
Flavonoid Synthesis-Related Genes Determine the Color of Flower Petals in Brassica napus L.
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results
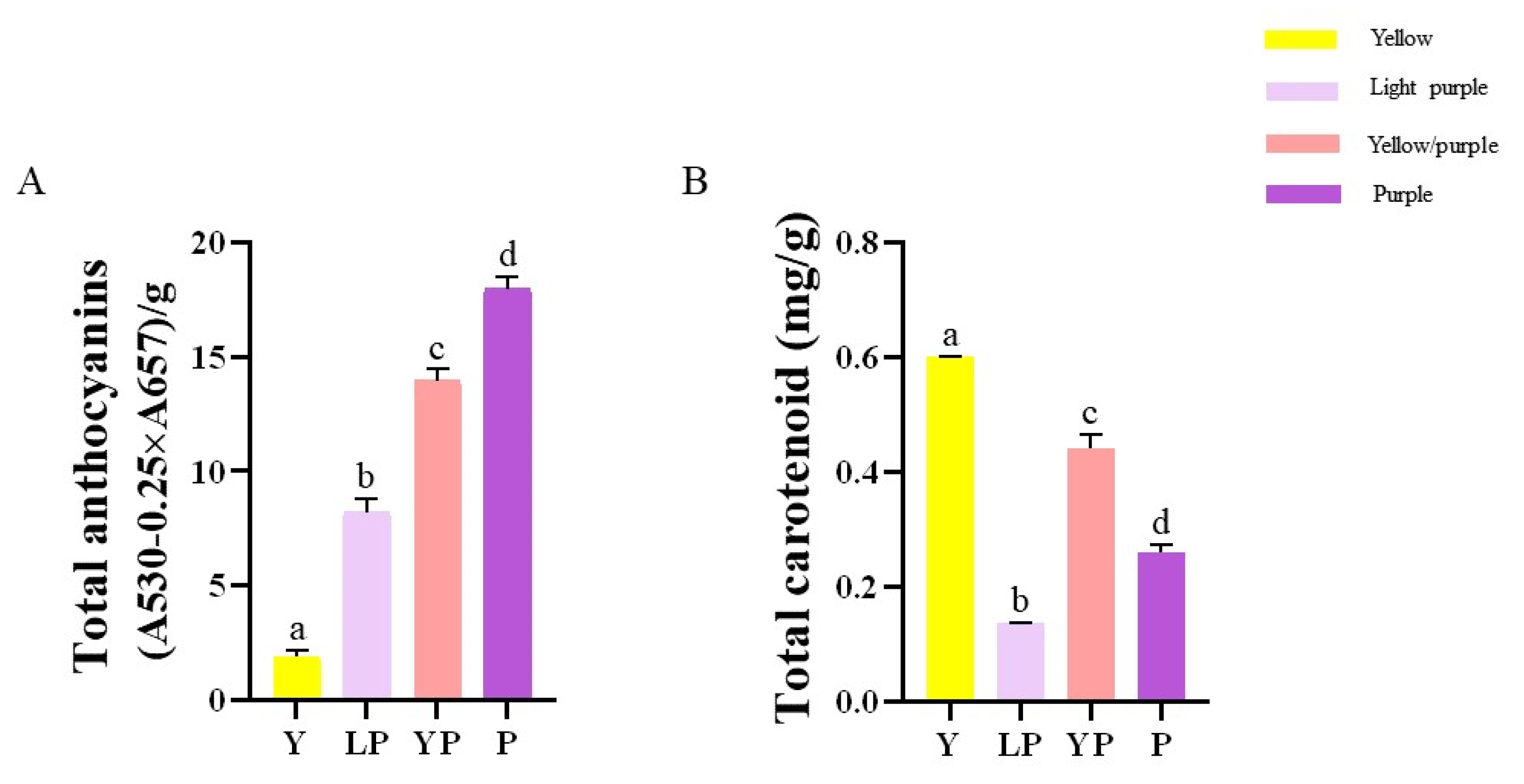
2.1. The Anthocyanin and Carotenoid Contents in the Petals of the Four Brassica napus L. Cultivars
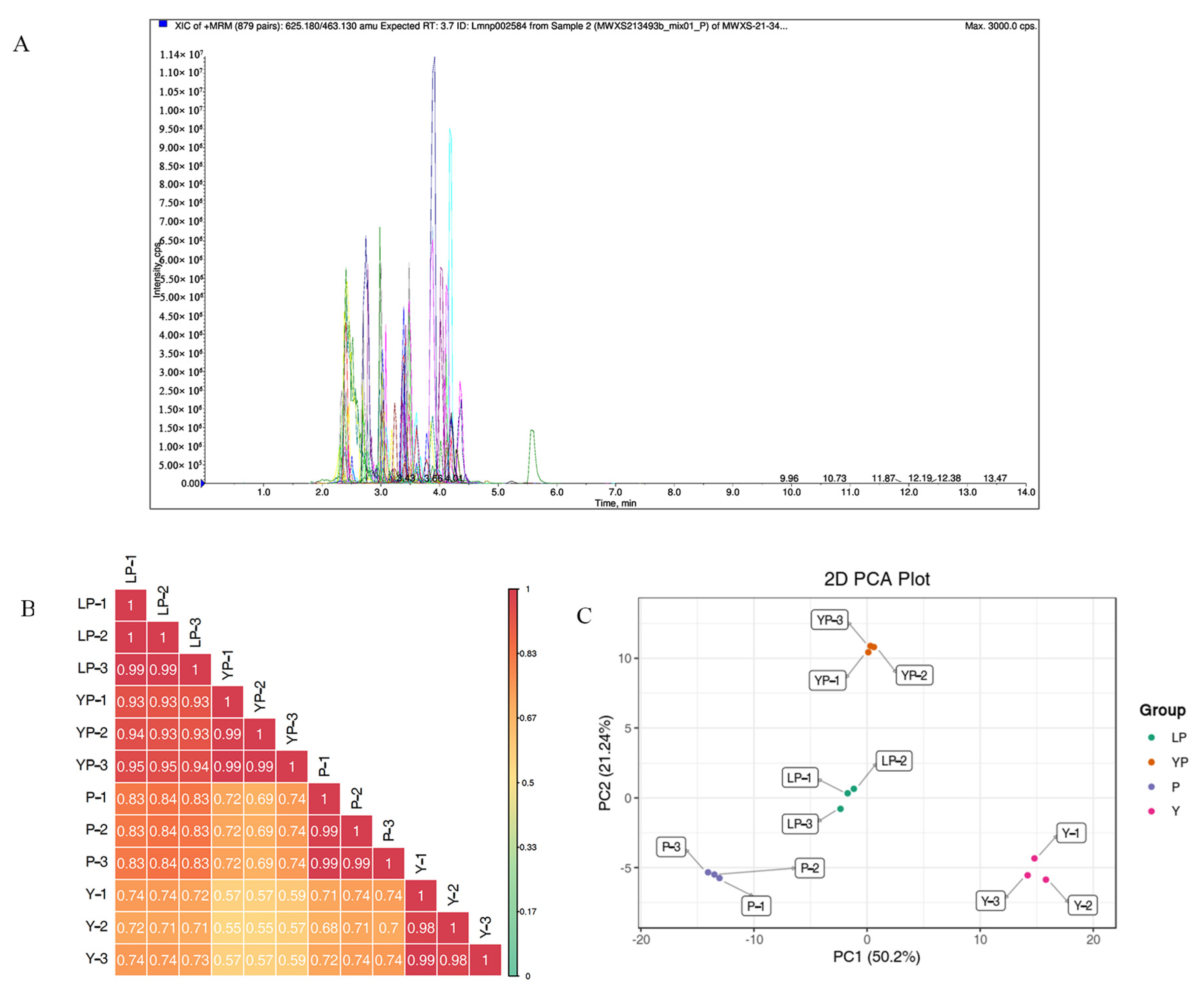
2.2. Widely Targeted Metabolomic Analysis of the Four Brassica napus L. Cultivars
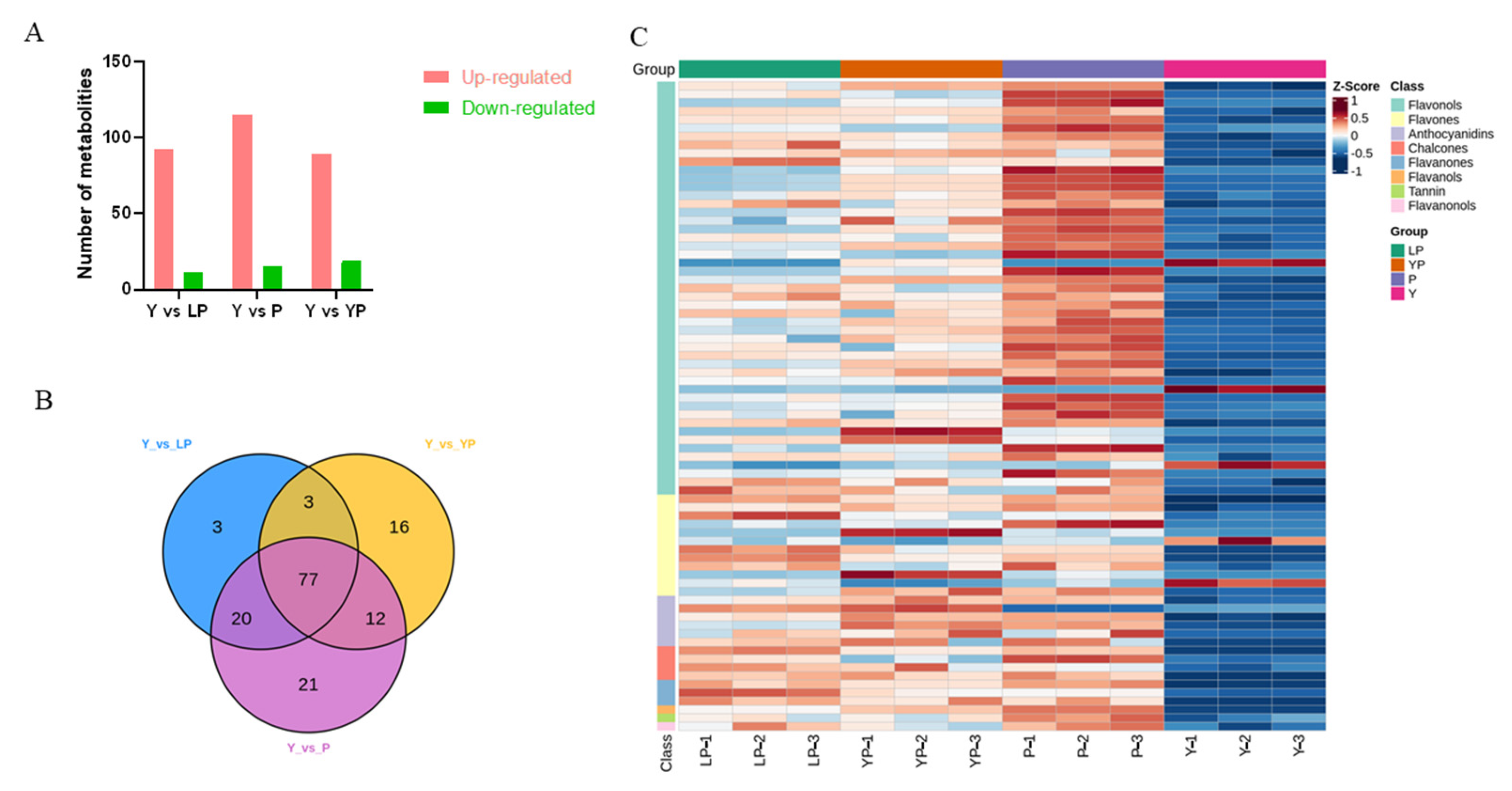
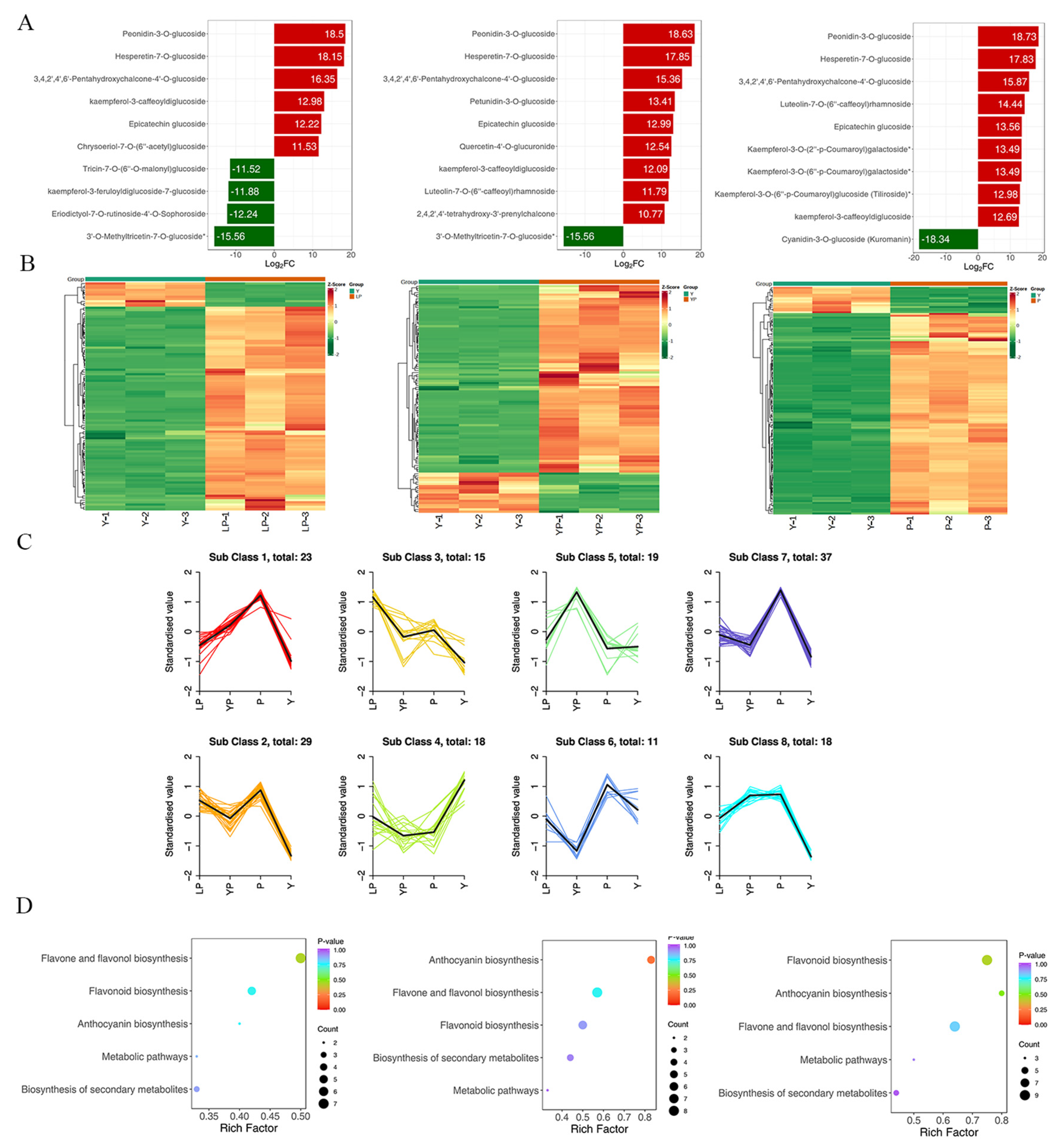
2.3. Metabolic Differences among the Four Petal Colours of Brassica napus L.
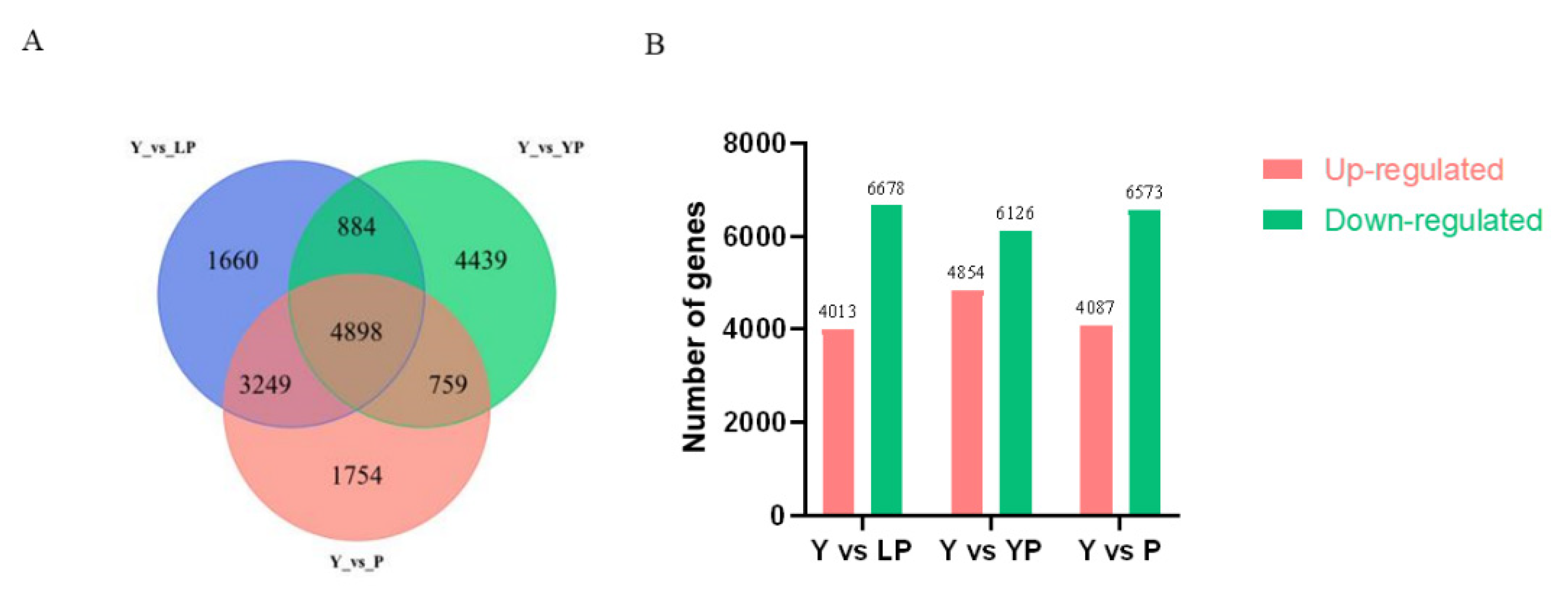
2.4. Transcriptome and Differential Gene Expression Analyses
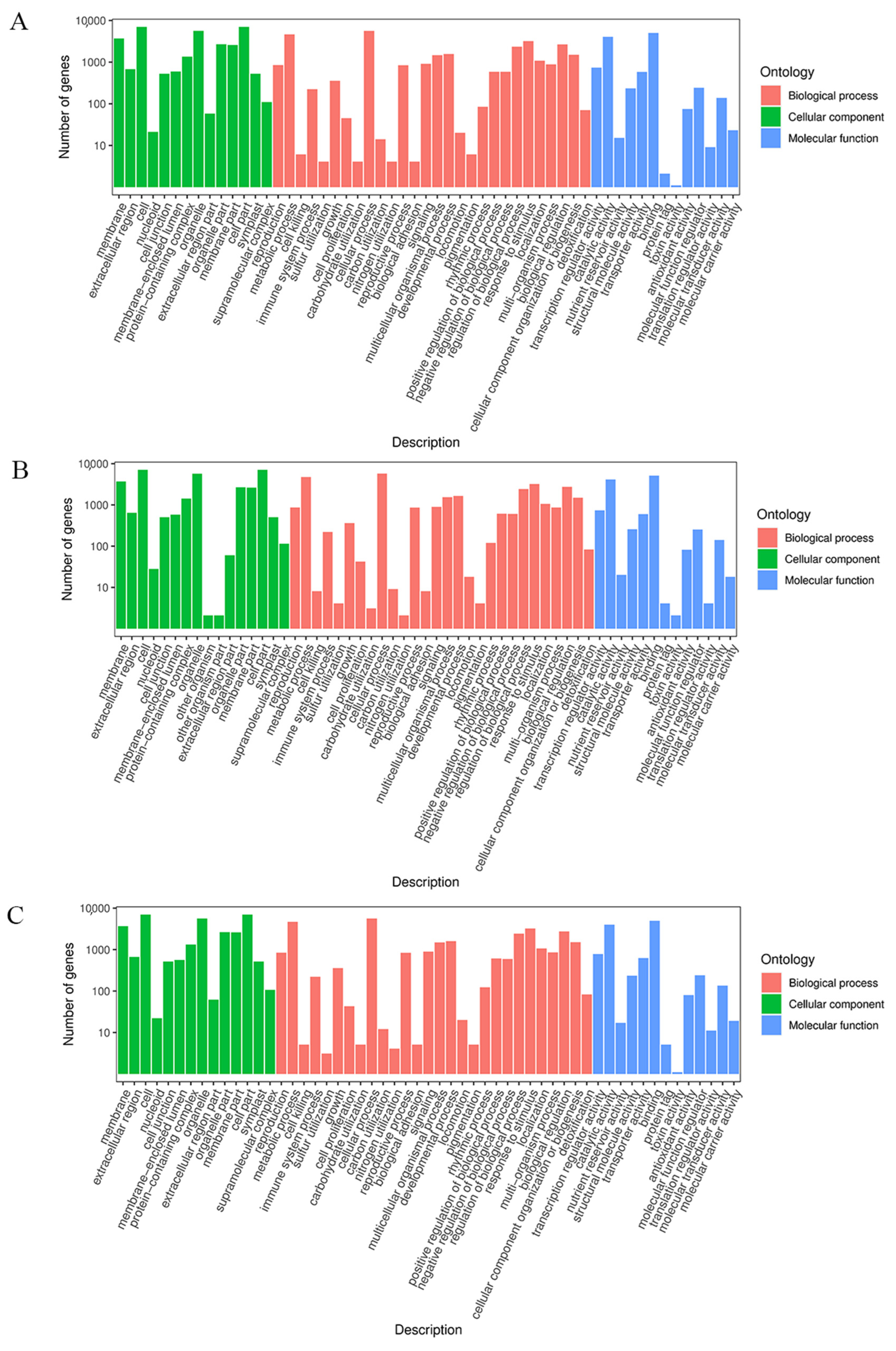
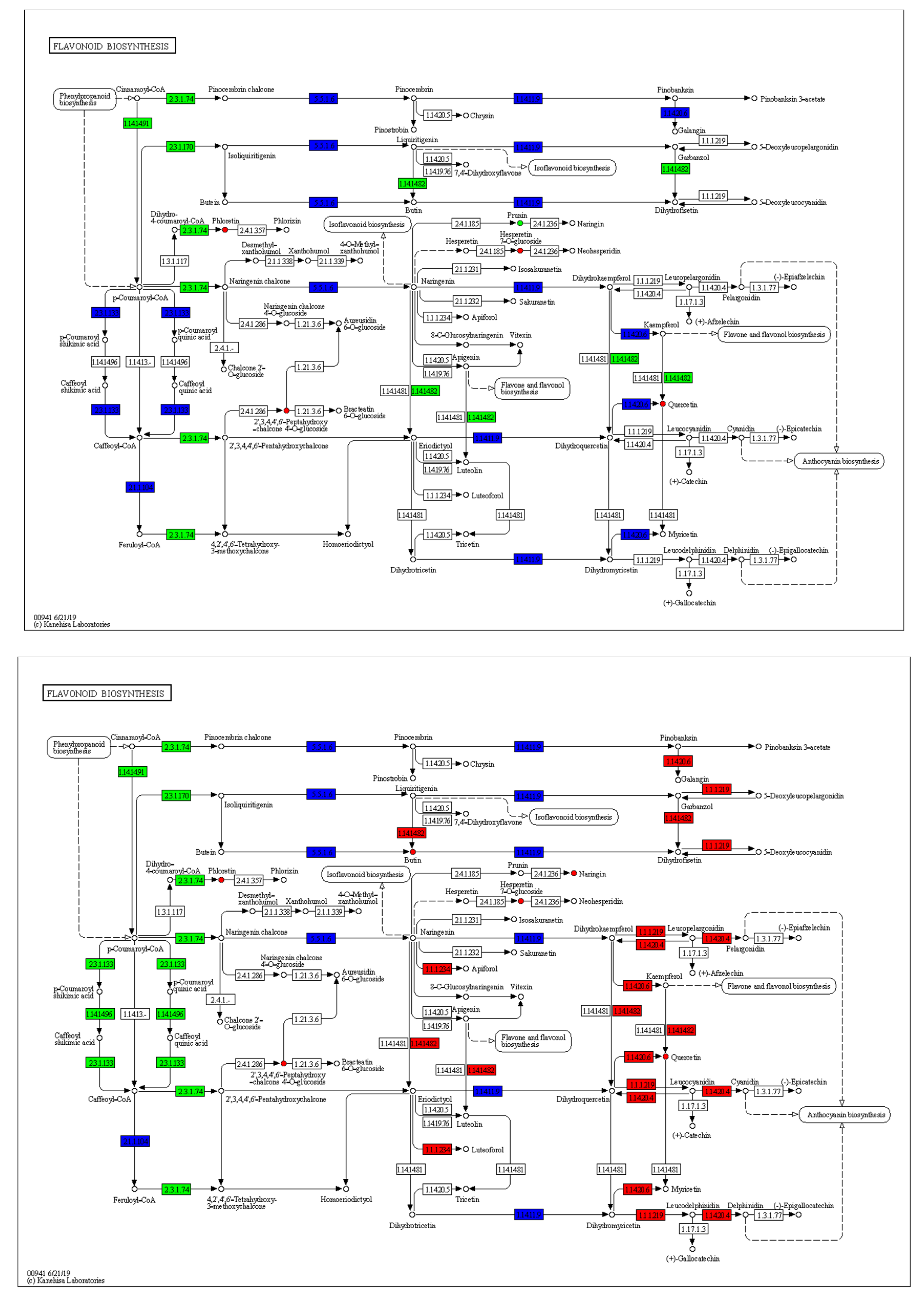
2.5. GO and KEGG Pathway Analysis of the DEGs
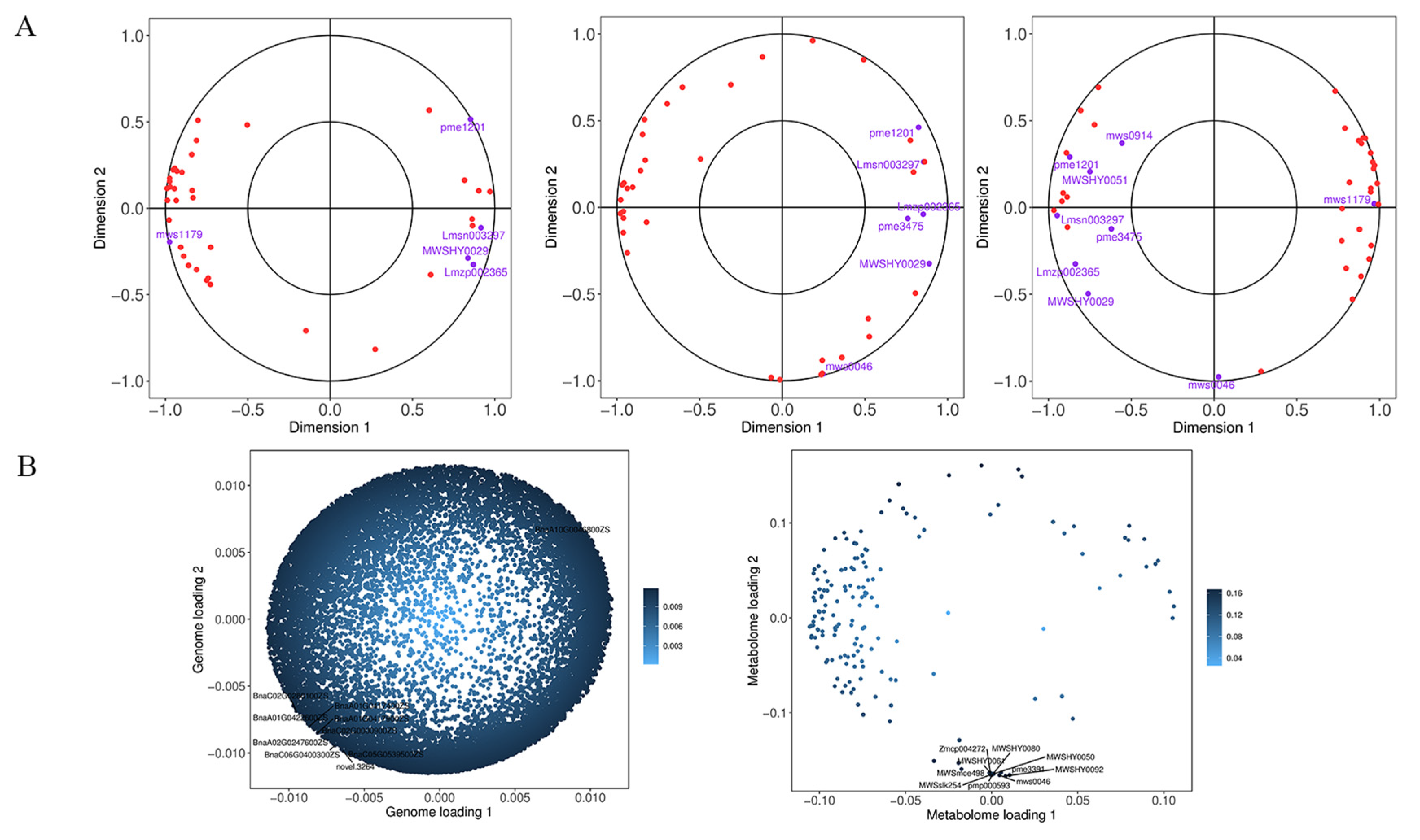
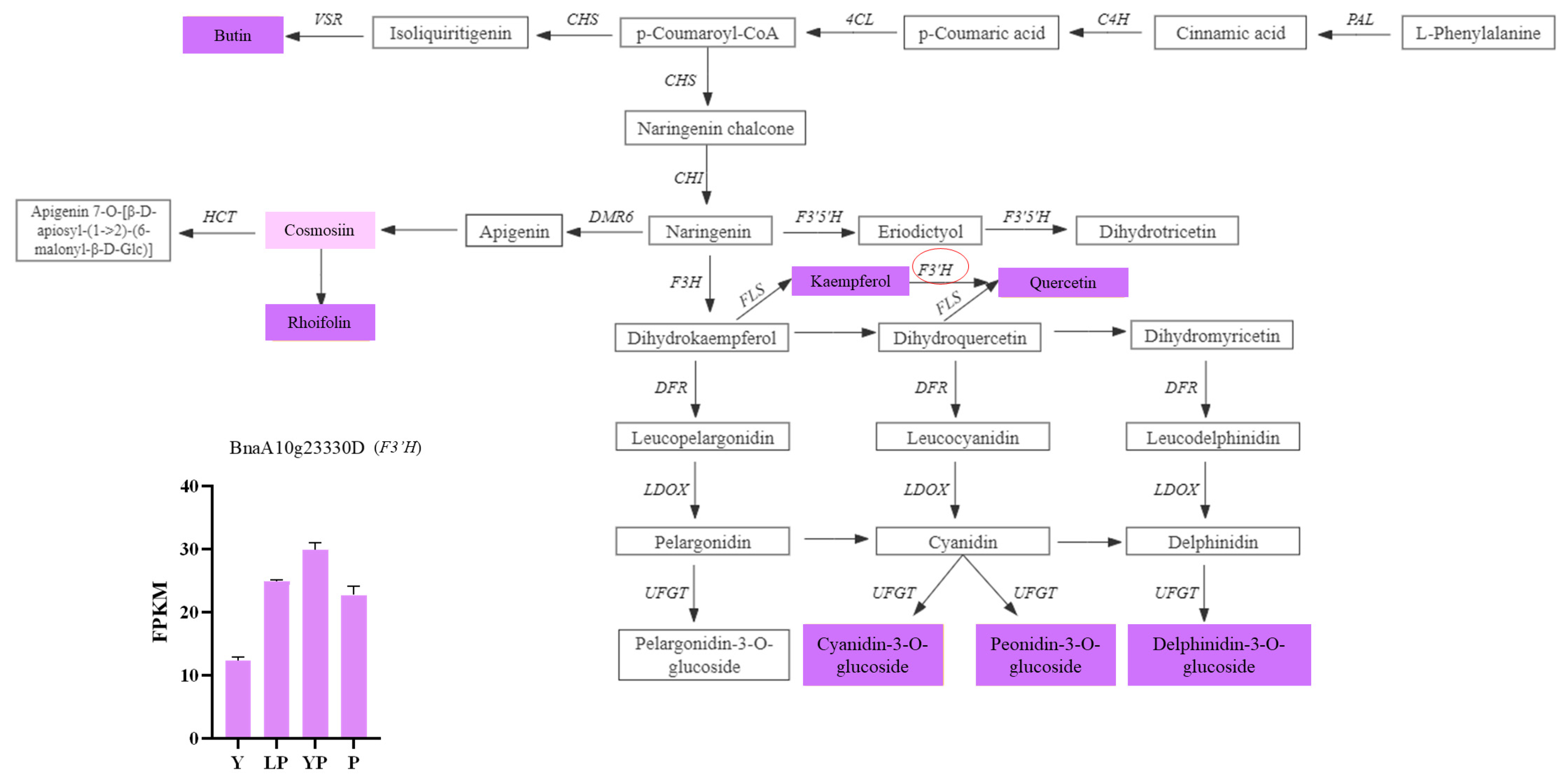
2.6. Combined Transcriptomic and Metabolomic Analyses
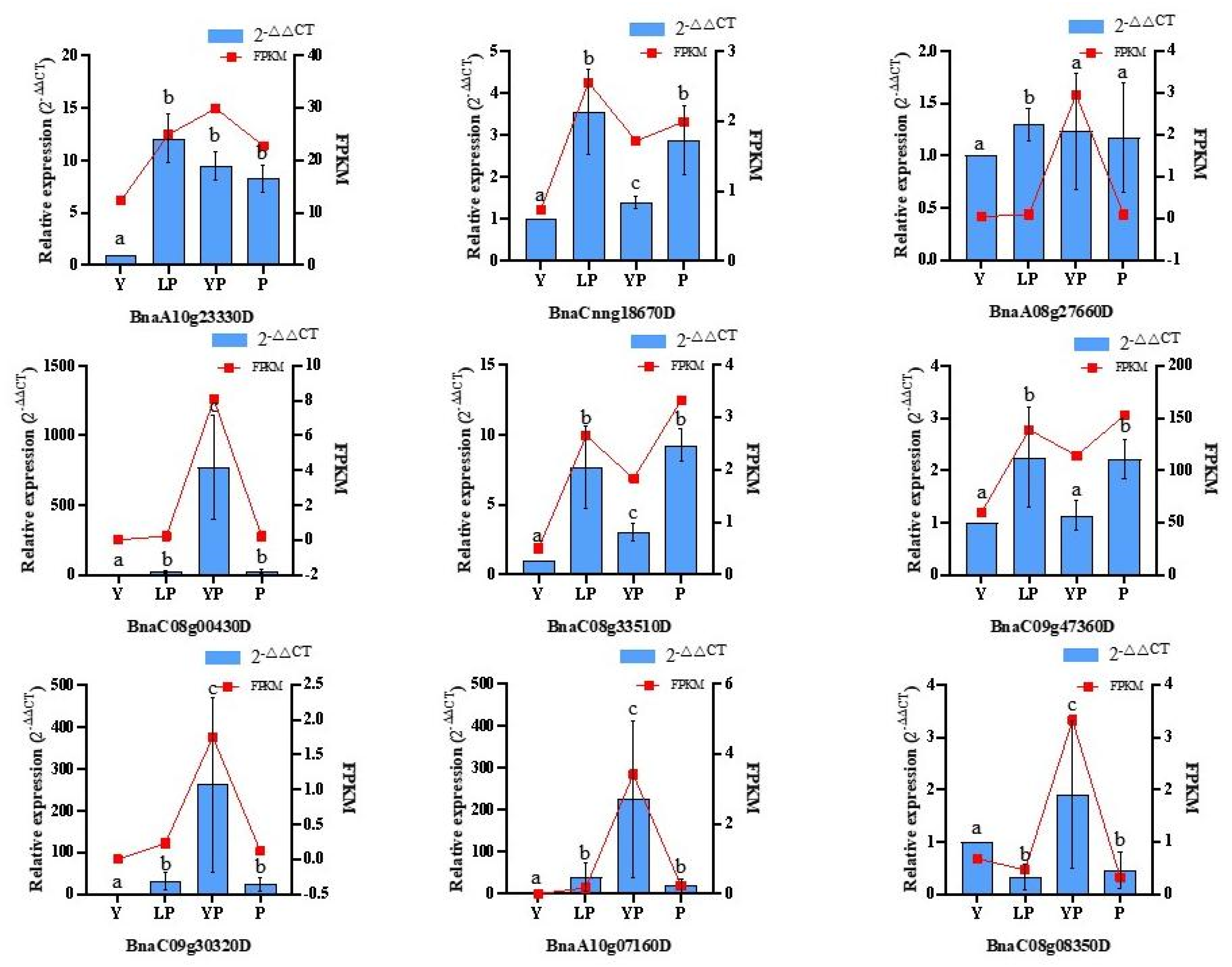
2.7. qRT-PCR Analysis of DEGs in the Flavonoid Metabolic Pathway
3. Materials and Methods
3.1. Plant Materials
3.2. Metabolomic Analysis
3.2.1. Determination of Carotenoid and Anthocyanin Contents
3.2.2. Metabolite Extraction and Ultra Performance Liquid Chromatography-MS/MS Conditions
3.2.3. Qualitative and Quantitative Metabolite Analyses
3.3. Transcriptome Sequencing and Data Analysis
3.3.1. RNA Extraction and RNA-Seq
3.3.2. Transcriptome Sequencing
3.3.3. qPCR Validation of the DEGs
3.3.4. Transcriptome Data Analysis
3.4. Integrated Metabolomic and Transcriptomic Analyses
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Zhang, Y.X.; Zhao, F.; Zhang, H.L. Rapeseed industry development status quo, problems and countermeasures of China. World Agric. 2015, 96–99, 203. [Google Scholar]
- Chen, Z.B.; Yu, J. Research strategies based on the analysis of rapeseed production in China. Chin. J. Oil Crop Sci. 2010, 32, 303–308. [Google Scholar]
- Taylor, L.P.; Grotewold, E. Flavonoids as developmental regulators. Front. Plant Sci. 2005, 8, 317–323. [Google Scholar] [CrossRef] [PubMed]
- Chen, D.Z.; Liu, Y.; Fu, W.Q.; Ge, X.H.; Li, Z.Y. Progress on genetics and breeding of rapeseed (Brassica napus L.) with coloured flowers. Chin. J. Oil Crop Sci. 2019, 41, 309–316. [Google Scholar]
- Yin, C.F.; Guan, C.Y. A Review of Rapaseed Flower Color. Crop Res. 2013, 27, 403–410. [Google Scholar]
- Tian, L.S.; Niu, Y.Z.; Yu, Q.Q.; Guo, S.X.; Li, L. Genetic Analysis of White Flower Color with Mixed Model of Major Gene Plus Polygene in Brassica napus L. Sci. Agric. Sin. 2009, 42, 3987–3995. [Google Scholar]
- Zhang, B.; Liu, C.; Wang, Y.; Yao, X.; Wang, F.; Wu, J.; King, G.J.; Liu, K. Disruption of a CAROTENOID CLEAVAGE DIOXYGENASE 4 gene converts flower colour from white to yellow in Brassica species. New Phytol. 2015, 206, 1513–1526. [Google Scholar] [CrossRef]
- Zhang, X.; Li, R.; Chen, L.; Niu, S.; Gao, J.; Wen, J.; Yi, B.; Ma, C.; Tu, J.; Fu, T.; et al. Fine-mapping and candidate gene analysis of the Brassica juncea white-flowered mutant Bjpc2 using the whole-genome resequencing. Mol. Genet. Genom. 2018, 293, 359–370. [Google Scholar] [CrossRef]
- Jia, L.; Wang, J.; Wang, R.; Duan, M.; Qiao, C.; Chen, X.; Ma, G.; Zhou, X.; Zhu, M.; Jing, F.; et al. Comparative transcriptomic and metabolomic analyses of carotenoid biosynthesis reveal the basis of white petal color in Brassica napus. Planta 2021, 253, 8. [Google Scholar] [CrossRef]
- Lee, S.; Lee, S.C.; Byun, D.H.; Lee, D.Y.; Park, J.Y.; Lee, J.H.; Lee, H.O.; Sung, S.H.; Yang, T.J. Association of molecular markers derived from the BrCRTISO1 gene with prolycopene-enriched orange-colored leaves in Brassica rapa. Theor. Appl. Genet. 2014, 127, 179–191. [Google Scholar] [CrossRef]
- Wu, N.R. Transcriptome Analysis of Different Flower Colors in Brassica napus. Master’s Thesis, Hunan Agriculture University, Changsha, China, 2018. [Google Scholar]
- Xu, J.; Qian, X.; Wang, X.; Li, R.; Cheng, X.; Yang, Y.; Fu, J.; Zhang, S.; King, G.J.; Wu, J.; et al. construction of an integrated genetic linkage map for the A genome of Brassica napus using SSR markers derived from sequenced BACs in B. rapa. BMC Genom. 2010, 11, 594. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Qiao, C.L. Transcriptomic Analysis of Orange-red Petals and Genetic Research on the Anthocyanins Metabolic Pathway in Brassica napus L. Master’s Thesis, Southwest University, Chongqing, China, 2020. [Google Scholar]
- Zhang, W.; Ning, G.; Lv, H.; Liao, L.; Bao, M. Single MYB-type transcription factor AtCAPRICE: A new efficient tool to engineer the production of anthocyanin in tobacco. Biochem. Biophys. Res. Comm 2009, 388, 742–747. [Google Scholar] [CrossRef] [PubMed]
- Neff, M.M.; Chory, J. Genetic interactions between phytochrome, A.; phytochrome, B.; and cryptochrome 1 during Arabidopsis development. Plant Physiol. 1998, 118, 27–35. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, Q.; Liu, X.; Liu, H.; Fu, Y.; Cheng, Y.; Zhang, L.; Shi, W.; Zhang, Y.; Chen, J. Transcriptomic and Metabolomic Analysis of Wheat Kernels in Response to the Feeding of Orange Wheat Blossom Midges (Sitodiplosis mosellana) in the Field. Agric. Food Chem. 2022, 70, 1477–1493. [Google Scholar] [CrossRef]
- Chen, W.C.; Zhang, J.Q.; Zheng, S. Metabolite profiling and transcriptome analyses reveal novel regulatory mechanisms of melatonin biosynthesis in hickory. Hortic. Res. 2021, 8, 196. [Google Scholar] [CrossRef]
- Zhao, H. A Comparative Analysis of the Differences in the Regulation Mechanism of the Color of Chinese Cabbage and Brassica napus; Southwest University: Chongqing, China, 2019. [Google Scholar]
- Sharma, N.; Rahman, M.H.; Strelkov, S.; Thiagarajah, M.; Bansal, V.K.; Kav, N.N. Proteome-level changes in two Brassica napus lines exhibiting differential responses to the fungal pathogen Alternaria brassicae. Plant Sci. 2007, 172, 95–110. [Google Scholar] [CrossRef]
- Liu, Y.J.; Ye, S.H.; Yuan, G.G.; Ma, X.; Heng, S.; Yi, B.; Ma, C.; Shen, J.; Tu, J.; Fu, T.; et al. Gene silencing of BnaA09.ZEP and BnaC09.ZEP confers orange color in Brassica napus flowers. Plant J. 2020, 104, 932–949. [Google Scholar] [CrossRef]
- Ye, S.H.; Hua, S.J.; Ma, T.T.; Ma, X.; Chen, Y.; Wu, L.; Zhao, L.; Yi, B.; Ma, C.; Tu, J.; et al. Genetic and multi-omics analyses reveal BnaA07.PAP2In-184-317 as the key gene conferring anthocyanin-based color in Brassica napus flowers. J. Exp. Bot. 2022, 73, 6630–6645. [Google Scholar] [CrossRef]
- Vogt, T. Phenylpropanoid biosynthesis. Mol. Plant. 2010, 3, 2–20. [Google Scholar] [CrossRef] [Green Version]
- Petroni, K.; Tonelli, C. Recent advances on the regulation of anthocyanin synthesis in reproductive organs. Plant Sci. 2011, 181, 219–229. [Google Scholar] [CrossRef]
- Nishihara, M.; Higuchi, A.; Watanabe, A.; Tasaki, K. Application of the CRISPR/Cas9 system for modification of flower color in Torenia fournieri. BMC Plant Biol. 2018, 18, 331–339. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fu, W.Q.; Chen, D.Z.; Pan, Q.; Li, F.F.; Zhao, Z.G.; Ge, X.H.; Li, Z.Y. Production of red-flowered oilseed rape via the ectopic expression of Orychophragmus violaceus OvPAP2. Plant Biotechnol. 2018, 16, 367–380. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ryoji, T.; Fan, Y.; Di, S.K.; Murai, Y.; Iwashina, T.; Anai, T. Genetic and Chemical Analysis of Deep Purple Flower in Soybean. Crop Sci. 2017, 57, 1893–1898. [Google Scholar]
- Liu, D.; Wei, X.; Sun, D.; Yang, S.; Su, H.; Wang, Z.; Zhao, Y.; Li, L.; Liang, J.; Yang, L.; et al. An SNP Mutation of Gene RsPP Converts Petal Color from Purple to White in Radish (Raphanus sativus L.). Front. Plant Sci. 2021, 12, 643579. [Google Scholar] [CrossRef]
- Silva, S.; Costa, E.-M.; Calhau, C.; Morais, R.-M.; Pintado, M.-E. Anthocyanin extraction from plant tissues: A review. Crit. Rev. Food Sci. Nutr. 2017, 57, 3072–3083. [Google Scholar] [CrossRef]
- Gómez, J.D.; Vital, C.E.; Oliveira, M.G.A.; Ramos, H.J.O. Broad range flavonoid profiling by LC/MS of soybean genotypes contrasting for resistance to Anticarsia gemmatalis (Lepidoptera: Noctuidae). PLoS ONE 2018, 13, e0205010. [Google Scholar] [CrossRef]
- Du, Z.; Sha, X. Demethoxycurcumin inhibited human epithelia ovarian cancer cells’ growth via up-regulating miR-551a. Tumor Boil. 2017, 39, 1010428317694302. [Google Scholar] [CrossRef] [Green Version]
- Mol, J.; Grotewold, E.; Koes, R. How genes paint flowers and seeds. Trends Plant Sci. 1998, 3, 212–217. [Google Scholar] [CrossRef]
- Iwashina, T. Contribution to flower colors of flavonoids including anthocyanins: A review. Nat. Prod. Commun. 2015, 10, 529–544. [Google Scholar] [CrossRef] [Green Version]
- Aida, R.; Yoshida, K.; Kondo, T.; Kishimoto, S.; Shibata, M. Copigmentation gives bluer flowers on transgenic torenia plants with the antisense dihydroflavonol-4-reductase gene. Plant Sci. 2020, 160, 49–56. [Google Scholar] [CrossRef]
- Fukui, Y.; Tanaka, Y.; Kusumi, T.; Iwashita, T.; Nomoto, K. A rationale for the shift in color towards blue in transgenic carnation flowers expressing the flavonoid 3′,5′-hydroxylase gene. Phytochemistry 2003, 63, 15–23. [Google Scholar] [CrossRef] [PubMed]
- Yoshida, K.; Mori, M.; Kondo, T. Blue flower color development by anthocyanins: From chemical structure to cell physiology. Nat. Prod. Rep. 2009, 26, 884–915. [Google Scholar] [CrossRef] [PubMed]
- Okitsu, N.; Matsui, K.; Horikawa, M.; Sugahara, K.; Tanaka, Y. Identification and characterization of novel Nemophila menziesii flavone glucosyltransferases that catalyze biosynthesis of flavone 7,4′-O-diglucoside, a key component of blue metalloanthocyanins. Plant Cell Physiol. 2018, 59, 2075–2085. [Google Scholar] [CrossRef]
- Fu, J.W. The Study of the Subject Aboat Butin on Acuting in Rats Which Movement Function the Hind Legs and NF-kb, Caspase-3 Factors Influence of Expression after Spinal Cord Injury. Master’s Thesis, Dalian Medical University, Dalian, China, 2012. [Google Scholar]
- Xia, C. Study on the Renal Protective Effect of Phloretin Nanoparticles on Diabetic Rats. Master’s Thesis, Zhejiang University, Hangzhou, China, 2021. [Google Scholar]
- Han, Y.; Vimolmangkang, S.; Soria-Guerra, R.E.; Rosales-Mendoza, S.; Zheng, D.; Lygin, A.V.; Korban, S.S. Ectopic Expression ofApple F3′H Genes Contributes to Anthocyanin Accumulation in the Arabidopsis tt7 Mutant Grown Under Nitrogen Stress. Plant Physiol. 2010, 153, 806–820. [Google Scholar] [CrossRef] [Green Version]
- Rao, M.J.; Xu, Y.; Tang, X.; Huang, Y.; Xu, Q. CsCYT75B1, a Citrus CYTOCHROME P450 Gene, Is Involved in Accumulation of Antioxidant Flavonoids and Induces Drought Tolerance in Transgenic Arabidopsis. Antioxid. Redox Signal. 2020, 9, 161. [Google Scholar] [CrossRef] [Green Version]
- Himi, E.; Taketa, S. Barley Ant17, encoding flavanone 3-hydroxylase (F3H), is a promising target locus for attaining anthocyanin/proanthocyanidin-free plants without pleiotropic reduction of grain dormancy. Genome 2015, 58, 43–53. [Google Scholar] [CrossRef] [PubMed]
- Jung, C.S.; Griffiths, H.M.; De Jong, D.M.; Cheng, S.; Bodis, M.; Kim, T.S.; De Jong, W.S. The potato developer (D) locus encodes an R2R3 MYB transcription factor that regulates expression of multiple anthocyanin structural genes in tuber skin. Theor. Appl. Genet. 2009, 120, 45–57. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, Y.; Shen, S.; Yin, N.; Liu, H.; Du, D.; Li, J. Comparative analysis of the orange versus yellow petal of rapeseed (Brassica napus) using UPLC-HESI-MS/MS and transcriptome analysis. Plant Breed. 2022, 141, 77–87. [Google Scholar] [CrossRef]
- Yang, G.B.; Li, L.J.; Wei, M.; Li, J.; Yang, F. SmMYB113 Is a Key Transcription Factor Responsible for Compositional Variation of Anthocyanin and Color Diversity Among Eggplant Peels. Front. Plant Sci. 2022, 13, 843996. [Google Scholar] [CrossRef]
- Hao, P.; Liu, H.; Lin, B.; Ren, Y.; Huang, L.; Jiang, L.; Hua, S. BnaA03.ANS Identified by Metabolomics and RNA-seq Partly Played Irreplaceable Role in Pigmentation of Red Rapeseed (Brassica napus) Petal. Front. Plant Sci. 2022, 13, 940765. [Google Scholar] [CrossRef]












Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Li, S.; Li, X.; Wang, X.; Chang, T.; Peng, Z.; Guan, C.; Guan, M. Flavonoid Synthesis-Related Genes Determine the Color of Flower Petals in Brassica napus L. Int. J. Mol. Sci. 2023, 24, 6472. https://doi.org/10.3390/ijms24076472
Li S, Li X, Wang X, Chang T, Peng Z, Guan C, Guan M. Flavonoid Synthesis-Related Genes Determine the Color of Flower Petals in Brassica napus L. International Journal of Molecular Sciences. 2023; 24(7):6472. https://doi.org/10.3390/ijms24076472
Chicago/Turabian StyleLi, Shijun, Xi Li, Xiaodan Wang, Tao Chang, Zechuan Peng, Chunyun Guan, and Mei Guan. 2023. "Flavonoid Synthesis-Related Genes Determine the Color of Flower Petals in Brassica napus L." International Journal of Molecular Sciences 24, no. 7: 6472. https://doi.org/10.3390/ijms24076472