
Acute Ozone-Induced Transcriptional Changes in Markers of Oxidative Stress and Glucocorticoid Signaling in the Rat Hippocampus and Hypothalamus Are Sex-Specific
 , ,
, ,
Abstract
:1. Introduction
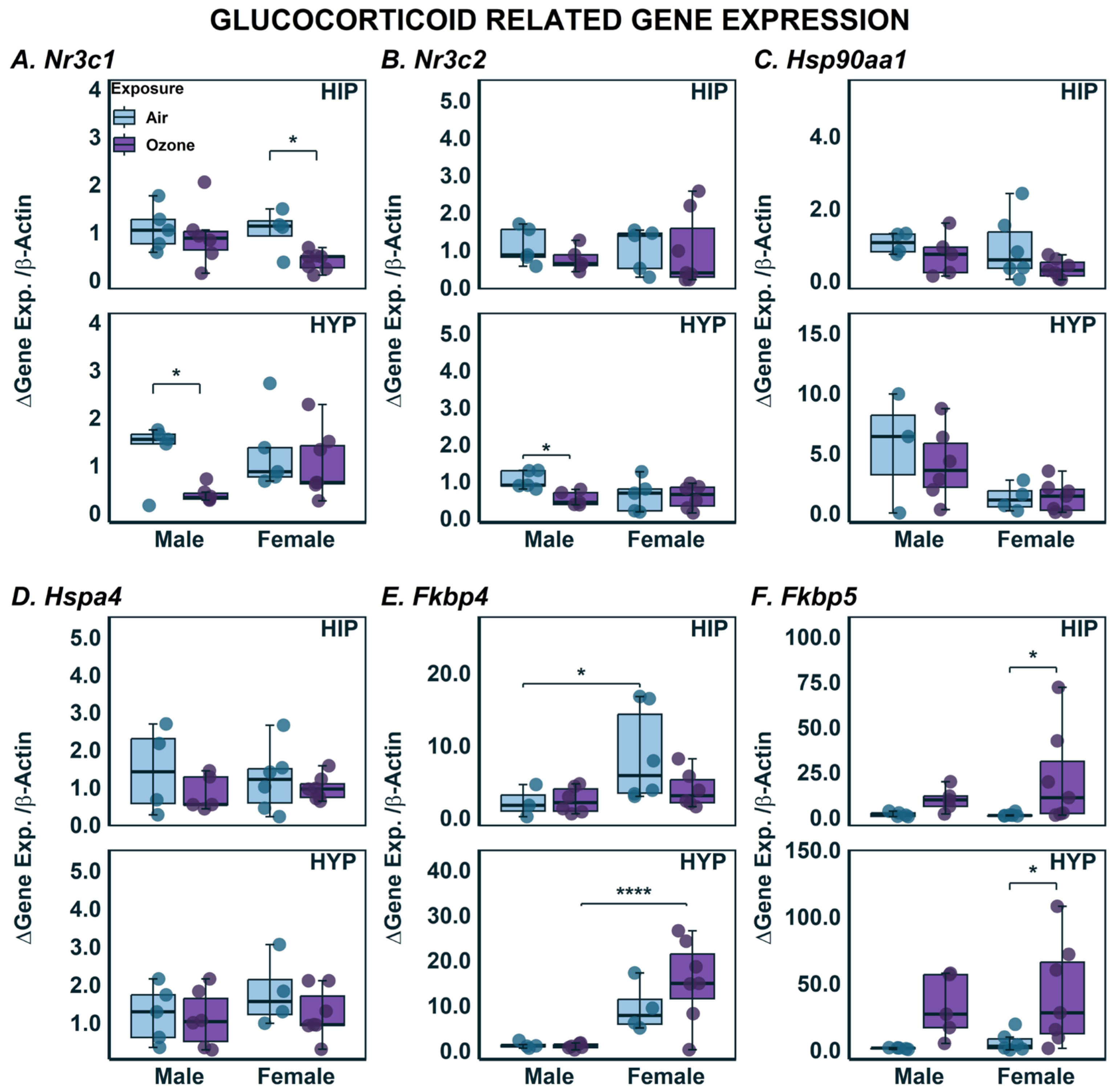
2. Results
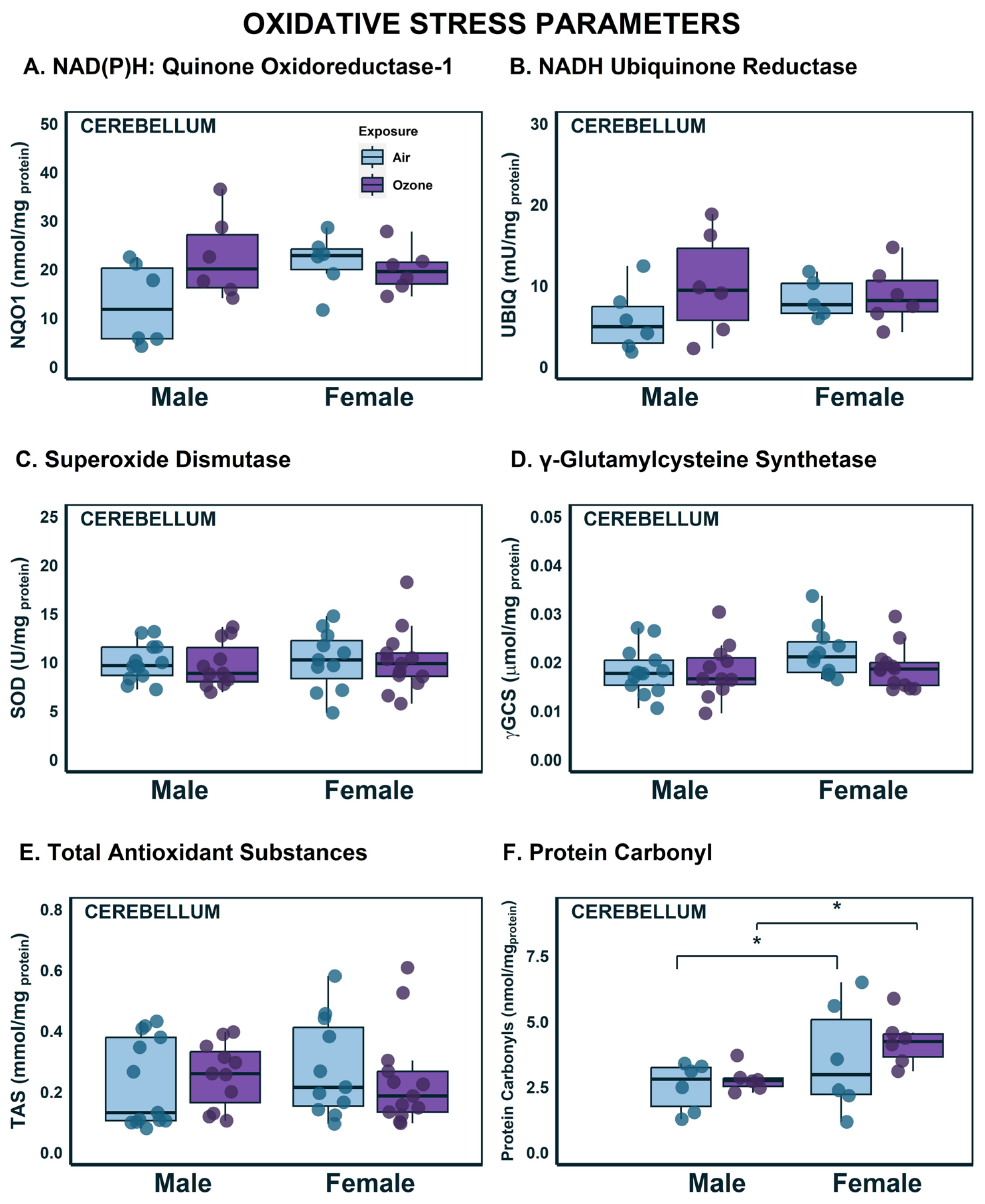
2.1. Effects on OS Parameters in the Cerebellum
2.2. Effects on Complex Enzyme Activities in the Cerebellum
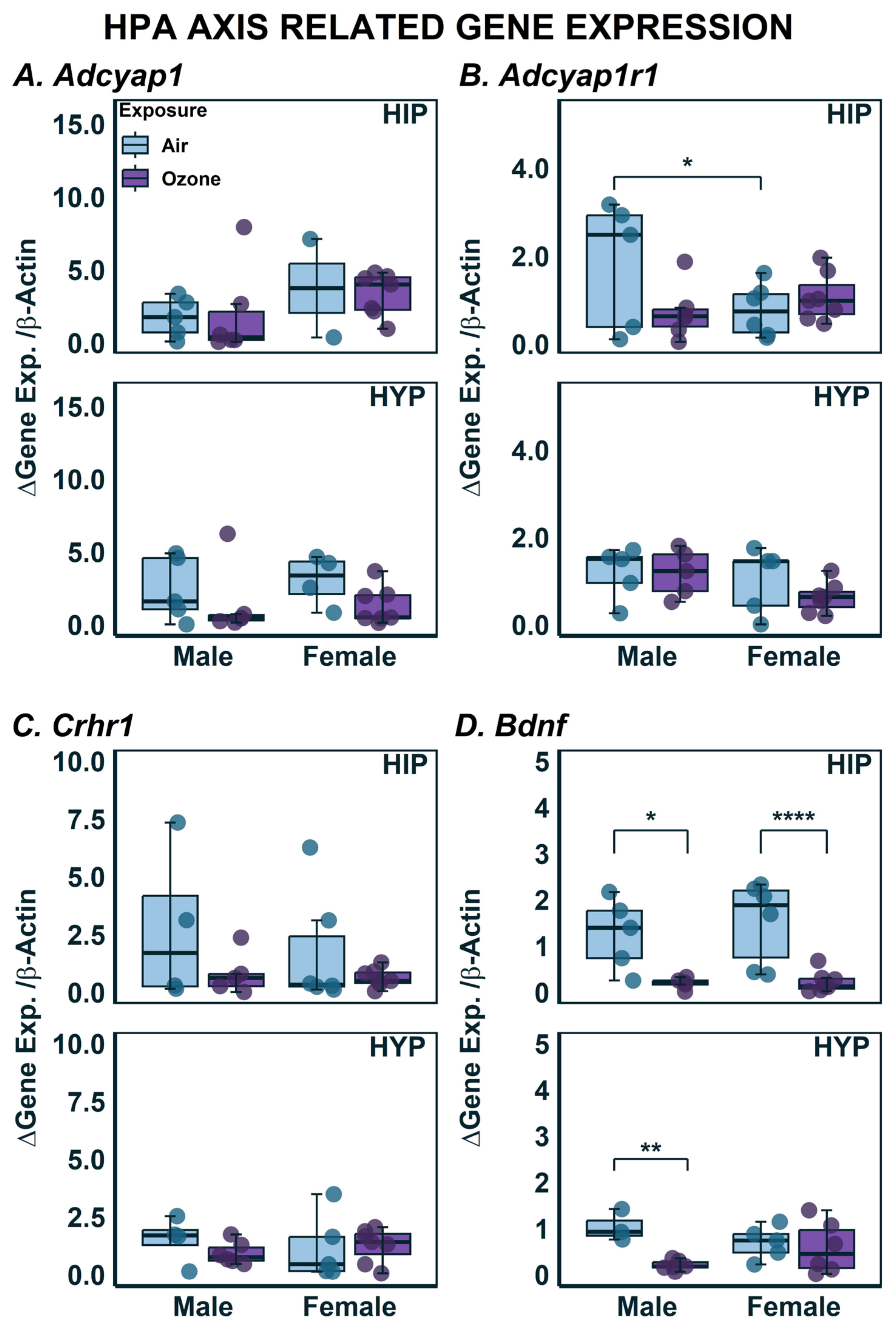
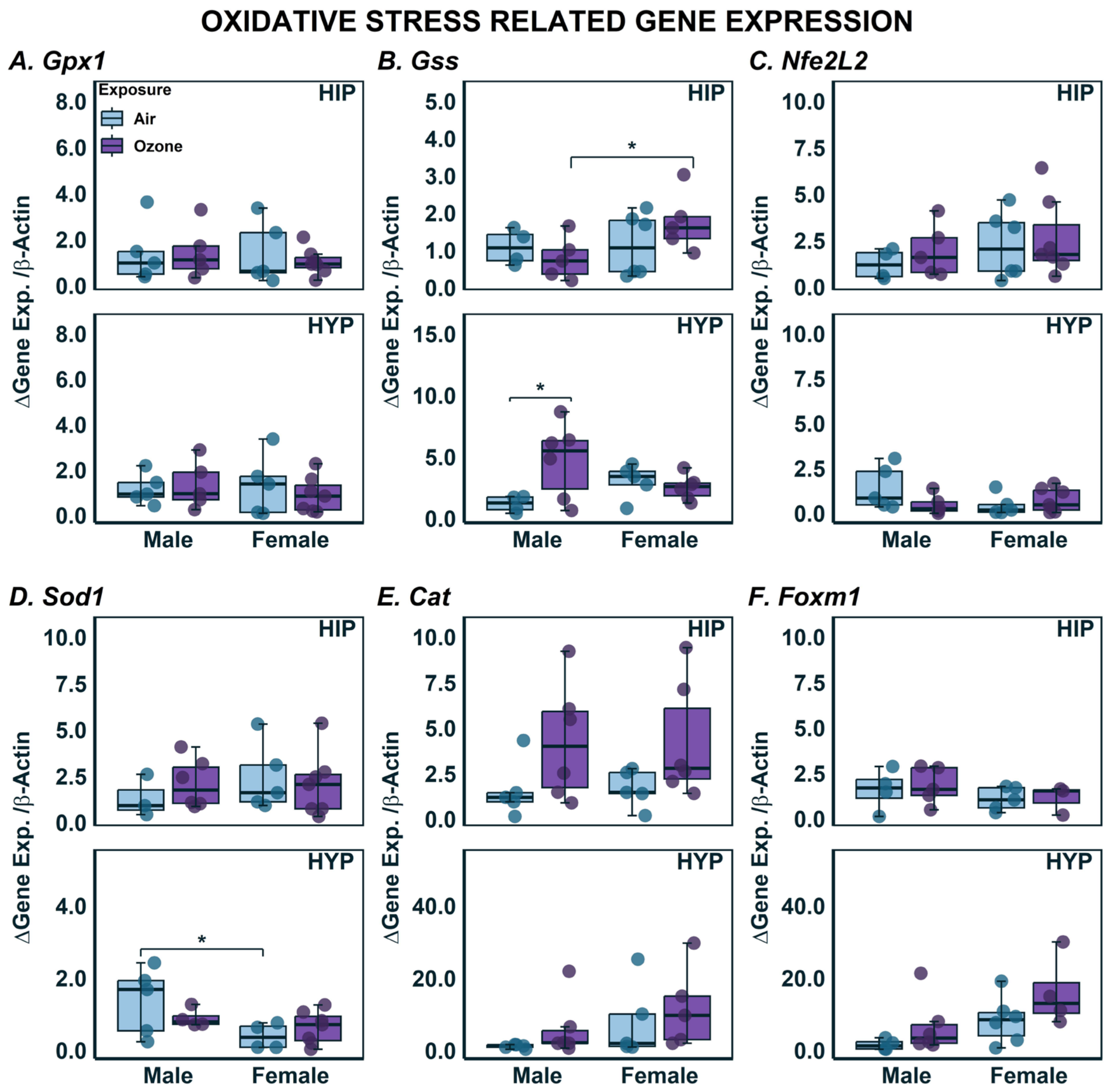
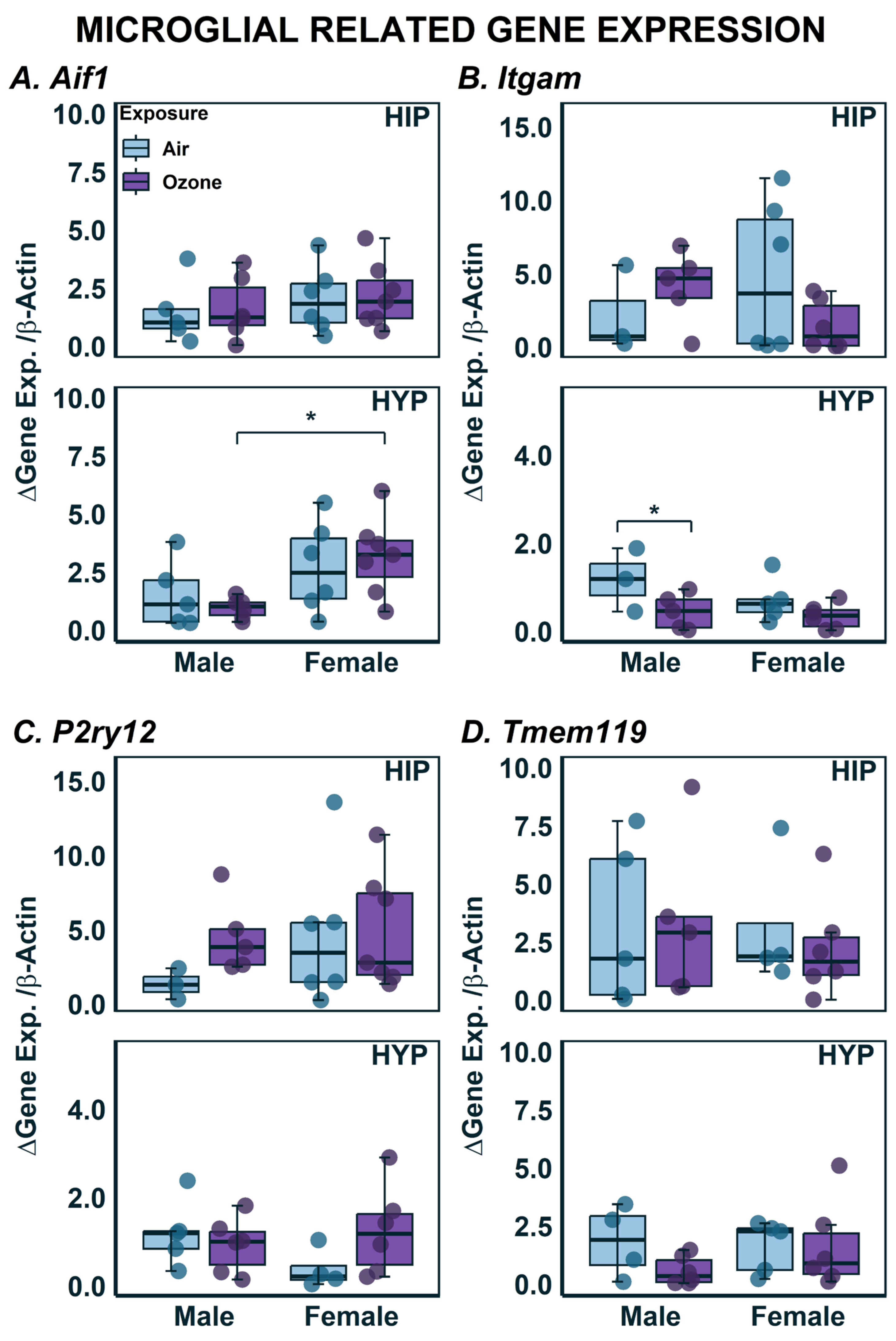
2.3. Effects on Gene Expression in HYP and HIP
3. Discussion
4. Materials and Methods
4.1. Animals
4.2. Ozone Generation & Exposure
4.3. Necropsy & Tissue Isolation
4.4. Tissue Preparation for OS Measures in Cerebellum
4.5. Markers of Cellular Damage
4.6. Assessment of Cerebellum Mitochondrial Complex I, II, and IV Enzyme Activities
4.7. Quantitative Polymerase Chain Reaction
4.8. Statistical Analysis
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Birukova, A.; Cyphert-Daly, J.; Cumming, R.I.; Yu, Y.R.; Gowdy, K.M.; Que, L.G.; Tighe, R.M. Sex Modifies Acute Ozone-Mediated Airway Physiologic Responses. Toxicol. Sci. 2019, 169, 499–510. [Google Scholar] [CrossRef]
- Klein, S.L.; Flanagan, K.L. Sex differences in immune responses. Nat. Rev. Immunol. 2016, 16, 626–638. [Google Scholar] [CrossRef] [PubMed]
- Shimamoto, A.; Rappeneau, V. Sex-dependent mental illnesses and mitochondria. Schizophr. Res. 2017, 187, 38–46. [Google Scholar] [CrossRef]
- Shin, H.H.; Maquiling, A.; Thomson, E.M.; Park, I.W.; Stieb, D.M.; Dehghani, P. Sex-difference in air pollution-related acute circulatory and respiratory mortality and hospitalization. Sci. Total Environ. 2022, 806, 150515. [Google Scholar] [CrossRef]
- Gordon, C.J.; Phillips, P.M.; Johnstone, A.F.; Beasley, T.E.; Ledbetter, A.D.; Schladweiler, M.C.; Snow, S.J.; Kodavanti, U.P. Effect of high-fructose and high-fat diets on pulmonary sensitivity, motor activity, and body composition of brown Norway rats exposed to ozone. Inhal. Toxicol. 2016, 28, 203–215. [Google Scholar] [CrossRef] [PubMed]
- Snow, S.J.; Broniowska, K.; Karoly, E.D.; Henriquez, A.R.; Phillips, P.M.; Ledbetter, A.D.; Schladweiler, M.C.; Miller, C.N.; Gordon, C.J.; Kodavanti, U.P. Offspring susceptibility to metabolic alterations due to maternal high-fat diet and the impact of inhaled ozone used as a stressor. Sci. Rep. 2020, 10, 16353. [Google Scholar] [CrossRef]
- Landrigan, P.J.; Fuller, R.; Acosta, N.J.R.; Adeyi, O.; Arnold, R.; Basu, N.; Baldé, A.B.; Bertollini, R.; Bose-O’Reilly, S.; Boufford, J.I.; et al. The Lancet Commission on pollution and health. Lancet 2018, 391, 462–512. [Google Scholar] [CrossRef] [Green Version]
- Gordon, C.J.; Phillips, P.M.; Beasley, T.E.; Ledbetter, A.; Aydin, C.; Snow, S.J.; Kodavanti, U.P.; Johnstone, A.F. Pulmonary sensitivity to ozone exposure in sedentary versus chronically trained, female rats. Inhal. Toxicol. 2016, 28, 293–302. [Google Scholar] [CrossRef]
- Koman, P.D.; Mancuso, P. Ozone Exposure, Cardiopulmonary Health, and Obesity: A Substantive Review. Chem. Res. Toxicol. 2017, 30, 1384–1395. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Manisalidis, I.; Stavropoulou, E.; Stavropoulos, A.; Bezirtzoglou, E. Environmental and Health Impacts of Air Pollution: A Review. Front. Public Health 2020, 8, 14. [Google Scholar] [CrossRef] [Green Version]
- Paz, C. Some consequences of ozone exposure on health. Arch. Med. Res. 1997, 28, 163–170. [Google Scholar]
- Pryor, W.A. How far does ozone penetrate into the pulmonary air/tissue boundary before it reacts? Free. Radic. Biol. Med. 1992, 12, 83–88. [Google Scholar] [CrossRef] [PubMed]
- Erickson, M.A.; Jude, J.; Zhao, H.; Rhea, E.M.; Salameh, T.S.; Jester, W.; Pu, S.; Harrowitz, J.; Nguyen, N.; Banks, W.A.; et al. Serum amyloid A: An ozone-induced circulating factor with potentially important functions in the lung-brain axis. FASEB J. 2017, 31, 3950–3965. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mumaw, C.L.; Levesque, S.; McGraw, C.; Robertson, S.; Lucas, S.; Stafflinger, J.E.; Campen, M.J.; Hall, P.; Norenberg, J.P.; Anderson, T.; et al. Microglial priming through the lung-brain axis: The role of air pollution-induced circulating factors. FASEB J. 2016, 30, 1880–1891. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gonzalez-Guevara, E.; Martinez-Lazcano, J.C.; Custodio, V.; Hernandez-Ceron, M.; Rubio, C.; Paz, C. Exposure to ozone induces a systemic inflammatory response: Possible source of the neurological alterations induced by this gas. Inhal. Toxicol. 2014, 26, 485–491. [Google Scholar] [CrossRef]
- Henriquez, A.; House, J.; Miller, D.B.; Snow, S.J.; Fisher, A.; Ren, H.; Schladweiler, M.C.; Ledbetter, A.D.; Wright, F.; Kodavanti, U.P. Adrenal-derived stress hormones modulate ozone-induced lung injury and inflammation. Toxicol. Appl. Pharmacol. 2017, 329, 249–258. [Google Scholar] [CrossRef]
- Henriquez, A.R.; Snow, S.J.; Schladweiler, M.C.; Miller, C.N.; Dye, J.A.; Ledbetter, A.D.; Richards, J.E.; Mauge-Lewis, K.; McGee, M.A.; Kodavanti, U.P. Adrenergic and glucocorticoid receptor antagonists reduce ozone-induced lung injury and inflammation. Toxicol. Appl. Pharmacol. 2018, 339, 161–171. [Google Scholar] [CrossRef]
- Kodavanti, U.P. Stretching the stress boundary: Linking air pollution health effects to a neurohormonal stress response. Biochim. Biophys. Acta 2016, 1860, 2880–2890. [Google Scholar] [CrossRef]
- Miller, D.B.; Snow, S.J.; Schladweiler, M.C.; Richards, J.E.; Ghio, A.J.; Ledbetter, A.D.; Kodavanti, U.P. Acute Ozone-Induced Pulmonary and Systemic Metabolic Effects Are Diminished in Adrenalectomized Rats. Toxicol. Sci. 2016, 150, 312–322. [Google Scholar] [CrossRef] [Green Version]
- Snow, S.J.; Henriquez, A.R.; Costa, D.L.; Kodavanti, U.P. Neuroendocrine Regulation of Air Pollution Health Effects: Emerging Insights. Toxicol. Sci. 2018, 164, 9–20. [Google Scholar] [CrossRef]
- Thomas, J.; Guenette, J.; Thomson, E.M. Stress axis variability is associated with differential ozone-induced lung inflammatory signaling and injury biomarker response. Environ. Res. 2018, 167, 751–758. [Google Scholar] [CrossRef]
- Thomson, E.M. Air Pollution, Stress, and Allostatic Load: Linking Systemic and Central Nervous System Impacts. J. Alzheimers Dis. 2019, 69, 597–614. [Google Scholar] [CrossRef] [Green Version]
- Thomson, E.M.; Pal, S.; Guenette, J.; Wade, M.G.; Atlas, E.; Holloway, A.C.; Williams, A.; Vincent, R. Ozone Inhalation Provokes Glucocorticoid-Dependent and -Independent Effects on Inflammatory and Metabolic Pathways. Toxicol. Sci. 2016, 152, 17–28. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Thomson, E.M.; Pilon, S.; Guenette, J.; Williams, A.; Holloway, A.C. Ozone modifies the metabolic and endocrine response to glucose: Reproduction of effects with the stress hormone corticosterone. Toxicol. Appl. Pharmacol. 2018, 342, 31–38. [Google Scholar] [CrossRef]
- Wigger, A.; Neumann, I.D. Periodic Maternal Deprivation Induces Gender-Dependent Alterations in Behavioral and Neuroendocrine Responses to Emotional Stress in Adult Rats. Physiol. Behav. 1999, 66, 293–302. [Google Scholar] [CrossRef]
- Viau, V. Functional cross-talk between the hypothalamic-pituitary-gonadal and -adrenal axes. J. Neuroendocrinol. 2002, 14, 506–513. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kajantie, E.; Phillips, D.I. The effects of sex and hormonal status on the physiological response to acute psychosocial stress. Psychoneuroendocrinology 2006, 31, 151–178. [Google Scholar] [CrossRef]
- Gordon, C.J.; Gottipolu, R.R.; Kenyon, E.M.; Thomas, R.; Schladweiler, M.C.; Mack, C.M.; Shannahan, J.H.; Wallenborn, J.G.; Nyska, A.; MacPhail, R.C.; et al. Aging and Susceptibility to Toluene in Rats: A Pharmacokinetic, Biomarker, and Physiological Approach. J. Toxicol. Environ. Health Part A 2009, 73, 301–318. [Google Scholar] [CrossRef]
- Gordon, C.J.; Jarema, K.A.; Lehmann, J.R.; Ledbetter, A.D.; Schladweiler, M.C.; Schmid, J.E.; Ward, W.O.; Kodavanti, U.P.; Nyska, A.; MacPhail, R.C. Susceptibility of adult and senescent Brown Norway rats to repeated ozone exposure: An assessment of behavior, serum biochemistry and cardiopulmonary function. Inhal. Toxicol. 2013, 25, 141–159. [Google Scholar] [CrossRef]
- Gordon, C.J.; Phillips, P.M.; Johnstone, A.F. Impact of genetic strain on body fat loss, food consumption, metabolism, ventilation, and motor activity in free running female rats. Physiol. Behav. 2016, 153, 56–63. [Google Scholar] [CrossRef] [PubMed]
- Gordon, C.J.; Phillips, P.M.; Johnstone, A.F.M.; Schmid, J.; Schladweiler, M.C.; Ledbetter, A.; Snow, S.J.; Kodavanti, U.P. Effects of maternal high-fat diet and sedentary lifestyle on susceptibility of adult offspring to ozone exposure in rats. Inhal. Toxicol. 2017, 29, 239–254. [Google Scholar] [CrossRef] [PubMed]
- Gordon, C.J.; Phillips, P.M.; Ledbetter, A.; Snow, S.J.; Schladweiler, M.C.; Johnstone, A.F.; Kodavanti, U.P. Active vs. sedentary lifestyle from weaning to adulthood and susceptibility to ozone in rats. Am. J. Physiol. Lung Cell. Mol. Physiol. 2017, 312, L100–L109. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kodavanti, P.R.S.; Valdez, M.; Richards, J.E.; Agina-Obu, D.I.; Phillips, P.M.; Jarema, K.A.; Kodavanti, U.P. Ozone-induced changes in oxidative stress parameters in brain regions of adult, middle-age, and senescent Brown Norway rats. Toxicol. Appl. Pharmacol. 2021, 410, 115351. [Google Scholar] [CrossRef]
- Valdez, M.; Valdez, J.M.; Freeborn, D.; Johnstone, A.F.M.; Kodavanti, P.R.S. The effects of ozone exposure and sedentary lifestyle on neuronal microglia and mitochondrial bioenergetics of female Long-Evans rats. Toxicol. Appl. Pharmacol. 2020, 408, 115254. [Google Scholar] [CrossRef] [PubMed]
- Henriquez, A.R.; House, J.S.; Snow, S.J.; Miller, C.N.; Schladweiler, M.C.; Fisher, A.; Ren, H.; Valdez, M.; Kodavanti, P.R.; Kodavanti, U.P. Ozone-induced dysregulation of neuroendocrine axes requires adrenal-derived stress hormones. Toxicol. Sci. 2019, 172, 38–50. [Google Scholar] [CrossRef] [PubMed]
- Valdez, M.C.; Freeborn, D.; Valdez, J.M.; Johnstone, A.F.M.; Snow, S.J.; Tennant, A.H.; Kodavanti, U.P.; Kodavanti, P.R.S. Mitochondrial Bioenergetics in Brain Following Ozone Exposure in Rats Maintained on Coconut, Fish and Olive Oil-Rich Diets. Int. J. Mol. Sci. 2019, 20, 6303. [Google Scholar] [CrossRef] [Green Version]
- Valdez, J.M.; Johnstone, A.F.M.; Richards, J.E.; Schmid, J.E.; Royland, J.E.; Kodavanti, P.R.S. Interaction of Diet and Ozone Exposure on Oxidative Stress Parameters within Specific Brain Regions of Male Brown Norway Rats. Int. J. Mol. Sci. 2018, 20, 11. [Google Scholar] [CrossRef] [Green Version]
- Gackiere, F.; Saliba, L.; Baude, A.; Bosler, O.; Strube, C. Ozone inhalation activates stress-responsive regions of the CNS. J. Neurochem. 2011, 117, 961–972. [Google Scholar] [CrossRef]
- Rivas-Arancibia, S.; Zimbron, L.F.; Rodriguez-Martinez, E.; Maldonado, P.D.; Borgonio Perez, G.; Sepulveda-Parada, M. Oxidative stress-dependent changes in immune responses and cell death in the substantia nigra after ozone exposure in rat. Front. Aging Neurosci. 2015, 7, 65. [Google Scholar] [CrossRef]
- Gomez-Crisostomo, N.P.; Rodriguez Martinez, E.; Rivas-Arancibia, S. Oxidative stress activates the transcription factors FoxO 1a and FoxO 3a in the hippocampus of rats exposed to low doses of ozone. Oxid. Med. Cell. Longev. 2014, 2014, 805764. [Google Scholar] [CrossRef] [Green Version]
- Kadiiska, M.B.; Basu, S.; Brot, N.; Cooper, C.; Csallany, A.S.; Davies, M.J.; George, M.M.; Murray, D.M.; Roberts, L.J., 2nd; Shigenaga, M.K.; et al. Biomarkers of oxidative stress study V: Ozone exposure of rats and its effect on lipids, proteins, and DNA in plasma and urine. Free Radic. Biol. Med. 2013, 61, 408–415. [Google Scholar] [CrossRef] [Green Version]
- Schieber, M.; Chandel, N.S. ROS function in redox signaling and oxidative stress. Curr. Biol. 2014, 24, R453–R462. [Google Scholar] [CrossRef] [Green Version]
- Kodavanti, P.R.S. Reactive Oxygen Species and Antioxidants in Neurotoxicology. In Target Organ Toxicity Series (Neurotoxicology); Tilson, H.A., Harry, G.J., Eds.; Taylor and Francis: Abingdon, UK, 1999; pp. 157–178. [Google Scholar]
- Mercurio, D.; Fumagalli, S.; Schafer, M.K.; Pedragosa, J.; Ngassam, L.D.C.; Wilhelmi, V.; Winterberg, S.; Planas, A.M.; Weihe, E.; De Simoni, M.G. Protein Expression of the Microglial Marker Tmem119 Decreases in Association with Morphological Changes and Location in a Mouse Model of Traumatic Brain Injury. Front. Cell. Neurosci. 2022, 16, 820127. [Google Scholar] [CrossRef]
- Henriquez, A.R.; Snow, S.J.; Dye, J.A.; Schladweiler, M.C.; Alewel, D.I.; Miller, C.N.; Kodavanti, U.P. The contribution of the neuroendocrine system to adaption after repeated daily ozone exposure in rats. Toxicol. Appl. Pharmacol. 2022, 447, 116085. [Google Scholar] [CrossRef]
- Kodavanti, U.P. Susceptibility Variations in Air Pollution Health Effects: Incorporating Neuroendocrine Activation. Toxicol. Pathol. 2019, 47, 962–975. [Google Scholar] [CrossRef]
- Greve, H.J.; Dunbar, A.L.; Lombo, C.G.; Ahmed, C.; Thang, M.; Messenger, E.J.; Mumaw, C.L.; Johnson, J.A.; Kodavanti, U.P.; Oblak, A.L.; et al. The bidirectional lung brain-axis of amyloid-beta pathology: Ozone dysregulates the peri-plaque microenvironment. Brain 2022, 146, 991–1005. [Google Scholar] [CrossRef]
- Rivas-Arancibia, S.; Guevara-Guzman, R.; Lopez-Vidal, Y.; Rodriguez-Martinez, E.; Zanardo-Gomes, M.; Angoa-Perez, M.; Raisman-Vozari, R. Oxidative stress caused by ozone exposure induces loss of brain repair in the hippocampus of adult rats. Toxicol. Sci. 2010, 113, 187–197. [Google Scholar] [CrossRef] [Green Version]
- Tyler, C.R.; Noor, S.; Young, T.L.; Rivero, V.; Sanchez, B.; Lucas, S.; Caldwell, K.K.; Milligan, E.D.; Campen, M.J. Aging Exacerbates Neuroinflammatory Outcomes Induced by Acute Ozone Exposure. Toxicol. Sci. 2018, 163, 123–139. [Google Scholar] [CrossRef] [Green Version]
- Nery-Flores, S.D.; Ramirez-Herrera, M.A.; Mendoza-Magana, M.L.; Romero-Prado, M.M.J.; Ramirez-Vazquez, J.J.; Banuelos-Pineda, J.; Espinoza-Gutierrez, H.A.; Ramirez-Mendoza, A.A.; Tostado, M.C. Dietary Curcumin Prevented Astrocytosis, Microgliosis, and Apoptosis Caused by Acute and Chronic Exposure to Ozone. Molecules 2019, 24, 2839. [Google Scholar] [CrossRef] [Green Version]
- Henriquez, A.R.; Snow, S.J.; Jackson, T.W.; House, J.S.; Motsinger-Reif, A.A.; Ward-Caviness, C.K.; Schladweiler, M.C.; Alewel, D.I.; Miller, C.N.; Farraj, A.K.; et al. Stress Drivers of Glucose Dynamics during Ozone Exposure Measured Using Radiotelemetry in Rats. Environ. Health Perspect. 2022, 130, 127006. [Google Scholar] [CrossRef]
- Herman, J.P. The neuroendocrinology of stress: Glucocorticoid signaling mechanisms. Psychoneuroendocrinology 2022, 137, 105641. [Google Scholar] [CrossRef] [PubMed]
- Moisan, M.P. Sexual Dimorphism in Glucocorticoid Stress Response. Int. J. Mol. Sci. 2021, 22, 3139. [Google Scholar] [CrossRef] [PubMed]
- Sze, Y.; Brunton, P.J. Sex, stress and steroids. Eur. J. Neurosci. 2020, 52, 2487–2515. [Google Scholar] [CrossRef] [PubMed]
- Akhter, H.; Ballinger, C.; Liu, N.; van Groen, T.; Postlethwait, E.M.; Liu, R.M. Cyclic Ozone Exposure Induces Gender-Dependent Neuropathology and Memory Decline in an Animal Model of Alzheimer’s Disease. Toxicol. Sci. 2015, 147, 222–234. [Google Scholar] [CrossRef] [Green Version]
- Dorado-Martínez, C.; Paredes-carbajal, C.; Mascher, D.; Borgonio-Pérez, G.; Rivas-arancibia, S. Effects of Different Ozone Doses on Memory, Motor Activity and Lipid Peroxidation Levels, in Rats. Int. J. Neurosci. 2001, 108, 149–161. [Google Scholar] [CrossRef]
- Aoyama, K. Glutathione in the Brain. Int. J. Mol. Sci. 2021, 22, 5010. [Google Scholar] [CrossRef]
- Zheleznyakova, G.Y.; Cao, H.; Schioth, H.B. BDNF DNA methylation changes as a biomarker of psychiatric disorders: Literature review and open access database analysis. Behav. Brain Funct. 2016, 12, 17. [Google Scholar] [CrossRef] [Green Version]
- Notaras, M.; van den Buuse, M. Neurobiology of BDNF in fear memory, sensitivity to stress, and stress-related disorders. Mol. Psychiatry 2020, 25, 2251–2274. [Google Scholar] [CrossRef]
- Linz, R.; Puhlmann, L.M.C.; Apostolakou, F.; Mantzou, E.; Papassotiriou, I.; Chrousos, G.P.; Engert, V.; Singer, T. Acute psychosocial stress increases serum BDNF levels: An antagonistic relation to cortisol but no group differences after mental training. Neuropsychopharmacology 2019, 44, 1797–1804. [Google Scholar] [CrossRef]
- Zhu, L.; Wang, Y.; Lin, X.; Zhao, X.; Fu, Z.J. Effects of Ozone on Hippocampus BDNF and Fos Expressions in Rats with Chronic Compression of Dorsal Root Ganglia. Biomed. Res. Int. 2021, 2021, 5572915. [Google Scholar] [CrossRef]
- Rose, M.; Filiatreault, A.; Guenette, J.; Williams, A.; Thomson, E.M. Ozone increases plasma kynurenine-tryptophan ratio and impacts hippocampal serotonin receptor and neurotrophic factor expression: Role of stress hormones. Environ. Res. 2020, 185, 109483. [Google Scholar] [CrossRef]
- Woodburn, S.C.; Bollinger, J.L.; Wohleb, E.S. The semantics of microglia activation: Neuroinflammation, homeostasis, and stress. J. Neuroinflamm. 2021, 18, 258. [Google Scholar] [CrossRef]
- Torres-Rodriguez, O.; Ortiz-Nazario, E.; Rivera-Escobales, Y.; Velazquez, B.; Colon, M.; Porter, J.T. Sex-dependent effects of microglial reduction on impaired fear extinction induced by single prolonged stress. Front. Behav. Neurosci. 2022, 16, 1014767. [Google Scholar] [CrossRef]
- Xu, Y.; Zhang, J.; Fang, Z.; Zhang, H. The effects of social defeat stress on hippocampal glial cells and interleukin-6 in adolescence and adulthood. NeuroReport 2022, 33, 828–834. [Google Scholar] [CrossRef] [PubMed]
- Hou, L.; Qu, X.; Qiu, X.; Huang, R.; Zhao, X.; Wang, Q. Integrin CD11b mediates locus coeruleus noradrenergic neurodegeneration in a mouse Parkinson’s disease model. J. Neuroinflamm. 2020, 17, 148. [Google Scholar] [CrossRef]
- Koorneef, L.L.; Viho, E.M.G.; Wahl, L.F.; Meijer, O.C. Do Corticosteroid Receptor mRNA Levels Predict the Expression of Their Target Genes? J. Endocr. Soc. 2022, 7, bvac188. [Google Scholar] [CrossRef]
- Gray, J.D.; Kogan, J.F.; Marrocco, J.; McEwen, B.S. Genomic and epigenomic mechanisms of glucocorticoids in the brain. Nat. Rev. Endocrinol. 2017, 13, 661–673. [Google Scholar] [CrossRef]
- Elakovic, I.; Djordjevic, A.; Adzic, M.; Djordjevic, J.; Radojcic, M.; Matic, G. Gender-specific response of brain corticosteroid receptors to stress and fluoxetine. Brain Res. 2011, 1384, 61–68. [Google Scholar] [CrossRef]
- Plieger, T.; Felten, A.; Splittgerber, H.; Duke, E.; Reuter, M. The role of genetic variation in the glucocorticoid receptor (NR3C1) and mineralocorticoid receptor (NR3C2) in the association between cortisol response and cognition under acute stress. Psychoneuroendocrinology 2018, 87, 173–180. [Google Scholar] [CrossRef]
- Ayyar, V.S.; DuBois, D.C.; Nakamura, T.; Almon, R.R.; Jusko, W.J. Modeling Corticosteroid Pharmacokinetics and Pharmacodynamics, Part II: Sex Differences in Methylprednisolone Pharmacokinetics and Corticosterone Suppression. J. Pharmacol. Exp. Ther. 2019, 370, 327–336. [Google Scholar] [CrossRef]
- Hausl, A.S.; Brix, L.M.; Hartmann, J.; Pohlmann, M.L.; Lopez, J.P.; Menegaz, D.; Brivio, E.; Engelhardt, C.; Roeh, S.; Bajaj, T.; et al. The co-chaperone Fkbp5 shapes the acute stress response in the paraventricular nucleus of the hypothalamus of male mice. Mol. Psychiatry 2021, 26, 3060–3076. [Google Scholar] [CrossRef] [PubMed]
- Mendonca, M.S.; Mangiavacchi, P.M.; Rios, A.F.L. Regulatory functions of FKBP5 intronic regions associated with psychiatric disorders. J. Psychiatr. Res. 2021, 143, 1–8. [Google Scholar] [CrossRef] [PubMed]
- Davies, T.H.; Ning, Y.M.; Sanchez, E.R. A new first step in activation of steroid receptors: Hormone-induced switching of FKBP51 and FKBP52 immunophilins. J. Biol. Chem. 2002, 277, 4597–4600. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Habara, M.; Sato, Y.; Goshima, T.; Sakurai, M.; Imai, H.; Shimizu, H.; Katayama, Y.; Hanaki, S.; Masaki, T.; Morimoto, M.; et al. FKBP52 and FKBP51 differentially regulate the stability of estrogen receptor in breast cancer. Proc. Natl. Acad. Sci. USA 2022, 119, e2110256119. [Google Scholar] [CrossRef] [PubMed]
- Zundel, C.G.; Ryan, P.; Brokamp, C.; Heeter, A.; Huang, Y.; Strawn, J.R.; Marusak, H.A. Air pollution, depressive and anxiety disorders, and brain effects: A systematic review. Neurotoxicology 2022, 93, 272–300. [Google Scholar] [CrossRef] [PubMed]
- Calderon-Garciduenas, L.; Mora-Tiscareno, A.; Chung, C.J.; Valencia, G.; Fordham, L.A.; Garcia, R.; Osnaya, N.; Romero, L.; Acuna, H.; Villarreal-Calderon, A.; et al. Exposure to air pollution is associated with lung hyperinflation in healthy children and adolescents in Southwest Mexico City: A pilot study. Inhal. Toxicol. 2000, 12, 537–561. [Google Scholar] [CrossRef]
- Xu, J.; Zhang, Y.; Fu, J.S.; Zheng, S.; Wang, W. Process analysis of typical summertime ozone episodes over the Beijing area. Sci. Total Environ. 2008, 399, 147–157. [Google Scholar] [CrossRef]
- Perdigones, B.C.; Lee, S.; Cohen, R.C.; Park, J.H.; Min, K.E. Two Decades of Changes in Summertime Ozone Production in California’s South Coast Air Basin. Environ. Sci. Technol. 2022, 56, 10586–10595. [Google Scholar] [CrossRef]
- Devlin, R.B.; Duncan, K.E.; Jardim, M.; Schmitt, M.T.; Rappold, A.G.; Diaz-Sanchez, D. Controlled exposure of healthy young volunteers to ozone causes cardiovascular effects. Circulation 2012, 126, 104–111. [Google Scholar] [CrossRef]
- Hatch, G.E.; Slade, R.; Harris, L.P.; McDonnell, W.F.; Devlin, R.B.; Koren, H.S.; Costa, D.L.; McKee, J. Ozone dose and effect in humans and rats. A comparison using oxygen-18 labeling and bronchoalveolar lavage. Am. J. Respir. Crit. Care Med. 1994, 150, 676–683. [Google Scholar] [CrossRef]
- Glowinski, J.; Iversen, L.L. Regional Studies of Catecholamines in the Rat Brain-I. J. Neurochem. 1966, 13, 655–669. [Google Scholar] [CrossRef]
- Lind, C.; Cadenas, E.; Hochstein, P.; Ernster, L. [30] DT-diaphorase: Purification, properties, and function. In Methods in Enzymology; Elsevier: Amsterdam, The Netherlands, 1990; Volume 186, pp. 287–301. [Google Scholar]
- Cormier, A.; Morin, C.; Zini, R.; Tillement, J.-P.; Lagrue, G. In vitro effects of nicotine on mitochondrial respiration and superoxide anion generation. Brain Res. 2001, 900, 72–79. [Google Scholar] [CrossRef]
- Pfaffl, M.W. A new mathematical model for relative quantification in real-time RT-PCR. Nucleic Acids Res. 2001, 29, e45. [Google Scholar] [CrossRef]
- RStudio. RStudio: Integrated Development Environment for R. Available online: http://www.rstudio.com/ (accessed on 31 January 2023).
- Wickham, H.; François, R.; Henry, L.; Müller, K. dplyr: A Grammar of Data Manipulation. Available online: https://CRAN.R-project.org/package=dplyr (accessed on 31 January 2023).
- Lenth, R.; Singmann, H.; Love, J.; Buerkner, P.; Herve, M. emmeans: Estimated Marginal Means, Aka Least-Squares Means. Available online: https://github.com/rvlenth/emmeans (accessed on 31 January 2023).
- Wickham, H. ggplot2: Elegant Graphics for Data Analysis. Available online: https://ggplot2.tidyverse.org (accessed on 31 January 2023).
- Wilke, C.O. Cowplot: Streamlined Plot Theme and Plot Annotations for ‘ggplot2’. Available online: https://wilkelab.org/cowplot (accessed on 31 January 2023).







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Hippocampus | ||||||||
|---|---|---|---|---|---|---|---|---|
| Male v. Female | Air v. Ozone | |||||||
| # Males | # Females | Significance | Significance | |||||
| Gene | Air | Ozone | Air | Ozone | Air | Ozone | Male | Female |
| Glucocorticoid Genes | ||||||||
| Fkbp4 | 3 | 6 | 6 | 6 | 3.8 × 10−2 (↑) | - | - | - |
| Fkbp5 | 5 | 5 | 6 | 7 | - | - | - | 2.9 × 10−2 (↑) |
| Nr3c1 | 5 | 6 | 4 | 7 | - | - | - | 4.1 × 10−2 (↓) |
| Nr3c2 | 5 | 5 | 5 | 7 | - | - | - | - |
| Hsp90aa1 | 4 | 5 | 6 | 7 | - | - | - | - |
| Hspa4 | 4 | 5 | 6 | 7 | - | - | - | - |
| HPA Axis Genes | ||||||||
| Bdnf | 5 | 5 | 6 | 7 | - | - | 1.0 × 10−2 (↓) | 9.0 × 10−4 (↓) |
| Adcyap1 | 5 | 6 | 2 | 7 | - | - | - | - |
| Adcyap1r1 | 5 | 6 | 6 | 7 | - | - | 4.3 × 10−2 (↓) | - |
| Crhr1 | 4 | 5 | 6 | 7 | - | - | - | - |
| Microglial Genes | ||||||||
| Aif1 | 5 | 6 | 6 | 7 | - | - | - | - |
| Itgam | 3 | 5 | 6 | 6 | - | - | - | - |
| P2ry12 | 3 | 5 | 6 | 7 | - | - | - | - |
| Tmem119 | 5 | 5 | 5 | 6 | - | - | - | - |
| Oxidative Stress Genes | ||||||||
| Cat | 5 | 6 | 5 | 6 | - | - | - | - |
| Foxm1 | 4 | 5 | 5 | 3 | - | - | - | - |
| Gpx1 | 5 | 5 | 5 | 7 | - | - | - | - |
| Gss | 4 | 5 | 6 | 5 | - | 4.7 × 10−2 (↑) | - | - |
| Nfe2l2 | 4 | 5 | 6 | 7 | - | - | - | - |
| Sod1 | 3 | 6 | 5 | 7 | - | - | - | - |
| Hypothalamus | ||||||||
| Glucocorticoid Genes | ||||||||
| Fkbp4 | 4 | 6 | 4 | 7 | - | 4.0 × 10−4 (↑) | - | - |
| Fkbp5 | 5 | 5 | 6 | 7 | - | - | - | 1.8 × 10−2 (↑) |
| Nr3c1 | 5 | 6 | 5 | 7 | - | - | 2.8 × 10−2 (↓) | - |
| Nr3c2 | 5 | 5 | 5 | 6 | - | - | 2.4 × 10−2 (↓) | - |
| Hsp90aa1 | 3 | 6 | 4 | 7 | - | - | - | - |
| Hspa4 | 5 | 6 | 4 | 7 | - | - | - | - |
| HPA Axis Genes | ||||||||
| Bdnf | 3 | 6 | 5 | 6 | - | - | 7.4 × 10−3 (↓) | - |
| Adcyap1 | 5 | 6 | 4 | 7 | - | - | - | - |
| Adcyap1r1 | 5 | 5 | 5 | 7 | - | - | - | - |
| Crhr1 | 4 | 6 | 5 | 7 | - | - | - | - |
| Microglial Genes | ||||||||
| Aif1 | 5 | 6 | 6 | 7 | - | 1.5 × 10−2 (↑) | - | - |
| Itgam | 3 | 5 | 5 | 6 | - | - | 4.4 × 10−2 (↓) | - |
| P2ry12 | 5 | 6 | 4 | 6 | - | - | - | - |
| Tmem119 | 4 | 6 | 5 | 6 | - | - | - | - |
| Oxidative Stress Genes | ||||||||
| Cat | 5 | 6 | 5 | 5 | - | - | - | - |
| Foxm1 | 4 | 6 | 6 | 4 | - | - | - | - |
| Gpx1 | 5 | 5 | 5 | 7 | - | - | - | - |
| Gss | 4 | 6 | 5 | 6 | - | - | 1.0 × 10−2 (↑) | - |
| Nfe2l2 | 5 | 5 | 5 | 7 | - | - | - | - |
| Sod1 | 5 | 4 | 4 | 7 | 2.3 × 10−2 (↓) | - | - | - |
| Gene | Accession Number | Forward Primer Sequence | Reverse Primer Sequence | Product Length | Efficiency | Product Length |
|---|---|---|---|---|---|---|
| Actb | NM_031144.3 | GTGTGGATTGGTGGCTCTATC | AACGCAGCTCAGTAACAGTC | 137 | 96.184 | 137 |
| Adcyap1 | NM_016989.2 | GAAGAAGAGGCTTACGATCAGG | GTCCAAGACTTTGCGGTAGG | 177 | 97.664 | 177 |
| Adcyap1r1 | NM_001270579.1 | GGAAGTGAGGTCTTGCTCTATG | TCCTGACACTTGCTGCTTAC | 127 | 101.797 | 127 |
| Aif1 | NM_017196 | ATCGTCATCTCCCCACCTAA | GATCATCGAGGAAGTGCTTGT | 145 | 92.694 | 145 |
| Bdnf | NM_001270630.1 | GGTCGATTAGGTGGCTTCATAG | CGGAAACAGAACGAACAGAAAC | 160 | 98.044 | 160 |
| Cat | NM_012520 | TCCCAACTACTACCCCAACA | AAGTGACGTTGTCTTCATTAGCA | 121 | 103.621 | 121 |
| Crhr1 | NM_030999 | GGTATACACTGACTACATCTACCAG | CAGCCTTCCTGTACTGAATGG | 143 | 100.945 | 143 |
| Fkbp4 | NM_001191863.1 | TCATCAAGAGAGAGGGTACAGG | TGGTTGCCACAGCAATATCC | 183 | 103.394 | 183 |
| Fkbp5 | NM_001012174 | CACCAGTAACAATGAAGAAAACCC | CCTCACTAGTCCCCACTCTT | 116 | 108.288 | 116 |
| Foxm1 | NM_031633 | GGCTTGGAAAGATGAGTTCTGA | AACCTTAACCCGATTCTGCTC | 101 | 103.778 | 101 |
| Gpx1 | NM_030826.4 | ACATCAGGAGAATGGCAAGAATG | CATTCACCTCGCACTTCTCAAAC | 110 | 104.432 | 110 |
| Gss | NM_012962 | GACAGGAAGATCCATGTAATCCG | ATCTCGGAAGTAAACCACAGC | 120 | 101.884 | 120 |
| Hsp90aa1 | NM_175761.2 | AAACAGCACTCCTGTCTTCC | GCCTAGTCTACTTCTTCCATGC | 199 | 103.447 | 199 |
| Hspa4 | NM_153629.1 | ACCACCTCAAGCAAAGAAGG | CCGTTCCTTCTCCAGTTTATCC | 154 | 97.097 | 154 |
| Itgam | NM_012711 | GATGTTCAAGCAGAATTTCGGT | GTATTGCCATCAGCGTCCAT | 117 | 106.195 | 117 |
| Nfe2l2 | NM_031789 | CAGCATGATGGACTTGGAATTG | AGTTGCTCTTGTCTCTCCTTTTC | 189 | 108.400 | 189 |
| Nr3c1 | NM_012576.2 | CCTTTGTTCTAAGCTAGGGAAGG | GTGGATGAGGATGGTTAGAATGG | 127 | 96.072 | 127 |
| Nr3c2 | NM_013131.1 | GGCAAATCTCAACAACTCAAGG | TGAAGTGGCATAGCTGAAGG | 142 | 105.798 | 142 |
| P2ry12 | NM_022800 | TCCGAGTCAACAGAATAACCAG | GATCTTGTAGTCTCTGCTGCAC | 113 | 94.093 | 113 |
| Sod1 | NM_017050 | GACAATACACAAGGCTGTACCAC | TTGCCCAAGTCATCTTGTTTCTC | 230 | 109.360 | 230 |
| Tmem119 | NM_001107155 | CGAGACAGTTGGACCGAGAC | ACAAGTAGCAGCAGAGACAGGAG | 141 | 99.560 | 141 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Valdez, M.C.; Freeborn, D.L.; Vulimiri, P.; Valdez, J.M.; Kodavanti, U.P.; Kodavanti, P.R.S. Acute Ozone-Induced Transcriptional Changes in Markers of Oxidative Stress and Glucocorticoid Signaling in the Rat Hippocampus and Hypothalamus Are Sex-Specific. Int. J. Mol. Sci. 2023, 24, 6404. https://doi.org/10.3390/ijms24076404
Valdez MC, Freeborn DL, Vulimiri P, Valdez JM, Kodavanti UP, Kodavanti PRS. Acute Ozone-Induced Transcriptional Changes in Markers of Oxidative Stress and Glucocorticoid Signaling in the Rat Hippocampus and Hypothalamus Are Sex-Specific. International Journal of Molecular Sciences. 2023; 24(7):6404. https://doi.org/10.3390/ijms24076404
Chicago/Turabian StyleValdez, Matthew C., Danielle L. Freeborn, Padmaja Vulimiri, Joseph M. Valdez, Urmila P. Kodavanti, and Prasada Rao S. Kodavanti. 2023. "Acute Ozone-Induced Transcriptional Changes in Markers of Oxidative Stress and Glucocorticoid Signaling in the Rat Hippocampus and Hypothalamus Are Sex-Specific" International Journal of Molecular Sciences 24, no. 7: 6404. https://doi.org/10.3390/ijms24076404