
Decreased SIRT1 Activity Is Involved in the Acute Injury Response of Chondrocytes to Ex Vivo Injurious Mechanical Overload
 and
and
Abstract
:1. Introduction
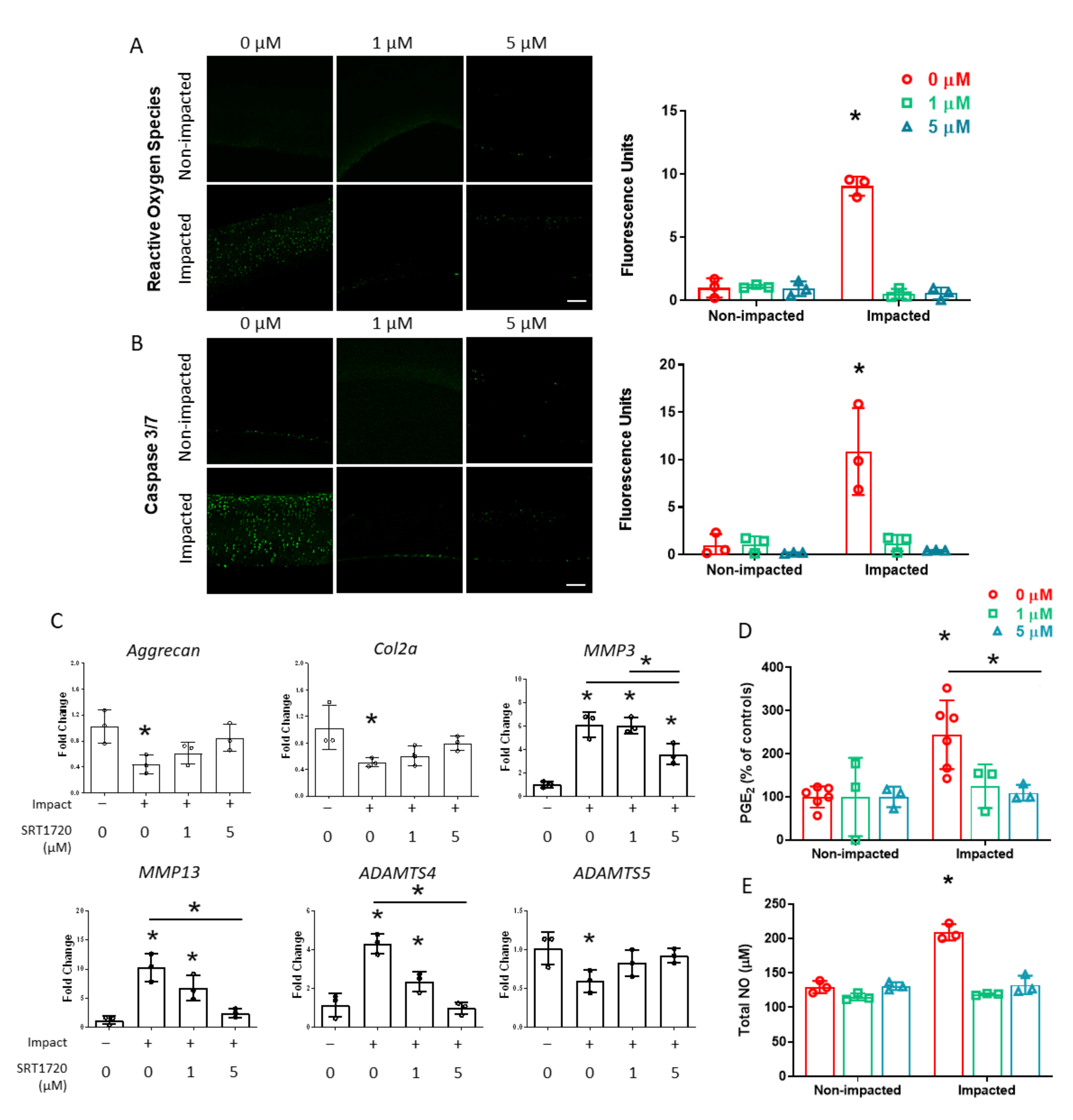
2. Results
2.1. Sublethal Cartilage Impact Is a Platform to Study Molecular Events in the Chondrocyte Injury Response
2.2. SIRT1 Enzyme Activity Decreases with Cartilage Impact and Is Blocked by SIRT1 Activator SRT1720
2.3. The Chondrocyte Injury Response 24 h Following Cartilage Impact Is Prevented with SIRT1 Activation
3. Discussion
4. Materials and Methods
4.1. Tissue Harvest and Impact
4.2. Fluorescent Staining and Confocal Imaging
4.3. SIRT1 Enzyme Activity
4.4. RNA Extraction and RT-PCR
4.5. PGE2 ELISA
4.6. Total NO Assay
4.7. Quantification and Statistical Analysis
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Cui, A.; Li, H.; Wang, D.; Zhong, J.; Chen, Y.; Lu, H. Global, Regional Prevalence, Incidence and Risk Factors of Knee Osteoarthritis in Population-Based Studies. EClinicalMedicine 2020, 29, 100587. [Google Scholar] [CrossRef] [PubMed]
- Anderson, D.D.; Chubinskaya, S.; Guilak, F.; Martin, J.A.; Oegema, T.R.; Olson, S.A.; Buckwalter, J.A. Post-traumatic Osteoarthritis: Improved Understanding and Opportunities for Early Intervention. J. Orthop. Res. 2011, 29, 802–809. [Google Scholar] [CrossRef] [PubMed]
- Gelber, A.C.; Hochberg, M.C.; Mead, L.A.; Wang, N.Y.; Wigley, F.M.; Klag, M.J. Joint Injury in Young Adults and Risk for Subsequent Knee and Hip Osteoarthritis. Ann. Intern. Med. 2000, 133, 321–328. [Google Scholar] [CrossRef] [PubMed]
- Wilder, F.V.; Hall, B.J.; Barrett, J.P.; Lemrow, N.B. History of Acute Knee Injury and Osteoarthritis of the Knee: A Prospective Epidemiological Assessment. The Clearwater Osteoarthritis Study. Osteoarthr. Cartil. 2002, 10, 611–616. [Google Scholar] [CrossRef]
- Muthuri, S.G.; McWilliams, D.F.; Doherty, M.; Zhang, W. History of Knee Injuries and Knee Osteoarthritis: A Meta-Analysis of Observational Studies. Osteoarthr. Cartil. 2011, 19, 1286–1293. [Google Scholar] [CrossRef]
- Snoeker, B.; Turkiewicz, A.; Magnusson, K.; Frobell, R.; Yu, D.; Peat, G.; Englund, M. Risk of Knee Osteoarthritis after Different Types of Knee Injuries in Young Adults: A Population-Based Cohort Study. Br. J. Sport. Med. 2020, 54, 725–730. [Google Scholar] [CrossRef]
- Buckwalter, J.A. The Role of Mechanical Forces in the Initiation and Progression of Osteoarthritis. HSS J. 2012, 8, 37–38. [Google Scholar] [CrossRef]
- Kramer, W.C.; Hendricks, K.J.; Wang, J. Pathogenetic Mechanisms of Posttraumatic Osteoarthritis: Opportunities for Early Intervention. Int. J. Clin. Exp. Med. 2011, 4, 285–298. [Google Scholar]
- McKinley, T.O.; Borrelli, J.; D’Lima, D.D.; Furman, B.D.; Giannoudis, P.V. Basic Science of Intra-Articular Fractures and Posttraumatic Osteoarthritis. J. Orthop. Trauma 2010, 24, 567–570. [Google Scholar] [CrossRef]
- D’Lima, D.D.; Hashimoto, S.; Chen, P.C.; Colwell, C.W.; Lotz, M.K. Human Chondrocyte Apoptosis in Response to Mechanical Injury. Osteoarthr. Cartil. 2001, 9, 712–719. [Google Scholar] [CrossRef]
- Brouillette, M.J.; Ramakrishnan, P.S.; Wagner, V.M.; Sauter, E.E.; Journot, B.J.; McKinley, T.O.; Martin, J.A. Strain-Dependent Oxidant Release in Articular Cartilage Originates from Mitochondria. Biomech. Model. Mechanobiol. 2014, 13, 565–572. [Google Scholar] [CrossRef]
- Natoli, R.M.; Scott, C.C.; Athanasiou, K.A. Temporal Effects of Impact on Articular Cartilage Cell Death, Gene Expression, Matrix Biochemistry, and Biomechanics. Ann. Biomed. Eng. 2008, 36, 780–792. [Google Scholar] [CrossRef]
- Dwivedi, G.; Flaman, L.; Alaybeyoglu, B.; Struglics, A.; Frank, E.H.; Chubinskya, S.; Trippel, S.B.; Rosen, V.; Cirit, M.; Grodzinsky, A.J. Inflammatory Cytokines and Mechanical Injury Induce Post-Traumatic Osteoarthritis-like Changes in a Human Cartilage-Bone-Synovium Microphysiological System. Arthritis Res. Ther. 2022, 24, 198. [Google Scholar] [CrossRef]
- Diekman, B.O.; Collins, J.A.; Loeser, R.F. Does Joint Injury Make Young Joints Old? J. Am. Acad. Orthop. Surg. 2018, 26, e455. [Google Scholar] [CrossRef]
- Viswanathan, M.; Guarente, L. Regulation of Caenorhabditis Elegans Lifespan by Sir-2.1 Transgenes. Nature 2011, 477, E1–E2. [Google Scholar] [CrossRef]
- Imai, S.I.; Guarente, L. It Takes Two to Tango: Nad+ and Sirtuins in Aging/Longevity Control. NPJ Aging Mech. Dis. 2016, 2, 16017. [Google Scholar] [CrossRef]
- Mouchiroud, L.; Houtkooper, R.H.; Moullan, N.; Katsyuba, E.; Ryu, D.; Cantó, C.; Mottis, A.; Jo, Y.S.; Viswanathan, M.; Schoonjans, K.; et al. XThe NAD+/Sirtuin Pathway Modulates Longevity through Activation of Mitochondrial UPR and FOXO Signaling. Cell 2013, 154, 430–441. [Google Scholar] [CrossRef]
- Satoh, A.; Brace, C.S.; Rensing, N.; Cliften, P.; Wozniak, D.F.; Herzog, E.D.; Yamada, K.A.; Imai, S.I. Sirt1 Extends Life Span and Delays Aging in Mice through the Regulation of Nk2 Homeobox 1 in the DMH and LH. Cell Metab. 2013, 18, 416–430. [Google Scholar] [CrossRef]
- Schmeisser, K.; Mansfeld, J.; Kuhlow, D.; Weimer, S.; Priebe, S.; Heiland, I.; Birringer, M.; Groth, M.; Segref, A.; Kanfi, Y.; et al. Role of Sirtuins in Lifespan Regulation Is Linked to Methylation of Nicotinamide. Nat. Chem. Biol. 2013, 9, 693–700. [Google Scholar] [CrossRef]
- Stumpferl, S.W.; Brand, S.E.; Jiang, J.C.; Korona, B.; Tiwari, A.; Dai, J.; Seo, J.G.; Jazwinski, S.M. Natural Genetic Variation in Yeast Longevity. Genome Res. 2012, 22, 1963–1973. [Google Scholar] [CrossRef]
- Gomes, A.P.; Price, N.L.; Ling, A.J.Y.; Moslehi, J.J.; Montgomery, M.K.; Rajman, L.; White, J.P.; Teodoro, J.S.; Wrann, C.D.; Hubbard, B.P.; et al. Declining NAD+ Induces a Pseudohypoxic State Disrupting Nuclear-Mitochondrial Communication during Aging. Cell 2013, 155, 1624–1638. [Google Scholar] [CrossRef] [PubMed]
- Massudi, H.; Grant, R.; Braidy, N.; Guest, J.; Farnsworth, B.; Guillemin, G.J. Age-Associated Changes in Oxidative Stress and NAD+ Metabolism in Human Tissue. PLoS ONE 2012, 7, e42357. [Google Scholar] [CrossRef] [PubMed]
- Houtkooper, R.H.; Pirinen, E.; Auwerx, J. Sirtuins as Regulators of Metabolism and Healthspan. Nat. Rev. Mol. Cell Biol. 2012, 13, 225–238. [Google Scholar] [CrossRef] [PubMed]
- Guarente, L. Sirtuins, Aging, and Medicine. N. Engl. J. Med. 2011, 364, 2235–2244. [Google Scholar] [CrossRef] [PubMed]
- Haigis, M.C.; Sinclair, D.A. Mammalian Sirtuins: Biological Insights and Disease Relevance. Annu. Rev. Pathol. Mech. Dis. 2010, 5, 253–295. [Google Scholar] [CrossRef]
- Matsuzaki, T.; Matsushita, T.; Takayama, K.; Matsumoto, T.; Nishida, K.; Kuroda, R.; Kurosaka, M. Disruption of Sirt1 in Chondrocytes Causes Accelerated Progression of Osteoarthritis under Mechanical Stress and during Ageing in Mice. Ann. Rheum. Dis. 2014, 73, 1397–1404. [Google Scholar] [CrossRef]
- Matsuzaki, T.; Matsushita, T.; Kubo, S.; Matsumoto, T.; Sasaki, H.; Takayama, K.; Kurosaka, M.; Kuroda, R. Disruption of Sirt1 in Chondrocytes Causes Accelerated Development of Osteoarthritis Induced by Joint Instability in the Mouse. Osteoarthr. Cartil. 2012, 20, S25. [Google Scholar] [CrossRef]
- Matsushita, T.; Sasaki, H.; Takayama, K.; Ishida, K.; Matsumoto, T.; Kubo, S.; Matsuzaki, T.; Nishida, K.; Kurosaka, M.; Kuroda, R. The Overexpression of SIRT1 Inhibited Osteoarthritic Gene Expression Changes Induced by Interleukin-1β in Human Chondrocytes. J. Orthop. Res. 2013, 31, 531–537. [Google Scholar] [CrossRef]
- Li, W.; Cai, L.; Zhang, Y.; Cui, L.; Shen, G. Intra-Articular Resveratrol Injection Prevents Osteoarthritis Progression in a Mouse Model by Activating SIRT1 and Thereby Silencing HIF-2α. J. Orthop. Res. 2015, 33, 1061–1070. [Google Scholar] [CrossRef]
- Qin, N.; Wei, L.; Li, W.; Yang, W.; Cai, L.; Qian, Z.; Wu, S. Local Intra-Articular Injection of Resveratrol Delays Cartilage Degeneration in C57BL/6 Mice by Inducing Autophagy via AMPK/MTOR Pathway. J. Pharmacol. Sci. 2017, 134, 166–174. [Google Scholar] [CrossRef]
- Nishida, K.; Matsushita, T.; Takayama, K.; Tanaka, T.; Miyaji, N.; Ibaraki, K.; Araki, D.; Kanzaki, N.; Matsumoto, T.; Kuroda, R. Intraperitoneal Injection of the SIRT1 Activator SRT1720 Attenuates the Progression of Experimental Osteoarthritis in Mice. Bone Jt. Res. 2018, 7, 252–262. [Google Scholar] [CrossRef]
- Wang, J.; Gao, J.S.; Chen, J.W.; Li, F.; Tian, J. Effect of Resveratrol on Cartilage Protection and Apoptosis Inhibition in Experimental Osteoarthritis of Rabbit. Rheumatol. Int. 2012, 32, 1541–1548. [Google Scholar] [CrossRef]
- Miyaji, N.; Nishida, K.; Tanaka, T.; Araki, D.; Kanzaki, N.; Hoshino, Y.; Kuroda, R.; Matsushita, T. Inhibition of Knee Osteoarthritis Progression in Mice by Administering SRT2014, an Activator of Silent Information Regulator 2 Ortholog 1. Cartilage 2021, 13, 1356S–1366S. [Google Scholar] [CrossRef]
- Hubbard, B.P.; Gomes, A.P.; Dai, H.; Li, J.; Case, A.W.; Considine, T.; Riera, T.V.; Lee, J.E.; Yen, E.S.; Lamming, D.W.; et al. Evidence for a Common Mechanism of SIRT1 Regulation by Allosteric Activators. Science 2013, 339, 1216–1219. [Google Scholar] [CrossRef]
- Joos, H.; Hogrefe, C.; Rieger, L.; Dürselen, L.; Ignatius, A.; Brenner, R.E. Single Impact Trauma in Human Early-Stage Osteoarthritic Cartilage: Implication of Prostaglandin D2 but No Additive Effect of IL-1β on Cell Survival. Int. J. Mol. Med. 2011, 28, 271–277. [Google Scholar] [CrossRef]
- Takayama, K.; Ishida, K.; Matsushita, T.; Fujita, N.; Hayashi, S.; Sasaki, K.; Tei, K.; Kubo, S.; Matsumoto, T.; Fujioka, H.; et al. SIRT1 Regulation of Apoptosis of Human Chondrocytes. Arthritis Rheum. 2009, 60, 2731–2740. [Google Scholar] [CrossRef]
- Song, R.H.; Tortorella, M.D.; Malfait, A.M.; Alston, J.T.; Yang, Z.; Arner, E.C.; Griggs, D.W. Aggrecan Degradation in Human Articular Cartilage Explants Is Mediated by Both ADAMTS-4 and ADAMTS-5. Arthritis Rheum. 2007, 56, 575–585. [Google Scholar] [CrossRef]
- Tortorella, M.D.; Malfait, A.M.; Deccico, C.; Arner, E. The Role of ADAM-TS4 (Aggrecanase-1) and ADAM-TS5 (Aggrecanase-2) in a Model of Cartilage Degradation. Osteoarthr. Cartil. 2001, 9, 539–552. [Google Scholar] [CrossRef]
- Garrity, J.T.; Stoker, A.M.; Sims, H.J.; Cook, J.L. Improved Osteochondral Allograft Preservation Using Serum-Free Media at Body Temperature. Am. J. Sport. Med. 2012, 40, 2542–2548. [Google Scholar] [CrossRef]
- Bonitsky, C.M.; McGann, M.E.; Selep, M.J.; Ovaert, T.C.; Trippel, S.B.; Wagner, D.R. Genipin Crosslinking Decreases the Mechanical Wear and Biochemical Degradation of Impacted Cartilage in Vitro. J. Orthop. Res. 2017, 35, 558–565. [Google Scholar] [CrossRef]
- Schindelin, J.; Arganda-Carreras, I.; Frise, E.; Kaynig, V.; Longair, M.; Pietzsch, T.; Preibisch, S.; Rueden, C.; Saalfeld, S.; Schmid, B.; et al. Fiji: An Open-Source Platform for Biological-Image Analysis. Nat. Methods 2012, 9, 676–682. [Google Scholar] [CrossRef] [PubMed]
- Le Bleu, H.K.; Kamal, F.A.; Kelly, M.; Ketz, J.P.; Zuscik, M.J.; Elbarbary, R.A. Extraction of High-Quality RNA from Human Articular Cartilage. Anal. Biochem. 2017, 518, 134–138. [Google Scholar] [CrossRef] [PubMed]
- Idrees, M.; Xu, L.; Song, S.H.; Joo, M.D.; Lee, K.L.; Muhammad, T.; El Sheikh, M.; Sidrat, T.; Kong, I.K. PTPN11 (SHP2) Is Indispensable for Growth Factors and Cytokine Signal Transduction During Bovine Oocyte Maturation and Blastocyst Development. Cells 2019, 8, 1272. [Google Scholar] [CrossRef] [PubMed]
- Tilwani, R.K.; Bader, D.L.; Chowdhury, T.T. Functional Biomaterials Biomechanical Conditioning Enhanced Matrix Synthesis in Nucleus Pulposus Cells Cultured in Agarose Constructs with TGFβ. J. Funct. Biomater 2012, 3, 23–36. [Google Scholar] [CrossRef]
- Shi, S.; Mercer, S.; Eckert, G.J.; Trippel, S.B. Regulation of Articular Chondrocyte Catabolic Genes by Growth Factor Interaction. J. Cell. Biochem. 2019, 120, 11127–11139. [Google Scholar] [CrossRef]
- Park, Y.; Lutolf, M.P.; Hubbell, J.A.; Hunziker, E.B.; Wong, M. Bovine Primary Chondrocyte Culture in Synthetic Matrix Metalloproteinase-Sensitive Poly(Ethylene Glycol)-Based Hydrogels as a Scaffold for Cartilage Repair. Tissue Eng. 2004, 10, 515–522. [Google Scholar] [CrossRef]
- Al-Sabah, A.; Stadnik, P.; Gilbert, S.J.; Duance, V.C.; Blain, E.J. Importance of Reference Gene Selection for Articular Cartilage Mechanobiology Studies. Osteoarthr. Cartil. 2016, 24, 719–730. [Google Scholar] [CrossRef]




{kind=link}
{kind=link}
{kind=link}
| Gene | Sequence | Reference |
|---|---|---|
| SIRT1 | 5′-CAACGGTTTCCATTCGTGTG-3′ | [43] |
| 5′-GTTCGAGGATCTGTGCCAAT-3′ | ||
| Aggrecan | 5′-TGGTGTTTGTGACTCTGAGG-3′ | [44] |
| 5′-GATGAAGTAGCAGGGGATGG-3′ | ||
| Col2a | 5′-AAACCCGAACCCAGAACC-3′ | [44] |
| 5′-AAGTCCGAACTGTGAGAGG-3′ | ||
| MMP3 | 5′-TGTGCTCAGCCTATCCACTG-3′ | [45] |
| 5′-AGCTTTCCTGTCACCTCCAA-3′ | ||
| MMP13 | 5′-CAATGTTTTTCCTCGAACTCTCAA-3′ | [46] |
| 5′-TTCCACTTCAGAATGAGTCAGATCA-3′ | ||
| ADAMTS4 | 5′-GAAGCAATGCACTGGTCTGA-3′ | [45] |
| 5′-CTAGGAGACAGTGCCCGAAG-3′ | ||
| ADAMTS5 | 5′-TGCAGATTCTTGCCACAGAC-3′ | [45] |
| 5′-CTTTTGGAGCCGACTTCTTG-3′ | ||
| 18s | 5′-GCAATTATTCCCCATGAACG-3′ | [47] |
| 5′-GCCTCACTAAACCATCCAA-3′ |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Karnik, S.; Noori-Dokht, H.; Williams, T.; Joukar, A.; Trippel, S.B.; Sankar, U.; Wagner, D.R. Decreased SIRT1 Activity Is Involved in the Acute Injury Response of Chondrocytes to Ex Vivo Injurious Mechanical Overload. Int. J. Mol. Sci. 2023, 24, 6521. https://doi.org/10.3390/ijms24076521
Karnik S, Noori-Dokht H, Williams T, Joukar A, Trippel SB, Sankar U, Wagner DR. Decreased SIRT1 Activity Is Involved in the Acute Injury Response of Chondrocytes to Ex Vivo Injurious Mechanical Overload. International Journal of Molecular Sciences. 2023; 24(7):6521. https://doi.org/10.3390/ijms24076521
Chicago/Turabian StyleKarnik, Sonali, Hessam Noori-Dokht, Taylor Williams, Amin Joukar, Stephen B. Trippel, Uma Sankar, and Diane R. Wagner. 2023. "Decreased SIRT1 Activity Is Involved in the Acute Injury Response of Chondrocytes to Ex Vivo Injurious Mechanical Overload" International Journal of Molecular Sciences 24, no. 7: 6521. https://doi.org/10.3390/ijms24076521