
Crosstalk between SOX Genes and Long Non-Coding RNAs in Glioblastoma
 , ,
, ,
Abstract
:1. Introduction
2. The Role of SOX Genes in Glioblastoma

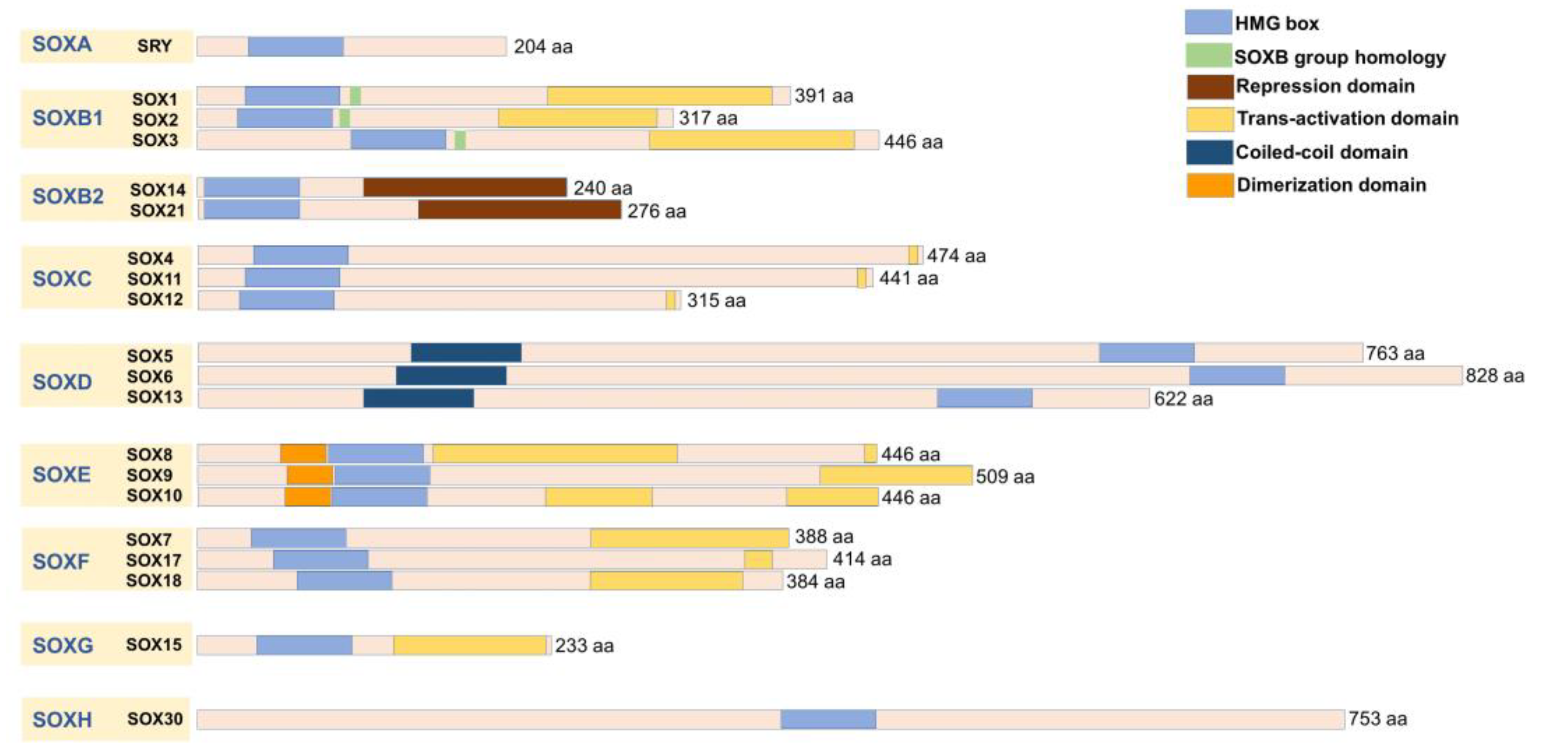{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| SOX Group | Members | Expression in GMB | Clinical Outcome | References |
|---|---|---|---|---|
| SOXA | SRY | Differentially expressed in GBM tissue samples | Downregulation was correlated with worse overall survival | [29] |
| SOXB1 | SOX1 | Overexpressed in GMB | Overexpression was correlated with poor prognosis | [31] |
| SOX2 | Overexpressed in GMB, a marker of undifferentiated proliferating cells of GBM | Overexpression was correlated with poor prognosis | [32,36] | |
| SOX3 | Elevated expression in GBM | Overexpression correlated with better overall survival; or with poor outcome; or no effect on survival | [38,40,41] | |
| SOXB2 | SOX14 | No data | No data | |
| SOX21 | Co-expressed with SOX2 in GBM, potential tumor suppressor | Elevated expression associated with low-risk score and better overall survival | [43] | |
| SOXC | SOX4 | Overexpressed in primary GBM tissues | High expression was correlated with a good prognosis | [46] |
| SOX11 | Overexpressed in malignant gliomas | High expression marked as a significant favorable prognostic indicator | [49] | |
| SOX12 | No data | No data | ||
| SOXD | SOX5 | Low expression in GBM, suggested to act as a tumor suppressor | One study corelated its overexpression with poor prognosis | [51] |
| SOX6 | Downregulated or overexpressed | High expression correlated with lower survival rate | [53] | |
| SOX13 | Downregulated in the majority of GBM samples | No data | [53] | |
| SOXE | SOX8 | Downregulated, could serve for predicting the differentiation status of glioma subtypes | No data | [53] |
| SOX9 | High expression in GBM | High expression was correlated with poor clinical outcome | [55] | |
| SOX10 | Downregulated in GBM | Low expression was correlated with shorter survival rate | [58] | |
| SOXF | SOX7 | Downregulated in GBM tissue samples and cell lines | No data | [59] |
| SOX17 | No data | No data | ||
| SOX18 | No data | No data | ||
| SOXG | SOX15 | Downregulated in GBM | Low expression was correlated with shorter survival | [62] |
| SOXH | SOX30 | No data | No data |
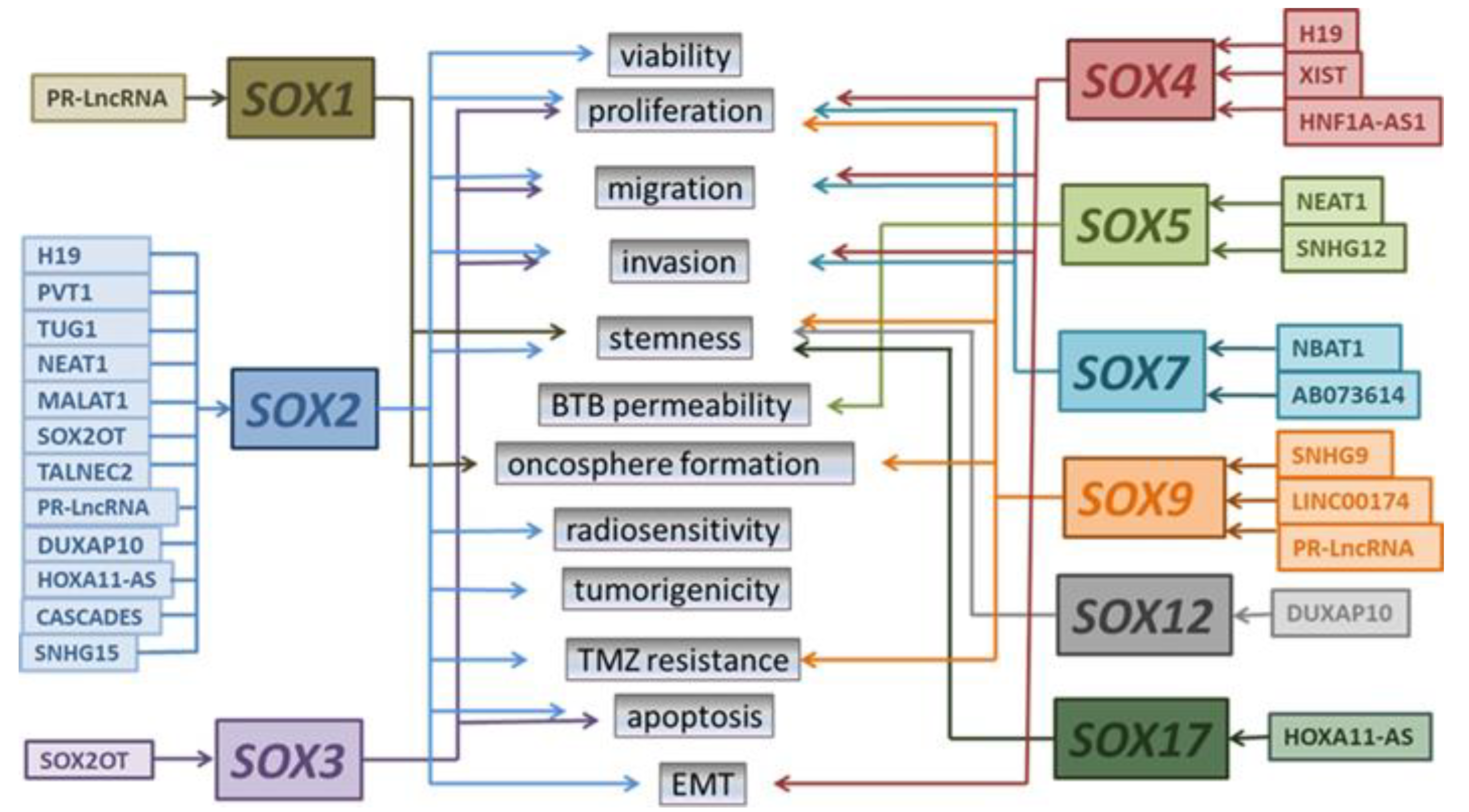
3. LncRNAs Acting through SOX Genes in Glioblastoma
3.1. LncRNAs Acting through SOX with Tumor Suppressor Roles in GBM
3.2. LncRNAs Acting through SOX with Oncogenic Roles in GBM
4. SOX Overlapping Transcripts and SOX Antisense RNA
5. Potential of lncRNAs for Prognosis and Therapy in GBM
5.1. LncRNAs as Prognostic Markers in GBM
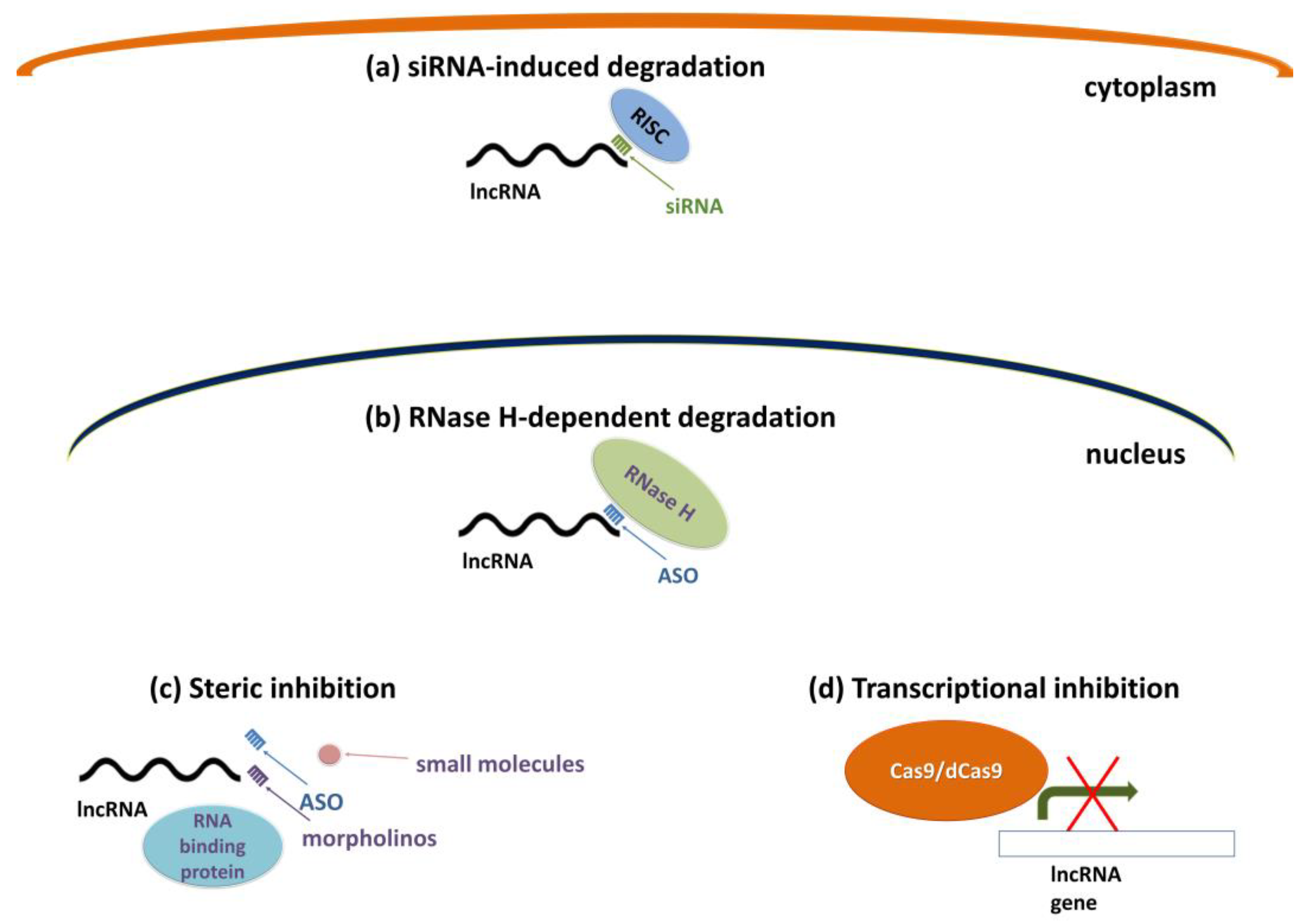
5.2. Approaches for Therapeutic Targeting of lncRNAs
6. Future Directions
7. Concluding Remarks
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Louis, D.N.; Ohgaki, H.; Wiestler, O.D.; Cavenee, W.K.; Burger, P.C.; Jouvet, A.; Scheithauer, B.W.; Kleihues, P. The 2007 WHO classification of tumours of the central nervous system. Acta Neuropathol. 2007, 114, 97–109. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Delgado-Lopez, P.D.; Corrales-Garcia, E.M. Survival in glioblastoma: A review on the impact of treatment modalities. Clin. Transl. Oncol. 2016, 18, 1062–1071. [Google Scholar] [CrossRef]
- Grimm, D.; Bauer, J.; Wise, P.; Krüger, M.; Simonsen, U.; Wehland, M.; Infanger, M.; Corydon, T.J. The role of SOX family members in solid tumours and metastasis. Semin. Cancer Biol. 2020, 67, 122–153. [Google Scholar] [CrossRef]
- Kim, S.-H.; Lim, K.-H.; Yang, S.; Joo, J.-Y. Long non-coding RNAs in brain tumors: Roles and potential as therapeutic targets. J. Hematol. Oncol. 2021, 14, 77. [Google Scholar] [CrossRef] [PubMed]
- Li, B.; Zhao, H.; Song, J.; Wang, F.; Chen, M. LINC00174 down-regulation decreases chemoresistance to temozolomide in human glioma cells by regulating miR-138-5p/SOX9 axis. Hum. Cell 2020, 33, 159–174. [Google Scholar] [CrossRef]
- Li, Z.; Jiang, P.; Li, J.; Peng, M.; Zhao, X.; Zhang, X.; Chen, K.; Zhang, Y.; Liu, H.; Gan, L.; et al. Tumor-derived exosomal lnc-Sox2ot promotes EMT and stemness by acting as a ceRNA in pancreatic ductal adenocarcinoma. Oncogene 2018, 37, 3822–3838. [Google Scholar] [CrossRef]
- Liu, B.; Zhou, J.; Wang, C.; Chi, Y.; Wei, Q.; Fu, Z.; Lian, C.; Huang, Q.; Liao, C.; Yang, Z.; et al. LncRNA SOX2OT promotes temozolomide resistance by elevating SOX2 expression via ALKBH5-mediated epigenetic regulation in glioblastoma. Cell Death Dis. 2020, 11, 384. [Google Scholar] [CrossRef] [PubMed]
- Schepers, G.E.; Teasdale, R.D.; Koopman, P. Twenty pairs of sox: Extent, homology, and nomenclature of the mouse and human sox transcription factor gene families. Dev. Cell 2002, 3, 167–170. [Google Scholar] [CrossRef] [Green Version]
- Pevny, L.H.; Lovell-Badge, R. Sox genes find their feet. Curr. Opin. Genet. Dev. 1997, 7, 338–344. [Google Scholar] [CrossRef]
- Bowles, J.; Schepers, G.; Koopman, P. Phylogeny of the SOX family of developmental transcription factors based on sequence and structural indicators. Dev. Biol. 2000, 227, 239–255. [Google Scholar] [CrossRef] [Green Version]
- Kamachi, Y.; Uchikawa, M.; Kondoh, H. Pairing SOX off: With partners in the regulation of embryonic development. Trends Genet. 2000, 16, 182–187. [Google Scholar] [CrossRef]
- Williams, C.A.C.; Soufi, A.; Pollard, S.M. Post-translational modification of SOX family proteins: Key biochemical targets in cancer? Semin. Cancer Biol. 2020, 67, 30–38. [Google Scholar] [CrossRef]
- Kamachi, Y.; Kondoh, H. Sox proteins: Regulators of cell fate specification and differentiation. Development 2013, 140, 4129–4144. [Google Scholar] [CrossRef] [Green Version]
- Hu, J.; Li, K.; Li, Z.; Gao, C.; Guo, F.; Wang, Y.; Xue, F. Sex-determining region Y box-containing genes: Regulators and biomarkers in gynecological cancers. Cancer Biol. Med. 2019, 16, 462–474. [Google Scholar] [CrossRef] [PubMed]
- She, Z.Y.; Yang, W.X. SOX family transcription factors involved in diverse cellular events during development. Eur. J. Cell Biol. 2015, 94, 547–563. [Google Scholar] [CrossRef]
- Miao, Q.; Hill, M.C.; Chen, F.; Mo, Q.; Ku, A.T.; Ramos, C.; Sock, E.; Lefebvre, V.; Nguyen, H. SOX11 and SOX4 drive the reactivation of an embryonic gene program during murine wound repair. Nat. Commun. 2019, 10, 4042. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sarkar, A.; Hochedlinger, K. The sox family of transcription factors: Versatile regulators of stem and progenitor cell fate. Cell Stem Cell 2013, 12, 15–30. [Google Scholar] [CrossRef] [Green Version]
- Lai, H.; Cheng, X.; Liu, Q.; Luo, W.; Liu, M.; Zhang, M.; Miao, J.; Ji, Z.; Lin, G.N.; Song, W.; et al. Single-cell RNA sequencing reveals the epithelial cell heterogeneity and invasive subpopulation in human bladder cancer. Int. J. Cancer 2021, 149, 2099–2115. [Google Scholar] [CrossRef]
- Thomas, E.; Thankan, R.S.; Purushottamachar, P.; Huang, W.; Kane, M.A.; Zhang, Y.; Ambulos, N.; Weber, D.J.; Njar, V.C.O. Transcriptome profiling reveals that VNPP433-3β, the lead next-generation galeterone analog inhibits prostate cancer stem cells by downregulating epithelial-mesenchymal transition and stem cell markers. Mol. Carcinog. 2022, 61, 643–654. [Google Scholar] [CrossRef] [PubMed]
- Gu, W.; Wang, B.; Wan, F.; Wu, J.; Lu, X.; Wang, H.; Zhu, Y.; Zhang, H.; Shi, G.; Dai, B.; et al. SOX2 and SOX12 are predictive of prognosis in patients with clear cell renal cell carcinoma. Oncol. Lett. 2018, 15, 4564–4570. [Google Scholar] [CrossRef]
- Li, M.M.; Tang, Y.Q.; Gong, Y.F.; Cheng, W.; Li, H.L.; Kong, F.E.; Zhu, W.J.; Liu, S.S.; Huang, L.; Guan, X.Y.; et al. Development of an oncogenic dedifferentiation SOX signature with prognostic significance in hepatocellular carcinoma. BMC Cancer 2019, 19, 851. [Google Scholar] [CrossRef] [Green Version]
- Seok, J.; Gil, M.; Dayem, A.A.; Saha, S.K.; Cho, S.G. Multi-Omics Analysis of SOX4, SOX11, and SOX12 Expression and the Associated Pathways in Human Cancers. J. Pers. Med. 2021, 11, 823. [Google Scholar] [CrossRef] [PubMed]
- Chen, H.; Chen, X.; Zeng, F.; Fu, A.; Huang, M. Prognostic value of SOX9 in cervical cancer: Bioinformatics and experimental approaches. Front. Genet. 2022, 13, 939328. [Google Scholar] [CrossRef]
- Chen, Y.C.; Sahoo, S.; Brien, R.; Jung, S.; Humphries, B.; Lee, W.; Cheng, Y.H.; Zhang, Z.; Luker, K.E.; Wicha, M.S.; et al. Single-cell RNA-sequencing of migratory breast cancer cells: Discovering genes associated with cancer metastasis. Analyst 2019, 144, 7296–7309. [Google Scholar] [CrossRef]
- Otálora-Otálora, B.A.; López-Kleine, L.; Rojas, A. Lung Cancer Gene Regulatory Network of Transcription Factors Related to the Hallmarks of Cancer. Curr. Issues Mol. Biol. 2023, 45, 434–464. [Google Scholar] [CrossRef]
- Ostrom, Q.T.; Gittleman, H.; Truitt, G.; Boscia, A.; Kruchko, C.; Barnholtz-Sloan, J.S. CBTRUS Statistical Report: Primary Brain and Other Central Nervous System Tumors Diagnosed in the United States in 2011-2015. Neuro Oncol. 2018, 20, iv1–iv86. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gittleman, H.; Ostrom, Q.T.; Stetson, L.C.; Waite, K.; Hodges, T.R.; Wright, C.H.; Wright, J.; Rubin, J.B.; Berens, M.E.; Lathia, J.; et al. Sex is an important prognostic factor for glioblastoma but not for nonglioblastoma. Neurooncol. Pract. 2019, 6, 451–462. [Google Scholar] [CrossRef]
- Yuan, Y.; Liu, L.; Chen, H.; Wang, Y.; Xu, Y.; Mao, H.; Li, J.; Mills, G.B.; Shu, Y.; Li, L.; et al. Comprehensive Characterization of Molecular Differences in Cancer between Male and Female Patients. Cancer Cell 2016, 29, 711–722. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Łysiak, M.; Smits, A.; Roodakker, K.R.; Sandberg, E.; Dimberg, A.; Mudaisi, M.; Bratthäll, C.; Strandeus, M.; Milos, P.; Hallbeck, M.; et al. Deletions on Chromosome Y and Downregulation of the SRY Gene in Tumor Tissue Are Associated with Worse Survival of Glioblastoma Patients. Cancers 2021, 13, 1619. [Google Scholar] [CrossRef]
- Matteoni, S.; Abbruzzese, C.; Villani, V.; Malorni, W.; Pace, A.; Matarrese, P.; Paggi, M.G. The influence of patient sex on clinical approaches to malignant glioma. Cancer Lett. 2020, 468, 41–47. [Google Scholar] [CrossRef] [PubMed]
- Garcia, I.; Aldaregia, J.; Marjanovic Vicentic, J.; Aldaz, P.; Moreno-Cugnon, L.; Torres-Bayona, S.; Carrasco-Garcia, E.; Garros-Regulez, L.; Egaña, L.; Rubio, A.; et al. Oncogenic activity of SOX1 in glioblastoma. Sci. Rep. 2017, 7, 46575. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Annovazzi, L.; Mellai, M.; Caldera, V.; Valente, G.; Schiffer, D. SOX2 expression and amplification in gliomas and glioma cell lines. Cancer Genom. Proteom. 2011, 8, 139–147. [Google Scholar]
- Leiss, L.; Mutlu, E.; Øyan, A.; Yan, T.; Tsinkalovsky, O.; Sleire, L.; Petersen, K.; Rahman, M.A.; Johannessen, M.; Mitra, S.S.; et al. Tumour-associated glial host cells display a stem-like phenotype with a distinct gene expression profile and promote growth of GBM xenografts. BMC Cancer 2017, 17, 108. [Google Scholar] [CrossRef] [Green Version]
- Phi, J.H.; Park, S.H.; Kim, S.K.; Paek, S.H.; Kim, J.H.; Lee, Y.J.; Cho, B.K.; Park, C.K.; Lee, D.H.; Wang, K.C. Sox2 expression in brain tumors: A reflection of the neuroglial differentiation pathway. Am. J. Surg. Pathol. 2008, 32, 103–112. [Google Scholar] [CrossRef] [PubMed]
- Schmitz, M.; Temme, A.; Senner, V.; Ebner, R.; Schwind, S.; Stevanovic, S.; Wehner, R.; Schackert, G.; Schackert, H.K.; Fussel, M.; et al. Identification of SOX2 as a novel glioma-associated antigen and potential target for T cell-based immunotherapy. Br. J. Cancer 2007, 96, 1293–1301. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Garros-Regulez, L.; Garcia, I.; Carrasco-Garcia, E.; Lantero, A.; Aldaz, P.; Moreno-Cugnon, L.; Arrizabalaga, O.; Undabeitia, J.; Torres-Bayona, S.; Villanua, J.; et al. Targeting SOX2 as a Therapeutic Strategy in Glioblastoma. Front. Oncol. 2016, 6, 222. [Google Scholar] [CrossRef] [Green Version]
- Fang, X.; Yoon, J.G.; Li, L.; Yu, W.; Shao, J.; Hua, D.; Zheng, S.; Hood, L.; Goodlett, D.R.; Foltz, G.; et al. The SOX2 response program in glioblastoma multiforme: An integrated ChIP-seq, expression microarray, and microRNA analysis. BMC Genom. 2011, 12, 11. [Google Scholar] [CrossRef] [Green Version]
- Pan, C.; Liang, L.; Wang, Z.; Zhang, B.; Li, Q.; Tian, Y.; Yu, Y.; Chen, Z.; Wang, X.; Liu, H. Expression and significance of SOX B1 genes in glioblastoma multiforme patients. J. Cell. Mol. Med. 2022, 26, 789–799. [Google Scholar] [CrossRef]
- Ge, Y.; Zhou, F.; Chen, H.; Cui, C.; Liu, D.; Li, Q.; Yang, Z.; Wu, G.; Sun, S.; Gu, J.; et al. Sox2 is translationally activated by eukaryotic initiation factor 4E in human glioma-initiating cells. Biochem. Biophys. Res. Commun. 2010, 397, 711–717. [Google Scholar] [CrossRef]
- Lu, S.; Yu, Z.; Zhang, X.; Sui, L. MiR-483 Targeted SOX3 to Suppress Glioma Cell Migration, Invasion and Promote Cell Apoptosis. OncoTargets Ther. 2020, 13, 2153–2161. [Google Scholar] [CrossRef] [Green Version]
- Marjanovic Vicentic, J.; Drakulic, D.; Garcia, I.; Vukovic, V.; Aldaz, P.; Puskas, N.; Nikolic, I.; Tasic, G.; Raicevic, S.; Garros-Regulez, L.; et al. SOX3 can promote the malignant behavior of glioblastoma cells. Cell. Oncol. 2019, 42, 41–54. [Google Scholar] [CrossRef]
- Sandberg, M.; Källström, M.; Muhr, J. Sox21 promotes the progression of vertebrate neurogenesis. Nat. Neurosci. 2005, 8, 995–1001. [Google Scholar] [CrossRef]
- Ferletta, M.; Caglayan, D.; Mokvist, L.; Jiang, Y.; Kastemar, M.; Uhrbom, L.; Westermark, B. Forced expression of Sox21 inhibits Sox2 and induces apoptosis in human glioma cells. Int. J. Cancer 2011, 129, 45–60. [Google Scholar] [CrossRef]
- Uchikawa, M.; Kamachi, Y.; Kondoh, H. Two distinct subgroups of Group B Sox genes for transcriptional activators and repressors: Their expression during embryonic organogenesis of the chicken. Mech. Dev. 1999, 84, 103–120. [Google Scholar] [CrossRef]
- Caglayan, D.; Lundin, E.; Kastemar, M.; Westermark, B.; Ferletta, M. Sox21 inhibits glioma progression in vivo by forming complexes with Sox2 and stimulating aberrant differentiation. Int. J. Cancer 2013, 133, 1345–1356. [Google Scholar] [CrossRef] [PubMed]
- Zhang, J.; Jiang, H.; Shao, J.; Mao, R.; Liu, J.; Ma, Y.; Fang, X.; Zhao, N.; Zheng, S.; Lin, B. SOX4 inhibits GBM cell growth and induces G0/G1 cell cycle arrest through Akt-p53 axis. BMC Neurol. 2014, 14, 207. [Google Scholar] [CrossRef] [Green Version]
- Han, W.; Hu, P.; Wu, F.; Wang, S.; Hu, Y.; Li, S.; Jiang, T.; Qiang, B.; Peng, X. FHL3 links cell growth and self-renewal by modulating SOX4 in glioma. Cell Death Differ. 2019, 26, 796–811. [Google Scholar] [CrossRef] [PubMed]
- Hide, T.; Takezaki, T.; Nakatani, Y.; Nakamura, H.; Kuratsu, J.; Kondo, T. Sox11 prevents tumorigenesis of glioma-initiating cells by inducing neuronal differentiation. Cancer Res. 2009, 69, 7953–7959. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Korkolopoulou, P.; Levidou, G.; El-Habr, E.A.; Adamopoulos, C.; Fragkou, P.; Boviatsis, E.; Themistocleous, M.S.; Petraki, K.; Vrettakos, G.; Sakalidou, M.; et al. Sox11 expression in astrocytic gliomas: Correlation with nestin/c-Met/IDH1-R132H expression phenotypes, p-Stat-3 and survival. Br. J. Cancer 2013, 108, 2142–2152. [Google Scholar] [CrossRef] [Green Version]
- Weigle, B.; Ebner, R.; Temme, A.; Schwind, S.; Schmitz, M.; Kiessling, A.; Rieger, M.A.; Schackert, G.; Schackert, H.K.; Rieber, E.P. Highly specific overexpression of the transcription factor SOX11 in human malignant gliomas. Oncol. Rep. 2005, 13, 139–144. [Google Scholar] [CrossRef]
- Tchougounova, E.; Jiang, Y.; Bråsäter, D.; Lindberg, N.; Kastemar, M.; Asplund, A.; Westermark, B.; Uhrbom, L. Sox5 can suppress platelet-derived growth factor B-induced glioma development in Ink4a-deficient mice through induction of acute cellular senescence. Oncogene 2009, 28, 1537–1548. [Google Scholar] [CrossRef] [Green Version]
- Chen, Y.; Liu, W.; Chao, T.; Zhang, Y.; Yan, X.; Gong, Y.; Qiang, B.; Yuan, J.; Sun, M.; Peng, X. MicroRNA-21 down-regulates the expression of tumor suppressor PDCD4 in human glioblastoma cell T98G. Cancer Lett. 2008, 272, 197–205. [Google Scholar] [CrossRef]
- Schlierf, B.; Friedrich, R.P.; Roerig, P.; Felsberg, J.; Reifenberger, G.; Wegner, M. Expression of SoxE and SoxD genes in human gliomas. Neuropathol. Appl. Neurobiol. 2007, 33, 621–630. [Google Scholar] [CrossRef]
- Uhlen, M.; Zhang, C.; Lee, S.; Sjöstedt, E.; Fagerberg, L.; Bidkhori, G.; Benfeitas, R.; Arif, M.; Liu, Z.; Edfors, F.; et al. A pathology atlas of the human cancer transcriptome. Science 2017, 357, eaan2507. [Google Scholar] [CrossRef] [Green Version]
- Wang, L.; He, S.; Yuan, J.; Mao, X.; Cao, Y.; Zong, J.; Tu, Y.; Zhang, Y. Oncogenic role of SOX9 expression in human malignant glioma. Med. Oncol. 2012, 29, 3484–3490. [Google Scholar] [CrossRef]
- Wang, Z.; Xu, X.; Liu, N.; Cheng, Y.; Jin, W.; Zhang, P.; Wang, X.; Yang, H.; Liu, H.; Tu, Y. SOX9-PDK1 axis is essential for glioma stem cell self-renewal and temozolomide resistance. Oncotarget 2017, 9, 192–204. [Google Scholar] [CrossRef] [Green Version]
- Sun, Y.; Jing, Y.; Zhang, Y. Serum lncRNA-ANRIL and SOX9 expression levels in glioma patients and their relationship with poor prognosis. World J. Surg. Oncol. 2021, 19, 287. [Google Scholar] [CrossRef] [PubMed]
- Etcheverry, A.; Aubry, M.; de Tayrac, M.; Vauleon, E.; Boniface, R.; Guenot, F.; Saikali, S.; Hamlat, A.; Riffaud, L.; Menei, P.; et al. DNA methylation in glioblastoma: Impact on gene expression and clinical outcome. BMC Genom. 2010, 11, 701. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Xiuju, C.; Zhen, W.; Yanchao, S. SOX7 inhibits tumor progression of glioblastoma and is regulated by miRNA-24. Open Med. 2016, 11, 133–137. [Google Scholar] [CrossRef] [PubMed]
- Zhao, T.; Yang, H.; Tian, Y.; Xie, Q.; Lu, Y.; Wang, Y.; Su, N.; Dong, B.; Liu, X.; Wang, C.; et al. SOX7 is associated with the suppression of human glioma by HMG-box dependent regulation of Wnt/β-catenin signaling. Cancer Lett. 2016, 375, 100–107. [Google Scholar] [CrossRef] [PubMed]
- Majchrzak-Celińska, A.; Słocińska, M.; Barciszewska, A.M.; Nowak, S.; Baer-Dubowska, W. Wnt pathway antagonists, SFRP1, SFRP2, SOX17, and PPP2R2B, are methylated in gliomas and SFRP1 methylation predicts shorter survival. J. Appl. Genet. 2016, 57, 189–197. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, D.; Guo, S.; Wang, H.; Hu, Y. SOX15 exerts antitumor function in glioma by inhibiting cell proliferation and invasion via downregulation of Wnt/beta-catenin signaling. Life Sci. 2020, 255, 117792. [Google Scholar] [CrossRef] [PubMed]
- Brosnan, C.A.; Voinnet, O. The long and the short of noncoding RNAs. Curr. Opin. Cell Biol. 2009, 21, 416–425. [Google Scholar] [CrossRef] [PubMed]
- Statello, L.; Guo, C.J.; Chen, L.L.; Huarte, M. Gene regulation by long non-coding RNAs and its biological functions. Nat. Rev. Mol. Cell Biol. 2021, 22, 96–118. [Google Scholar] [CrossRef] [PubMed]
- Ma, L.; Bajic, V.B.; Zhang, Z. On the classification of long non-coding RNAs. RNA Biol. 2013, 10, 925–933. [Google Scholar] [CrossRef] [Green Version]
- Momtazmanesh, S.; Rezaei, N. Long Non-Coding RNAs in Diagnosis, Treatment, Prognosis, and Progression of Glioma: A State-of-the-Art Review. Front. Oncol. 2021, 11, 712786. [Google Scholar] [CrossRef]
- Mahinfar, P.; Baradaran, B.; Davoudian, S.; Vahidian, F.; Cho, W.C.; Mansoori, B. Long Non-Coding RNAs in Multidrug Resistance of Glioblastoma. Genes 2021, 12, 455. [Google Scholar] [CrossRef]
- Melendez-Zajgla, J.; Maldonado, V. The Role of lncRNAs in the Stem Phenotype of Pancreatic Ductal Adenocarcinoma. Int. J. Mol. Sci. 2021, 22, 6374. [Google Scholar] [CrossRef]
- Blokhin, I.; Khorkova, O.; Hsiao, J.; Wahlestedt, C. Developments in lncRNA drug discovery: Where are we heading? Expert Opin. Drug Discov. 2018, 13, 837–849. [Google Scholar] [CrossRef]
- Campos-Parra, A.D.; López-Urrutia, E.; Orozco Moreno, L.T.; López-Camarillo, C.; Meza-Menchaca, T.; Figueroa González, G.; Bustamante Montes, L.P.; Pérez-Plasencia, C. Long Non-Coding RNAs as New Master Regulators of Resistance to Systemic Treatments in Breast Cancer. Int. J. Mol. Sci. 2018, 19, 2711. [Google Scholar] [CrossRef] [Green Version]
- Saleembhasha, A.; Mishra, S. Long non-coding RNAs as pan-cancer master gene regulators of associated protein-coding genes: A systems biology approach. PeerJ 2019, 7, e6388. [Google Scholar] [CrossRef] [Green Version]
- Stackhouse, C.T.; Gillespie, G.Y.; Willey, C.D. Exploring the Roles of lncRNAs in GBM Pathophysiology and Their Therapeutic Potential. Cells 2020, 9, 2369. [Google Scholar] [CrossRef] [PubMed]
- Rezaei, O.; Tamizkar, K.H.; Sharifi, G.; Taheri, M.; Ghafouri-Fard, S. Emerging Role of Long Non-Coding RNAs in the Pathobiology of Glioblastoma. Front. Oncol. 2020, 10, 625884. [Google Scholar] [CrossRef]
- Torres-Bayona, S.; Aldaz, P.; Auzmendi-Iriarte, J.; Saenz-Antonanzas, A.; Garcia, I.; Arrazola, M.; Gerovska, D.; Undabeitia, J.; Querejeta, A.; Egana, L.; et al. PR-LncRNA signature regulates glioma cell activity through expression of SOX factors. Sci. Rep. 2018, 8, 12746. [Google Scholar] [CrossRef] [Green Version]
- Guan, N.; Wang, R.; Feng, X.; Li, C.; Guo, W. Long non-coding RNA NBAT1 inhibits the progression of glioma through the miR-21/SOX7 axis. Oncol. Lett. 2020, 20, 3024–3034. [Google Scholar] [CrossRef] [PubMed]
- Xiong, Z.; Wang, L.; Wang, Q.; Yuan, Y. LncRNA MALAT1/miR-129 axis promotes glioma tumorigenesis by targeting SOX2. J. Cell. Mol. Med. 2018, 22, 3929–3940. [Google Scholar] [CrossRef]
- Zhou, K.; Zhang, C.; Yao, H.; Zhang, X.; Zhou, Y.; Che, Y.; Huang, Y. Knockdown of long non-coding RNA NEAT1 inhibits glioma cell migration and invasion via modulation of SOX2 targeted by miR-132. Mol. Cancer 2018, 17, 105. [Google Scholar] [CrossRef] [Green Version]
- Guo, J.; Cai, H.; Zheng, J.; Liu, X.; Liu, Y.; Ma, J.; Que, Z.; Gong, W.; Gao, Y.; Tao, W.; et al. Long non-coding RNA NEAT1 regulates permeability of the blood-tumor barrier via miR-181d-5p-mediated expression changes in ZO-1, occludin, and claudin-5. Biochim Biophys Acta Mol. Basis Dis. 2017, 1863, 2240–2254. [Google Scholar] [CrossRef]
- Katsushima, K.; Natsume, A.; Ohka, F.; Shinjo, K.; Hatanaka, A.; Ichimura, N.; Sato, S.; Takahashi, S.; Kimura, H.; Totoki, Y.; et al. Targeting the Notch-regulated non-coding RNA TUG1 for glioma treatment. Nat. Commun. 2016, 7, 13616. [Google Scholar] [CrossRef] [PubMed]
- Brodie, S.; Lee, H.K.; Jiang, W.; Cazacu, S.; Xiang, C.; Poisson, L.M.; Datta, I.; Kalkanis, S.; Ginsberg, D.; Brodie, C. The novel long non-coding RNA TALNEC2, regulates tumor cell growth and the stemness and radiation response of glioma stem cells. Oncotarget 2017, 8, 31785–31801. [Google Scholar] [CrossRef] [Green Version]
- Gong, R.; Li, Z.Q.; Fu, K.; Ma, C.; Wang, W.; Chen, J.C. Long Noncoding RNA PVT1 Promotes Stemness and Temozolomide Resistance through miR-365/ELF4/SOX2 Axis in Glioma. Exp. Neurobiol. 2021, 30, 244–255. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Z.; Zhou, J.; Zhang, J.; Duan, R.; Pu, P.; Han, L. Downregulation of lncRNA-HOXA11-AS modulates proliferation and stemness in Glioma cells. Chin. Neurosurg. J. 2017, 3, 25. [Google Scholar] [CrossRef] [Green Version]
- Li, W.; Jiang, P.; Sun, X.; Xu, S.; Ma, X.; Zhan, R. Suppressing H19 Modulates Tumorigenicity and Stemness in U251 and U87MG Glioma Cells. Cell. Mol. Neurobiol. 2016, 36, 1219–1227. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hu, Q.; Yin, J.; Zeng, A.; Jin, X.; Zhang, Z.; Yan, W.; You, Y. H19 Functions as a Competing Endogenous RNA to Regulate EMT by Sponging miR-130a-3p in Glioma. Cell Physiol. Biochem. 2018, 50, 233–245. [Google Scholar] [CrossRef] [PubMed]
- Shahzad, U.; Li, C.; Johnston, M.; Wang, J.J.; Sabha, N.; Varn, F.S.; Riemenschneider, A.; Krumholtz, S.; Meda, P.; Smith, C.A.; et al. CASCADES, a novel SOX2 super-enhancer associated long noncoding RNA, regulates cancer stem cell specification and differentiation in glioblastoma multiforme. bioRxiv 2020. [Google Scholar] [CrossRef]
- Luo, C.; Quan, Z.; Zhong, B.; Zhang, M.; Zhou, B.; Wang, S.; Luo, X.; Tang, C. lncRNA XIST promotes glioma proliferation and metastasis through miR-133a/SOX4. Exp. Ther. Med. 2020, 19, 1641–1648. [Google Scholar] [CrossRef] [Green Version]
- Wu, J.; Li, R.; Li, L.; Gu, Y.; Zhan, H.; Zhou, C.; Zhong, C. MYC-activated lncRNA HNF1A-AS1 overexpression facilitates glioma progression via cooperating with miR-32-5p/SOX4 axis. Cancer Med. 2020, 9, 6387–6398. [Google Scholar] [CrossRef]
- Wang, C.J.; Chao, C.R.; Zhao, W.F.; Liu, H.M.; Feng, J.S.; Cui, Y.X. Long noncoding RNA SNHG9 facilitates growth of glioma stem-like cells via miR-326/SOX9 axis. J. Gene Med. 2022, 24, e3334. [Google Scholar] [CrossRef]
- Liu, X.; Zheng, J.; Xue, Y.; Qu, C.; Chen, J.; Wang, Z.; Li, Z.; Zhang, L.; Liu, Y. Inhibition of TDP43-Mediated SNHG12-miR-195-SOX5 Feedback Loop Impeded Malignant Biological Behaviors of Glioma Cells. Mol. Ther. Nucleic Acids 2018, 10, 142–158. [Google Scholar] [CrossRef] [Green Version]
- Li, Z.; Zhang, J.; Zheng, H.; Li, C.; Xiong, J.; Wang, W.; Bao, H.; Jin, H.; Liang, P. Modulating lncRNA SNHG15/CDK6/miR-627 circuit by palbociclib, overcomes temozolomide resistance and reduces M2-polarization of glioma associated microglia in glioblastoma multiforme. J. Exp. Clin. Cancer Res. 2019, 38, 380. [Google Scholar] [CrossRef]
- Li, Y.; Zhu, G.; Zeng, W.; Wang, J.; Li, Z.; Wang, B.; Tian, B.; Lu, D.; Zhang, X.; Gao, G.; et al. Long noncoding RNA AB073614 promotes the malignance of glioma by activating Wnt/β-catenin signaling through downregulating SOX7. Oncotarget 2017, 8, 65577–65587. [Google Scholar] [CrossRef] [Green Version]
- Wu, B.; Yang, C.; Fang, Y.; Ding, W.; Zhang, Y. Long noncoding RNA DUXAP10 promotes the stemness of glioma cells by recruiting HuR to enhance Sox12 mRNA stability. Environ. Toxicol. 2021, 36, 840–849. [Google Scholar] [CrossRef]
- Su, R.; Cao, S.; Ma, J.; Liu, Y.; Liu, X.; Zheng, J.; Chen, J.; Liu, L.; Cai, H.; Li, Z.; et al. Knockdown of SOX2OT inhibits the malignant biological behaviors of glioblastoma stem cells via up-regulating the expression of miR-194-5p and miR-122. Mol. Cancer 2017, 16, 171. [Google Scholar] [CrossRef] [Green Version]
- Sanchez, Y.; Segura, V.; Marin-Bejar, O.; Athie, A.; Marchese, F.P.; Gonzalez, J.; Bujanda, L.; Guo, S.; Matheu, A.; Huarte, M. Genome-wide analysis of the human p53 transcriptional network unveils a lncRNA tumour suppressor signature. Nat. Commun. 2014, 5, 5812. [Google Scholar] [CrossRef] [Green Version]
- Bradner, J.E.; Hnisz, D.; Young, R.A. Transcriptional Addiction in Cancer. Cell 2017, 168, 629–643. [Google Scholar] [CrossRef] [Green Version]
- Huang, J.; Li, K.; Cai, W.; Liu, X.; Zhang, Y.; Orkin, S.H.; Xu, J.; Yuan, G.C. Dissecting super-enhancer hierarchy based on chromatin interactions. Nat. Commun. 2018, 9, 943. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sengupta, S.; George, R.E. Super-Enhancer-Driven Transcriptional Dependencies in Cancer. Trends Cancer 2017, 3, 269–281. [Google Scholar] [CrossRef] [Green Version]
- Chang, H.C.; Huang, H.C.; Juan, H.F.; Hsu, C.L. Investigating the role of super-enhancer RNAs underlying embryonic stem cell differentiation. BMC Genom. 2019, 20, 896. [Google Scholar] [CrossRef] [Green Version]
- Lee, J.H.; Xiong, F.; Li, W. Enhancer RNAs in cancer: Regulation, mechanisms and therapeutic potential. RNA Biol. 2020, 17, 1550–1559. [Google Scholar] [CrossRef]
- Madden, E.; Logue, S.E.; Healy, S.J.; Manie, S.; Samali, A. The role of the unfolded protein response in cancer progression: From oncogenesis to chemoresistance. Biol. Cell 2019, 111, 1–17. [Google Scholar] [CrossRef] [Green Version]
- Wang, X.; Jiang, Q.; Zhang, C.; Yang, Q.; Wang, L.; Zhang, J.; Wang, L.; Chen, X.; Hou, X.; Han, D.; et al. Long noncoding RNA SNHG12 is a potential diagnostic and prognostic biomarker in various tumors. Chin. Neurosurg. J. 2021, 7, 37. [Google Scholar] [CrossRef] [PubMed]
- Kent, W.J.; Sugnet, C.W.; Furey, T.S.; Roskin, K.M.; Pringle, T.H.; Zahler, A.M.; Haussler, D. The human genome browser at UCSC. Genome Res. 2002, 12, 996–1006. [Google Scholar] [CrossRef] [Green Version]
- Ahmad, A.; Strohbuecker, S.; Tufarelli, C.; Sottile, V. Expression of a SOX1 overlapping transcript in neural differentiation and cancer models. Cell. Mol. Life Sci. 2017, 74, 4245–4258. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Xi, J.; Xu, Y.; Guo, Z.; Li, J.; Wu, Y.; Sun, Q.; Wang, Y.; Chen, M.; Zhu, S.; Bian, S.; et al. LncRNA SOX1-OT V1 acts as a decoy of HDAC10 to promote SOX1-dependent hESC neuronal differentiation. EMBO Rep. 2022, 23, e53015. [Google Scholar] [CrossRef]
- Amaral, P.P.; Neyt, C.; Wilkins, S.J.; Askarian-Amiri, M.E.; Sunkin, S.M.; Perkins, A.C.; Mattick, J.S. Complex architecture and regulated expression of the Sox2ot locus during vertebrate development. RNA 2009, 15, 2013–2027. [Google Scholar] [CrossRef] [Green Version]
- Shahryari, A.; Rafiee, M.R.; Fouani, Y.; Oliae, N.A.; Samaei, N.M.; Shafiee, M.; Semnani, S.; Vasei, M.; Mowla, S.J. Two novel splice variants of SOX2OT, SOX2OT-S1, and SOX2OT-S2 are coupregulated with SOX2 and OCT4 in esophageal squamous cell carcinoma. Stem Cells 2014, 32, 126–134. [Google Scholar] [CrossRef]
- Wang, Y.; Wu, N.; Luo, X.; Zhang, X.; Liao, Q.; Wang, J. SOX2OT, a novel tumor-related long non-coding RNA. Biomed. Pharmacother. 2020, 123, 109725. [Google Scholar] [CrossRef]
- Saghaeian Jazi, M.; Samaei, N.M.; Ghanei, M.; Shadmehr, M.B.; Mowla, S.J. Identification of new SOX2OT transcript variants highly expressed in human cancer cell lines and down regulated in stem cell differentiation. Mol. Biol. Rep. 2016, 43, 65–72. [Google Scholar] [CrossRef]
- Messemaker, T.C.; van Leeuwen, S.M.; van den Berg, P.R.; ‘t Jong, A.E.J.; Palstra, R.-J.; Hoeben, R.C.; Semrau, S.; Mikkers, H.M.M. Allele-specific repression of Sox2 through the long non-coding RNA Sox2ot. Sci. Rep. 2018, 8, 386. [Google Scholar] [CrossRef] [Green Version]
- Knauss, J.L.; Miao, N.; Kim, S.-N.; Nie, Y.; Shi, Y.; Wu, T.; Pinto, H.B.; Donohoe, M.E.; Sun, T. Long noncoding RNA Sox2ot and transcription factor YY1 co-regulate the differentiation of cortical neural progenitors by repressing Sox2. Cell Death Dis. 2018, 9, 799. [Google Scholar] [CrossRef] [Green Version]
- Li, P.Y.; Wang, P.; Gao, S.G.; Dong, D.Y. Long Noncoding RNA SOX2-OT: Regulations, Functions, and Roles on Mental Illnesses, Cancers, and Diabetic Complications. BioMed Res. Int. 2020, 2020, 2901589. [Google Scholar] [CrossRef] [PubMed]
- Shyamsundar, R.; Kim, Y.H.; Higgins, J.P.; Montgomery, K.; Jorden, M.; Sethuraman, A.; van de Rijn, M.; Botstein, D.; Brown, P.O.; Pollack, J.R. A DNA microarray survey of gene expression in normal human tissues. Genome Biol. 2005, 6, R22. [Google Scholar] [CrossRef] [Green Version]
- Yanai, I.; Benjamin, H.; Shmoish, M.; Chalifa-Caspi, V.; Shklar, M.; Ophir, R.; Bar-Even, A.; Horn-Saban, S.; Safran, M.; Domany, E.; et al. Genome-wide midrange transcription profiles reveal expression level relationships in human tissue specification. Bioinformatics 2005, 21, 650–659. [Google Scholar] [CrossRef] [Green Version]
- Askarian-Amiri, M.E.; Seyfoddin, V.; Smart, C.E.; Wang, J.; Kim, J.E.; Hansji, H.; Baguley, B.C.; Finlay, G.J.; Leung, E.Y. Emerging role of long non-coding RNA SOX2OT in SOX2 regulation in breast cancer. PLoS ONE 2014, 9, e102140. [Google Scholar] [CrossRef]
- Zhang, J.J.; Zhu, Y.; Zhang, X.F.; Liu, D.F.; Wang, Y.; Yang, C.; Shi, G.D.; Peng, Y.P.; Zhang, K.; Tian, L.; et al. Yin Yang-1 suppresses pancreatic ductal adenocarcinoma cell proliferation and tumor growth by regulating SOX2OT-SOX2 axis. Cancer Lett. 2017, 408, 144–154. [Google Scholar] [CrossRef]
- Wei, C.X.; Wong, H.; Xu, F.; Liu, Z.; Ran, L.; Jiang, R.D. IRF4-induced upregulation of lncRNA SOX2-OT promotes cell proliferation and metastasis in cholangiocarcinoma by regulating SOX2 and PI3K/AKT signaling. Eur. Rev. Med. Pharmacol. Sci. 2018, 22, 8169–8178. [Google Scholar] [CrossRef]
- Wang, Z.; Tan, M.; Chen, G.; Li, Z.; Lu, X. LncRNA SOX2-OT is a novel prognostic biomarker for osteosarcoma patients and regulates osteosarcoma cells proliferation and motility through modulating SOX2. IUBMB Life 2017, 69, 867–876. [Google Scholar] [CrossRef] [Green Version]
- Chang, X.; Zhang, H.; Yang, Q.; Pang, L. LncRNA SOX2OT affects cervical cancer cell growth, migration and invasion by regulating SOX2. Cell Cycle 2020, 19, 1391–1403. [Google Scholar] [CrossRef]
- Zhan, Y.; Chen, Z.; He, S.; Gong, Y.; He, A.; Li, Y.; Zhang, L.; Zhang, X.; Fang, D.; Li, X.; et al. Long non-coding RNA SOX2OT promotes the stemness phenotype of bladder cancer cells by modulating SOX2. Mol. Cancer 2020, 19, 25. [Google Scholar] [CrossRef] [Green Version]
- Feng, Y.; Xu, Y.; Gao, Y.; Chen, Y.; Wang, X.; Chen, Z. A novel lncRNA SOX2OT promotes the malignancy of human colorectal cancer by interacting with miR-194-5p/SOX5 axis. Cell Death Dis. 2021, 12, 499. [Google Scholar] [CrossRef]
- Wu, Y.; Chen, X.; Liang, Y.; Li, J.; Zhang, K.; Dai, L.; Guan, X.; Wang, K.; Bai, Y. Overexpression of long non-coding RNA SOX2OT promotes esophageal squamous cell carcinoma growth. Cancer Cell Int. 2018, 18, 76. [Google Scholar] [CrossRef] [PubMed]
- Hou, Z.; Zhao, W.; Zhou, J.; Shen, L.; Zhan, P.; Xu, C.; Chang, C.; Bi, H.; Zou, J.; Yao, X.; et al. A long noncoding RNA Sox2ot regulates lung cancer cell proliferation and is a prognostic indicator of poor survival. Int. J. Biochem. Cell Biol. 2014, 53, 380–388. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Y.; Yang, R.; Lian, J.; Xu, H. LncRNA Sox2ot overexpression serves as a poor prognostic biomarker in gastric cancer. Am. J. Transl. Res. 2016, 8, 5035–5043. [Google Scholar]
- Zou, J.H.; Li, C.Y.; Bao, J.; Zheng, G.Q. High expression of long noncoding RNA Sox2ot is associated with the aggressive progression and poor outcome of gastric cancer. Eur. Rev. Med. Pharmacol. Sci. 2016, 20, 4482–4486. [Google Scholar]
- Shi, X.M.; Teng, F. Up-regulation of long non-coding RNA Sox2ot promotes hepatocellular carcinoma cell metastasis and correlates with poor prognosis. Int. J. Clin. Exp. Pathol. 2015, 8, 4008–4014. [Google Scholar]
- Sun, J.; Wei, X.; Xu, L. Upregulation of lncRNA Sox2ot indicates a poor prognosis for patients with hepatocellular carcinoma and promotes cell invasion. Oncol. Lett. 2018, 16, 1189–1195. [Google Scholar] [CrossRef]
- Han, L.; Zhang, W.; Zhang, B.; Zhan, L. Long non-coding RNA SOX2OT promotes cell proliferation and motility in human ovarian cancer. Exp. Ther. Med. 2018, 15, 2182–2188. [Google Scholar] [CrossRef]
- Zhang, E.; Li, X. LncRNA SOX2-OT regulates proliferation and metastasis of nasopharyngeal carcinoma cells through miR-146b-5p/HNRNPA2B1 pathway. J. Cell. Biochem. 2019, 120, 16575–16588. [Google Scholar] [CrossRef]
- Song, X.; Wang, H.; Wu, J.; Sun, Y. Long Noncoding RNA SOX2-OT Knockdown Inhibits Proliferation and Metastasis of Prostate Cancer Cells Through Modulating the miR-452-5p/HMGB3 Axis and Inactivating Wnt/β-Catenin Pathway. Cancer Biother. Radiopharm. 2020, 35, 682–695. [Google Scholar] [CrossRef]
- Fantes, J.; Ragge, N.K.; Lynch, S.A.; McGill, N.I.; Collin, J.R.; Howard-Peebles, P.N.; Hayward, C.; Vivian, A.J.; Williamson, K.; van Heyningen, V.; et al. Mutations in SOX2 cause anophthalmia. Nat. Genet. 2003, 33, 461–463. [Google Scholar] [CrossRef] [Green Version]
- Zeng, H.; Xu, N.; Liu, Y.; Liu, B.; Yang, Z.; Fu, Z.; Lian, C.; Guo, H. Genomic profiling of long non-coding RNA and mRNA expression associated with acquired temozolomide resistance in glioblastoma cells. Int. J. Oncol. 2017, 51, 445–455. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, H.; Hu, Q.; Tong, Y.; Li, S.; Chen, M.; Wang, B.; Li, H. LncRNA SOX2-OT regulates miR-192-5p/RAB2A axis and ERK pathway to promote glioblastoma cell growth. Cell Cycle 2021, 20, 2010–2020. [Google Scholar] [CrossRef] [PubMed]
- Saghaeian Jazi, M.; Samaei, N.M.; Mowla, S.J.; Arefnezhad, B.; Kouhsar, M. SOX2OT knockdown derived changes in mitotic regulatory gene network of cancer cells. Cancer Cell Int. 2018, 18, 129. [Google Scholar] [CrossRef] [Green Version]
- Liang, Y.; Zhang, D.; Zheng, T.; Yang, G.; Wang, J.; Meng, F.; Liu, Y.; Zhang, G.; Zhang, L.; Han, J.; et al. lncRNA-SOX2OT promotes hepatocellular carcinoma invasion and metastasis through miR-122-5p-mediated activation of PKM2. Oncogenesis 2020, 9, 54. [Google Scholar] [CrossRef]
- Tianhua, Y.; Dianqiu, L.; Xuanhe, Z.; Zhe, Z.; Dongmei, G. Long non-coding RNA Sox2 overlapping transcript (SOX2OT) promotes multiple myeloma progression via microRNA-143-3p/c-MET axis. J. Cell. Mol. Med. 2020, 24, 5185–5194. [Google Scholar] [CrossRef] [Green Version]
- Zhang, W.; Yang, S.; Chen, D.; Yuwen, D.; Zhang, J.; Wei, X.; Han, X.; Guan, X. SOX2-OT induced by PAI-1 promotes triple-negative breast cancer cells metastasis by sponging miR-942-5p and activating PI3K/Akt signaling. Cell. Mol. Life Sci. 2022, 79, 59. [Google Scholar] [CrossRef]
- Zhu, K.; Yuan, Y.; Wen, J.; Chen, D.; Zhu, W.; Ouyang, Z.; Wang, W. LncRNA Sox2OT-V7 promotes doxorubicin-induced autophagy and chemoresistance in osteosarcoma via tumor-suppressive miR-142/miR-22. Aging 2020, 12, 6644–6666. [Google Scholar] [CrossRef]
- Chen, H.; Lu, Q.; Fei, X.; Shen, L.; Jiang, D.; Dai, D. miR-22 inhibits the proliferation, motility, and invasion of human glioblastoma cells by directly targeting SIRT1. Tumour Biol. 2016, 37, 6761–6768. [Google Scholar] [CrossRef]
- Cheng, Z.X.; Song, Y.X.; Wang, Z.Y.; Wang, Y.; Dong, Y. miR-144-3p serves as a tumor suppressor by targeting FZD7 and predicts the prognosis of human glioblastoma. Eur. Rev. Med. Pharmacol. Sci. 2017, 21, 4079–4086. [Google Scholar]
- Ding, C.Q.; Deng, W.S.; Yin, X.F.; Ding, X.D. MiR-122 inhibits cell proliferation and induces apoptosis by targeting runt-related transcription factors 2 in human glioma. Eur. Rev. Med. Pharmacol. Sci. 2018, 22, 4925–4933. [Google Scholar] [CrossRef]
- Gheidari, F.; Arefian, E.; Jamshidi Adegani, F.; Fallah Atanaki, F.; Soleimani, M. The miR-142 Suppresses U-87 Glioblastoma Cell Growth by Targeting EGFR Oncogenic Signaling Pathway. Iran. J. Pharm. Res. 2021, 20, 202–212. [Google Scholar] [CrossRef] [PubMed]
- Li, Y.; Wang, Y.; Yu, L.; Sun, C.; Cheng, D.; Yu, S.; Wang, Q.; Yan, Y.; Kang, C.; Jin, S.; et al. miR-146b-5p inhibits glioma migration and invasion by targeting MMP16. Cancer Lett. 2013, 339, 260–269. [Google Scholar] [CrossRef] [PubMed]
- Liu, L.; Chen, K.; Wu, J.; Shi, L.; Hu, B.; Cheng, S.; Li, M.; Song, L. Downregulation of miR-452 promotes stem-like traits and tumorigenicity of gliomas. Clin. Cancer Res. 2013, 19, 3429–3438. [Google Scholar] [CrossRef] [Green Version]
- Liu, N.Z.; Li, T.; Liu, C.M.; Liu, F.R.; Wang, Y.X. Hsa_circ_0000337 promotes proliferation, migration and invasion in glioma by competitively binding miRNA-942-5p and thus upregulates MAT2A. Eur. Rev. Med. Pharmacol. Sci. 2020, 24, 12251–12257. [Google Scholar] [CrossRef]
- Qin, Y.; Chen, W.; Liu, B.; Zhou, L.; Deng, L.; Niu, W.; Bao, D.; Cheng, C.; Li, D.; Liu, S.; et al. MiR-200c Inhibits the Tumor Progression of Glioma via Targeting Moesin. Theranostics 2017, 7, 1663–1673. [Google Scholar] [CrossRef]
- Wang, W.; Chen, D.; Zhu, K. SOX2OT variant 7 contributes to the synergistic interaction between EGCG and Doxorubicin to kill osteosarcoma via autophagy and stemness inhibition. J. Exp. Clin. Cancer Res. 2018, 37, 37. [Google Scholar] [CrossRef] [Green Version]
- Yang, C.M.; Wang, T.H.; Chen, H.C.; Li, S.C.; Lee, M.C.; Liou, H.H.; Liu, P.F.; Tseng, Y.K.; Shiue, Y.L.; Ger, L.P.; et al. Aberrant DNA hypermethylation-silenced SOX21-AS1 gene expression and its clinical importance in oral cancer. Clin. Epigenet. 2016, 8, 129. [Google Scholar] [CrossRef] [Green Version]
- Zhao, Z.; Zhang, K.-N.; Wang, Q.; Li, G.; Zeng, F.; Zhang, Y.; Wu, F.; Chai, R.; Wang, Z.; Zhang, C.; et al. Chinese Glioma Genome Atlas (CGGA): A Comprehensive Resource with Functional Genomic Data from Chinese Glioma Patients. Genom. Proteom. Bioinform. 2021, 19, 1–12. [Google Scholar] [CrossRef]
- Cao, Y.; Zhu, H.; Tan, J.; Yin, W.; Zhou, Q.; Xin, Z.; Wu, Z.; Jiang, Z.; Guo, Y.; Kuang, Y.; et al. Development of an Immune-Related LncRNA Prognostic Signature for Glioma. Front. Genet. 2021, 12, 678436. [Google Scholar] [CrossRef]
- Zhou, M.; Zhang, Z.; Zhao, H.; Bao, S.; Cheng, L.; Sun, J. An Immune-Related Six-lncRNA Signature to Improve Prognosis Prediction of Glioblastoma Multiforme. Mol. Neurobiol. 2018, 55, 3684–3697. [Google Scholar] [CrossRef]
- Luan, F.; Chen, W.; Chen, M.; Yan, J.; Chen, H.; Yu, H.; Liu, T.; Mo, L. An autophagy-related long non-coding RNA signature for glioma. FEBS Open Bio 2019, 9, 653–667. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tao, C.; Luo, H.; Chen, L.; Li, J.; Zhu, X.; Huang, K. Identification of an epithelial-mesenchymal transition related long non-coding RNA (LncRNA) signature in Glioma. Bioengineered 2021, 12, 4016–4031. [Google Scholar] [CrossRef]
- Tanzhu, G.; Li, N.; Li, Z.; Zhou, R.; Shen, L. Molecular Subtypes and Prognostic Signature of Pyroptosis-Related lncRNAs in Glioma Patients. Front. Oncol. 2022, 12, 779168. [Google Scholar] [CrossRef]
- Zheng, J.; Zhou, Z.; Qiu, Y.; Wang, M.; Yu, H.; Wu, Z.; Wang, X.; Jiang, X. A Prognostic Ferroptosis-Related lncRNAs Signature Associated With Immune Landscape and Radiotherapy Response in Glioma. Front. Cell Dev. Biol. 2021, 9, 675555. [Google Scholar] [CrossRef]
- Cheng, M.; Sun, L.; Huang, K.; Yue, X.; Chen, J.; Zhang, Z.; Zhao, B.; Bian, E. A Signature of Nine lncRNA Methylated Genes Predicts Survival in Patients With Glioma. Front. Oncol. 2021, 11, 646409. [Google Scholar] [CrossRef]
- Liu, G.; Liu, D.; Huang, J.; Li, J.; Wang, C.; Liu, G.; Ge, S.; Gong, H. Comprehensive analysis of ceRNA network related to lincRNA in glioblastoma and prediction of clinical prognosis. BMC Cancer 2021, 21, 98. [Google Scholar] [CrossRef]
- Peng, Q.; Li, R.; Li, Y.; Xu, X.; Ni, W.; Lin, H.; Ning, L. Prediction of a competing endogenous RNA co-expression network as a prognostic marker in glioblastoma. J. Cell. Mol. Med. 2020, 24, 13346–13355. [Google Scholar] [CrossRef]
- Wang, H.; Zhang, H.; Zeng, J.; Tan, Y. ceRNA network analysis reveals prognostic markers for glioblastoma. Oncol. Lett. 2019, 17, 5545–5557. [Google Scholar] [CrossRef]
- Zhu, X.; Jiang, L.; Yang, H.; Chen, T.; Wu, X.; Lv, K. Analyzing the lncRNA, miRNA, and mRNA-associated ceRNA networks to reveal potential prognostic biomarkers for glioblastoma multiforme. Cancer Cell Int. 2020, 20, 393. [Google Scholar] [CrossRef]
- Li, Q.; Yu, Q.; Ji, J.; Wang, P.; Li, D. Comparison and analysis of lncRNA-mediated ceRNA regulation in different molecular subtypes of glioblastoma. Mol. Omics 2019, 15, 406–419. [Google Scholar] [CrossRef]
- Arun, G.; Diermeier, S.D.; Spector, D.L. Therapeutic Targeting of Long Non-Coding RNAs in Cancer. Trends Mol. Med. 2018, 24, 257–277. [Google Scholar] [CrossRef] [PubMed]
- Hammond, S.M.; Boettcher, S.; Caudy, A.A.; Kobayashi, R.; Hannon, G.J. Argonaute2, a link between genetic and biochemical analyses of RNAi. Science 2001, 293, 1146–1150. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hannon, G.J.; Rossi, J.J. Unlocking the potential of the human genome with RNA interference. Nature 2004, 431, 371–378. [Google Scholar] [CrossRef] [PubMed]
- Liu, S.J.; Horlbeck, M.A.; Cho, S.W.; Birk, H.S.; Malatesta, M.; He, D.; Attenello, F.J.; Villalta, J.E.; Cho, M.Y.; Chen, Y.; et al. CRISPRi-based genome-scale identification of functional long noncoding RNA loci in human cells. Science 2017, 355, aah7111. [Google Scholar] [CrossRef] [Green Version]
- Kim, S.S.; Harford, J.B.; Moghe, M.; Rait, A.; Pirollo, K.F.; Chang, E.H. Targeted nanocomplex carrying siRNA against MALAT1 sensitizes glioblastoma to temozolomide. Nucleic Acids Res. 2018, 46, 1424–1440. [Google Scholar] [CrossRef] [Green Version]
- Cheng, H.; Zhao, H.; Xiao, X.; Huang, Q.; Zeng, W.; Tian, B.; Ma, T.; Lu, D.; Jin, Y.; Li, Y. Long Non-coding RNA MALAT1 Upregulates ZEB2 Expression to Promote Malignant Progression of Glioma by Attenuating miR-124. Mol. Neurobiol. 2021, 58, 1006–1016. [Google Scholar] [CrossRef]
- Arun, G.; Diermeier, S.; Akerman, M.; Chang, K.C.; Wilkinson, J.E.; Hearn, S.; Kim, Y.; MacLeod, A.R.; Krainer, A.R.; Norton, L.; et al. Differentiation of mammary tumors and reduction in metastasis upon Malat1 lncRNA loss. Genes Dev. 2016, 30, 34–51. [Google Scholar] [CrossRef] [Green Version]
- Gutschner, T.; Hämmerle, M.; Eissmann, M.; Hsu, J.; Kim, Y.; Hung, G.; Revenko, A.; Arun, G.; Stentrup, M.; Gross, M.; et al. The noncoding RNA MALAT1 is a critical regulator of the metastasis phenotype of lung cancer cells. Cancer Res. 2013, 73, 1180–1189. [Google Scholar] [CrossRef] [Green Version]
- Adriaens, C.; Standaert, L.; Barra, J.; Latil, M.; Verfaillie, A.; Kalev, P.; Boeckx, B.; Wijnhoven, P.W.; Radaelli, E.; Vermi, W.; et al. p53 induces formation of NEAT1 lncRNA-containing paraspeckles that modulate replication stress response and chemosensitivity. Nat. Med. 2016, 22, 861–868. [Google Scholar] [CrossRef]
- Chen, Q.; Cai, J.; Wang, Q.; Wang, Y.; Liu, M.; Yang, J.; Zhou, J.; Kang, C.; Li, M.; Jiang, C. Long Noncoding RNA NEAT1, Regulated by the EGFR Pathway, Contributes to Glioblastoma Progression Through the WNT/β-Catenin Pathway by Scaffolding EZH2. Clin. Cancer Res. 2018, 24, 684–695. [Google Scholar] [CrossRef] [Green Version]
- Winkle, M.; El-Daly, S.M.; Fabbri, M.; Calin, G.A. Noncoding RNA therapeutics—Challenges and potential solutions. Nat. Rev. Drug Discov. 2021, 20, 629–651. [Google Scholar] [CrossRef]
- Adams, D.; Gonzalez-Duarte, A.; O’Riordan, W.D.; Yang, C.C.; Ueda, M.; Kristen, A.V.; Tournev, I.; Schmidt, H.H.; Coelho, T.; Berk, J.L.; et al. Patisiran, an RNAi Therapeutic, for Hereditary Transthyretin Amyloidosis. N. Engl. J. Med. 2018, 379, 11–21. [Google Scholar] [CrossRef]
- Finkel, R.S.; Chiriboga, C.A.; Vajsar, J.; Day, J.W.; Montes, J.; De Vivo, D.C.; Yamashita, M.; Rigo, F.; Hung, G.; Schneider, E.; et al. Treatment of infantile-onset spinal muscular atrophy with nusinersen: A phase 2, open-label, dose-escalation study. Lancet 2016, 388, 3017–3026. [Google Scholar] [CrossRef]
- Sardh, E.; Harper, P.; Balwani, M.; Stein, P.; Rees, D.; Bissell, D.M.; Desnick, R.; Parker, C.; Phillips, J.; Bonkovsky, H.L.; et al. Phase 1 Trial of an RNA Interference Therapy for Acute Intermittent Porphyria. N. Engl. J. Med. 2019, 380, 549–558. [Google Scholar] [CrossRef]
- Yang, J. Patisiran for the treatment of hereditary transthyretin-mediated amyloidosis. Expert Rev. Clin. Pharmacol. 2019, 12, 95–99. [Google Scholar] [CrossRef]
- Hüser, L.; Novak, D.; Umansky, V.; Altevogt, P.; Utikal, J. Targeting SOX2 in anticancer therapy. Expert Opin. Ther. Targets 2018, 22, 983–991. [Google Scholar] [CrossRef]
- Chen, Y.; Wu, J.J.; Lin, X.B.; Bao, Y.; Chen, Z.H.; Zhang, C.R.; Cai, Z.; Zhou, J.Y.; Ding, M.H.; Wu, X.J.; et al. Differential lncRNA expression profiles in recurrent gliomas compared with primary gliomas identified by microarray analysis. Int. J. Clin. Exp. Med. 2015, 8, 5033–5043. [Google Scholar]
- Nandeesh, B.N.; Naskar, S.; Shashtri, A.H.; Arivazhagan, A.; Santosh, V. Recurrent Glioblastomas Exhibit Higher Expression of Biomarkers with Stem-like Properties. J. Neurosci. Rural Pract. 2018, 9, 86–91. [Google Scholar] [CrossRef] [Green Version]





| LncRNA | SOX Targets | Mechanism of Action | Glioma Cells Properties Affected by Modulation of SOX Expression | Reference |
|---|---|---|---|---|
| lncRNA with tumor suppressor roles | ||||
| PR-LncRNA | SOX1 SOX2 SOX9 | decreases expression of SOX1, SOX2, and SOX9 | decreases the oncosphere formation ability and self-renewal potential of GSCs | [74] |
| NBAT1 | SOX7 | NBAT1/miR-21/SOX7 axis | inhibits proliferation, migration, and invasion of GBM cells | [75] |
| lncRNA with oncogenic roles | ||||
| MALAT1 | SOX2 | MALAT1/miR-129/SOX2 axis | enhances viability and proliferation of GSCs | [76] |
| NEAT1 | SOX2 | NEAT1/miR-132/SOX2 axis | promotes viability, migration, and invasion of GBM cells | [77] |
| SOX5 | NEAT1/miR-181d-5p/SOX5 axis that reduces expression of tight junction proteins ZO-1, occludin, and claudin-5 | impaired BTB permeability in GECs | [78] | |
| TUG1 | SOX2 | TUG1/miR-145/SOX2 axis | maintains the stemness and tumorigenicity of GSCs | [79] |
| TALNEC2 | SOX2 | TALNEC2/miR-21/SOX2 axis TALNEC2/miR-191/SOX2 axis | promotes mesenchymal transformation and stemness and decreases radiosensitivity of GSCs | [80] |
| PVT1 | SOX2 | PVT1/miR-365/ELF4/SOX2 axis | promotes proliferation, migration, invasion, and temozolomide (TMZ) resistance of GBM cells | [81] |
| HOXA11-AS | SOX2 SOX17 | increases expression of SOX2 and SOX17 | promotes stemness of GBM cells and glioma progression in vivo | [82] |
| H19 | SOX2 | increases expression of SOX2 | promotes proliferation, migration, stemness, and TMZ resistance in GBM cells | [83] |
| SOX4 | H19/miR-130a-3p/SOX4 axis | promotes migration, invasion, and neurosphere formation, and facilitates EMT | [84] | |
| CASCADES | SOX2 | SOX2 super-enhancer associated lncRNA that modulates activity of SOX2 in a positive feedback loop | promotes stemness of GSCs | [85] |
| XIST | SOX4 | XIST/miR-133a/SOX4 axis | promotes proliferation, invasion, migration, and EMT of GBM cells | [86] |
| HNF1A-AS1 | SOX4 | HNF1A-AS1/miR-32-5p/SOX4 axis | promotes proliferation, migration, and invasion, and inhibits apoptosis of GBM cells | [87] |
| SNHG9 | SOX9 | SNHG9/miR-326/SOX9 axis | promotes the growth of GSCs | [88] |
| SNHG12 | SOX5 | TDP43/SNHG12/miR-195/SOX5 axis that promotes expression of oncogene Gelsolin SOX5 activates SNHG12 forming a feedback loop | promotes proliferation, migration, and invasion and inhibits apoptosis of GBM cells | [89] |
| SNHG15 | SOX2 | SNHG15/miR-627-5p/SOX2 axis | promotes GBM tumorigenesis, decreases sensitivity towards TMZ treatment | [90] |
| AB073614 | SOX7 | AB073614 induces Wnt/β-catenin signaling activity by downregulation of SOX7 expression | promotes proliferation, migration, and invasion of GBM cells | [91] |
| LINC00174 | SOX9 | LINC00174/miR-138-5p/SOX9 axis | promotes proliferation, cell cycle progression, and increases chemoresistance to TMZ in GBM cells | [5] |
| DUXAP10 | SOX12 SOX2 | recruiting HuR to the cytoplasm enhancing SOX12 mRNA stability increases expression of SOX2 | promotes stemness of GBM cells facilitates growth of GSCs | [92] |
| SOX2OT | SOX2 | SOX2OT binds RNA demethylase ALKBH5 which regulates SOX2 expression via RNA demethylation | inhibits cell apoptosis, promotes cell proliferation, and TMZ resistance | [7] |
| SOX2OT/SOX2/Wnt5a/β-catenin axis | [7] | |||
| SOX2OT/miR-194-5p/SOX3 axis | ||||
| SOX3 | SOX2OT/miR-122/SOX3 axis | inhibits proliferation, migration, and invasion of GSCs, and promotes apoptosis of GSCs | [93] | |
| Tumor Cells | Signaling Axis | Cell Properties Affected by SOX2OT | Reference |
|---|---|---|---|
| breast cancer cells | SOX2OT/SOX2 | Ectopic expression of SOX2OT reduces proliferation and increases breast cancer cell anchorage-independent growth | [114] |
| pancreatic ductal adenocarcinoma cells | SOX2OT/miR-200/SOX2 | SOX2OT promotes EMT, stem cell-like properties, invasion and metastasis | [6] |
| YY1/SOX2OT/SOX2 | Ectopic expression of SOX2OT promotes cell proliferation and colony formation capacity | [115] | |
| cholangiocarcinoma cells | IRF4/SOX2OT/SOX2/PI3K/AKT | SOX2OT overexpression increases cell proliferation, decreases apoptosis rate and enhances migratory and invasion abilities in vitro, and metastatic ability in vivo | [116] |
| osteosarcoma cells | SOX2OT/SOX2 | SOX2OT overexpression promotes cell proliferation, increases colony formation ability, elevates migration, and invasion capabilities, and increases the expression of cancer stem cell biomarkers | [117] |
| cervical cancer cells | SOX2OT/SOX2 | SOX2OT knockdown suppresses cell viability, arrests cell cycle and ameliorates migration and invasion abilities of cells | [118] |
| bladder cancer cells | SOX2OT/miR-200c/SOX2 | SOX2OT knockdown inhibits the stemness phenotype (self-renewal, migration, invasion, and tumorigenicity) of bladder cancer stem cells | [119] |
| colorectal cancer cells | SOX2OT/miR-194-5p/SOX5 | SOX2OT silencing suppresses cell proliferation, migration, and invasion in vitro, and inhibits tumorigenesis in vivo | [120] |
| esophageal squamous cell carcinoma cells | SOX2/SOX2OT | SOX2OT overexpression promotes cell growth | [121] |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Stevanovic, M.; Kovacevic-Grujicic, N.; Petrovic, I.; Drakulic, D.; Milivojevic, M.; Mojsin, M. Crosstalk between SOX Genes and Long Non-Coding RNAs in Glioblastoma. Int. J. Mol. Sci. 2023, 24, 6392. https://doi.org/10.3390/ijms24076392
Stevanovic M, Kovacevic-Grujicic N, Petrovic I, Drakulic D, Milivojevic M, Mojsin M. Crosstalk between SOX Genes and Long Non-Coding RNAs in Glioblastoma. International Journal of Molecular Sciences. 2023; 24(7):6392. https://doi.org/10.3390/ijms24076392
Chicago/Turabian StyleStevanovic, Milena, Natasa Kovacevic-Grujicic, Isidora Petrovic, Danijela Drakulic, Milena Milivojevic, and Marija Mojsin. 2023. "Crosstalk between SOX Genes and Long Non-Coding RNAs in Glioblastoma" International Journal of Molecular Sciences 24, no. 7: 6392. https://doi.org/10.3390/ijms24076392