
Molecular Cytological Analysis and Specific Marker Development in Wheat-Psathyrostachys huashanica Keng 3Ns Additional Line with Elongated Glume
 and
and
Abstract
:1. Introduction
2. Results
2.1. Subsection Agronomic Traits
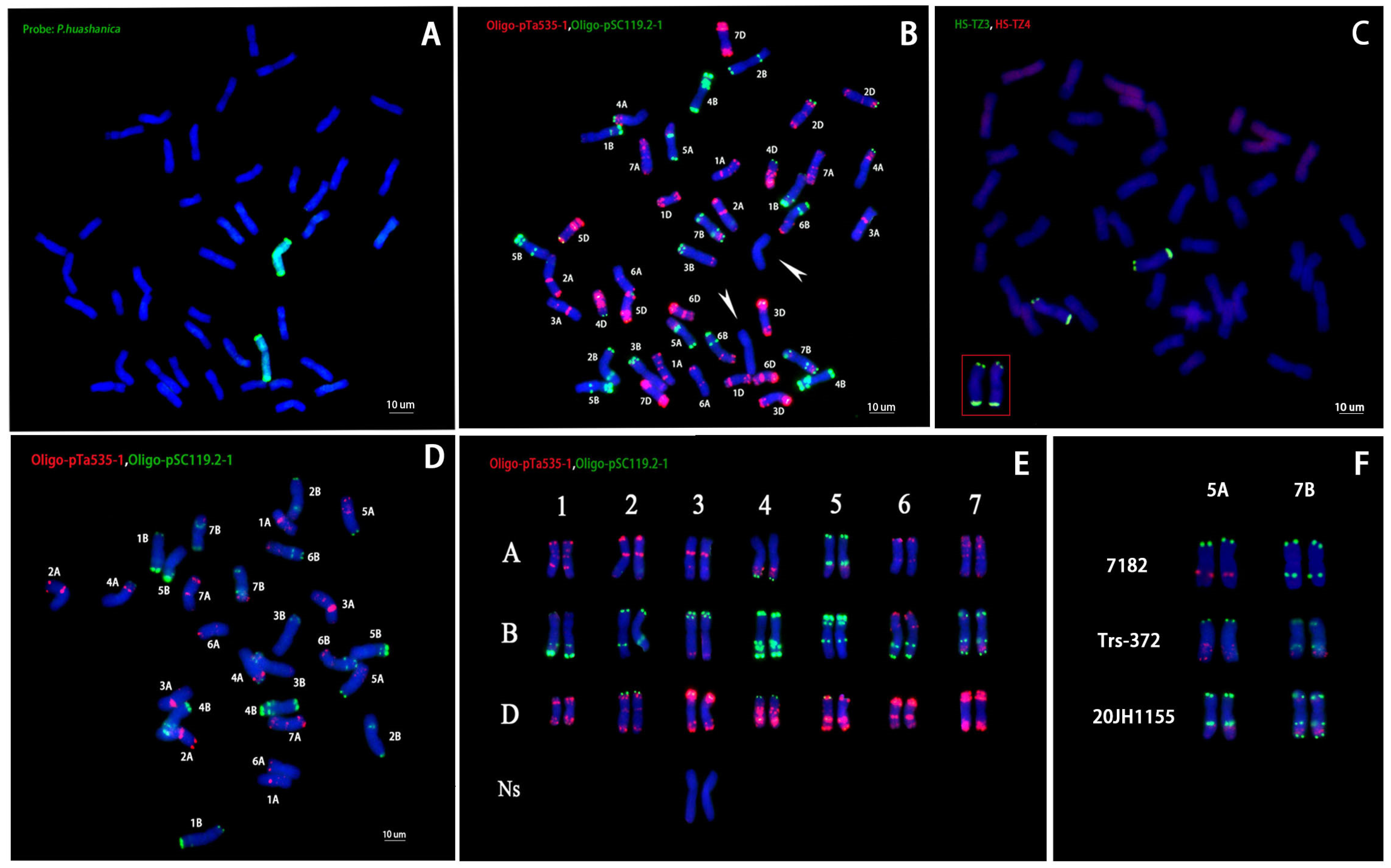
2.2. Observation Cytologenetics and In Situ Hybridization of 20JH1155
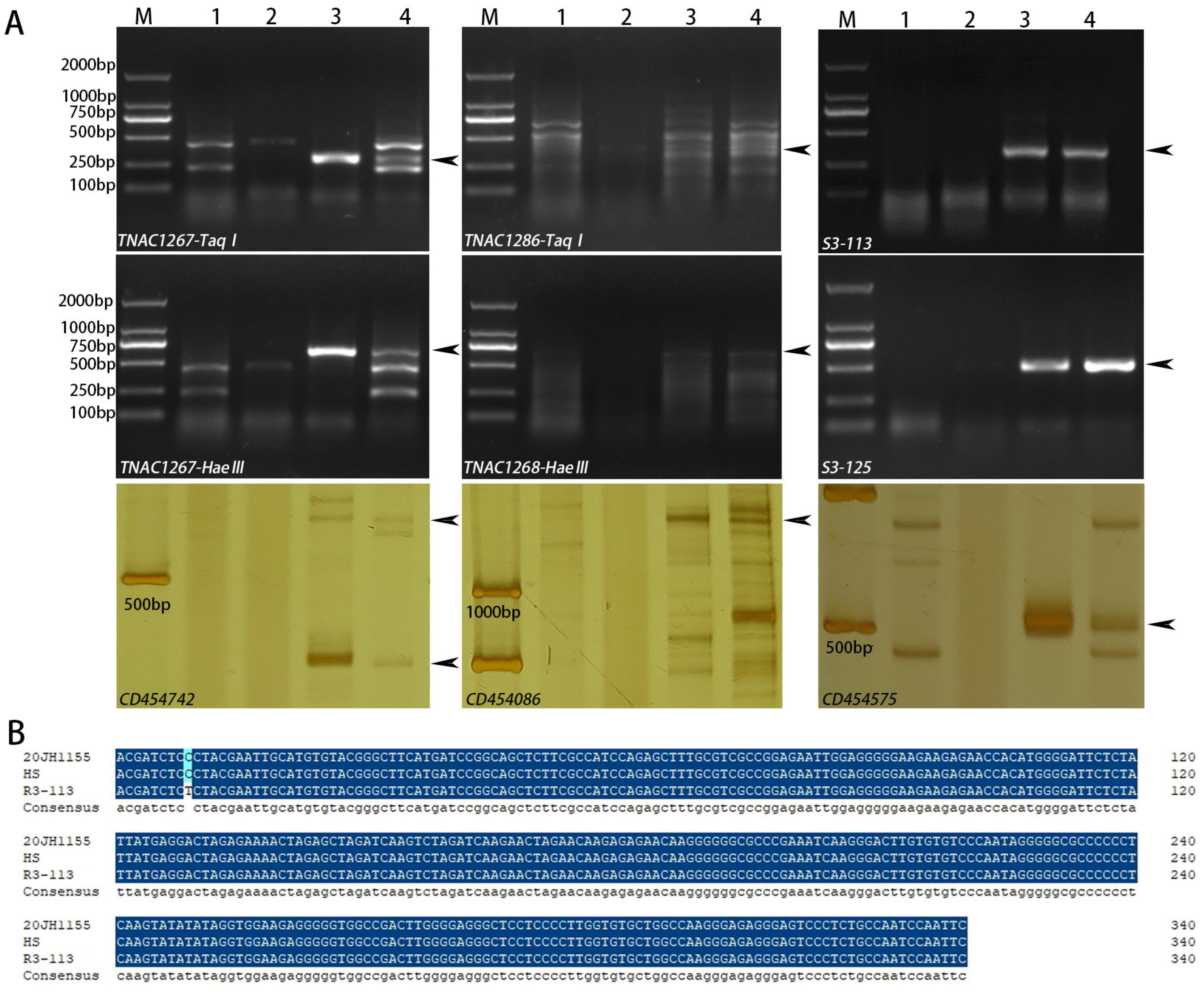
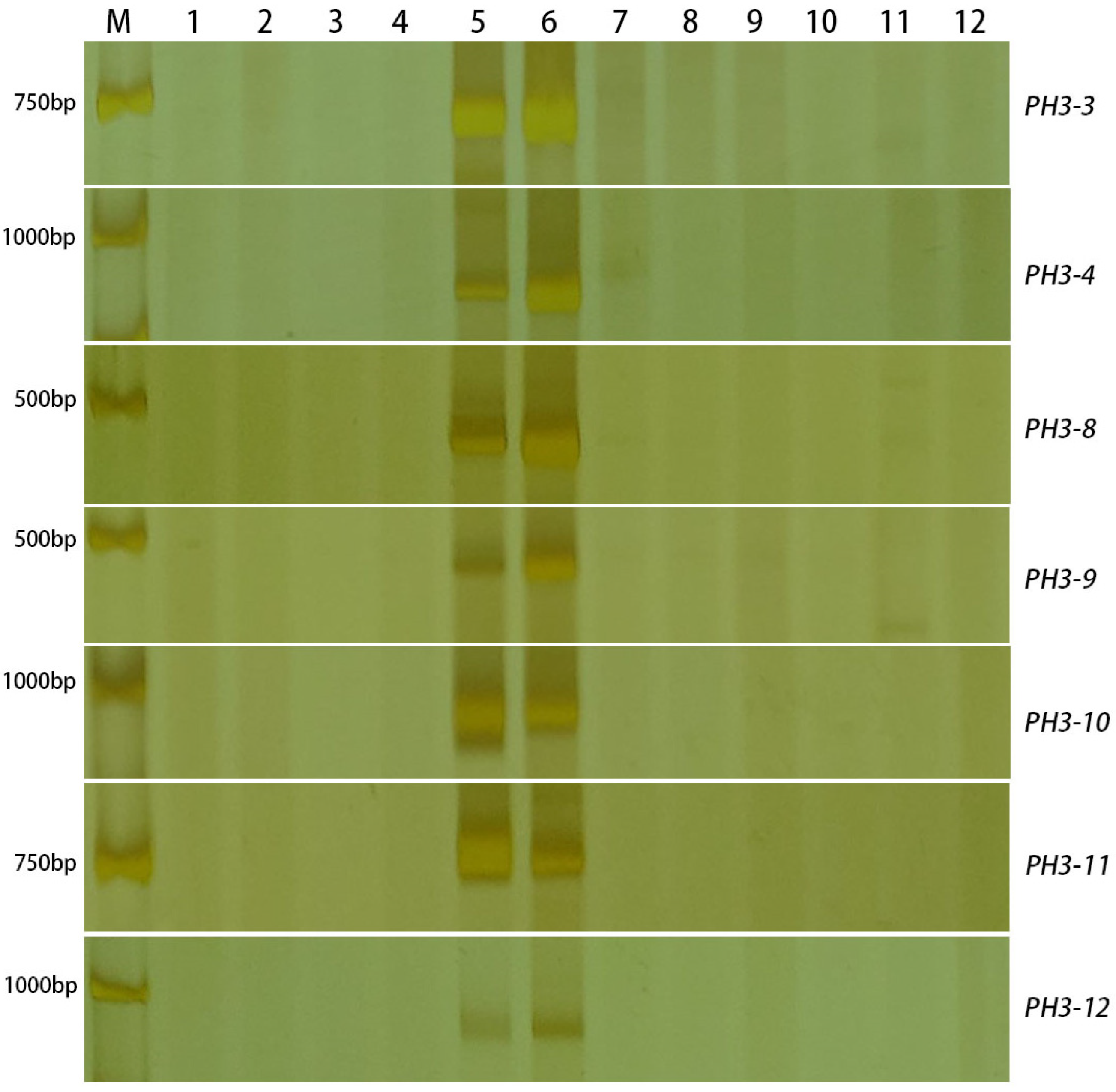
2.3. Molecular Marker Analysis
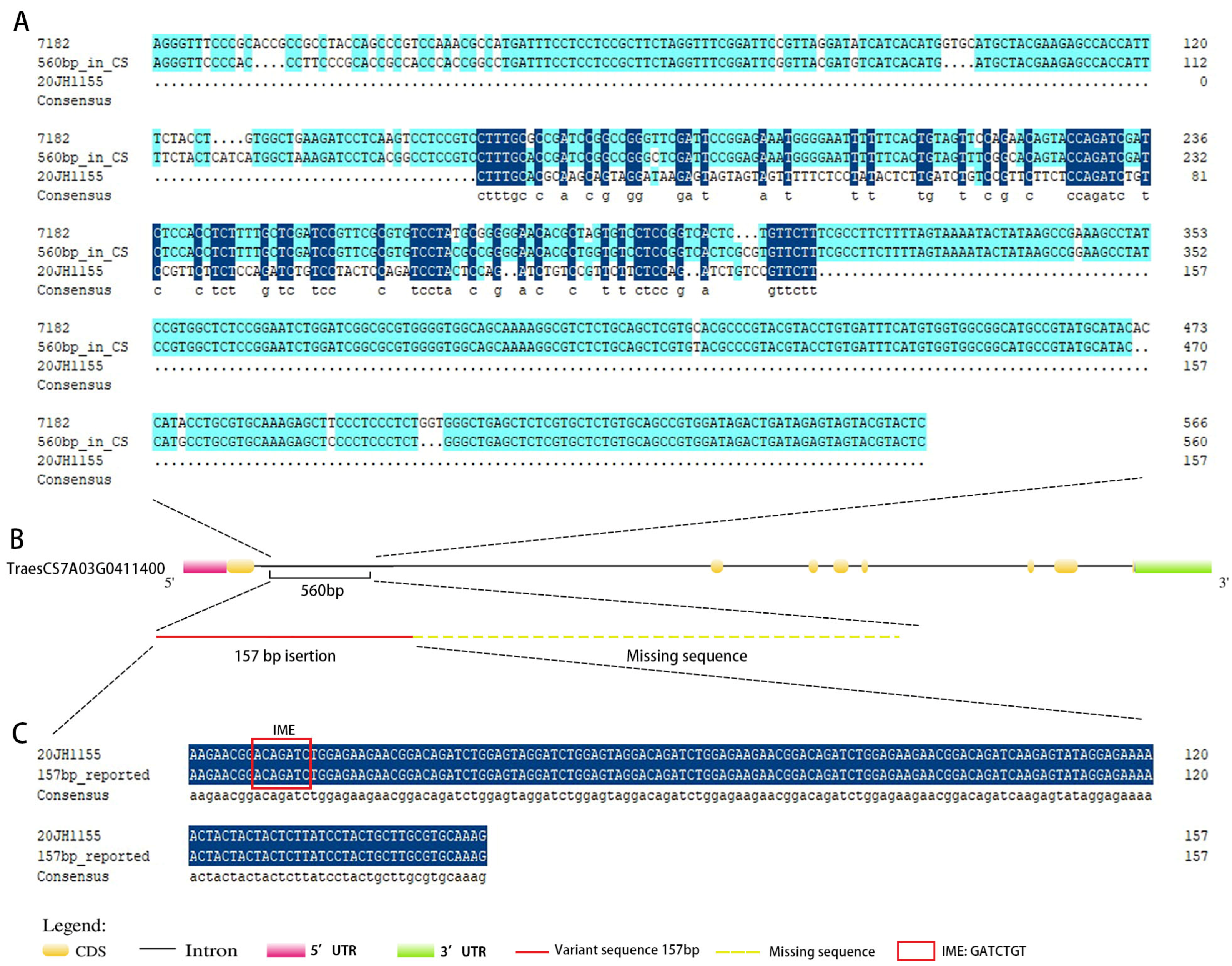
2.4. Identification of the Variant Section of VRT-A2 in 20JH1155
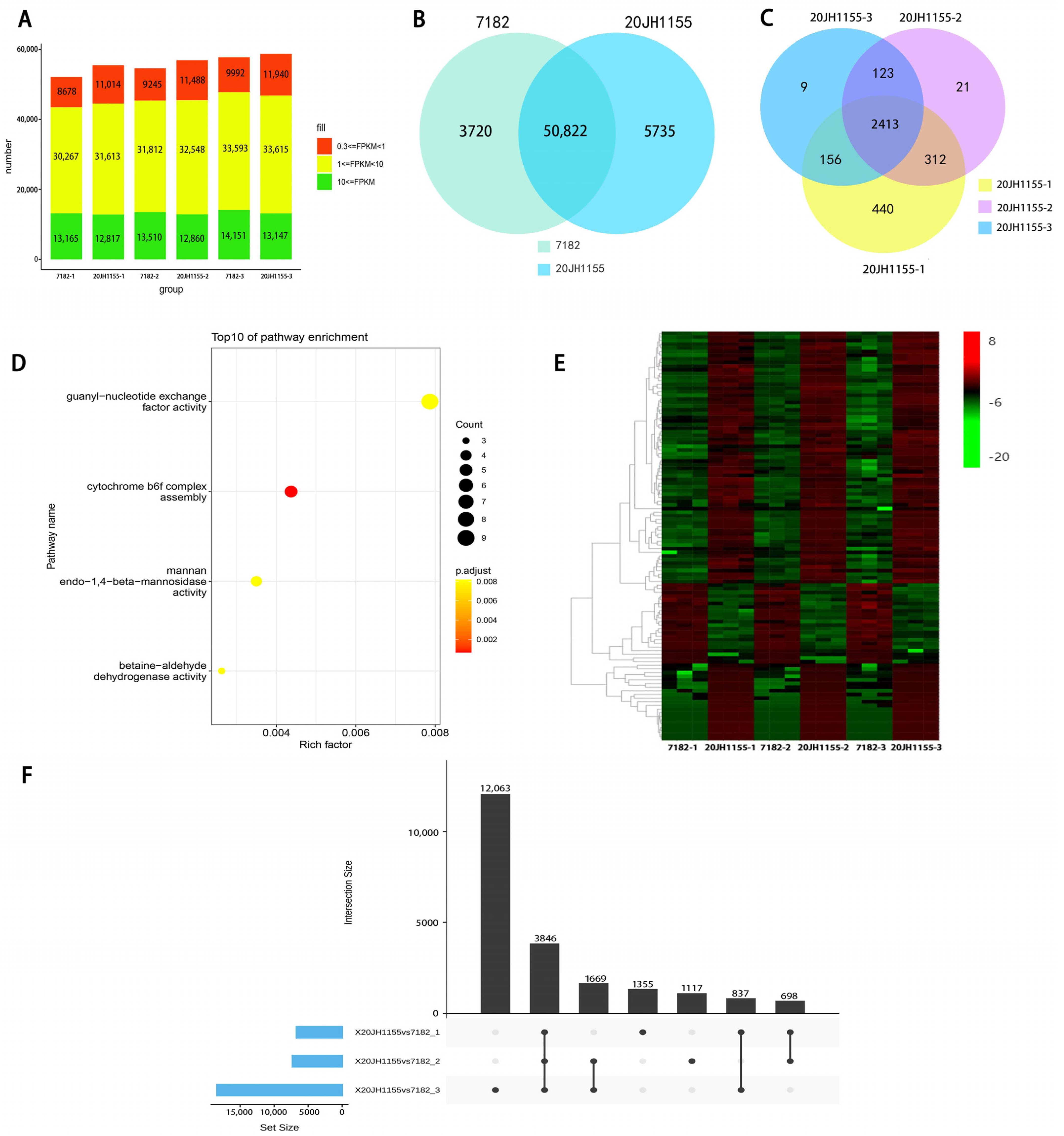
2.5. RNA Expression Level Detection
2.6. Development and Evaluation of P. huashanica Chromosomes Molecular Makers
3. Discussion
4. Materials and Methods
4.1. Plant Materials
4.2. Assessment of Agronomic Traits and Disease Resistance
4.3. Cytological Identification
4.4. GISH and FISH Analysis
4.5. Molecular Marker Analysis
4.6. Variant Sequence Cloning and Analysis of 20JH1155
4.7. RNA Sequencing and Transcriptome Analysis
4.8. Molecular Marker Development Based on RNA-Seq
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Shiferaw, B.; Smale, M.; Braun, H.-J.; Duveiller, E.; Reynolds, M.; Muricho, G. Crops that feed the world 10. Past successes and future challenges to the role played by wheat in global food security. Food Secur. 2013, 5, 291–317. [Google Scholar] [CrossRef] [Green Version]
- Caceres, M.; Pupilli, F.; Ceccarelli, M.; Vaccino, P.; Sarri, V.; De Pace, C.; Cionini, P. Cryptic Introgression of Dasypyrum villosum Parental DNA in Wheat Lines Derived from Intergeneric Hybridization. Cytogenet. Genome Res. 2012, 136, 75–81. [Google Scholar] [CrossRef] [PubMed]
- Nemeth, C.; Yang, C.-Y.; Kasprzak, P.; Hubbart, S.; Scholefield, D.; Mehra, S.; Skipper, E.; King, I.; King, J. Generation of amphidiploids from hybrids of wheat and related species from the genera Aegilops, Secale, Thinopyrum, and Triticum as a source of genetic variation for wheat improvement. Genome 2015, 58, 71–79. [Google Scholar] [CrossRef] [PubMed]
- Harlan, J.R.; de Wet, J.M.J. Toward a rational classification of cultivated plants. Taxon 1971, 20, 509–517. [Google Scholar] [CrossRef]
- Rosyara, U.; Kishii, M.; Payne, T.; Sansaloni, C.P.; Singh, R.P.; Braun, H.-J.; Dreisigacker, S. Genetic Contribution of Synthetic Hexaploid Wheat to CIMMYT’s Spring Bread Wheat Breeding Germplasm. Sci. Rep. 2019, 9, 12355. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kishii, M.; Dou, Q.; Garg, M.; Ito, M.; Tanaka, H.; Tsujimoto, H. Production of wheat-Psathyrostachy huashanica chronomose addition lines. Genes Genet. 2010, 85, 281–286. [Google Scholar] [CrossRef] [Green Version]
- Bai, S.-S.; Zhang, H.-B.; Han, J.; Wu, J.-H.; Li, J.-C.; Geng, X.-X.; Lü, B.-Y.; Xie, S.-F.; Han, D.-J.; Zhao, J.-X.; et al. Identification of genetic locus with resistance to take-all in the wheat-Psathyrostachys huashanica Keng introgression line H148. J. Integr. Agric. 2021, 20, 3101–3113. [Google Scholar] [CrossRef]
- E McMillan, V.; Gutteridge, R.J.; E Hammond-Kosack, K. Identifying variation in resistance to the take-all fungus, Gaeumannomyces graminis var. tritici, between different ancestral and modern wheat species. BMC Plant Biol. 2014, 14, 12. [Google Scholar] [CrossRef] [Green Version]
- Du, W.; Wang, J.; Wang, L.; Zhang, J.; Chen, X.; Zhao, J.; Yang, Q.; Wu, J. Development and Characterization of a Psathyrostachys huashanica Keng 7Ns Chromosome Addition Line with Leaf Rust Resistance. PLoS ONE 2013, 8, e70879. [Google Scholar] [CrossRef] [Green Version]
- Du, W.; Wang, J.; Lu, M.; Sun, S.; Chen, X.; Zhao, J.; Yang, Q.; Wu, J. Characterization of a wheat-Psathyrostachys huashanica Keng 4Ns disomic addition line for enhanced tiller numbers and stripe rust resistance. Planta 2013, 239, 97–105. [Google Scholar] [CrossRef]
- Li, C.; Lin, H.; Chen, A.; Lau, M.; Jernstedt, J.; Dubcovsky, J. Wheat VRN1, FUL2 and FUL3 play critical and redundant roles in spikelet development and spike determinacy. Development 2019, 146, dev175398. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gol, L.; Tomé, F.; von Korff, M. Floral transitions in wheat and barley interactions between photoperiod, abiotic stresses, and nutrient status. J. Exp. Bot. 2017, 68, 1399–1410. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Whingwiri, E.E.; Stern, W.R. Floret survival in wheat: Significance of the time of floret initiation relative to terminal spikelet formation. J. Agric. Sci. 1982, 98, 257–268. [Google Scholar] [CrossRef]
- Okamoto, Y.; Takumi, S. Pleiotropic effects of the elongated glume gene P1 on grain and spikelet shape-related traits in tetraploid wheat. Euphytica 2013, 194, 207–218. [Google Scholar] [CrossRef]
- Davies, B.; Egea-Cortines, M.; de Andrade Silva, E.; Saedler, H.; Sommer, H. Multiple interactions amongst floral homeotic MADS box proteins. EMBO J. 1996, 15, 4330–4343. [Google Scholar] [CrossRef]
- Engledow, F.L. The inheritance of glume-length and grain-length in a wheat cross. J. Genet. 1920, 10, 109–134. [Google Scholar] [CrossRef] [Green Version]
- Biffen, R.H. Mendel’s Laws of Inheritance and Wheat Breeding. J. Agric. Sci. 1905, 1, 4–48. [Google Scholar] [CrossRef] [Green Version]
- Akond, M.A.S.M.G.; Watanabe, N.; Furuta, Y. Exploration of genetic diversity among Xinjiang Triticum and Triticum polonicum by AFLP markers. J. Appl. Genet. 2007, 48, 25–33. [Google Scholar] [CrossRef]
- Watanabe, N.; Yotani, Y.; Furuta, Y. The inheritance and chromosomal location of a gene for long glume in durum wheat. Euphytica 1996, 91, 235–239. [Google Scholar] [CrossRef]
- Kane, N.A.; Danyluk, J.; Tardif, G.; Ouellet, F.; Laliberté, J.-F.; Limin, A.E.; Fowler, D.B.; Sarhan, F. TaVRT-2, a Member of the StMADS-11 Clade of Flowering Repressors, Is Regulated by Vernalization and Photoperiod in Wheat. Plant Physiol. 2005, 138, 2354–2363. [Google Scholar] [CrossRef] [Green Version]
- Adamski, N.M.; Simmonds, J.; Brinton, J.F.; Backhaus, A.E.; Chen, Y.; Smedley, M.; Hayta, S.; Florio, T.; Crane, P.; Scott, P.; et al. Ectopic expression of Triticum polonicum VRT-A2 underlies elongated glumes and grains in hexaploid wheat in a dosage-dependent manner. Plant Cell 2021, 33, 2296–2319. [Google Scholar] [CrossRef] [PubMed]
- Xiao, J.; Chen, Y.; Lu, Y.; Liu, Z.; Si, D.; Xu, T.; Sun, L.; Wang, Z.; Yuan, C.; Sun, H.; et al. A natural variation of an SVP MADS-box transcription factor in Triticum petropavlovskyi leads to its ectopic expression and contributes to elongated glume. Mol. Plant 2021, 14, 1408–1411. [Google Scholar] [CrossRef] [PubMed]
- Liu, J.; Chen, Z.; Wang, Z.; Zhang, Z.; Xie, X.; Wang, Z.; Chai, L.; Song, L.; Cheng, X.; Feng, M.; et al. Ectopic expression of VRT-A2 underlies the origin of Triticum polonicum and Triticum petropavlovskyi with long outer glumes and grains. Mol. Plant 2021, 14, 1472–1488. [Google Scholar] [CrossRef] [PubMed]
- Schilling, S.; Kennedy, A.; Pan, S.; Jermiin, L.S.; Melzer, R. Genome-wide analysis of MIKC -type MADS -box genes in wheat: Pervasive duplications, functional conservation and putative neofunctionalization. New Phytol. 2020, 225, 511–529. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, J.-C.; Li, J.-J.; Zhao, L.; Zhao, J.-X.; Wu, J.; Chen, X.-H.; Zhang, L.-Y.; Dong, P.-H.; Wang, L.-M.; Zhao, D.-H.; et al. Rapid identification of Psathyrostachys huashanica Keng chromosomes in wheat background based on ND-FISH and SNP array methods. J. Integr. Agric. 2023. [Google Scholar] [CrossRef]
- Tang, Z.; Yang, Z.; Fu, S. Oligonucleotides replacing the roles of repetitive sequences pAs1, pSc119.2, pTa-535, pTa71, CCS1, and pAWRC.1 for FISH analysis. J. Appl. Genet. 2014, 55, 313–318. [Google Scholar] [CrossRef]
- Elleuch, H.; Belbahri, L.; Boetti, H.; David, H.; Thomassetb, B.; David, A. Rice salT promoter is activated in Papaver somniferum and Nicotiana tabacum transgenic cells in the absence of exogenous ABA. Enzym. Microb. Technol. 2001, 28, 106–113. [Google Scholar] [CrossRef]
- Hu, T.; Tian, Y.; Zhu, J.; Wang, Y.; Jing, R.; Lei, J.; Sun, Y.; Yu, Y.; Li, J.; Chen, X.; et al. OsNDUFA9 encoding a mitochondrial complex I subunit is essential for embryo development and starch synthesis in rice. Plant Cell Rep. 2018, 37, 1667–1679. [Google Scholar] [CrossRef]
- Mondol, P.C.; Xu, D.; Duan, L.; Shi, J.; Wang, C.; Chen, X.; Chen, M.; Hu, J.; Liang, W.; Zhang, D. Defective Pollen Wall 3 (DPW3), a novel alpha integrin-like protein, is required for pollen wall formation in rice. New Phytol. 2019, 225, 807–822. [Google Scholar] [CrossRef]
- Liu, Y.; Huang, S.; Han, J.; Hou, C.; Zheng, D.; Zhang, Z.; Wu, J. Development and Molecular Cytogenetic Identification of a New Wheat–Psathyrostachys huashanica Keng Translocation Line Resistant to Powdery Mildew. Front. Plant Sci. 2021, 12, 689502. [Google Scholar] [CrossRef]
- Tan, B.; Zhao, L.; Li, L.; Zhang, H.; Zhu, W.; Xu, L.; Wang, Y.; Zeng, J.; Fan, X.; Sha, L.; et al. Identification of a Wheat-Psathyrostachys huashanica 7Ns Ditelosomic Addition Line Conferring Early Maturation by Cytological Analysis and Newly Developed Molecular and FISH Markers. Front. Plant Sci. 2021, 12, 784001. [Google Scholar] [CrossRef]
- Li, J.; Li, J.; Jiang, S.; Zhao, L.; Xiang, L.; Fu, Y.; Liu, S.; Yang, Q.; Wu, J.; Chen, X. Addition of Psathyrostachys huashanica HMW glutenin subunit expresses positive contribution to protein polymerization and gluten microstructure of receptor wheat. Food Chem. 2023, 405 (Pt A), 134739. [Google Scholar] [CrossRef]
- Zhao, J.; Liu, Y.; Cheng, X.; Pang, Y.; Li, J.; Su, Z.; Wu, J.; Yang, Q.; Bai, G.; Chen, X. Development and identification of a dwarf wheat-Leymus mollis double substitution line with resistance to yellow rust and Fusarium head blight. Crops J. 2019, 7, 516–526. [Google Scholar] [CrossRef]
- Kane, N.A.; Agharbaoui, Z.; Diallo, A.O.; Adam, H.; Tominaga, Y.; Ouellet, F.; Sarhan, F. TaVRT2 represses transcription of the wheat vernalization gene TaVRN1. Plant J. 2007, 51, 670–680. [Google Scholar] [CrossRef]
- Millet, E. Relationships Between Grain Weight and the Size of Floret Cavity in the Wheat Spike. Ann. Bot. 1986, 58, 417–423. [Google Scholar] [CrossRef]
- Rozhkov, R.V. Inheritance of glume length and grain length in hybridisation of Triticum polonicum with hard wheat. TSitologiia i Genet. 2006, 40, 21–26. [Google Scholar]
- Metzlaff, M.; Troebner, W.; Baldauf, F.; Schlegel, R.; Cullum, J. Wheat specific repetitive DNA sequences- construction and characterization of four different genomic clones. Theor. Appl. Genet. 1986, 72, 207–210. [Google Scholar] [CrossRef]
- Afzal, N.; Wang, R.-N.; Dong, P.-B.; Wang, N.; Liu, H.-Y.; Li, Z.-H. The whole chloroplast genome of Psathyrostachys huashanica (Gramineae), a rare and endangered herb species endemic to China. Mitochondrial DNA Part B 2019, 4, 622–623. [Google Scholar] [CrossRef] [Green Version]
- Elshafei, A.A.; Motawei, M.I.; Esmail, R.M.; Al-Doss, A.A.; Hussien, A.M.; Ibrahim, E.I.; Amer, M.A. Molecular breeding for rust resistance in wheat genotypes. Mol. Biol. Rep. 2021, 48, 731–742. [Google Scholar] [CrossRef]
- Su, J.N.; Guo, J.; Wang, C.J.; Jin, F.; Zhao, J.X.; Yang, Q.H.; Chen, X.H.; Wu, J. Specific SCAR Markers on Chromosome 3Ns of Psathyrostachys huashanica Keng. J. Triticeae Crops 2015, 35, 1–6. [Google Scholar]
- Gao, Z.; Yun, L.; Li, Z.; Liu, Q.; Zhang, C.; Ma, Y.; Shi, F. Hybrid purity identification using EST-SSR markers and heterosis analysis of quantitative traits of Russian wildrye. PeerJ 2022, 10, e14442. [Google Scholar] [CrossRef]
- Said, M.; Hřibová, E.; Danilova, T.V.; Karafiátová, M.; Čížková, J.; Friebe, B.; Doležel, J.; Gill, B.S.; Vrána, J. The Agropyron cristatum karyotype, chromosome structure and cross-genome homoeology as revealed by fluorescence in situ hybridization with tandem repeats and wheat single-gene probes. Theor. Appl. Genet. 2018, 131, 2213–2227. [Google Scholar] [CrossRef] [Green Version]
- Pertea, M.; Kim, D.; Pertea, G.M.; Leek, J.T.; Salzberg, S.L. Transcript-level expression analysis of RNA-seq experiments with HISAT, StringTie and Ballgown. Nat. Protoc. 2016, 11, 1650–1667. [Google Scholar] [CrossRef]
- Burks, D.J.; Azad, R.K. RNA-Seq Data Analysis Pipeline for Plants: Transcriptome Assembly, Alignment, and Differential Expression Analysis. Methods Mol. Biol. 2022, 2396, 47–60. [Google Scholar] [CrossRef]
- Nakayama, R.; Safi, M.T.; Ahmadzai, W.; Sato, K.; Kawaura, K. Comparative transcriptome analysis of synthetic and common wheat in response to salt stress. Sci. Rep. 2022, 12, 11534. [Google Scholar] [CrossRef]
- Buttar, Z.A.; Shalmani, A.; Niaz, M.; Wang, C.; Hussain, S.; Wang, C. Transcriptome Analysis Reveals Potential Mechanism in Storage Protein Trafficking within Developing Grains of Common Wheat. Int. J. Mol. Sci. 2022, 23, 14851. [Google Scholar] [CrossRef]
- Camacho, J.P.; Sharbel, T.F.; Beukeboom, L.W. B-chromosome evolution. Philos. Trans. R. Soc. London. Ser. B Biol. Sci. 2000, 355, 163–178. [Google Scholar] [CrossRef] [Green Version]
- Hyde, L.; Osman, K.; Winfield, M.; Sanchez-Moran, E.; Higgins, J.D.; Henderson, I.R.; Sparks, C.; Franklin, F.C.H.; Edwards, K.J. Identification, characterization, and rescue of CRISPR /Cas9 generated wheat SPO11-1 mutants. Plant Biotechnol. J. 2023, 21, 405–418. [Google Scholar] [CrossRef]
- Türkösi, E.; Ivanizs, L.; Farkas, A.; Gaál, E.; Kruppa, K.; Kovács, P.; Szakács, É; Szőke-Pázsi, K.; Said, M.; Cápal, P.; et al. Transfer of the ph1b Deletion Chromosome 5B From Chinese Spring Wheat Into a Winter Wheat Line and Induction of Chromosome Rearrangements in Wheat-Aegilops biuncialis Hybrids. Front. Plant Sci. 2022, 13, 875676. [Google Scholar] [CrossRef]
- Li, J.; Zhao, L.; Cheng, X.; Bai, G.; Li, M.; Wu, J.; Yang, Q.; Chen, X.; Yang, Z.; Zhao, J. Molecular cytogenetic characterization of a novel wheat–Psathyrostachys huashanica Keng T3DS-5NsL*5NsS and T5DL-3DS*3DL dual translocation line with powdery mildew resistance. BMC Plant Biol. 2020, 20, 163. [Google Scholar] [CrossRef] [Green Version]
- Wang, Y.; Cao, Q.; Zhang, J.; Wang, S.; Chen, C.; Wang, C.; Zhang, H.; Wang, Y.; Ji, W. Cytogenetic Analysis and Molecular Marker Development for a New Wheat–Thinopyrum ponticum 1Js (1D) Disomic Substitution Line With Resistance to Stripe Rust and Powdery Mildew. Front. Plant Sci. 2020, 11, 1282. [Google Scholar] [CrossRef]
- Du, X.; Feng, X.; Li, R.; Jin, Y.; Shang, L.; Zhao, J.; Wang, C.; Li, T.; Chen, C.; Tian, Z.; et al. Cytogenetic identification and molecular marker development of a novel wheat-Leymus mollis 4Ns(4D) alien disomic substitution line with resistance to stripe rust and Fusarium head blight. Front. Plant Sci. 2022, 13, 1012939. [Google Scholar] [CrossRef] [PubMed]
- Ngernmuen, A.; Suktrakul, W.; Damchuay, K.; Longya, A.; Kate-Ngam, S.; Jantasuriyarat, C. Substantial enhancement of high polymorphic SSR marker development using in silico method from 18 available rice blast fungus genome sequences and its application in genetic diversity assessment. Biologia 2019, 74, 1181–1189. [Google Scholar] [CrossRef]
- Ando, K.; Krishnan, V.; Rynearson, S.; Rouse, M.N.; Danilova, T.; Friebe, B.; See, D.; Pumphrey, M.O. Introgression of a novel Ug99-effective stem rust resistance gene into wheat and devolopment of Dasypyrum villosum chromosome specific markers via genotyping-by-sequencing(GBS). Plant Dis. 2019, 103, 33. [Google Scholar] [CrossRef]
- Katsu, K.; Suzuki, T.; Fujino, K.; Morishita, T.; Noda, T. Development of a DNA marker for variety discrimination specific to ‘Manten-Kirari’ based on an NGS-RNA sequence in Tartary buckwheat (Fagopyrum tataricum). Food Chem. 2019, 295, 51–57. [Google Scholar] [CrossRef]
- Kim, S.H.; Kim, D.Y.; Yacoubi, I.; Seo, Y.W. Development of single-nucleotide polymorphism markers of salinity tolerance for Tunisian durum wheat using RNA sequencing. Acta Agric. Scand. Sect. B—Soil Plant Sci. 2020, 71, 28–44. [Google Scholar] [CrossRef]
- Ghimire, B.; Sapkota, S.; Bahri, B.A.; Martinez-Espinoza, A.D.; Buck, J.W.; Mergoum, M. Fusarium Head Blight and Rust Diseases in Soft Red Winter Wheat in the Southeast United States: State of the Art, Challenges and Future Perspective for Breeding. Front. Plant Sci. 2020, 11, 1080. [Google Scholar] [CrossRef]
- Large, E.C. Growth stages in cereals illustration of the feekes scale. Plant Pathol. 1954, 3, 128–129. [Google Scholar] [CrossRef]
- Shahjaman; Mollah, M.H.; Rahman, R.; Islam, S.S.; Mollah, N.H. Robust identification of differentially expressed genes from RNA-seq data. Genomics 2020, 112, 2000–2010. [Google Scholar] [CrossRef]
- Heslop-Harrison, J. Relationships between species of Leymus, Psathyrostachys, and Hordeum (Poaceae, Triticeae) inferred from Southern hybridization of genomic and cloned DNA probes. Plant Syst. Evol. 1994, 189, 217–231. [Google Scholar] [CrossRef]
- Giorgi, D.; D’Ovidio, R.; A Tanzarella, O.; Ceoloni, C.; Porceddu, E. Isolation and characterization of S genome specific sequences from Aegilops sect. sitopsis species. Genome 2003, 46, 478–489. [Google Scholar] [CrossRef] [PubMed]
- Bödvarsdóttir, S.K.; Anamthawat-Jónsson, K. Isolation, characterization, and analysis of Leymus-specific DNA sequences. Genome 2003, 46, 673–682. [Google Scholar] [CrossRef] [PubMed]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Material | Plant Height (cm) | Spike Length (mm) | Glume length (mm) | Tiller Number | Spikelet Per Spike | TGW (g) | Grain Length (mm) | Grain Width (mm) | Awn Type |
|---|---|---|---|---|---|---|---|---|---|
| 7182 | 81.8 ± 4.2 a | 11.1 ± 1.1 ab | 10.7 ± 0.9 b | 15 ± 3 a | 21 ± 1 a | 36.63 ± 0.73 c | 6.08 ± 0.03 c | 3.19 ± 0.03 b | long |
| Trs-372 | 81.7 ± 2.5 a | 10.5 ± 0.5 b | 11.3 ± 0.5 b | 12 ± 3 b | 19 ± 1 b | 53.13 ± 0.31 a | 7.97 ± 0.02 a | 3.31 ± 0.01 ab | long |
| 20JH1155 | 70.5 ± 2.6 b | 11.5 ± 4.9 a | 15.7 ± 0.8 a | 8 ± 2 c | 19 ± 1 b | 47.26 ± 0.38 b | 7.25 ± 0.03 b | 3.36 ± 0.02 a | zero |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Pang, J.; Huang, C.; Wang, Y.; Wen, X.; Deng, P.; Li, T.; Wang, C.; Liu, X.; Chen, C.; Zhao, J.; et al. Molecular Cytological Analysis and Specific Marker Development in Wheat-Psathyrostachys huashanica Keng 3Ns Additional Line with Elongated Glume. Int. J. Mol. Sci. 2023, 24, 6726. https://doi.org/10.3390/ijms24076726
Pang J, Huang C, Wang Y, Wen X, Deng P, Li T, Wang C, Liu X, Chen C, Zhao J, et al. Molecular Cytological Analysis and Specific Marker Development in Wheat-Psathyrostachys huashanica Keng 3Ns Additional Line with Elongated Glume. International Journal of Molecular Sciences. 2023; 24(7):6726. https://doi.org/10.3390/ijms24076726
Chicago/Turabian StylePang, Jingyu, Chenxi Huang, Yuesheng Wang, Xinyu Wen, Pingchuan Deng, Tingdong Li, Changyou Wang, Xinlun Liu, Chunhuan Chen, Jixin Zhao, and et al. 2023. "Molecular Cytological Analysis and Specific Marker Development in Wheat-Psathyrostachys huashanica Keng 3Ns Additional Line with Elongated Glume" International Journal of Molecular Sciences 24, no. 7: 6726. https://doi.org/10.3390/ijms24076726