
Human Male Genital Tract Microbiota
 and
and
Abstract
:1. Genital Microbiota
2. Initial Studies on Bacteria Colonising the MGT
3. Microbiota of the MGT
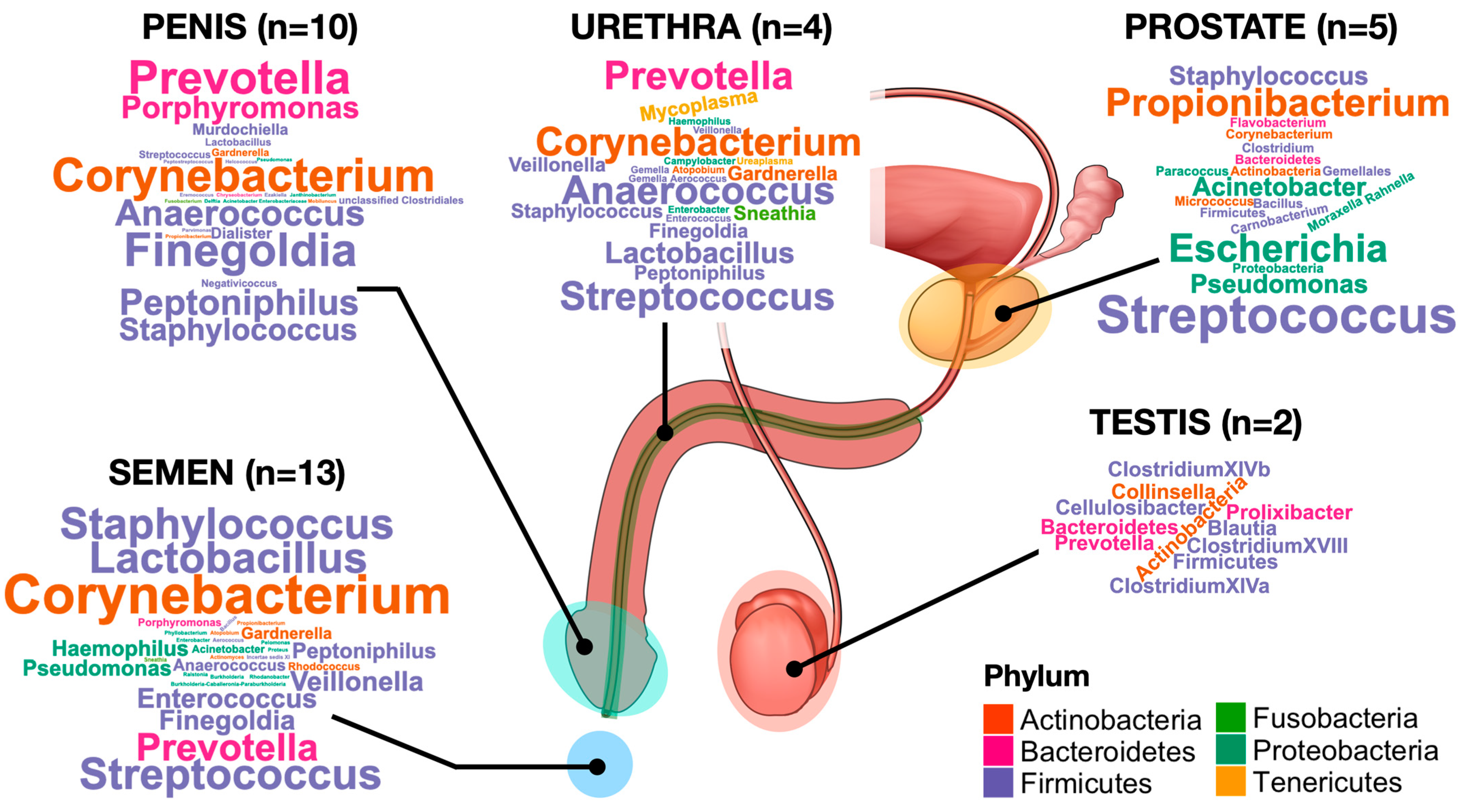
3.1. Testes, Epididymis and Vas Deferens
3.2. Accessory Glands
3.3. Colonisation of the Urethra
3.4. Penis Glans and Coronary Sulcus
3.5. Semen Microbiota
4. How May Genital Microbiota Impair Male Fertility?
5. Future Directions and Missing Gaps
Author Contributions
Funding
Conflicts of Interest
References
- Moeller, A.H.; Caro-Quintero, A.; Mjungu, D.; Georgiev, A.V.; Lonsdorf, E.V.; Muller, M.N.; Pusey, A.E.; Peeters, M.; Hahn, B.H.; Ochman, H. Cospeciation of Gut Microbiota with Hominids. Science 2016, 353, 380–382. [Google Scholar] [CrossRef] [Green Version]
- van de Guchte, M.; Blottière, H.M.; Doré, J. Humans as Holobionts: Implications for Prevention and Therapy. Microbiome 2018, 6, 81. [Google Scholar] [CrossRef]
- Lloyd-Price, J.; Arze, C.; Ananthakrishnan, A.N.; Schirmer, M.; Avila-Pacheco, J.; Poon, T.W.; Andrews, E.; Ajami, N.J.; Bonham, K.S.; Brislawn, C.J.; et al. Multi-Omics of the Gut Microbial Ecosystem in Inflammatory Bowel Diseases. Nature 2019, 569, 655–662. [Google Scholar] [CrossRef]
- Valdes, A.M.; Walter, J.; Segal, E.; Spector, T.D. Role of the Gut Microbiota in Nutrition and Health. BMJ 2018, 361, 2179. [Google Scholar] [CrossRef] [Green Version]
- Gilbert, J.A.; Blaser, M.J.; Caporaso, J.G.; Jansson, J.K.; Lynch, S.V.; Knight, R. Current Understanding of the Human Microbiome. Nat. Med. 2018, 24, 392–400. [Google Scholar] [CrossRef]
- Dominguez-Bello, M.G.; Godoy-Vitorino, F.; Knight, R.; Blaser, M.J. Role of the Microbiome in Human Development. Gut 2019, 68, 1108–1114. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gupta, P.; Singh, M.P.; Goyal, K. Diversity of Vaginal Microbiome in Pregnancy: Deciphering the Obscurity. Front. Public. Health 2020, 8, 326. [Google Scholar] [CrossRef] [PubMed]
- O’Callaghan, J.L.; Turner, R.; Dekker Nitert, M.; Barrett, H.L.; Clifton, V.; Pelzer, E.S. Re-Assessing Microbiomes in the Low-Biomass Reproductive Niche. BJOG 2020, 127, 147–158. [Google Scholar] [CrossRef]
- Heil, B.A.; Paccamonti, D.L.; Sones, J.L. Role for the Mammalian Female Reproductive Tract Microbiome in Pregnancy Outcomes. Physiol. Genom. 2019, 51, 390–399. [Google Scholar] [CrossRef] [PubMed]
- Amabebe, E.; Anumba, D.O.C. The Vaginal Microenvironment: The Physiologic Role of Lactobacilli. Front. Med. 2018, 5, 181. [Google Scholar] [CrossRef] [Green Version]
- Peric, A.; Weiss, J.; Vulliemoz, N.; Baud, D.; Stojanov, M. Bacterial Colonization of the Female Upper Genital Tract. Int. J. Mol. Sci. 2019, 20, 3405. [Google Scholar] [CrossRef] [Green Version]
- Onywera, H.; Williamson, A.-L.; Ponomarenko, J.; Meiring, T.L. The Penile Microbiota in Uncircumcised and Circumcised Men: Relationships With HIV and Human Papillomavirus Infections and Cervicovaginal Microbiota. Front. Med. (Lausanne) 2020, 7, 383. [Google Scholar] [CrossRef] [PubMed]
- Baud, D.; Pattaroni, C.; Vulliemoz, N.; Castella, V.; Marsland, B.J.; Stojanov, M. Sperm Microbiota and Its Impact on Semen Parameters. Front. Microbiol. 2019, 10, 234. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ding, N.; Zhang, X.; Zhang, X.D.; Jing, J.; Liu, S.S.; Mu, Y.P.; Peng, L.L.; Yan, Y.J.; Xiao, G.M.; Bi, X.Y.; et al. Impairment of Spermatogenesis and Sperm Motility by the High-Fat Diet-Induced Dysbiosis of Gut Microbes. Gut 2020, 69, 1608–1619. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Alfano, M.; Ferrarese, R.; Locatelli, I.; Ventimiglia, E.; Ippolito, S.; Gallina, P.; Cesana, D.; Canducci, F.; Pagliardini, L.; Viganò, P.; et al. Testicular Microbiome in Azoospermic Men-First Evidence of the Impact of an Altered Microenvironment. Hum. Reprod. 2018, 33, 1212–1217. [Google Scholar] [CrossRef]
- Putman, S.B.; Chanmugam, A.S. Urethritis, Prostatitis, and Epididymitis. In Infectious Diseases Emergencies; Chanmugam, A.S., Rothman, R., Desai, S., Putman, S., Eds.; Oxford University Press: Oxford, UK, 2016; ISBN 978-0-19-997680-5. [Google Scholar]
- Lam, J.C.; Lang, R.; Stokes, W. How I Manage Bacterial Prostatitis. Clin. Microbiol. Infect. 2023, 29, 32–37. [Google Scholar] [CrossRef]
- Bacterial Prostatitis: Current Opinion in Infectious Diseases. Available online: https://journals.lww.com/co-infectiousdiseases/Fulltext/2016/02000/Bacterial_prostatitis.15.aspx (accessed on 12 February 2023).
- Farahani, L.; Tharakan, T.; Yap, T.; Ramsay, J.W.; Jayasena, C.N.; Minhas, S. The Semen Microbiome and Its Impact on Sperm Function and Male Fertility: A Systematic Review and Meta-Analysis. Andrology 2021, 9, 115–144. [Google Scholar] [CrossRef]
- Jarvi, K.; Lacroix, J.M.; Jain, A.; Dumitru, I.; Heritz, D.; Mittelman, M.W. Polymerase Chain Reaction-Based Detection of Bacteria in Semen. Fertil. Steril. 1996, 66, 463–467. [Google Scholar] [CrossRef]
- Lacroix, J.-M.; Jarvi, K.; Batra, S.D.; Heritz, D.M.; Mittelman, M.W. PCR-Based Technique for the Detection of Bacteria in Semen and Urine. J. Microbiol. Methods 1996, 26, 61–71. [Google Scholar] [CrossRef]
- Theiler, T.; Olaru, I.D.; Kilzer, C.; Schuler, F.; Schaumburg, F. Ejaculate for Microbiological Culture: To Wash or Not To Wash? Microbiol. Spectr. 2022, 10, e0326922. [Google Scholar] [CrossRef]
- James, E.R.; Carrell, D.T.; Aston, K.I.; Jenkins, T.G.; Yeste, M.; Salas-Huetos, A. The Role of the Epididymis and the Contribution of Epididymosomes to Mammalian Reproduction. Int. J. Mol. Sci. 2020, 21, 5377. [Google Scholar] [CrossRef] [PubMed]
- Wang, S.; Zhang, K.; Yao, Y.; Li, J.; Deng, S. Bacterial Infections Affect Male Fertility: A Focus on the Oxidative Stress-Autophagy Axis. Front. Cell. Dev. Biol. 2021, 9, 727812. [Google Scholar] [CrossRef] [PubMed]
- Nistal, M.; González-Peramato, P. Infectious Disease of the Testis. In Encyclopedia of Pathology; van Krieken, J.H.J.M., Ed.; Encyclopedia of Pathology; Springer International Publishing: Cham, Switzerland, 2019; pp. 1–5. ISBN 978-3-319-28845-1. [Google Scholar]
- Molina, N.M.; Plaza-Díaz, J.; Vilchez-Vargas, R.; Sola-Leyva, A.; Vargas, E.; Mendoza-Tesarik, R.; Galán-Lázaro, M.; Mendoza-Ladrón de Guevara, N.; Tesarik, J.; Altmäe, S. Assessing the Testicular Sperm Microbiome: A Low-Biomass Site with Abundant Contamination. Reprod. BioMed. Online 2021, 43, 523–531. [Google Scholar] [CrossRef]
- Weng, S.-L.; Chiu, C.-M.; Lin, F.-M.; Huang, W.-C.; Liang, C.; Yang, T.; Yang, T.-L.; Liu, C.-Y.; Wu, W.-Y.; Chang, Y.-A.; et al. Bacterial Communities in Semen from Men of Infertile Couples: Metagenomic Sequencing Reveals Relationships of Seminal Microbiota to Semen Quality. PLoS ONE 2014, 9, e110152. [Google Scholar] [CrossRef] [Green Version]
- Suarez Arbelaez, M.C.; Israeli, J.M.; Tipton, C.D.; Loloi, J.; Deebel, N.; Leong, J.Y.; Ramasamy, R. Pilot Study: Next-Generation Sequencing of the Semen Microbiome in Vasectomized Versus Nonvasectomized Men. Eur. Urol. Focus. 2022, 11, 10. [Google Scholar] [CrossRef]
- Rizzo, A.; Santoni, M.; Mollica, V.; Fiorentino, M.; Brandi, G.; Massari, F. Microbiota and Prostate Cancer. Semin. Cancer Biol. 2022, 86, 1058–1065. [Google Scholar] [CrossRef] [PubMed]
- Sung, H.; Ferlay, J.; Siegel, R.L.; Laversanne, M.; Soerjomataram, I.; Jemal, A.; Bray, F. Global Cancer Statistics 2020: GLOBOCAN Estimates of Incidence and Mortality Worldwide for 36 Cancers in 185 Countries. CA Cancer J. Clin. 2021, 71, 209–249. [Google Scholar] [CrossRef]
- Verze, P.; Cai, T.; Lorenzetti, S. The Role of the Prostate in Male Fertility, Health and Disease. Nat. Rev. Urol. 2016, 13, 379–386. [Google Scholar] [CrossRef] [Green Version]
- Lee, J.C. Microbiology of the Prostate. Curr. Urol. Rep. 2000, 1, 159–163. [Google Scholar] [CrossRef]
- Berger, R.E.; Krieger, J.N.; Rothman, I.; Muller, C.H.; Hillier, S.L. Bacteria in the Prostate Tissue of Men With Idiopathic Prostatic Inflammation. J. Urol. 1997, 157, 863–865. [Google Scholar] [CrossRef]
- Cavarretta, I.; Ferrarese, R.; Cazzaniga, W.; Saita, D.; Lucianò, R.; Ceresola, E.R.; Locatelli, I.; Visconti, L.; Lavorgna, G.; Briganti, A.; et al. The Microbiome of the Prostate Tumor Microenvironment. Eur. Urol. 2017, 72, 625–631. [Google Scholar] [CrossRef] [PubMed]
- Yow, M.A.; Tabrizi, S.N.; Severi, G.; Bolton, D.M.; Pedersen, J.; Giles, G.G.; Southey, M.C. Australian Prostate Cancer BioResource Characterisation of Microbial Communities within Aggressive Prostate Cancer Tissues. Infect. Agents Cancer 2017, 12, 4. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Feng, Y.; Ramnarine, V.R.; Bell, R.; Volik, S.; Davicioni, E.; Hayes, V.M.; Ren, S.; Collins, C.C. Metagenomic and Metatranscriptomic Analysis of Human Prostate Microbiota from Patients with Prostate Cancer. BMC Genom. 2019, 20, 146. [Google Scholar] [CrossRef] [Green Version]
- Jain, S.; Samal, A.G.; Das, B.; Pradhan, B.; Sahu, N.; Mohapatra, D.; Behera, P.K.; Satpathi, P.S.; Mohanty, A.K.; Satpathi, S.; et al. Escherichia Coli, a Common Constituent of Benign Prostate Hyperplasia-Associated Microbiota Induces Inflammation and DNA Damage in Prostate Epithelial Cells. Prostate 2020, 80, 1341–1352. [Google Scholar] [CrossRef] [PubMed]
- Wu, Y.; Jiang, H.; Tan, M.; Lu, X. Screening for Chronic Prostatitis Pathogens Using High-Throughput next-Generation Sequencing. Prostate 2020, 80, 577–587. [Google Scholar] [CrossRef] [Green Version]
- Okada, K.; Takezawa, K.; Tsujimura, G.; Imanaka, T.; Kuribayashi, S.; Ueda, N.; Hatano, K.; Fukuhara, S.; Kiuchi, H.; Fujita, K.; et al. Localization and Potential Role of Prostate Microbiota. Front. Cell. Infect. Microbiol. 2022, 12, 1776. [Google Scholar] [CrossRef]
- Lei, H.; Han, H.; Feng, Y.; Zhang, X.; Xin, Z.; Tian, L. Altered Microbiota Profile in Seminal Vesicles of Men Presenting with Refractory Hematospermia. Mol. Biol. Rep. 2022, 50, 2381–2389. [Google Scholar] [CrossRef]
- Dong, Q.; Nelson, D.E.; Toh, E.; Diao, L.; Gao, X.; Fortenberry, J.D.; Van Der Pol, B. The Microbial Communities in Male First Catch Urine Are Highly Similar to Those in Paired Urethral Swab Specimens. PLoS ONE 2011, 6, e19709. [Google Scholar] [CrossRef] [Green Version]
- Hrbacek, J.; Morais, D.; Cermak, P.; Hanacek, V.; Zachoval, R. Alpha-Diversity and Microbial Community Structure of the Male Urinary Microbiota Depend on Urine Sampling Method. Sci. Rep. 2021, 11, 23758. [Google Scholar] [CrossRef]
- Plummer, E.L.; Ratten, L.K.; Vodstrcil, L.A.; Murray, G.L.; Danielewski, J.A.; Fairley, C.K.; Garland, S.M.; Chow, E.P.F.; Bradshaw, C.S. The Urethral Microbiota of Men with and without Idiopathic Urethritis. mBio 2022, 13, e0221322. [Google Scholar] [CrossRef]
- Roth, R.S.; Liden, M.; Huttner, A. The Urobiome in Men and Women: A Clinical Review. Clin. Microbiol. Infect. 2022. [Google Scholar] [CrossRef]
- Nelson, D.E.; Dong, Q.; Pol, B.V.D.; Toh, E.; Fan, B.; Katz, B.P.; Mi, D.; Rong, R.; Weinstock, G.M.; Sodergren, E.; et al. Bacterial Communities of the Coronal Sulcus and Distal Urethra of Adolescent Males. PLoS ONE 2012, 7, e36298. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zozaya, M.; Ferris, M.J.; Siren, J.D.; Lillis, R.; Myers, L.; Nsuami, M.J.; Eren, A.M.; Brown, J.; Taylor, C.M.; Martin, D.H. Bacterial Communities in Penile Skin, Male Urethra, and Vaginas of Heterosexual Couples with and without Bacterial Vaginosis. Microbiome 2016, 4, 16. [Google Scholar] [CrossRef] [Green Version]
- Liu, C.M.; Hungate, B.A.; Tobian, A.A.R.; Ravel, J.; Prodger, J.L.; Serwadda, D.; Kigozi, G.; Galiwango, R.M.; Nalugoda, F.; Keim, P.; et al. Penile Microbiota and Female Partner Bacterial Vaginosis in Rakai, Uganda. mBio 2015, 6, e00589-15. [Google Scholar] [CrossRef] [Green Version]
- Onywera, H.; Williamson, A.-L.; Cozzuto, L.; Bonnin, S.; Mbulawa, Z.Z.A.; Coetzee, D.; Ponomarenko, J.; Meiring, T.L. The Penile Microbiota of Black South African Men: Relationship with Human Papillomavirus and HIV Infection. BMC Microbiol. 2020, 20, 78. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Plummer, E.L.; Vodstrcil, L.A.; Doyle, M.; Danielewski, J.A.; Murray, G.L.; Fehler, G.; Fairley, C.K.; Bulach, D.M.; Garland, S.M.; Chow, E.P.F.; et al. A Prospective, Open-Label Pilot Study of Concurrent Male Partner Treatment for Bacterial Vaginosis. mBio 2021, 12, e02323-21. [Google Scholar] [CrossRef]
- Byrd, A.L.; Belkaid, Y.; Segre, J.A. The Human Skin Microbiome. Nat. Rev. Microbiol. 2018, 16, 143–155. [Google Scholar] [CrossRef]
- Gonçalves, M.F.M.; Fernandes, Â.R.; Rodrigues, A.G.; Lisboa, C. Microbiome in Male Genital Mucosa (Prepuce, Glans, and Coronal Sulcus): A Systematic Review. Microorganisms 2022, 10, 2312. [Google Scholar] [CrossRef] [PubMed]
- Liu, C.M.; Osborne, B.J.W.; Hungate, B.A.; Shahabi, K.; Huibner, S.; Lester, R.; Dwan, M.G.; Kovacs, C.; Contente-Cuomo, T.L.; Benko, E.; et al. The Semen Microbiome and Its Relationship with Local Immunology and Viral Load in HIV Infection. PLoS Pathog. 2014, 10, e1004262. [Google Scholar] [CrossRef]
- Liu, C.M.; Prodger, J.L.; Tobian, A.A.R.; Abraham, A.G.; Kigozi, G.; Hungate, B.A.; Aziz, M.; Nalugoda, F.; Sariya, S.; Serwadda, D.; et al. Penile Anaerobic Dysbiosis as a Risk Factor for HIV Infection. mBio 2017, 8, e00996-17. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Prodger, J.L.; Abraham, A.G.; Tobian, A.A.R.; Park, D.E.; Aziz, M.; Roach, K.; Gray, R.H.; Buchanan, L.; Kigozi, G.; Galiwango, R.M.; et al. Penile Bacteria Associated with HIV Seroconversion, Inflammation, and Immune Cells. JCI Insight 2021, 6, e147363. [Google Scholar] [CrossRef] [PubMed]
- Price, L.B.; Liu, C.M.; Johnson, K.E.; Aziz, M.; Lau, M.K.; Bowers, J.; Ravel, J.; Keim, P.S.; Serwadda, D.; Wawer, M.J.; et al. The Effects of Circumcision on the Penis Microbiome. PLoS ONE 2010, 5, e8422. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liu, C.M.; Hungate, B.A.; Tobian, A.A.R.; Serwadda, D.; Ravel, J.; Lester, R.; Kigozi, G.; Aziz, M.; Galiwango, R.M.; Nalugoda, F.; et al. Male Circumcision Significantly Reduces Prevalence and Load of Genital Anaerobic Bacteria. mBio 2013, 4, e00076. [Google Scholar] [CrossRef] [Green Version]
- Mehta, S.D.; Nandi, D.; Agingu, W.; Green, S.J.; Bhaumik, D.K.; Bailey, R.C.; Otieno, F. Vaginal and Penile Microbiome Associations With Herpes Simplex Virus Type 2 in Women and Their Male Sex Partners. J. Infect. Dis. 2022, 226, 644–654. [Google Scholar] [CrossRef]
- Watchorn, R.E.; van den Munckhof, E.H.A.; Quint, K.D.; Eliahoo, J.; de Koning, M.N.C.; Quint, W.G.V.; Bunker, C.B. Balanopreputial Sac and Urine Microbiota in Patients with Male Genital Lichen Sclerosus. Int. J. Dermatol. 2021, 60, 201–207. [Google Scholar] [CrossRef] [PubMed]
- Mehta, S.D.; Zhao, D.; Green, S.J.; Agingu, W.; Otieno, F.; Bhaumik, R.; Bhaumik, D.; Bailey, R.C. The Microbiome Composition of a Man’s Penis Predicts Incident Bacterial Vaginosis in His Female Sex Partner With High Accuracy. Front. Cell. Infect. Microbiol. 2020, 10, 433. [Google Scholar] [CrossRef]
- Gray, R.H.; Kigozi, G.; Serwadda, D.; Makumbi, F.; Nalugoda, F.; Watya, S.; Moulton, L.; Chen, M.Z.; Sewankambo, N.K.; Kiwanuka, N.; et al. The Effects of Male Circumcision on Female Partners’ Genital Tract Symptoms and Vaginal Infections in a Randomized Trial in Rakai, Uganda. Am. J. Obs. Gynecol. 2009, 200, 42.e1-7. [Google Scholar] [CrossRef] [Green Version]
- Prodger, J.L.; Kaul, R. The Biology of How Circumcision Reduces HIV Susceptibility: Broader Implications for the Prevention Field. AIDS Res. Ther. 2017, 14, 49. [Google Scholar] [CrossRef] [Green Version]
- Tuominen, H.; Rautava, J.; Kero, K.; Syrjänen, S.; Collado, M.C.; Rautava, S. HPV Infection and Bacterial Microbiota in the Semen from Healthy Men. BMC Infect. Dis. 2021, 21, 373. [Google Scholar] [CrossRef]
- Mändar, R.; Punab, M.; Korrovits, P.; Türk, S.; Ausmees, K.; Lapp, E.; Preem, J.-K.; Oopkaup, K.; Salumets, A.; Truu, J. Seminal Microbiome in Men with and without Prostatitis. Int. J. Urol. 2017, 24, 211–216. [Google Scholar] [CrossRef] [Green Version]
- Hou, D.; Zhou, X.; Zhong, X.; Settles, M.L.; Herring, J.; Wang, L.; Abdo, Z.; Forney, L.J.; Xu, C. Microbiota of the Seminal Fluid from Healthy and Infertile Men. Fertil. Steril. 2013, 100, 1261–1269. [Google Scholar] [CrossRef] [Green Version]
- Mändar, R.; Punab, M.; Borovkova, N.; Lapp, E.; Kiiker, R.; Korrovits, P.; Metspalu, A.; Krjutškov, K.; Nõlvak, H.; Preem, J.-K.; et al. Complementary Seminovaginal Microbiome in Couples. Res. Microbiol. 2015, 166, 440–447. [Google Scholar] [CrossRef] [PubMed]
- Chen, H.; Luo, T.; Chen, T.; Wang, G. Seminal Bacterial Composition in Patients with Obstructive and Non-obstructive Azoospermia. Exp. Ther. Med. 2018, 15, 2884–2890. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Monteiro, C.; Marques, P.I.; Cavadas, B.; Damião, I.; Almeida, V.; Barros, N.; Barros, A.; Carvalho, F.; Gomes, S.; Seixas, S. Characterization of Microbiota in Male Infertility Cases Uncovers Differences in Seminal Hyperviscosity and Oligoasthenoteratozoospermia Possibly Correlated with Increased Prevalence of Infectious Bacteria. Am. J. Reprod. Immunol. 2018, 79, e12838. [Google Scholar] [CrossRef]
- Yang, H.; Zhang, J.; Xue, Z.; Zhao, C.; Lei, L.; Wen, Y.; Dong, Y.; Yang, J.; Zhang, L. Potential Pathogenic Bacteria in Seminal Microbiota of Patients with Different Types of Dysspermatism. Sci. Rep. 2020, 10, 6876. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Amato, V.; Papaleo, E.; Pasciuta, R.; Viganò, P.; Ferrarese, R.; Clementi, N.; Sanchez, A.M.; Quaranta, L.; Burioni, R.; Ambrosi, A.; et al. Differential Composition of Vaginal Microbiome, but Not of Seminal Microbiome, Is Associated With Successful Intrauterine Insemination in Couples With Idiopathic Infertility: A Prospective Observational Study. Open. Forum Infect. Dis. 2020, 7, ofz525. [Google Scholar] [CrossRef] [PubMed]
- Štšepetova, J.; Baranova, J.; Simm, J.; Parm, Ü.; Rööp, T.; Sokmann, S.; Korrovits, P.; Jaagura, M.; Rosenstein, K.; Salumets, A.; et al. The Complex Microbiome from Native Semen to Embryo Culture Environment in Human in Vitro Fertilization Procedure. Reprod. Biol. Endocrinol. 2020, 18, 3. [Google Scholar] [CrossRef]
- Lundy, S.D.; Sangwan, N.; Parekh, N.V.; Selvam, M.K.P.; Gupta, S.; McCaffrey, P.; Bessoff, K.; Vala, A.; Agarwal, A.; Sabanegh, E.S.; et al. Functional and Taxonomic Dysbiosis of the Gut, Urine, and Semen Microbiomes in Male Infertility. Eur. Urol. 2021. [Google Scholar] [CrossRef]
- Yao, Y.; Qiu, X.-J.; Wang, D.-S.; Luo, J.-K.; Tang, T.; Li, Y.-H.; Zhang, C.-H.; Liu, H.; Zhou, L.; Zhao, L.-L. Semen Microbiota in Normal and Leukocytospermic Males. Asian J. 2021, 24, 398–405. [Google Scholar] [CrossRef]
- Bukharin, O.; Perunova, N.; Ivanova, E.; Chaynikova, I.; Bekpergenova, A.; Bondarenko, T.; Kuzmin, M. Semen Microbiota and Cytokines of Healthy and Infertile Men. Asian J. 2022, 24, 353. [Google Scholar] [CrossRef]
- Onderdonk, A.B.; Delaney, M.L.; Fichorova, R.N. The Human Microbiome during Bacterial Vaginosis. Clin. Microbiol. Rev. 2016, 29, 223–238. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Griswold, M.D. Spermatogenesis: The Commitment to Meiosis. Physiol. Rev. 2016, 96, 1–17. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sullivan, R.; Mieusset, R. The Human Epididymis: Its Function in Sperm Maturation. Hum. Reprod. Update 2016, 22, 574–587. [Google Scholar] [CrossRef] [Green Version]
- Hosseinzadeh, S.; Eley, A.; Pacey, A.A. Semen Quality of Men with Asymptomatic Chlamydial Infection. J. Androl. 2004, 25, 104–109. [Google Scholar] [CrossRef]
- Ahmadi, M.H.; Mirsalehian, A.; Sadighi Gilani, M.A.; Bahador, A.; Talebi, M. Asymptomatic Infection With Mycoplasma Hominis Negatively Affects Semen Parameters and Leads to Male Infertility as Confirmed by Improved Semen Parameters After Antibiotic Treatment. Urology 2017, 100, 97–102. [Google Scholar] [CrossRef] [PubMed]
- Nunez-Calonge, R.; Caballero, P.; Redondo, C.; Baquero, F.; Martinez-Ferrer, M.; Meseguer, M.A. Ureaplasma Urealyticum Reduces Motility and Induces Membrane Alterations in Human Spermatozoa. Hum. Reprod. 1998, 13, 2756–2761. [Google Scholar] [CrossRef]
- Satta, A.; Stivala, A.; Garozzo, A.; Morello, A.; Perdichizzi, A.; Vicari, E.; Salmeri, M.; Calogero, A.E. Experimental Chlamydia Trachomatis Infection Causes Apoptosis in Human Sperm. Hum. Reprod. 2006, 21, 134–137. [Google Scholar] [CrossRef] [Green Version]
- Hosseinzadeh, S.; Brewis, I.A.; Eley, A.; Pacey, A.A. Co-Incubation of Human Spermatozoa with Chlamydia Trachomatis Serovar E Causes Premature Sperm Death. Hum. Reprod. 2001, 16, 293–299. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Baud, D.; Vulliemoz, N.; Ammerdorffer, A.; Gyger, J.; Greub, G.; Castella, V.; Stojanov, M. Waddlia Chondrophila, a Chlamydia-Related Bacterium, Has a Negative Impact on Human Spermatozoa. Hum. Reprod. 2017, 33, 3–10. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Huang, C.; Long, X.; Jing, S.; Fan, L.; Xu, K.; Wang, S.; Zhu, W. Ureaplasma Urealyticum and Mycoplasma Hominis Infections and Semen Quality in 19,098 Infertile Men in China. World J. Urol. 2016, 34, 1039–1044. [Google Scholar] [CrossRef]
- Eley, A.; Pacey, A.A.; Galdiero, M.; Galdiero, M.; Galdiero, F. Can Chlamydia Trachomatis Directly Damage Your Sperm? Lancet Infect. Dis. 2005, 5, 53–57. [Google Scholar] [CrossRef] [PubMed]
- Svenstrup, H.F.; Fedder, J.; Abraham-Peskir, J.; Birkelund, S.; Christiansen, G. Mycoplasma Genitalium Attaches to Human Spermatozoa. Hum. Reprod. 2003, 18, 2103–2109. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wølner-Hanssen, P.; Mårdh, P.-A. In Vitro Tests of the Adherence of Chlamydia Trachomatis to Human Spermatozoa. Fertil. Steril. 1984, 42, 102–107. [Google Scholar] [CrossRef]
- Erbengi, T. Ultrastructural Observations on the Entry of Chlamydia Trachomatis into Human Spermatozoa. Hum. Reprod. 1993, 8, 416–421. [Google Scholar] [CrossRef]
- Gorga, F.; Galdiero, M.; Buommino, E.; Galdiero, E. Porins and Lipopolysaccharide Induce Apoptosis in Human Spermatozoa. Clin. Diagn. Lab. Immunol. 2001, 8, 206–208. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, Z.; Zhang, D.; He, Y.; Ding, Z.; Mao, F.; Luo, T.; Zhang, X. Lipopolysaccharide Compromises Human Sperm Function by Reducing Intracellular CAMP. Tohoku J. Exp. Med. 2016, 238, 105–112. [Google Scholar] [CrossRef] [Green Version]
- Boguen, R.; Treulen, F.; Uribe, P.; Villegas, J.V. Ability of Escherichia Coli to Produce Hemolysis Leads to a Greater Pathogenic Effect on Human Sperm. Fertil. Steril. 2015, 103, 1155–1161. [Google Scholar] [CrossRef]
- Qiang, H.; Jiang, M.-S.; Lin, J.-Y.; He, W.-M. Influence of Enterococci on Human Sperm Membrane in Vitro. Asian J. 2007, 9, 77–81. [Google Scholar] [CrossRef]
- Prabha, V.; Gupta, T.; Kaur, S.; Kaur, N.; Kala, S.; Singh, A. Isolation of a Spermatozoal Immobilization Factor from Staphylococcus Aureus Filtrates. Can. J. Microbiol. 2009, 55, 874–878. [Google Scholar] [CrossRef]
- Answal, M.; Prabha, V. Escherichia Coli Recombinant Sperm Immobilizing Factor RecX as a Potential Vaginal Contraceptive. Reprod. Biol. Endocrinol. 2018, 16, 88. [Google Scholar] [CrossRef] [Green Version]
- Oghbaei, H.; Rastgar Rezaei, Y.; Nikanfar, S.; Zarezadeh, R.; Sadegi, M.; Latifi, Z.; Nouri, M.; Fattahi, A.; Ahmadi, Y.; Bleisinger, N. Effects of Bacteria on Male Fertility: Spermatogenesis and Sperm Function. Life Sci. 2020, 256, 117891. [Google Scholar] [CrossRef] [PubMed]
- Tvrdá, E.; Ďuračka, M.; Benko, F.; Lukáč, N. Bacteriospermia—A Formidable Player in Male Subfertility. Open. Life Sci. 2022, 17, 1001–1029. [Google Scholar] [CrossRef]
- Eini, F.; Kutenaei, M.A.; Zareei, F.; Dastjerdi, Z.S.; Shirzeyli, M.H.; Salehi, E. Effect of Bacterial Infection on Sperm Quality and DNA Fragmentation in Subfertile Men with Leukocytospermia. BMC Mol. Cell. Biol. 2021, 22, 42. [Google Scholar] [CrossRef] [PubMed]
- Fraczek, M.; Hryhorowicz, M.; Gill, K.; Zarzycka, M.; Gaczarzewicz, D.; Jedrzejczak, P.; Bilinska, B.; Piasecka, M.; Kurpisz, M. The Effect of Bacteriospermia and Leukocytospermia on Conventional and Nonconventional Semen Parameters in Healthy Young Normozoospermic Males. J. Reprod. Immunol. 2016, 118, 18–27. [Google Scholar] [CrossRef]
- Eldamnhoury, E.M.; Elatrash, G.A.; Rashwan, H.M.; El-Sakka, A.I. Association between Leukocytospermia and Semen Interleukin-6 and Tumor Necrosis Factor-Alpha in Infertile Men. Andrology 2018, 6, 775–780. [Google Scholar] [CrossRef] [Green Version]
- Visconti, A.; Le Roy, C.I.; Rosa, F.; Rossi, N.; Martin, T.C.; Mohney, R.P.; Li, W.; de Rinaldis, E.; Bell, J.T.; Venter, J.C.; et al. Interplay between the Human Gut Microbiome and Host Metabolism. Nat. Commun. 2019, 10, 4505. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Franzosa, E.A.; Sirota-Madi, A.; Avila-Pacheco, J.; Fornelos, N.; Haiser, H.J.; Reinker, S.; Vatanen, T.; Hall, A.B.; Mallick, H.; McIver, L.J.; et al. Gut Microbiome Structure and Metabolic Activity in Inflammatory Bowel Disease. Nat. Microbiol. 2019, 4, 293–305. [Google Scholar] [CrossRef]
- O’Hanlon, D.E.; Come, R.A.; Moench, T.R. Vaginal PH Measured in Vivo: Lactobacilli Determine PH and Lactic Acid Concentration. BMC Microbiol. 2019, 19, 13. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- O’Hanlon Vaginal PH and Microbicidal Lactic Acid When Lactobacilli Dominate the Microbiota. Available online: https://pubmed.ncbi.nlm.nih.gov/24223212/ (accessed on 16 September 2020).
- Wang, Y.-X.; Wu, Y.; Chen, H.-G.; Duan, P.; Wang, L.; Shen, H.-Q.; Lu, W.-Q.; Sun, B.; Wang, Q.; Zhang, B.; et al. Seminal Plasma Metabolome in Relation to Semen Quality and Urinary Phthalate Metabolites among Chinese Adult Men. Environ. Int. 2019, 129, 354–363. [Google Scholar] [CrossRef]
- Koeth, R.A.; Wang, Z.; Levison, B.S.; Buffa, J.A.; Org, E.; Sheehy, B.T.; Britt, E.B.; Fu, X.; Wu, Y.; Li, L.; et al. Intestinal Microbiota Metabolism of L-Carnitine, a Nutrient in Red Meat, Promotes Atherosclerosis. Nat. Med. 2013, 19, 576–585. [Google Scholar] [CrossRef] [Green Version]
- Hulme, H.; Meikle, L.M.; Strittmatter, N.; van der Hooft, J.J.J.; Swales, J.; Bragg, R.A.; Villar, V.H.; Ormsby, M.J.; Barnes, S.; Brown, S.L.; et al. Microbiome-Derived Carnitine Mimics as Previously Unknown Mediators of Gut-Brain Axis Communication. Sci. Adv. 2020, 6, eaax6328. [Google Scholar] [CrossRef] [Green Version]
- Perez-Carrasco, V.; Soriano-Lerma, A.; Soriano, M.; Gutiérrez-Fernández, J.; Garcia-Salcedo, J.A. Urinary Microbiome: Yin and Yang of the Urinary Tract. Front. Cell. Infect. Microbiol. 2021, 11, 617002. [Google Scholar] [CrossRef] [PubMed]
- Man, W.H.; de Steenhuijsen Piters, W.A.A.; Bogaert, D. The Microbiota of the Respiratory Tract: Gatekeeper to Respiratory Health. Nat. Rev. Microbiol. 2017, 15, 259–270. [Google Scholar] [CrossRef] [PubMed]
- Eisenhofer, R.; Minich, J.J.; Marotz, C.; Cooper, A.; Knight, R.; Weyrich, L.S. Contamination in Low Microbial Biomass Microbiome Studies: Issues and Recommendations. Trends Microbiol. 2019, 27, 105–117. [Google Scholar] [CrossRef]
- Karstens, L.; Asquith, M.; Davin, S.; Fair, D.; Gregory, W.T.; Wolfe, A.J.; Braun, J.; McWeeney, S. Controlling for Contaminants in Low-Biomass 16S RRNA Gene Sequencing Experiments. mSystems 2019, 4, e00290-19. [Google Scholar] [CrossRef] [Green Version]
- McKnight, D.T.; Huerlimann, R.; Bower, D.S.; Schwarzkopf, L.; Alford, R.A.; Zenger, K.R. MicroDecon: A Highly Accurate Read-Subtraction Tool for the Post-Sequencing Removal of Contamination in Metabarcoding Studies. Environ. DNA 2019, 1, 14–25. [Google Scholar] [CrossRef]
- Davis, N.M.; Proctor, D.M.; Holmes, S.P.; Relman, D.A.; Callahan, B.J. Simple Statistical Identification and Removal of Contaminant Sequences in Marker-Gene and Metagenomics Data. Microbiome 2018, 6, 226. [Google Scholar] [CrossRef] [Green Version]


{kind=link}
| Study | Sequencing Method | Sample Size | Negative Controls | Bacterial Count | Main Findings |
|---|---|---|---|---|---|
| Cavarretta et al., 2017 [34] | Pyrosequencing (V3–V5) | 16 | No | No |
|
| Yow et al., 2017 [35] | Illumina (V2–V3 and V4) | 10 | Yes | No |
|
| Feng et al., 2019 [36] | Illumina (shotgun) | 65 | No | No |
|
| Jain et al., 2020 [37] | Illumina (V3) | 20 | No | No |
|
| Wu et al., 2020 [38] | Illumina (V4) | 63 | No | No |
|
| Study | Sequencing Method | Sample Size | Negative Controls | Bacterial Count | Main Findings |
|---|---|---|---|---|---|
| Price et al., 2010 [55] | Pyrosequencing (V3–V4) | 12 | No | No |
|
| Nelson et al., 2012 [45] | Pyrosequencing (V1–V3, V3–V5 and V6–V9) | 18 | No |
| |
| Liu et al., 2013 [56] | Pyrosequencing (V3–V6) | 156 | No | Yes |
|
| Liu et al., 2015 [47] | Pyrosequencing (V3–V6) | 165 | No | Yes |
|
| Liu et al., 2017 [53] | Illumina (V3–V4) | 182 | No | Yes |
|
| Mehta et al., 2020 [57] | Illumina (V3–V4) | 231 | No | No |
|
| Onywera et al., 2020 [48] | Illumina (V3–V4) | 238 | Yes | No |
|
| Plummer et al., 2021 [49] | Illumina (V3–V4) | 34 | Yes | No |
|
| Watchorn et al., 2021 [58] | Illumina (V3–V4) | 40 | No | Yes |
|
| Prodger et al., 2021 [54] | Pyrosequencing (V3–V6) | 188 | No | Yes |
|
| Mehta et al., 2020 [59] | Illumina (V3–V4) | 168 | Yes | No |
|
| Study | Sequencing Method | Sample Size | Negative Controls | Bacterial Count | Main Findings |
|---|---|---|---|---|---|
| Hou et al., 2013 [64] | Pyrosequencing (V1–V2) | 77 | No | No |
|
| Weng et al., 2014 [27] | Illumina (V4) | 96 | No | No |
|
| Liu et al., 2014 [52] | Pyrosequencing (V3–V6) | 49 | No | No |
|
| Mändar et al., 2015 [65] | Illumina (V6) | 23 | No | Yes |
|
| Mändar et al., 2017 [63] | Illumina (V6) | 67 | No | No |
|
| Chen et al., 2018 [66] | Illumina (V4) | 17 | No | No |
|
| Monteiro et al., 2018 [67] | Ion Torrent (V3–V6) | 118 | No | No |
|
| Baud et al., 2019 [13] | Illumina (V1–V2) | 94 | Yes | Yes |
|
| Yang et al., 2020 [68] | Illumina (V1–V2) | 159 | No | No |
|
| Amato et al., 2020 [69] | Illumina (V3–V4) | 23 | No | Yes |
|
| Štšepetova et al., 2020 [70] | Pyrosequencing (NA) | 50 | Yes | Yes |
|
| Lundy et al., 2021 [71] | Illumina (V3–V4) | 37 | No | NA |
|
| Tuominen et al., 2021 [62] | Illumina (V3–V4) | 31 | No | No |
|
| Yao et al., 2021 [72] | Illumina (V3–V4) | 87 | No | No |
|
| Bukharin et al., 2022 [73] | Illumina (NA) | 72 | No | NA |
|
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zuber, A.; Peric, A.; Pluchino, N.; Baud, D.; Stojanov, M. Human Male Genital Tract Microbiota. Int. J. Mol. Sci. 2023, 24, 6939. https://doi.org/10.3390/ijms24086939
Zuber A, Peric A, Pluchino N, Baud D, Stojanov M. Human Male Genital Tract Microbiota. International Journal of Molecular Sciences. 2023; 24(8):6939. https://doi.org/10.3390/ijms24086939
Chicago/Turabian StyleZuber, Arnaud, Adriana Peric, Nicola Pluchino, David Baud, and Milos Stojanov. 2023. "Human Male Genital Tract Microbiota" International Journal of Molecular Sciences 24, no. 8: 6939. https://doi.org/10.3390/ijms24086939