Human Blood Constitute a Rare Population of Mast Cell Progenitors. Blood 2016, 127, 383–391. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Grootens, J.; Ungerstedt, J.S.; Nilsson, G.; Dahlin, J.S. Deciphering the Differentiation Trajectory from Hematopoietic Stem Cells to Mast Cells. Blood Adv. 2018, 2, 2273–2281. [Google Scholar] [CrossRef] [PubMed]
- Kounis, N.G.; Koniari, I.; Velissaris, D.; Tzanis, G.; Hahalis, G. Kounis Syndrome—Not a Single-Organ Arterial Disorder but a Multisystem and Multidisciplinary Disease. Balk. Med. J. 2019, 36, 212. [Google Scholar] [CrossRef] [PubMed]
- Fong, M.; Crane, J.S. Histology, Mast Cells. In StatPearls; StatPearls Publishing: Treasure Island, FL, USA, 2022. [Google Scholar]
- Elieh Ali Komi, D.; Wöhrl, S.; Bielory, L. Mast Cell Biology at Molecular Level: A Comprehensive Review. Clinic. Rev. Allergy Immunol. 2020, 58, 342–365. [Google Scholar] [CrossRef]
- Dahlin, J.S.; Maurer, M.; Metcalfe, D.D.; Pejler, G.; Sagi-Eisenberg, R.; Nilsson, G. The Ingenious Mast Cell: Contemporary Insights into Mast Cell Behavior and Function. Allergy 2022, 77, 83–99. [Google Scholar] [CrossRef]
- Caughey, G.H. Mast Cell Tryptases and Chymases in Inflammation and Host Defense. Immunol. Rev. 2007, 217, 141–154. [Google Scholar] [CrossRef] [Green Version]
- Gonzalez-Quintela, A.; Vizcaino, L.; Gude, F.; Rey, J.; Meijide, L.; Fernandez-Merino, C.; Linneberg, A.; Vidal, C. Factors Influencing Serum Total Tryptase Concentrations in a General Adult Population. Clin. Chem. Lab. Med. 2010, 48, 701–706. [Google Scholar] [CrossRef]
- Fellinger, C.; Hemmer, W.; Wöhrl, S.; Sesztak-Greinecker, G.; Jarisch, R.; Wantke, F. Clinical Characteristics and Risk Profile of Patients with Elevated Baseline Serum Tryptase. Allergol. Immunopathol. 2014, 42, 544–552. [Google Scholar] [CrossRef]
- Lyons, J.J. Hereditary Alpha Tryptasemia. Immunol. Allergy Clin. N. Am. 2018, 38, 483–495. [Google Scholar] [CrossRef]
- Nguyen, A.V.; Soulika, A.M. The Dynamics of the Skin’s Immune System. Int. J. Mol. Sci. 2019, 20, 1811. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mast Cells as Regulators of Skin Inflammation and Immunity. Available online: http://www.medicaljournals.se/acta/content/html/10.2340/00015555-1197 (accessed on 13 November 2022).
- Zhao, W.; Kepley, C.L.; Morel, P.A.; Okumoto, L.M.; Fukuoka, Y.; Schwartz, L.B. FcγRIIa, Not FcγRIIb, Is Constitutively and Functionally Expressed on Skin-Derived Human Mast Cells. J. Immunol. 2006, 177, 694–701. [Google Scholar] [CrossRef] [PubMed]
- Tkaczyk, C.; Okayama, Y.; Woolhiser, M.R.; Hagaman, D.D.; Gilfillan, A.M.; Metcalfe, D.D. Activation of Human Mast Cells through the High Affinity IgG Receptor. Mol. Immunol. 2002, 38, 1289–1293. [Google Scholar] [CrossRef] [PubMed]
- Theoharides, T.C.; Kalogeromitros, D. The Critical Role of Mast Cells in Allergy and Inflammation. Ann. N. Y. Acad. Sci. 2006, 1088, 78–99. [Google Scholar] [CrossRef] [Green Version]
- Toyoda, M.; Makino, T.; Kagoura, M.; Morohashi, M. Immunolocalization of Substance P in Human Skin Mast Cells. Arch. Dermatol. Res. 2000, 292, 418–421. [Google Scholar] [CrossRef] [PubMed]
- Ødum, L.; Petersen, L.J.; Skov, P.S.; Ebskov, L.B. Pituitary Adenylate Cyclase Activating Polypeptide (PACAP) Is Localized in Human Dermal Neurons and Causes Histamine Release from Skin Mast Cells. Inflamm. Res. 1998, 47, 488–492. [Google Scholar] [CrossRef] [PubMed]
- Xiang, Z.; Nilsson, G. IgE Receptor-Mediated Release of Nerve Growth Factor by Mast Cells: IgE Receptor-Mediated Release of NGF. Clin. Exp. Allergy 2000, 30, 1379–1386. [Google Scholar] [CrossRef] [PubMed]
- Siiskonen, H.; Harvima, I. Mast Cells and Sensory Nerves Contribute to Neurogenic Inflammation and Pruritus in Chronic Skin Inflammation. Front. Cell. Neurosci. 2019, 13, 422. [Google Scholar] [CrossRef]
- Kulka, M.; Sheen, C.H.; Tancowny, B.P.; Grammer, L.C.; Schleimer, R.P. Neuropeptides Activate Human Mast Cell Degranulation and Chemokine Production. Immunology 2008, 123, 398–410. [Google Scholar] [CrossRef]
- Cao, J.; Papadopoulou, N.; Kempuraj, D.; Boucher, W.S.; Sugimoto, K.; Cetrulo, C.L.; Theoharides, T.C. Human Mast Cells Express Corticotropin-Releasing Hormone (CRH) Receptors and CRH Leads to Selective Secretion of Vascular Endothelial Growth Factor. J. Immunol. 2005, 174, 7665–7675. [Google Scholar] [CrossRef] [Green Version]
- Alysandratos, K.; Asadi, S.; Angelidou, A.; Zhang, B.; Sismanopoulos, N.; Yang, H.; Critchfield, A.; Theoharides, T.C. Neurotensin and CRH Interactions Augment Human Mast Cell Activation. PLoS ONE 2012, 7, e48934. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kounis, N.G. Kounis Syndrome (Allergic Angina and Allergic Myocardial Infarction): A Natural Paradigm? Int. J. Cardiol. 2006, 110, 7–14. [Google Scholar] [CrossRef] [PubMed]
- Hara, M.; Ono, K.; Wada, H.; Sasayama, S.; Matsumori, A. Preformed Angiotensin II Is Present in Human Mast Cells.6. Cardiovasc Drugs Ther. 2004; 18, 420–415. [Google Scholar]
- Cookson, H.; Grattan, C. An Update on Mast Cell Disorders. Clin. Med. 2016, 16, 580–583. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Patel, N.; Mohammadi, A.; Rhatigan, R. A Comparative Analysis of Mast Cell Quantification in Five Common Dermatoses: Lichen Simplex Chronicus, Psoriasis, Lichen Planus, Lupus, and Insect Bite/Allergic Contact Dermatitis/Nummular Dermatitis. ISRN Dermatol. 2012, 2012, 759630. [Google Scholar] [CrossRef] [Green Version]
- Porebski, G.; Kwiecien, K.; Pawica, M.; Kwitniewski, M. Mas-Related G Protein-Coupled Receptor-X2 (MRGPRX2) in Drug Hypersensitivity Reactions. Front. Immunol. 2018, 9, 3027. [Google Scholar] [CrossRef] [Green Version]
- Valent, P.; Hartmann, K.; Bonadonna, P.; Niedoszytko, M.; Triggiani, M.; Arock, M.; Brockow, K. Mast Cell Activation Syndromes: Collegium Internationale Allergologicum Update 2022. Int. Arch. Allergy Immunol. 2022, 183, 693–705. [Google Scholar] [CrossRef]
- Omraninava, M.; Eslami, M.M.; Aslani, S.; Razi, B.; Imani, D.; Feyzinia, S. Interleukin 13 Gene Polymorphism and Susceptibility to Asthma: A Meta-Regression and Meta-Analysis. Eur. Ann. Allergy Clin. Immunol. 2022, 54, 150. [Google Scholar] [CrossRef]
- Chatila, T.A. Interleukin-4 Receptor Signaling Pathways in Asthma Pathogenesis. Trends Mol. Med. 2004, 10, 493–499. [Google Scholar] [CrossRef]
- Kim, S.-H.; Ye, Y.-M.; Palikhe, N.S.; Kim, J.-E.; Park, H.-S. Genetic and Ethnic Risk Factors Associated with Drug Hypersensitivity. Curr. Opin. Allergy Clin. Immunol. 2010, 10, 280–290. [Google Scholar] [CrossRef]
- Rausz, E.; Szilágyi, A.; Nedoszytko, B.; Lange, M.; Niedoszytko, M.; Lautner-Csorba, O.; Falus, A.; Aladzsity, I.; Kokai, M.; Valent, P.; et al. Comparative Analysis of IL6 and IL6 Receptor Gene Polymorphisms in Mastocytosis. Br. J. Haematol. 2013, 160, 216–219. [Google Scholar] [CrossRef] [Green Version]
- Nedoszytko, B.; Niedoszytko, M.; Lange, M.; van Doormaal, J.; Gleń, J.; Zabłotna, M.; Renke, J.; Vales, A.; Buljubasic, F.; Jassem, E.; et al. Interleukin-13 Promoter Gene Polymorphism -1112C/T Is Associated with the Systemic Form of Mastocytosis. Allergy 2009, 64, 287–294. [Google Scholar] [CrossRef] [PubMed]
- Grootens, J.; Ungerstedt, J.S.; Ekoff, M.; Rönnberg, E.; Klimkowska, M.; Amini, R.-M.; Arock, M.; Söderlund, S.; Mattsson, M.; Nilsson, G.; et al. Single-Cell Analysis Reveals the KIT D816V Mutation in Haematopoietic Stem and Progenitor Cells in Systemic Mastocytosis. EBioMedicine 2019, 43, 150–158. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Furitsu, T.; Tsujimura, T.; Tono, T.; Ikeda, H.; Kitayama, H.; Koshimizu, U.; Sugahara, H.; Butterfield, J.H.; Ashman, L.K.; Kanayama, Y. Identification of Mutations in the Coding Sequence of the Proto-Oncogene c-Kit in a Human Mast Cell Leukemia Cell Line Causing Ligand-Independent Activation of c-Kit Product. J. Clin. Investig. 1993, 92, 1736–1744. [Google Scholar] [CrossRef] [PubMed]
- Garcia-Montero, A.C.; Jara-Acevedo, M.; Teodosio, C.; Sanchez, M.L.; Nunez, R.; Prados, A.; Aldanondo, I.; Sanchez, L.; Dominguez, M.; Botana, L.M.; et al. KIT Mutation in Mast Cells and Other Bone Marrow Hematopoietic Cell Lineages in Systemic Mast Cell Disorders: A Prospective Study of the Spanish Network on Mastocytosis (REMA) in a Series of 113 Patients. Blood 2006, 108, 2366–2372. [Google Scholar] [CrossRef]
- Varney, V.A.; Nicholas, A.; Warner, A.; Sumar, N. IgE-Mediated Systemic Anaphylaxis And Its Association with Gene Polymorphisms Of ACE, Angiotensinogen And Chymase. JAA 2019, 12, 343–361. [Google Scholar] [CrossRef] [Green Version]
- Theoharides, T.C.; Valent, P.; Akin, C. Mast Cells, Mastocytosis, and Related Disorders. N. Engl. J. Med. 2015, 373, 163–172. [Google Scholar] [CrossRef]
- Valent, P.; Akin, C.; Arock, M.; Brockow, K.; Butterfield, J.H.; Carter, M.C.; Castells, M.; Escribano, L.; Hartmann, K.; Lieberman, P.; et al. Definitions, Criteria and Global Classification of Mast Cell Disorders with Special Reference to Mast Cell Activation Syndromes: A Consensus Proposal. Int. Arch. Allergy Immunol. 2012, 157, 215–225. [Google Scholar] [CrossRef] [Green Version]
- Niedoszytko, M.; Ratajska, M.; Chełmińska, M.; Makowiecki, M.; Malek, E.; Siemińska, A.; Limon, J.; Jassem, E. The Angiotensinogen AGT p.M235T Gene Polymorphism May Be Responsible for the Development of Severe Anaphylactic Reactions to Insect Venom Allergens. Int. Arch. Allergy Immunol. 2010, 153, 166–172. [Google Scholar] [CrossRef]
- Lyons, J.J.; Chovanec, J.; O’Connell, M.P.; Liu, Y.; Šelb, J.; Zanotti, R.; Bai, Y.; Kim, J.; Le, Q.T.; DiMaggio, T.; et al. Heritable Risk for Severe Anaphylaxis Associated with Increased α-Tryptase–Encoding Germline Copy Number at TPSAB1. J. Allergy Clin. Immunol. 2021, 147, 622–632. [Google Scholar] [CrossRef]
- Greiner, G.; Sprinzl, B.; Górska, A.; Ratzinger, F.; Gurbisz, M.; Witzeneder, N.; Schmetterer, K.G.; Gisslinger, B.; Uyanik, G.; Hadzijusufovic, E.; et al. Hereditary α Tryptasemia Is a Valid Genetic Biomarker for Severe Mediator-Related Symptoms in Mastocytosis. Blood 2021, 137, 238–247. [Google Scholar] [CrossRef]
- Jarkvist, J.; Brockow, K.; Gülen, T. Low Frequency of IgE-Mediated Food Hypersensitivity in Mastocytosis. J. Allergy Clin. Immunol. Pract. 2020, 8, 3093–3101. [Google Scholar] [CrossRef] [PubMed]
- Choi, I.S.; Koh, Y.I.; Chung, S.-W.; Lim, H. Increased Releasability of Skin Mast Cells after Exercise in Patients with Exercise-Induced Asthma. J. Korean Med. Sci. 2004, 19, 724. [Google Scholar] [CrossRef] [PubMed]
- Carter, M.C.; Metcalfe, D.D.; Matito, A.; Escribano, L.; Butterfield, J.H.; Schwartz, L.B.; Bonadonna, P.; Zanotti, R.; Triggiani, M.; Castells, M.; et al. Adverse Reactions to Drugs and Biologics in Patients with Clonal Mast Cell Disorders: A Work Group Report of the Mast Cells Disorder Committee, American Academy of Allergy, Asthma & Immunology. J. Allergy Clin. Immunol. 2019, 143, 880–893. [Google Scholar] [CrossRef] [PubMed]
- Picard, M.; Giavina-Bianchi, P.; Mezzano, V.; Castells, M. Expanding Spectrum of Mast Cell Activation Disorders: Monoclonal and Idiopathic Mast Cell Activation Syndromes. Clin. Ther. 2013, 35, 548–562. [Google Scholar] [CrossRef]
- Alvarez-Twose, I.; González-de-Olano, D.; Sánchez-Muñoz, L.; Matito, A.; Jara-Acevedo, M.; Teodosio, C.; García-Montero, A.; Morgado, J.M.; Orfao, A.; Escribano, L. Validation of the REMA Score for Predicting Mast Cell Clonality and Systemic Mastocytosis in Patients with Systemic Mast Cell Activation Symptoms. Int. Arch. Allergy Immunol. 2012, 157, 275–280. [Google Scholar] [CrossRef] [Green Version]
- Hamilton, M.J.; Scarlata, K. Mast Cell Activation Syndrome—What It Is and Isn’t. Pract. Gastroenterol. 2020, 7, 26–32. [Google Scholar]
- Cardet, J.-C.; Castells, M.C.; Hamilton, M.J. Immunology and Clinical Manifestations of Non-Clonal Mast Cell Activation Syndrome. Curr. Allergy Asthma Rep. 2013, 13, 10–18. [Google Scholar] [CrossRef]
- Galli, S.J.; Gaudenzio, N.; Tsai, M. Mast Cells in Inflammation and Disease: Recent Progress and Ongoing Concerns. Annu. Rev. Immunol. 2020, 38, 49–77. [Google Scholar] [CrossRef]
- Le, Q.T.; Lyons, J.J.; Naranjo, A.N.; Olivera, A.; Lazarus, R.A.; Metcalfe, D.D.; Milner, J.D.; Schwartz, L.B. Impact of Naturally Forming Human α/β-Tryptase Heterotetramers in the Pathogenesis of Hereditary α-Tryptasemia. J. Exp. Med. 2019, 216, 2348–2361. [Google Scholar] [CrossRef]
- Naranjo, A.N.; Bandara, G.; Bai, Y.; Smelkinson, M.G.; Tobío, A.; Komarow, H.D.; Boyden, S.E.; Kastner, D.L.; Metcalfe, D.D.; Olivera, A. Critical signaling events in the mechanoactivation of human mast cells through p. C492Y-ADGRE2. J. Investig. Dermatol. 2020, 140, 2210–2220.e5. [Google Scholar] [CrossRef]
- Valent, P.; Akin, C.; Hartmann, K.; Nilsson, G.; Reiter, A.; Hermine, O.; Sotlar, K.; Sperr, W.R.; Escribano, L.; George, T.I.; et al. Mast Cells as a Unique Hematopoietic Lineage and Cell System: From Paul Ehrlich’s Visions to Precision Medicine Concepts. Theranostics 2020, 10, 10743–10768. [Google Scholar] [CrossRef] [PubMed]
- Brockow, K.; Jofer, C.; Behrendt, H.; Ring, J. Anaphylaxis in Patients with Mastocytosis: A Study on History, Clinical Features and Risk Factors in 120 Patients: Anaphylaxis in Patients with Mastocytosis. Allergy 2008, 63, 226–232. [Google Scholar] [CrossRef] [PubMed]
- Akin, C.; Valent, P.; Metcalfe, D.D. Mast Cell Activation Syndrome: Proposed Diagnostic Criteria. J. Allergy Clin. Immunol. 2010, 126, 1099–1104.e4. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Valent, P. Mast Cell Activation Syndromes: Definition and Classification. Allergy 2013, 68, 417–424. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Valent, P.; Akin, C.; Bonadonna, P.; Hartmann, K.; Brockow, K.; Niedoszytko, M.; Nedoszytko, B.; Siebenhaar, F.; Sperr, W.R.; Elberink, J.N.G.O.; et al. Proposed Diagnostic Algorithm for Patients with Suspected Mast Cell Activation Syndrome (MCAS). J. Allergy Clin. Immunol. Pract. 2019, 7, 1125–1133.e1. [Google Scholar] [CrossRef] [PubMed]
- Valent, P.; Akin, C.; Nedoszytko, B.; Bonadonna, P.; Hartmann, K.; Niedoszytko, M.; Brockow, K.; Siebenhaar, F.; Triggiani, M.; Arock, M.; et al. Diagnosis, Classification and Management of Mast Cell Activation Syndromes (MCAS) in the Era of Personalized Medicine. Int. J. Mol. Sci. 2020, 21, 9030. [Google Scholar] [CrossRef]
- Walker, M.E.; Hatfield, J.K.; Brown, M.A. New Insights into the Role of Mast Cells in Autoimmunity: Evidence for a Common Mechanism of Action? Biochim. Biophys. Acta (BBA) Mol. Basis Dis. 2012, 1822, 57–65. [Google Scholar] [CrossRef] [Green Version]
- Ramos-Casals, M.; Brito-Zerón, P.; López-Guillermo, A.; Khamashta, M.A.; Bosch, X. Adult Haemophagocytic Syndrome. Lancet 2014, 383, 1503–1516. [Google Scholar] [CrossRef]
- Sandler, R.D.; Carter, S.; Kaur, H.; Francis, S.; Tattersall, R.S.; Snowden, J.A. Haemophagocytic Lymphohistiocytosis (HLH) Following Allogeneic Haematopoietic Stem Cell Transplantation (HSCT)—Time to Reappraise with Modern Diagnostic and Treatment Strategies? Bone Marrow Transpl. 2020, 55, 307–316. [Google Scholar] [CrossRef]
- Parisi, R.; Symmons, D.P.M.; Griffiths, C.E.M.; Ashcroft, D.M. Global Epidemiology of Psoriasis: A Systematic Review of Incidence and Prevalence. J. Investig. Dermatol. 2013, 133, 377–385. [Google Scholar] [CrossRef] [Green Version]
- Tsoi, L.C.; Stuart, P.E.; Tian, C.; Gudjonsson, J.E.; Das, S.; Zawistowski, M.; Ellinghaus, E.; Barker, J.N.; Chandran, V.; Dand, N.; et al. Large Scale Meta-Analysis Characterizes Genetic Architecture for Common Psoriasis Associated Variants. Nat. Commun. 2017, 8, 15382. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liu, Y.; Helms, C.; Liao, W.; Zaba, L.C.; Duan, S.; Gardner, J.; Wise, C.; Miner, A.; Malloy, M.J.; Pullinger, C.R.; et al. A Genome-Wide Association Study of Psoriasis and Psoriatic Arthritis Identifies New Disease Loci. PLoS Genet. 2008, 4, e1000041. [Google Scholar] [CrossRef] [PubMed]
- Armstrong, A.W. Psoriasis Provoked or Exacerbated by Medications: Identifying Culprit Drugs. JAMA Dermatol. 2014, 150, 963. [Google Scholar] [CrossRef] [PubMed]
- Armstrong, A.W.; Read, C. Pathophysiology, Clinical Presentation, and Treatment of Psoriasis: A Review. JAMA 2020, 323, 1945. [Google Scholar] [CrossRef]
- Ji, Y.-Z.; Liu, S.-R. Koebner Phenomenon Leading to the Formation of New Psoriatic Lesions: Evidences and Mechanisms. Biosci. Rep. 2019, 39, BSR20193266. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Toruniowa, B.; Jablońska, S. Mast Cells in the Initial Stages of Psoriasis. Arch. Dermatol. Res. 1988, 280, 189–193. [Google Scholar] [CrossRef]
- Woźniak, E.; Owczarczyk-Saczonek, A.; Placek, W. Psychological Stress, Mast Cells, and Psoriasis—Is There Any Relationship? Int. J. Mol. Sci. 2021, 22, 13252. [Google Scholar] [CrossRef]
- Ortega, M.A.; García-Montero, C.; Fraile-Martinez, O.; Alvarez-Mon, M.A.; Gómez-Lahoz, A.M.; Lahera, G.; Monserrat, J.; Rodriguez-Jimenez, R.; Quintero, J.; Álvarez-Mon, M. Immune-Mediated Diseases from the Point of View of Psychoneuroimmunoendocrinology. Biology 2022, 11, 973. [Google Scholar] [CrossRef]
- Scholzen, T.; Armstrong, C.A.; Bunnett, N.W.; Luger, T.A.; Olerud, J.E.; Ansel, J.C. Neuropeptides in the Skin: Interactions between the Neuroendocrine and the Skin Immune Systems. Exp. Dermatol. 1998, 7, 81–96. [Google Scholar] [CrossRef]
- Schulze, E.; Witt, M.; Fink, T.; Hofer, A.; Funk, R.H.W. Immunohistochemical Detection of Human Skin Nerve Fibers. Acta Histochem. 1997, 99, 301–309. [Google Scholar] [CrossRef]
- Choi, J.E.; Di Nardo, A. Skin Neurogenic Inflammation. Semin. Immunopathol. 2018, 40, 249–259. [Google Scholar] [CrossRef]
- Joseph, T.; Kurian, J.; Warwick, D.J.; Friedmann, P.S. Unilateral Remission of Psoriasis Following Traumatic Nerve Palsy. Br. J. Dermatol. 2005, 152, 185–186. [Google Scholar] [CrossRef]
- Zhu, T.H.; Nakamura, M.; Farahnik, B.; Abrouk, M.; Lee, K.; Singh, R.; Gevorgyan, A.; Koo, J.; Bhutani, T. The Role of the Nervous System in the Pathophysiology of Psoriasis: A Review of Cases of Psoriasis Remission or Improvement Following Denervation Injury. Am. J. Clin. Dermatol. 2016, 17, 257–263. [Google Scholar] [CrossRef]
- Kawakami, T.; Ando, T.; Kimura, M.; Wilson, B.S.; Kawakami, Y. Mast Cells in Atopic Dermatitis. Curr. Opin. Immunol. 2009, 21, 666–678. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liu, F.-T.; Goodarzi, H.; Chen, H.-Y. IgE, Mast Cells, and Eosinophils in Atopic Dermatitis. Clinic. Rev. Allergy Immunol. 2011, 41, 298–310. [Google Scholar] [CrossRef] [PubMed]
- Kanbe, T.; Soma, Y.; Kawa, Y.; Kashima, M.; Mizoguchi, M. Serum Levels of Soluble Stem Cell Factor and Soluble KIT Are Elevated in Patients with Atopic Dermatitis and Correlate with the Disease Severity. Br. J. Dermatol. 2001, 144, 1148–1153. [Google Scholar] [CrossRef] [PubMed]
- Oh, M.; Zhu, Z.; Yu, J.; Zheng, T. Role of Mast Cells in the Development of Atopic Dermatitis Induced by IL-13. J. Allergy Clin. Immunol. 2012, 129, AB36. [Google Scholar] [CrossRef]
- Schwartz, H.J.; Yunginger, J.W.; Schwartz, L.B. Is Unrecognized Anaphylaxis a Cause of Sudden Unexpected Death? Clin. Exp. Allergy 1995, 25, 866–870. [Google Scholar] [CrossRef]
- Abdelghany, M.; Subedi, R.; Shah, S.; Kozman, H. Kounis Syndrome: A Review Article on Epidemiology, Diagnostic Findings, Management and Complications of Allergic Acute Coronary Syndrome. Int. J. Cardiol. 2017, 232, 1–4. [Google Scholar] [CrossRef]
- Forlani, D.; Scarano, G.; D’Alleva, A.; Di Marco, M.; Paloscia, L.; Gatta, A.; Della Valle, L.; Farinelli, A.; Lumaca, A.; Petrarca, C.; et al. Kounis Syndrome as First Manifestation of Allergic Sensitization. Case Rep. Med. 2019, 2019, 6317956. [Google Scholar] [CrossRef]
- Kounis, N.G. Serum Tryptase Levels and Kounis Syndrome. Int. J. Cardiol. 2007, 114, 407–408. [Google Scholar] [CrossRef] [PubMed]
- Fassio, F.; Losappio, L.; Antolin-Amerigo, D.; Peveri, S.; Pala, G.; Preziosi, D.; Massaro, I.; Giuliani, G.; Gasperini, C.; Caminati, M.; et al. Kounis Syndrome: A Concise Review with Focus on Management. Eur. J. Intern. Med. 2016, 30, 7–10. [Google Scholar] [CrossRef] [PubMed]
- Malfait, F.; Francomano, C.; Byers, P.; Belmont, J.; Berglund, B.; Black, J.; Bloom, L.; Bowen, J.M.; Brady, A.F.; Burrows, N.P.; et al. The 2017 International Classification of the Ehlers–Danlos Syndromes. Am. J. Med. Genet. Part C Semin. Med. Genet. 2017, 175, 8–26. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hakim, A.; Grahame, R. Joint Hypermobility. Best Pract. Res. Clin. Rheumatol. 2003, 17, 989–1004. [Google Scholar] [CrossRef]
- Seneviratne, S.L.; Maitland, A.; Afrin, L. Mast Cell Disorders in Ehlers-Danlos Syndrome. Am. J. Med. Genet. 2017, 175, 226–236. [Google Scholar] [CrossRef] [Green Version]
- Afrin, L.B.; Self, S.; Menk, J.; Lazarchick, J. Characterization of Mast Cell Activation Syndrome. Am. J. Med. Sci. 2017, 353, 207–215. [Google Scholar] [CrossRef] [Green Version]
- Luzgina, N.G.; Potapova, O.V.; Shkurupiy, V.A. Structural and Functional Peculiarities of Mast Cells in Undifferentiated Connective Tissue Dysplasia. Bull. Exp. Biol. Med. 2011, 150, 676–678. [Google Scholar] [CrossRef]
- Molderings, G.J. Transgenerational Transmission of Systemic Mast Cell Activation Disease—Genetic and Epigenetic Features. Transl. Res. 2016, 174, 86–97. [Google Scholar] [CrossRef]
- Zhang, B.; Li, Q.; Shi, C.; Zhang, X. Drug-Induced Pseudoallergy: A Review of the Causes and Mechanisms. Pharmacology 2018, 101, 104–110. [Google Scholar] [CrossRef]
- Hamilton, M.J. Nonclonal Mast Cell Activation Syndrome. Immunol. Allergy Clin. N. Am. 2018, 38, 469–481. [Google Scholar] [CrossRef]
- Pastorello, E.A.; Stafylaraki, C.; Mirone, C.; Preziosi, D.; Aversano, M.G.; Mascheri, A.; Losappio, L.M.; Ortolani, V.; Nichelatti, M.; Farioli, L. Anti-Amoxicillin Immunoglobulin E, Histamine-2 Receptor Antagonist Therapy and Mast Cell Activation Syndrome Are Risk Factors for Amoxicillin Anaphylaxis. Int. Arch. Allergy Immunol. 2015, 166, 280–286. [Google Scholar] [CrossRef] [PubMed]
- McNeil, B.D. Identification of a Mast-Cell-Specific Receptor Crucial for Pseudo-Allergic Drug Reactions. 18.
- Fica, A.; Muñoz, D.; Rojas, T.; Sanzana, C.; Muñoz, C.; Fica, A.; Muñoz, D.; Rojas, T.; Sanzana, C.; Muñoz, C. Penicillin Desensitization in Allergic Pregnant Women with Syphilis. Report of Two Cases. Rev. Médica De Chile 2020, 148, 344–348. [Google Scholar] [CrossRef] [PubMed]
- STD Treatment Guidelines. 2015. Available online: https://www.cdc.gov/std/tg2015/default.htm (accessed on 4 February 2021).
- Janier, M.; Unemo, M.; Dupin, N.; Tiplica, G.S.; Potočnik, M.; Patel, R. 2020 European Guideline on the Management of Syphilis. J. Eur. Acad Dermatol. Venereol. 2020, 35, 574–588. [Google Scholar] [CrossRef] [PubMed]
- Pham, M.N.; Ho, H.-E.; Desai, M. Penicillin Desensitization: Treatment of Syphilis in Pregnancy in Penicillin-Allergic Patients. Ann. Allergy Asthma Immunol. 2017, 118, 537–541. [Google Scholar] [CrossRef] [PubMed]
- Staso, P.; Leonov, A. Drug Desensitization in 17-Year-Old Male with Mast Cell Activation Syndrome, Pneumonia, and Antibiotic Hypersensitivities. AME Case Rep. 2017, 1, 7. [Google Scholar] [CrossRef] [PubMed]
- Azimi, E.; Reddy, V.B.; Lerner, E.A. MRGPRX2, Atopic Dermatitis, and Red Man Syndrome. Itch 2017, 2, e5. [Google Scholar] [CrossRef]
- Sivagnanam, S.; Deleu, D. Red Man Syndrome. Crit. Care 2003, 7, 119–120. [Google Scholar] [CrossRef] [Green Version]
- Arroyo-Mercado, F.; Khudyakov, A.; Chawla, G.S.; Cantres-Fonseca, O.; McFarlane, I.M. Red Man Syndrome with Oral Vancomycin: A Case Report. Am. J. Med. Case Rep. 2019, 7, 16–17. [Google Scholar] [CrossRef]
- PRIME PubMed | Red Man Syndrome and Infliximab. Available online: https://www.unboundmedicine.com/medline/citation/12544210/Red_man_syndrome_and_infliximab_ (accessed on 4 February 2021).
- Myers, A.L.; Gaedigk, A.; Dai, H.; James, L.P.; Jones, B.L.; Neville, K.A. Defining Risk Factors for Red Man Syndrome in Children and Adults. Pediatr. Infect. Dis. J. 2012, 31, 464–468. [Google Scholar] [CrossRef] [Green Version]
- Soong, L.; Sotoodian, B.; Brassard, A. Immediate-Type Infliximab Infusion Reaction Presenting as Purpura. JAAD Case Rep. 2018, 4, 596–598. [Google Scholar] [CrossRef]
- Nassau, S.; Fonacier, L. Allergic Contact Dermatitis. Med. Clin. N. Am. 2020, 104, 61–76. [Google Scholar] [CrossRef] [PubMed]
- Owen, J.L.; Vakharia, P.P.; Silverberg, J.I. The Role and Diagnosis of Allergic Contact Dermatitis in Patients with Atopic Dermatitis. Am. J. Clin. Dermatol. 2018, 19, 293–302. [Google Scholar] [CrossRef] [PubMed]
- Hernandez, J.D.; Yu, M.; Sibilano, R.; Tsai, M.; Galli, S.J. Development of Multiple Features of Antigen-Induced Asthma Pathology in a New Strain of Mast Cell Deficient BALB/c- Kit W-Sh/W-Sh Mice. Lab. Investig. 2020, 100, 516–526. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Morita, H.; Saito, H.; Matsumoto, K.; Nakae, S. Regulatory Roles of Mast Cells in Immune Responses. Semin. Immunopathol. 2016, 38, 623–629. [Google Scholar] [CrossRef]
- Aroni, K.; Tsagroni, E.; Kavantzas, N.; Patsouris, E.; Ioannidis, E. A Study of the Pathogenesis of Rosacea: How Angiogenesis and Mast Cells May Participate in a Complex Multifactorial Process. Arch. Dermatol. Res. 2008, 300, 125–131. [Google Scholar] [CrossRef]
- Wang, L.; Wang, Y.-J.; Hao, D.; Wen, X.; Du, D.; He, G.; Jiang, X. The Theranostics Role of Mast Cells in the Pathophysiology of Rosacea. Front. Med. 2019, 6, 324. [Google Scholar] [CrossRef] [Green Version]
- Muto, Y.; Wang, Z.; Vanderberghe, M.; Two, A.; Gallo, R.L.; Di Nardo, A. Mast Cells Are Key Mediators of Cathelicidin-Initiated Skin Inflammation in Rosacea. J. Investig. Dermatol. 2014, 134, 2728–2736. [Google Scholar] [CrossRef] [Green Version]
- Holmes, A.D.; Steinhoff, M. Integrative Concepts of Rosacea Pathophysiology, Clinical Presentation and New Therapeutics. Exp. Dermatol. 2017, 26, 659–667. [Google Scholar] [CrossRef] [Green Version]
- Ahn, C.S.; Huang, W.W. Rosacea Pathogenesis. Dermatol. Clin. 2018, 36, 81–86. [Google Scholar] [CrossRef]
- Systemic Mastocytosis in Adults: 2021 Update on Diagnosis, Risk Stratification and Management—Pardanani—2021—American Journal of Hematology—Wiley Online Library. Available online: https://onlinelibrary.wiley.com/doi/10.1002/ajh.26118 (accessed on 20 December 2022).
- Lange, M.; Hartmann, K.; Carter, M.C.; Siebenhaar, F.; Alvarez-Twose, I.; Torrado, I.; Brockow, K.; Renke, J.; Irga-Jaworska, N.; Plata-Nazar, K.; et al. Molecular Background, Clinical Features and Management of Pediatric Mastocytosis: Status 2021. IJMS 2021, 22, 2586. [Google Scholar] [CrossRef]
- Valent, P.; Akin, C.; Gleixner, K.V.; Sperr, W.R.; Reiter, A.; Arock, M.; Triggiani, M. Multidisciplinary Challenges in Mastocytosis and How to Address with Personalized Medicine Approaches. IJMS 2019, 20, 2976. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Swerdlow, S.H.; Campo, E.; Harris, N.L.; Jaffe, E.S.; Pileri, S.A.; Stein, H.; Thiele, J.; Vardiman, J.W. WHO Classification of Tumours of Haematopoietic and Lymphoid Tissues. Available online: https://hero.epa.gov/hero/index.cfm/reference/details/reference_id/786623 (accessed on 20 December 2022).
- Chatterjee, A.; Ghosh, J.; Kapur, R. Mastocytosis: A Mutated KIT Receptor Induced Myeloproliferative Disorder. Oncotarget 2015, 6, 18250–18264. [Google Scholar] [CrossRef] [PubMed]
- Valent, P.; Akin, C.; Metcalfe, D.D. Mastocytosis: 2016 Updated WHO Classification and Novel Emerging Treatment Concepts. Blood 2017, 129, 1420–1427. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hartmann, K.; Escribano, L.; Grattan, C.; Brockow, K.; Carter, M.C.; Alvarez-Twose, I.; Matito, A.; Broesby-Olsen, S.; Siebenhaar, F.; Lange, M.; et al. Cutaneous Manifestations in Patients with Mastocytosis: Consensus Report of the European Competence Network on Mastocytosis; the American Academy of Allergy, Asthma & Immunology; and the European Academy of Allergology and Clinical Immunology. J. Allergy Clin. Immunol. 2016, 137, 35–45. [Google Scholar] [CrossRef] [PubMed]
- Valent, P.; Horny, H.-P.; Escribano, L.; Longley, B.J.; Li, C.Y.; Schwartz, L.B.; Marone, G.; Nuñez, R.; Akin, C.; Sotlar, K.; et al. Diagnostic Criteria and Classification of Mastocytosis: A Consensus Proposal. Leuk. Res. 2001, 25, 603–625. [Google Scholar] [CrossRef] [PubMed]
- Brockow, K.; Plata-Nazar, K.; Lange, M.; Nedoszytko, B.; Niedoszytko, M.; Valent, P. Mediator-Related Symptoms and Anaphylaxis in Children with Mastocytosis. IJMS 2021, 22, 2684. [Google Scholar] [CrossRef]
- Komi, D.E.A.; Rambasek, T.; Wöhrl, S. Mastocytosis: From a Molecular Point of View. Clinic. Rev. Allergy Immunol. 2018, 54, 397–411. [Google Scholar] [CrossRef]
- Méni, C.; Bruneau, J.; Georgin-Lavialle, S.; Le Saché de Peufeilhoux, L.; Damaj, G.; Hadj-Rabia, S.; Fraitag, S.; Dubreuil, P.; Hermine, O.; Bodemer, C. Paediatric Mastocytosis: A Systematic Review of 1747 Cases. Br. J. Dermatol. 2015, 172, 642–651. [Google Scholar] [CrossRef]
- Barnes, M.; Van, L.; DeLong, L.; Lawley, L.P. Severity of Cutaneous Findings Predict the Presence of Systemic Symptoms in Pediatric Maculopapular Cutaneous Mastocytosis. Pediatr. Dermatol. 2014, 31, 271–275. [Google Scholar] [CrossRef]
- Krečak, I.; Karaman, I.; Jakelić, S. Darier’s Sign. Eur. J. Intern. Med. 2022, 97, 97–98. [Google Scholar] [CrossRef]
- Kwiatkowska, D.; Reich, A. Role of Mast Cells in the Pathogenesis of Pruritus in Mastocytosis. Acta Derm. Venereol. 2021, 101, 350. [Google Scholar] [CrossRef] [PubMed]
- Reich, A.; Szepietowski, J.C. Diagnostic Procedures of Itch. In Current Problems in Dermatology; Szepietowski, J., Weisshaar, E., Eds.; S. Karger AG: Basel, Switzerland, 2016; Volume 50, pp. 24–28. ISBN 978-3-318-05888-8. [Google Scholar]
- Carter, M.C.; Metcalfe, D.D.; Komarow, H.D. Mastocytosis. Immunol. Allergy Clin. N. Am. 2014, 34, 181–196. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mast Cells, Glia and Neuroinflammation: Partners in Crime?—PMC. Available online: https://www.ncbi.nlm.nih.gov/pmc/articles/PMC3930370/ (accessed on 20 December 2022).
- Pentland, A.P.; Mahoney, M.; Jacobs, S.C.; Holtzman, M.J. Enhanced Prostaglandin Synthesis after Ultraviolet Injury Is Mediated by Endogenous Histamine Stimulation. A Mechanism for Irradiation Erythema. J. Clin. Investig. 1990, 86, 566–574. [Google Scholar] [CrossRef] [Green Version]
- Matito, A.; Azaña, J.M.; Torrelo, A.; Alvarez-Twose, I. Cutaneous Mastocytosis in Adults and Children. Immunol. Allergy Clin. N. Am. 2018, 38, 351–363. [Google Scholar] [CrossRef] [PubMed]
- Kleewein, K.; Lang, R.; Diem, A.; Vogel, T.; Pohla-Gubo, G.; Bauer, J.W.; Hintner, H.; Laimer, M. Diffuse Cutaneous Mastocytosis Masquerading as Epidermolysis Bullosa. Pediatr. Dermatol. 2011, 28, 720–725. [Google Scholar] [CrossRef] [PubMed]
- Lange, M.; Niedoszytko, M.; Nedoszytko, B.; Łata, J.; Trzeciak, M.; Biernat, W. Diffuse Cutaneous Mastocytosis: Analysis of 10 Cases and a Brief Review of the Literature: Diffuse Cutaneous Mastocytosis. J. Eur. Acad. Dermatol. Venereol. 2011, 26, 1565–1571. [Google Scholar] [CrossRef] [PubMed]
- Lange, M.; Niedoszytko, M.; Renke, J.; Gleń, J.; Nedoszytko, B. Clinical Aspects of Paediatric Mastocytosis: A Review of 101 Cases: Clinical Aspects of Paediatric Mastocytosis. J. Eur. Acad. Dermatol. Venereol. 2013, 27, 97–102. [Google Scholar] [CrossRef]
- Brockow, K.; Ring, J.; Alvarez-Twose, I.; Orfao, A.; Escribano, L. Extensive Blistering Is a Predictor for Severe Complications in Children with Mastocytosis. Allergy 2012, 67, 1323–1324. [Google Scholar] [CrossRef]
- Hoermann, G.; Cerny-Reiterer, S.; Perné, A.; Klauser, M.; Hoetzenecker, K.; Klein, K.; Müllauer, L.; Gröger, M.; Nijman, S.M.B.; Klepetko, W.; et al. Identification of Oncostatin M as a STAT5-Dependent Mediator of Bone Marrow Remodeling in KIT D816V-Positive Systemic Mastocytosis. Am. J. Pathol. 2011, 178, 2344–2356. [Google Scholar] [CrossRef] [Green Version]
- Metcalfe, D.D. Mast Cells and Mastocytosis. Blood 2008, 112, 946–956. [Google Scholar] [CrossRef] [Green Version]
- Valent, P. KIT D816V and the Cytokine Storm in Mastocytosis: Production and Role of Interleukin-6. Haematologica 2020, 105, 5–6. [Google Scholar] [CrossRef] [PubMed]
- Tobío, A.; Bandara, G.; Morris, D.A.; Kim, D.-K.; O’Connell, M.P.; Komarow, H.D.; Carter, M.C.; Smrz, D.; Metcalfe, D.D.; Olivera, A. Oncogenic D816V-KIT Signaling in Mast Cells Causes Persistent IL-6 Production. Haematologica 2020, 105, 124–135. [Google Scholar] [CrossRef] [PubMed]
- Greiner, G.; Witzeneder, N.; Berger, A.; Schmetterer, K.; Eisenwort, G.; Schiefer, A.-I.; Roos, S.; Popow-Kraupp, T.; Müllauer, L.; Zuber, J.; et al. CCL2 Is a KIT D816V–Dependent Modulator of the Bone Marrow Microenvironment in Systemic Mastocytosis. Blood 2017, 129, 371–382. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ahn, Y.M.; Hong, G.U.; Kim, S.H.; Lee, H.J.; Baek, H.S.; Kim, M.N.; Park, K.Y.; Ro, J.Y. Transglutaminase 2 Expressed in Mast Cells Recruited into Skin or Bone Marrow Induces the Development of Pediatric Mastocytosis. Pediatr. Allergy Immunol. 2015, 26, 438–445. [Google Scholar] [CrossRef]
- Lauritano, D.; Ronconi, G.; Caraffa, A.; Enrica Gallenga, C.; Kritas, S.K.; Di Emidio, P.; Martinotti, S.; Tetè, G.; Ross, R.; Conti, P. New Aspect of Allergic Contact Dermatitis, an Inflammatory Skin Disorder Mediated by Mast Cells: Can IL-38 Help? Med. Hypotheses 2020, 139, 109687. [Google Scholar] [CrossRef]
- Ayasse, M.T.; Buddenkotte, J.; Alam, M.; Steinhoff, M. Role of Neuroimmune Circuits and Pruritus in Psoriasis. Exp. Dermatol. 2020, 29, 414–426. [Google Scholar] [CrossRef]
- Voss, M.; Kotrba, J.; Gaffal, E.; Katsoulis-Dimitriou, K.; Dudeck, A. Mast Cells in the Skin: Defenders of Integrity or Offenders in Inflammation? Int. J. Mol. Sci. 2021, 22, 4589. [Google Scholar] [CrossRef]


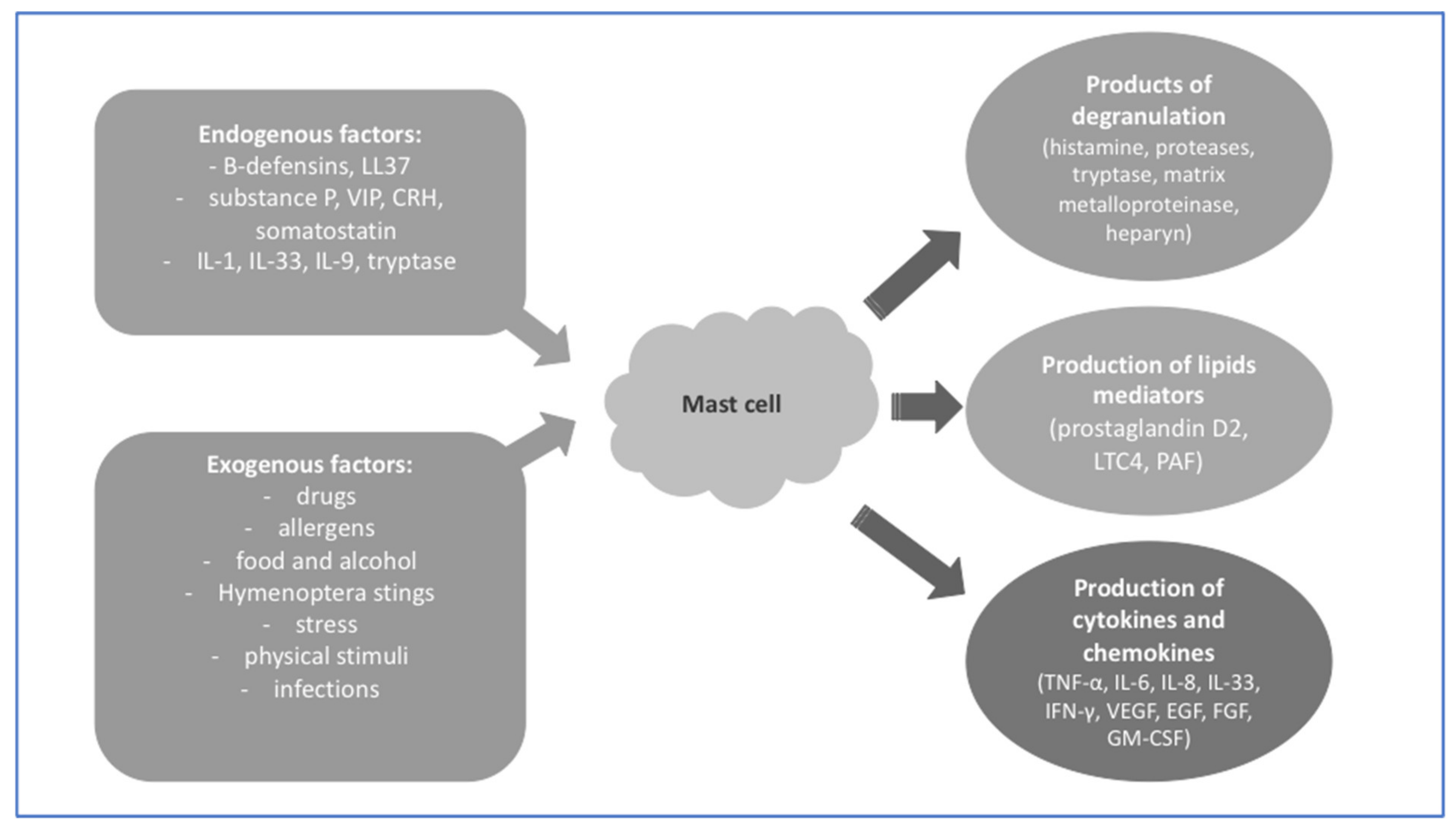{kind=link}
| Genetic factors | Genes encode IL-4 or IL-6, the production of IgE, and/or the regulation of the expression of the IgE receptor on MCs [30,31,32,33]; genes encode IL-13 [30,34]; the KIT mutation D816V [35,36,37]; genes encode angiotensinogen, angiotensin-converting enzymes, and chymase [38,39,40,41]; extra copies of the TPSAB1 gene encoding for alpha tryptase [42,43] |
| Environmental factors | Animal venoms (e.g., Hymenoptera insect, snake, black widow spider, scorpion); hypersensitivity to food (e.g., seafood, tuna fish, salt fish, uncooked anchovies, scombroid, food poisoning); alcohol; medications (e.g., antibiotics, non-steroidal anti-inflammatory drugs (NSAIDs), morphine, neuromuscular blocking agents, radiocontrast media, general anesthesia); physical stimuli (e.g., biopsy, endoscopy, vibration, exercise, pressure, friction, heat); infections (e.g., Ebstein–Barr virus (EBV), varicella zoster virus (VZV), herpes simplex virus (HSV), cytomegalovirus (CMV), human immunodeficiency virus (HIV), influenza, dengue, Ebola, Helicobacter pylori, helminthic parasites); stress [44,45,46,47,48,49,50]. |
| Comorbidities | HαT, SM, cutaneous mastocytosis, IgE-dependent allergy (especially Hymenoptera venom allergy) or other hypersensitivity, atopic disease, vibration urticaria, chronic inflammatory disorders, auto-immune disorders, rheumatic diseases, neurologic diseases, infectious disease [39,40,51,52,53,54,55] |
| Type of Medication | Mechanism of MC Activation |
|---|---|
| Acetylsalicylic acid, NSAID | Inhibition of cyclooxygenase-1 |
| Neuromuscular blocking agents, vespid mastoparan, opioids, icatibant, all fluoroquinolones | Activation of MRGPRX2 |
| Vancomycin, iodinated contrast media | Unknown |
| Liposomal drugs: AmBisome and Doxil, and the micelle-solubilized drug Taxol | Activating the complement system |
| Element of Pathogenesis | Mechanism | Effect |
|---|---|---|
| innate immune responses | a production of LL-37 cathelicidin and LL-37 stimulate MCs to release IL-6 and MMP9 | promote protease production and amplify inflammation |
| neurogenic inflammation | stimulation of MCs by neuropeptides (serotonin receptor 3A, pituitary adenylate cyclase-activating polypeptide, SP, vasoactive intestinal peptide) can cause MC degranulation and the release of histamine, tryptase, and other mediators (TNF-α, CXCL9, CXCL10, CXCL8) | promoting inflammation resulting in itching, flushing, erythema, and burning sensations |
| vasodilation and angiogenesis | a production of proangiogenic molecules (VEGF, fibroblast growth factor—FGF, histamine, and tryptase) | stimulation of the migration and proliferation of endothelial cells, leads to facilitating vascularization and angiogenesis; |
| histamine and serotonin can bind to the vascular receptors, causing vasodilation and increased vascular permeability; vasodilation leads to erythema, and increased vascular permeability causes flushing | ||
| fibrosing | histamine and tryptase enhance fibrosis through their chemotactic function on fibroblasts and MMPs; the proliferation of fibroblasts through VEGF and basic FGF | manifestation of phyma |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Woźniak, E.; Owczarczyk-Saczonek, A.; Lange, M.; Czarny, J.; Wygonowska, E.; Placek, W.; Nedoszytko, B. The Role of Mast Cells in the Induction and Maintenance of Inflammation in Selected Skin Diseases. Int. J. Mol. Sci. 2023, 24, 7021. https://doi.org/10.3390/ijms24087021
Woźniak E, Owczarczyk-Saczonek A, Lange M, Czarny J, Wygonowska E, Placek W, Nedoszytko B. The Role of Mast Cells in the Induction and Maintenance of Inflammation in Selected Skin Diseases. International Journal of Molecular Sciences. 2023; 24(8):7021. https://doi.org/10.3390/ijms24087021
Chicago/Turabian StyleWoźniak, Ewelina, Agnieszka Owczarczyk-Saczonek, Magdalena Lange, Justyna Czarny, Ewa Wygonowska, Waldemar Placek, and Bogusław Nedoszytko. 2023. "The Role of Mast Cells in the Induction and Maintenance of Inflammation in Selected Skin Diseases" International Journal of Molecular Sciences 24, no. 8: 7021. https://doi.org/10.3390/ijms24087021