
Nitrite Reductase Activity of Ferrous Nitrobindins: A Comparative Study
 ,
,  ,
, 
Abstract
:1. Introduction
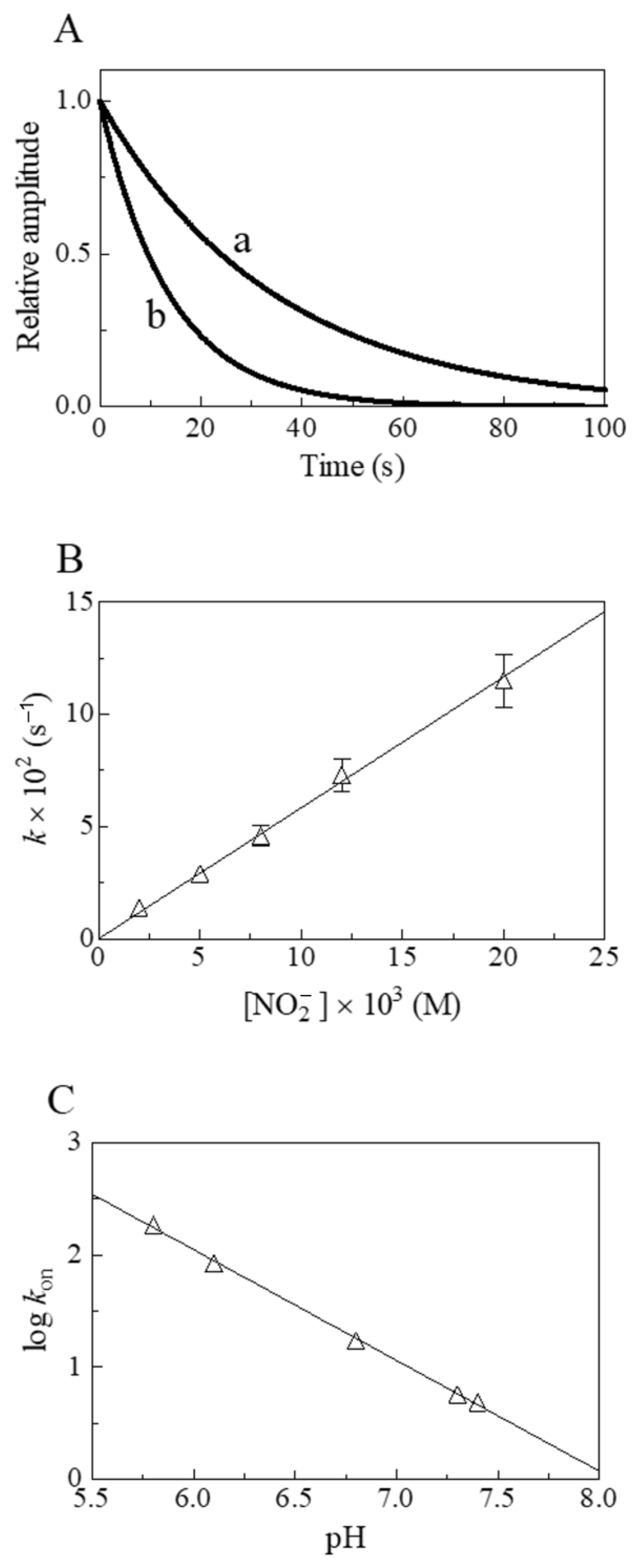
2. Results
3. Discussion
4. Materials and Methods
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Antonini, E.; Brunori, M. Hemoglobin and Myoglobin in Their Reactions with Ligands; North Holland Publishing Co.: Amsterdam, The Netherlands, 1971. [Google Scholar]
- Perutz, M.F. Regulation of oxygen affinity of hemoglobin: Influence of structure of the globin on the heme iron. Annu. Rev. Biochem. 1979, 48, 327–386. [Google Scholar] [CrossRef] [PubMed]
- Chan, M.K. Recent advances in heme-protein sensors. Curr. Opin. Chem. Biol. 2001, 5, 216–222. [Google Scholar] [CrossRef] [PubMed]
- Ascenzi, P.; di Masi, A.; Leboffe, L.; Fiocchetti, M.; Nuzzo, M.T.; Brunori, M.; Marino, M. Neuroglobin: From structure to function in health and disease. Mol. Aspects Med. 2016, 52, 1–48. [Google Scholar] [CrossRef]
- Domingues-Hamdi, E.; Vasseur, C.; Fournier, J.B.; Marden, M.C.; Wajcman, H.; Baudin-Creuza, V. Role of alpha-globin H helix in the building of tetrameric human hemoglobin: Interaction with alpha-hemoglobin stabilizing protein (AHSP) and heme molecule. PLoS ONE 2014, 9, e111395. [Google Scholar] [CrossRef] [PubMed]
- Vasseur, C.; Baudin-Creuza, V. Role of alpha-hemoglobin molecular chaperone in the hemoglobin formation and clinical expression of some hemoglobinopathies. Transfus. Clin. Biol. 2015, 22, 49–57. [Google Scholar] [CrossRef]
- Nardini, M.; Pesce, A.; Bolognesi, M. Truncated (2/2) hemoglobin: Unconventional structures and functional roles in vivo and in human pathogenesis. Mol. Aspects Med. 2022, 84, 101049. [Google Scholar] [CrossRef]
- Couture, M.; Yeh, S.R.; Wittenberg, B.A.; Wittenberg, J.B.; Ouellet, Y.; Rousseau, D.L.; Guertin, M. A cooperative oxygen-binding hemoglobin from Mycobacterium tuberculosis. Proc. Natl. Acad. Sci. USA 1999, 96, 11223–11228. [Google Scholar] [CrossRef] [Green Version]
- Wittenberg, J.B.; Bolognesi, M.; Wittenberg, B.A.; Guertin, M. Truncated hemoglobins: A new family of hemoglobins widely distributed in bacteria, unicellular eukaryotes, and plants. J. Biol. Chem. 2002, 277, 871–874. [Google Scholar] [CrossRef] [Green Version]
- Vuletich, D.A.; Lecomte, J.T. A phylogenetic and structural analysis of truncated hemoglobins. J. Mol. Evol. 2006, 62, 196–210. [Google Scholar] [CrossRef]
- Nardini, M.; Pesce, A.; Milani, M.; Bolognesi, M. Protein fold and structure in the truncated (2/2) globin family. Gene 2007, 398, 2–11. [Google Scholar] [CrossRef]
- Vinogradov, S.N.; Tinajero-Trejo, M.; Poole, R.K.; Hoogewijs, D. Bacterial and archaeal globins—A revised perspective. Biochim. Biophys. Acta 2013, 1834, 1789–1800. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- De Simone, G.; Ascenzi, P.; di Masi, A.; Polticelli, F. Nitrophorins and nitrobindins: Structure and function. Biomol. Concepts 2017, 8, 105–118. [Google Scholar] [CrossRef] [PubMed]
- Montfort, W.R.; Weichsel, A.; Andersen, J.F. Nitrophorins and related antihemostatic lipocalins from Rhodnius prolixus and other blood-sucking arthropods. Biochim. Biophys. Acta 2000, 1482, 110–118. [Google Scholar] [CrossRef] [PubMed]
- Andersen, J.F. Structure and mechanism in salivary proteins from blood-feeding arthropods. Toxicon 2010, 56, 1120–1129. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bianchetti, C.M.; Blouin, G.C.; Bitto, E.; Olson, J.S.; Phillips, G.N., Jr. The structure and NO binding properties of the nitrophorin-like heme-binding protein from Arabidopsis thaliana gene locus At1g79260.1. Proteins 2010, 78, 917–931. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bianchetti, C.M.; Bingman, C.A.; Phillips, G.N., Jr. Structure of the C-terminal heme-binding domain of THAP domain containing protein 4 from Homo sapiens. Proteins 2011, 79, 1337–1341. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- De Simone, G.; Ascenzi, P.; Polticelli, F. Nitrobindin: An Ubiquitous Family of All β-Barrel Heme-proteins. IUBMB Life 2016, 68, 423–428. [Google Scholar] [CrossRef] [Green Version]
- De Simone, G.; di Masi, A.; Vita, G.M.; Polticelli, F.; Pesce, A.; Nardini, M.; Bolognesi, M.; Ciaccio, C.; Coletta, M.; Turilli, E.S.; et al. Mycobacterial and Human Nitrobindins: Structure and Function. Antioxid. Redox Signal. 2020, 33, 229–246. [Google Scholar] [CrossRef]
- De Simone, G.; di Masi, A.; Ciaccio, C.; Coletta, M.; Ascenzi, P. NO Scavenging through Reductive Nitrosylation of Ferric Mycobacterium tuberculosis and Homo sapiens Nitrobindins. Int. J. Mol. Sci. 2020, 21, 9395. [Google Scholar] [CrossRef]
- De Simone, G.; Fattibene, P.; Sebastiani, F.; Smulevich, G.; Coletta, M.; Ascenzi, P. Dissociation of the proximal His-Fe bond upon NO binding to ferrous zebrafish nitrobindin. J. Inorg. Biochem. 2022, 236, 111962. [Google Scholar] [CrossRef]
- De Simone, G.; Sebastiani, F.; Smulevich, G.; Coletta, M.; Ascenzi, P. Nitrosylation of ferric zebrafish nitrobindin: A spectroscopic, kinetic, and thermodynamic study. J. Inorg. Biochem. 2022, 237, 111996. [Google Scholar] [CrossRef] [PubMed]
- De Simone, G.; di Masi, A.; Pesce, A.; Bolognesi, M.; Ciaccio, C.; Tognaccini, L.; Smulevich, G.; Abbruzzetti, S.; Viappiani, C.; Bruno, S.; et al. Mycobacterial and Human Ferrous Nitrobindins: Spectroscopic and Reactivity Properties. Int. J. Mol Sci. 2021, 22, 1674. [Google Scholar] [CrossRef] [PubMed]
- De Simone, G.; di Masi, A.; Fattibene, P.; Ciaccio, C.; Platas-Iglesias, C.; Coletta, M.; Pesce, A.; Ascenzi, P. Oxygen-mediated oxidation of ferrous nitrosylated nitrobindins. J. Inorg. Biochem. 2021, 224, 111579. [Google Scholar] [CrossRef]
- De Simone, G.; di Masi, A.; Polticelli, F.; Ascenzi, P. Human nitrobindin: The first example of an all-β-barrel ferric heme-protein that catalyzes peroxynitrite detoxification. FEBS Open Bio 2018, 8, 2002–2010. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- De Simone, G.; Coletta, A.; di Masi, A.; Coletta, M.; Ascenzi, P. The Balancing of Peroxynitrite Detoxification between Ferric Heme-Proteins and CO2: The Case of Zebrafish Nitrobindin. Antioxidants 2022, 11, 1932. [Google Scholar] [CrossRef] [PubMed]
- Ascenzi, P.; Tundo, G.R.; Fanali, G.; Coletta, M.; Fasano, M. Warfarin modulates the nitrite reductase activity of ferrous human serum heme-albumin. J. Biol. Inorg. Chem. 2013, 18, 939–946. [Google Scholar] [CrossRef]
- Ascenzi, P.; Sbardella, D.; Fiocchetti, M.; Santucci, R.; Coletta, M. NO2−-mediated nitrosylation of ferrous microperoxidase-11. J. Inorg. Biochem. 2015, 153, 121–127. [Google Scholar] [CrossRef]
- Ascenzi, P.; Sbardella, D.; Sinibaldi, F.; Santucci, R.; Coletta, M. The nitrite reductase activity of horse heart carboxymethylated-cytochrome c is modulated by cardiolipin. J. Biol. Inorg. Chem. 2016, 21, 421–432. [Google Scholar] [CrossRef]
- Ascenzi, P.; Tundo, G.R.; Coletta, M. The nitrite reductase activity of ferrous human hemoglobin:haptoglobin 1-1 and 2-2 complexes. J. Inorg. Biochem. 2018, 187, 116–122. [Google Scholar] [CrossRef]
- Toma, C.; De Cilla, S.; Palumbo, A.; Garhwal, D.P.; Grossini, E. Oxidative and Nitrosative Stress in Age-Related Macular Degeneration: A Review of Their Role in Different Stages of Disease. Antioxidants 2021, 10, 653. [Google Scholar] [CrossRef]
- Doyle, M.P.; Pickering, R.A.; DeWeert, T.M.; Hoekstra, J.W.; Pater, D. Kinetics and mechanism of the oxidation of human deoxyhemoglobin by nitrites. J. Biol. Chem. 1981, 256, 12393–12398. [Google Scholar] [CrossRef] [PubMed]
- Huang, K.T.; Keszler, A.; Patel, N.; Patel, R.P.; Gladwin, M.T.; Kim-Shapiro, D.B.; Hogg, N. The reaction between nitrite and deoxyhemoglobin. Reassessment of reaction kinetics and stoichiometry. J. Biol. Chem. 2005, 280, 31126–31131. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Huang, Z.; Shiva, S.; Kim-Shapiro, D.B.; Patel, R.P.; Ringwood, L.A.; Irby, C.E.; Huang, K.T.; Ho, C.; Hogg, N.; Schechter, A.N.; et al. Enzymatic function of hemoglobin as a nitrite reductase that produces NO under allosteric control. J. Clin. Investig. 2005, 115, 2099–2107. [Google Scholar] [CrossRef] [Green Version]
- Shiva, S.; Huang, Z.; Grubina, R.; Sun, J.; Ringwood, L.A.; MacArthur, P.H.; Xu, X.; Murphy, E.; Darley-Usmar, V.M.; Gladwin, M.T. Deoxymyoglobin is a nitrite reductase that generates nitric oxide and regulates mitochondrial respiration. Circ. Res. 2007, 100, 654–661. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Basu, S.; Azarova, N.A.; Font, M.D.; King, S.B.; Hogg, N.; Gladwin, M.T.; Shiva, S.; Kim-Shapiro, D.B. Nitrite reductase activity of cytochrome c. J. Biol. Chem. 2008, 283, 32590–32597. [Google Scholar] [CrossRef] [Green Version]
- Grubina, R.; Basu, S.; Tiso, M.; Kim-Shapiro, D.B.; Gladwin, M.T. Nitrite reductase activity of hemoglobin S (sickle) provides insight into contributions of heme redox potential versus ligand affinity. J. Biol. Chem. 2008, 283, 3628–3638. [Google Scholar] [CrossRef] [Green Version]
- Petersen, M.G.; Dewilde, S.; Fago, A. Reactions of ferrous neuroglobin and cytoglobin with nitrite under anaerobic conditions. J. Inorg. Biochem. 2008, 102, 1777–1782. [Google Scholar] [CrossRef]
- Salhany, J.M. Kinetics of reaction of nitrite with deoxy hemoglobin after rapid deoxygenation or predeoxygenation by dithionite measured in solution and bound to the cytoplasmic domain of band 3 (SLC4A1). Biochemistry 2008, 47, 6059–6072. [Google Scholar] [CrossRef]
- Sturms, R.; DiSpirito, A.A.; Hargrove, M.S. Plant and cyanobacterial hemoglobins reduce nitrite to nitric oxide under anoxic conditions. Biochemistry 2011, 50, 3873–3878. [Google Scholar] [CrossRef]
- Tiso, M.; Tejero, J.; Kenney, C.; Frizzell, S.; Gladwin, M.T. Nitrite reductase activity of nonsymbiotic hemoglobins from Arabidopsis thaliana. Biochemistry 2012, 51, 5285–5292. [Google Scholar] [CrossRef] [Green Version]
- Helbo, S.; Dewilde, S.; Williams, D.R.; Berghmans, H.; Berenbrink, M.; Cossins, A.R.; Fago, A. Functional differentiation of myoglobin isoforms in hypoxia-tolerant carp indicates tissue-specific protective roles. Am. J. Physiol. Regul. Integr. Comp. Physiol. 2012, 302, R693–R701. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, H.; Hemann, C.; Abdelghany, T.M.; El-Mahdy, M.A.; Zweier, J.L. Characterization of the mechanism and magnitude of cytoglobin-mediated nitrite reduction and nitric oxide generation under anaerobic conditions. J. Biol. Chem. 2012, 287, 36623–36633. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tiso, M.; Tejero, J.; Basu, S.; Azarov, I.; Wang, X.; Simplaceanu, V.; Frizzell, S.; Jayaraman, T.; Geary, L.; Shapiro, C.; et al. Human neuroglobin functions as a redox-regulated nitrite reductase. J. Biol. Chem. 2011, 286, 18277–18289. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ascenzi, P.; Marino, M.; Polticelli, F.; Santucci, R.; Coletta, M. Cardiolipin modulates allosterically the nitrite reductase activity of horse heart cytochrome c. J. Biol. Inorg. Chem. 2014, 19, 1195–1201. [Google Scholar] [CrossRef] [PubMed]
- Ascenzi, P.; Leboffe, L.; Pesce, A.; Ciaccio, C.; Sbardella, D.; Bolognesi, M.; Coletta, M. Nitrite-reductase and peroxynitrite isomerization activities of Methanosarcina acetivorans protoglobin. PLoS ONE 2014, 9, e95391. [Google Scholar] [CrossRef]
- Ascenzi, P.; di Masi, A.; Tundo, G.R.; Pesce, A.; Visca, P.; Coletta, M. Nitrosylation mechanisms of Mycobacterium tuberculosis and Campylobacter jejuni truncated hemoglobins N, O, and P. PLoS ONE 2014, 9, e102811. [Google Scholar] [CrossRef] [Green Version]
- Ciaccio, C.; Ocana-Calahorro, F.; Droghetti, E.; Tundo, G.R.; Sanz-Luque, E.; Polticelli, F.; Visca, P.; Smulevich, G.; Ascenzi, P.; Coletta, M. Functional and Spectroscopic Characterization of Chlamydomonas reinhardtii Truncated Hemoglobins. PLoS ONE 2015, 10, e0125005. [Google Scholar] [CrossRef] [Green Version]
- Smagghe, B.J.; Sarath, G.; Ross, E.; Hilbert, J.L.; Hargrove, M.S. Slow ligand binding kinetics dominate ferrous hexacoordinate hemoglobin reactivities and reveal differences between plants and other species. Biochemistry 2006, 45, 561–570. [Google Scholar] [CrossRef] [Green Version]
- Beckerson, P.; Reeder, B.J.; Wilson, M.T. Coupling of disulfide bond and distal histidine dissociation in human ferrous cytoglobin regulates ligand binding. FEBS Lett. 2015, 589, 507–512. [Google Scholar] [CrossRef] [Green Version]
- Couture, M.; Das, T.K.; Savard, P.Y.; Ouellet, Y.; Wittenberg, J.B.; Wittenberg, B.A.; Rousseau, D.L.; Guertin, M. Structural investigations of the hemoglobin of the cyanobacterium Synechocystis PCC6803 reveal a unique distal heme pocket. Eur. J. Biochem. 2000, 267, 4770–4780. [Google Scholar] [CrossRef]
- Bruno, S.; Faggiano, S.; Spyrakis, F.; Mozzarelli, A.; Abbruzzetti, S.; Grandi, E.; Viappiani, C.; Feis, A.; Mackowiak, S.; Smulevich, G.; et al. The reactivity with CO of AHb1 and AHb2 from Arabidopsis thaliana is controlled by the distal HisE7 and internal hydrophobic cavities. J. Am. Chem. Soc. 2007, 129, 2880–2889. [Google Scholar] [CrossRef] [PubMed]
- Tilleman, L.; Abbruzzetti, S.; Ciaccio, C.; De Sanctis, G.; Nardini, M.; Pesce, A.; Desmet, F.; Moens, L.; Van Doorslaer, S.; Bruno, S.; et al. Structural Bases for the Regulation of CO Binding in the Archaeal Protoglobin from Methanosarcina acetivorans. PLoS ONE 2015, 10, e0125959. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Smulevich, G.; Mantini, A.R.; Paoli, M.; Coletta, M.; Geraci, G. Resonance Raman studies of the heme active site of the homodimeric myoglobin from Nassa mutabilis: A peculiar case. Biochemistry 1995, 34, 7507–7516. [Google Scholar] [CrossRef] [PubMed]
- Egeberg, K.D.; Springer, B.A.; Martinis, S.A.; Sligar, S.G.; Morikis, D.; Champion, P.M. Alteration of sperm whale myoglobin heme axial ligation by site-directed mutagenesis. Biochemistry 1990, 29, 9783–9791. [Google Scholar] [CrossRef]
- Exertier, C.; Sebastiani, F.; Freda, I.; Gugole, E.; Cerutti, G.; Parisi, G.; Montemiglio, L.C.; Becucci, M.; Viappiani, C.; Bruno, S.; et al. Probing the Role of Murine Neuroglobin CDloop-D-Helix Unit in CO Ligand Binding and Structural Dynamics. ACS Chem. Biol. 2022, 17, 2099–2108. [Google Scholar] [CrossRef]
- Couture, M.; Burmester, T.; Hankeln, T.; Rousseau, D.L. The heme environment of mouse neuroglobin. Evidence for the presence of two conformations of the heme pocket. J. Biol. Chem. 2001, 276, 36377–36382. [Google Scholar] [CrossRef] [Green Version]
- Van Doorslaer, S.; Vinck, E.; Trandafir, F.; Ioanitescu, I.; Dewilde, S.; Moens, L. Tracing the structure-function relationship of neuroglobin and cytoglobin using resonance Raman and electron paramagnetic resonance spectroscopy. IUBMB Life 2004, 56, 665–670. [Google Scholar] [CrossRef]
- Reeder, B.J.; Ukeri, J. Strong modulation of nitrite reductase activity of cytoglobin by disulfide bond oxidation: Implications for nitric oxide homeostasis. Nitric Oxide 2018, 72, 16–23. [Google Scholar] [CrossRef]
- Makino, M.; Sugimoto, H.; Sawai, H.; Kawada, N.; Yoshizato, K.; Shiro, Y. High-resolution structure of human cytoglobin: Identification of extra N- and C-termini and a new dimerization mode. Acta Crystallogr. D Biol. Crystallogr. 2006, 62, 671–677. [Google Scholar] [CrossRef] [Green Version]
- Coletta, M.; Angeletti, M.; Ascone, I.; Boumis, G.; Castellano, A.C.; Dell’Ariccia, M.; Della Longa, S.; De Sanctis, G.; Priori, A.M.; Santucci, R.; et al. Heterotropic effectors exert more significant strain on monoligated than on unligated hemoglobin. Biophys. J. 1999, 76, 1532–1536. [Google Scholar] [CrossRef] [Green Version]
- Fermi, G.; Perutz, M.F.; Shaanan, B.; Fourme, R. The crystal structure of human deoxyhaemoglobin at 1.74 A resolution. J. Mol. Biol. 1984, 175, 159–174. [Google Scholar] [CrossRef] [PubMed]
- Sawicki, C.A.; Gibson, Q.H. Quaternary conformational changes in human hemoglobin studied by laser photolysis of carboxyhemoglobin. J. Biol. Chem. 1976, 251, 1533–1542. [Google Scholar] [CrossRef]
- Nagel, R.L.; Gibson, Q.H. Kinetics of the reaction of carbon monoxide with the hemoglobin-haptoglobin complex. J. Mol. Biol. 1966, 22, 249–255. [Google Scholar] [CrossRef]
- Ascenzi, P.; Coletta, A.; Cao, Y.; Trezza, V.; Leboffe, L.; Fanali, G.; Fasano, M.; Pesce, A.; Ciaccio, C.; Marini, S.; et al. Isoniazid inhibits the heme-based reactivity of Mycobacterium tuberculosis truncated hemoglobin N. PLoS ONE 2013, 8, e69762. [Google Scholar] [CrossRef] [Green Version]
- Ouellet, Y.; Milani, M.; Couture, M.; Bolognesi, M.; Guertin, M. Ligand interactions in the distal heme pocket of Mycobacterium tuberculosis truncated hemoglobin N: Roles of TyrB10 and GlnE11 residues. Biochemistry 2006, 45, 8770–8781. [Google Scholar] [CrossRef]
- Ouellet, H.; Juszczak, L.; Dantsker, D.; Samuni, U.; Ouellet, Y.H.; Savard, P.Y.; Wittenberg, J.B.; Wittenberg, B.A.; Friedman, J.M.; Guertin, M. Reactions of Mycobacterium tuberculosis truncated hemoglobin O with ligands reveal a novel ligand-inclusive hydrogen bond network. Biochemistry 2003, 42, 5764–5774. [Google Scholar] [CrossRef] [PubMed]
- Mukai, M.; Savard, P.Y.; Ouellet, H.; Guertin, M.; Yeh, S.R. Unique ligand-protein interactions in a new truncated hemoglobin from Mycobacterium tuberculosis. Biochemistry 2002, 41, 3897–3905. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Farres, J.; Rechsteiner, M.P.; Herold, S.; Frey, A.D.; Kallio, P.T. Ligand binding properties of bacterial hemoglobins and flavohemoglobins. Biochemistry 2005, 44, 4125–4134. [Google Scholar] [CrossRef] [PubMed]
- Wainwright, L.M.; Wang, Y.; Park, S.F.; Yeh, S.R.; Poole, R.K. Purification and spectroscopic characterization of Ctb, a group III truncated hemoglobin implicated in oxygen metabolism in the food-borne pathogen Campylobacter jejuni. Biochemistry 2006, 45, 6003–6011. [Google Scholar] [CrossRef] [Green Version]
- Bocedi, A.; De Sanctis, G.; Ciaccio, C.; Tundo, G.R.; Di Masi, A.; Fanali, G.; Nicoletti, F.P.; Fasano, M.; Smulevich, G.; Ascenzi, P.; et al. Reciprocal allosteric modulation of carbon monoxide and warfarin binding to ferrous human serum heme-albumin. PLoS ONE 2013, 8, e58842. [Google Scholar] [CrossRef] [Green Version]
- Wardell, M.; Wang, Z.; Ho, J.X.; Robert, J.; Ruker, F.; Ruble, J.; Carter, D.C. The atomic structure of human methemalbumin at 1.9 A. Biochem. Biophys. Res. Commun. 2002, 291, 813–819. [Google Scholar] [CrossRef] [PubMed]
- Banci, L.; Bertini, I.; Gray, H.B.; Luchinat, C.; Reddig, T.; Rosato, A.; Turano, P. Solution structure of oxidized horse heart cytochrome c. Biochemistry 1997, 36, 9867–9877. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kapetanaki, S.M.; Silkstone, G.; Husu, I.; Liebl, U.; Wilson, M.T.; Vos, M.H. Interaction of carbon monoxide with the apoptosis-inducing cytochrome c-cardiolipin complex. Biochemistry 2009, 48, 1613–1619. [Google Scholar] [CrossRef] [PubMed]
- Wilson, M.T.; Brunori, M.; Rotilio, G.C.; Antonini, E. Properties of modified cytochromes. II. Ligand binding to reduced carboxymethyl cytochrome c. J. Biol. Chem. 1973, 248, 8162–8169. [Google Scholar] [CrossRef] [PubMed]
- Silkstone, G.; Jasaitis, A.; Vos, M.H.; Wilson, M.T. Geminate carbon monoxide rebinding to a c-type haem. Dalton Trans. 2005, 21, 3489–3494. [Google Scholar] [CrossRef] [Green Version]
- Miksovska, J.; Norstrom, J.; Larsen, R.W. Thermodynamic profiles for CO photodissociation from heme model compounds: Effect of proximal ligands. Inorg. Chem. 2005, 44, 1006–1014. [Google Scholar] [CrossRef]
- Laberge, M.; Vreugdenhil, A.J.; Vanderkooi, J.M.; Butler, I.S. Microperoxidase-11: Molecular dynamics and Q-band excited resonance Raman of the oxidized, reduced and carbonyl forms. J. Biomol. Struct. Dyn. 1998, 15, 1039–1050. [Google Scholar] [CrossRef]
- Coletta, M.; Ascenzi, P.; Brunori, M. Kinetic evidence for a role of heme geometry on the modulation of carbon monoxide reactivity in human hemoglobin. J. Biol. Chem. 1988, 263, 18286–18289. [Google Scholar] [CrossRef]
- Perutz, M.F. Myoglobin and haemoglobin: Role of distal residues in reactions with haem ligands. Trends Biochem. Sci. 1989, 14, 42–44. [Google Scholar] [CrossRef]
- Coletta, M.; Ascenzi, P.; Traylor, T.G.; Brunori, M. Kinetics of carbon monoxide binding to monomeric hemoproteins. Role of the proximal histidine. J. Biol. Chem. 1985, 260, 4151–4155. [Google Scholar] [CrossRef]
- Hoy, J.A.; Kundu, S.; Trent, J.T., 3rd; Ramaswamy, S.; Hargrove, M.S. The crystal structure of Synechocystis hemoglobin with a covalent heme linkage. J. Biol. Chem. 2004, 279, 16535–16542. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Robinson, V.L.; Smith, B.B.; Arnone, A. A pH-dependent aquomet-to-hemichrome transition in crystalline horse methemoglobin. Biochemistry 2003, 42, 10113–10125. [Google Scholar] [CrossRef] [PubMed]
- Dordas, C. Nonsymbiotic hemoglobins and stress tolerance in plants. Plant Sci. 2009, 176, 433–440. [Google Scholar] [CrossRef] [PubMed]
- Mukhi, N.; Dhindwal, S.; Uppal, S.; Kumar, P.; Kaur, J.; Kundu, S. X-ray crystallographic structural characteristics of Arabidopsis hemoglobin I and their functional implications. Biochim. Biophys. Acta 2013, 1834, 1944–1956. [Google Scholar] [CrossRef] [PubMed]
- Barbosa, R.M.; Lopes Jesus, A.J.; Santos, R.M.; Pereira, C.L.; Marques, C.F.; Rocha, B.S.; Ferreira, N.R.; Ledo, A.; Laranjinha, J. Preparation, standardization, and measurement of nitric oxide solutions. Glob. J. Anal. Chem. 2011, 2, 272–284. [Google Scholar]
- Moore, E.G.; Gibson, Q.H. Cooperativity in the dissociation of nitric oxide from hemoglobin. J. Biol. Chem. 1976, 251, 2788–2794. [Google Scholar] [CrossRef]
- Canto, A.; Olivar, T.; Romero, F.J.; Miranda, M. Nitrosative Stress in Retinal Pathologies: Review. Antioxidants 2019, 8, 543. [Google Scholar] [CrossRef] [Green Version]
- Schmetterer, L.; Polak, K. Role of nitric oxide in the control of ocular blood flow. Prog. Retin. Eye Res. 2001, 20, 823–847. [Google Scholar] [CrossRef]
- Tummanapalli, S.S.; Kuppusamy, R.; Yeo, J.H.; Kumar, N.; New, E.J.; Willcox, M.D.P. The role of nitric oxide in ocular surface physiology and pathophysiology. Ocul. Surf. 2021, 21, 37–51. [Google Scholar] [CrossRef]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Heme Protein | kon(NO2−) (M−1 s−1) | kon(CO) (M−1 s−1) | Heme–Fe(II) Coordination |
|---|---|---|---|
| Mt-Nb(II) | 7.6 a | 5.5 × 104 b | 5C c |
| At-Nb(II) | 9.3 a | 2.3 × 105 b | n.a. |
| Dr-Nb(II) | 1.4 × 101 a | n.a. | 5C c |
| Hs-Nb(II) | 5.8 a | 1.0 × 105 b | 5C c |
| S-Hb(II) | 6.8 × 101 d | 3.0 × 105 e | 6C f |
| At-Hb(II) class 1 | 2.0 × 101 g | 4.6 × 105 h | 6C (60%) + 5C (40%) i |
| At-Hb(II) class 2 | 4.9 g | 1.0 × 106 h | 6C i |
| Rice nonsymbiotic Hb(II) class 1 | 8.3 × 101 d | 2.4 × 106 e | 6C (65%) + 5C (35%) j |
| Ma-Pgb(II) | 9.6 k 1.2 k | 2.1 × 107 l 1.1 × 106 l | 5C m |
| Cc-Mb(II) type 1 | 5.3 n | n.a. | n.a. |
| Cc-Mb(II) type 2 | 1.8 n | n.a. | n.a. |
| Efc-Mb(II) | 2.9 o | 5.5 × 105 p | 5C q |
| Pc-Mb(II) | 6.0 r | 5.0 × 105 h | 5C s |
| Mm-Ngb(II) | 5.1 t | 2.6 × 102 u | 6C v |
| Hs-Ngb(II) CysCD4-CysD5 | 1.2 × 10−1 w | 4.6 × 104 e | 6C x |
| Hs-Ngb(II) CysCD4/CysD5 | 6.2 × 10−2 y | 4.0 × 103 e | 6C x |
| Hs-Cygb(II) CysB2-CysE9 (monomer) | 3.2 × 101 z | 6.5 × 103 e | 6C x |
| Hs-Cygb(II) CysB2/CysE9 (monomer) | 6.3 × 10−1 aa | 1.2 × 103 e | 6C x |
| Hs-Cygb(II) CysB2/CysE9 (dimer) | 2.6 × 10−1 bb | n.a. | 6C cc |
| Hs-Hb(II) T-state | 1.2 × 10−1 r | 7.8 × 104 dd | 5C ee |
| Hs-Hb(II) R-state | 6.0 r | 1.1 × 107 ff | n.a. |
| Hs-Hp1-1:Hb(II) | 7.3 gg | 1.4 × 106 hh | n.a. |
| Hs-Hp2-2:Hb(II) | 1.2 × 101 gg | 5.2 × 106 hh | n.a. |
| Mt-trHbN(II) | 1.6 × 101 ii | 3.8 × 106 jj | 5C kk |
| Mt-trHbO(II) | 3.8 × 10−1 ii | 1.8 × 105 ll 1.4 × 104 ll | 5C mm |
| Cj-trHbP(II) q | 4.3 ii | 4.5 × 107 nn 1.9 × 106 nn | 5C oo |
| Hs-heme(II)-albumin | 1.3 pp | 4.0 × 106 qq | 5C rr |
| Efc-Cytc(II) | 7.3 × 10−2 ss | n.a. | 6C tt |
| Efc-Cytc(II) with CL | 2.6 ss | 1.0 × 107 uu | 5C vv |
| Carboxymethylated Efc-Cytc(II) | 1.6 ww | 1.6 × 106 xx | 5C yy |
| Carboxymethylated Efc-Cytc(II) with CL | 1.6 ww | n.a. | n.a. |
| Efc-MP-11(II) | 3.4 zz | 9.7 × 106 aaa | 5C bbb |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
De Simone, G.; di Masi, A.; Tundo, G.R.; Coletta, M.; Ascenzi, P. Nitrite Reductase Activity of Ferrous Nitrobindins: A Comparative Study. Int. J. Mol. Sci. 2023, 24, 6553. https://doi.org/10.3390/ijms24076553
De Simone G, di Masi A, Tundo GR, Coletta M, Ascenzi P. Nitrite Reductase Activity of Ferrous Nitrobindins: A Comparative Study. International Journal of Molecular Sciences. 2023; 24(7):6553. https://doi.org/10.3390/ijms24076553
Chicago/Turabian StyleDe Simone, Giovanna, Alessandra di Masi, Grazia R. Tundo, Massimo Coletta, and Paolo Ascenzi. 2023. "Nitrite Reductase Activity of Ferrous Nitrobindins: A Comparative Study" International Journal of Molecular Sciences 24, no. 7: 6553. https://doi.org/10.3390/ijms24076553