
Determination of Selected Isoquinoline Alkaloids from Chelidonium majus, Mahonia aquifolium and Sanguinaria canadensis Extracts by Liquid Chromatography and Their In Vitro and In Vivo Cytotoxic Activity against Human Cancer Cells
 , , , , , , ,
, , , , , , ,  and
and 
Abstract
:1. Introduction
2. Results and Discussion
2.1. Determination of the Alkaloid Contents in Plant Extracts by HPLC-DAD
2.2. LC-MS/MS
2.3. Investigation of the In Vitro Cytotoxic Activity of Plant Extracts against MCF-7, SCC-25, MDA-MB-231 and FaDu Cells
2.4. Investigation of In Vitro Anticancer Activity of Chelidonium majus Extracts against A375, SK-MEL-3 and G361 Cells
2.5. Investigation of In Vitro Anticancer Activity of Chelidonium majus Extracts by BrdU Incorporation
2.6. Correlation of Some Alkaloid Contents with the Cytotoxic Activity of Chelidonium majus Extracts
2.7. Comparison of the In Vitro Cytotoxic Activity of Chelidonium majus Extracts with Cytotoxic Activity of Anticancer Drugs
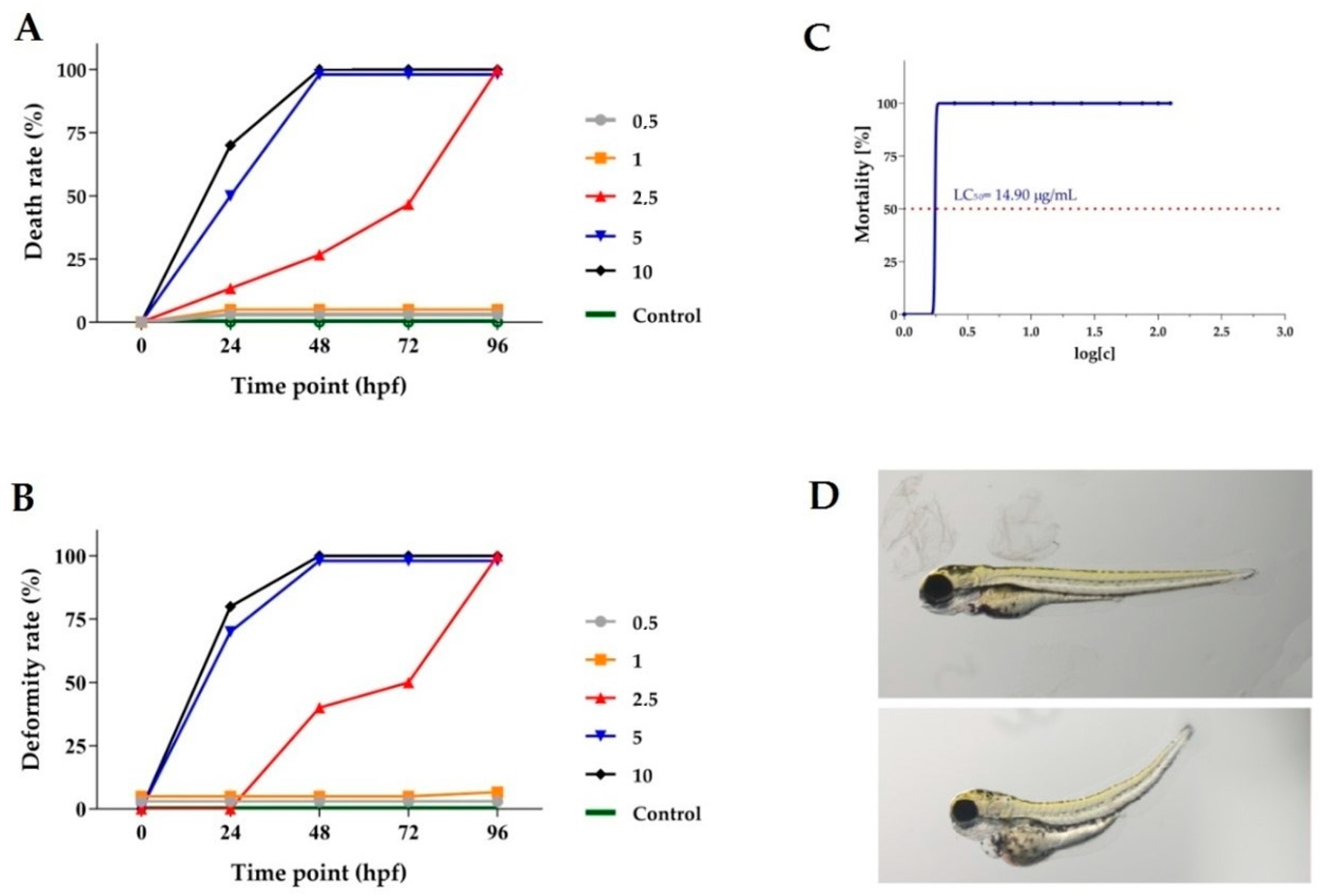
2.8. In Vivo Investigations of Toxicity of Chelidonium majus Root Extract
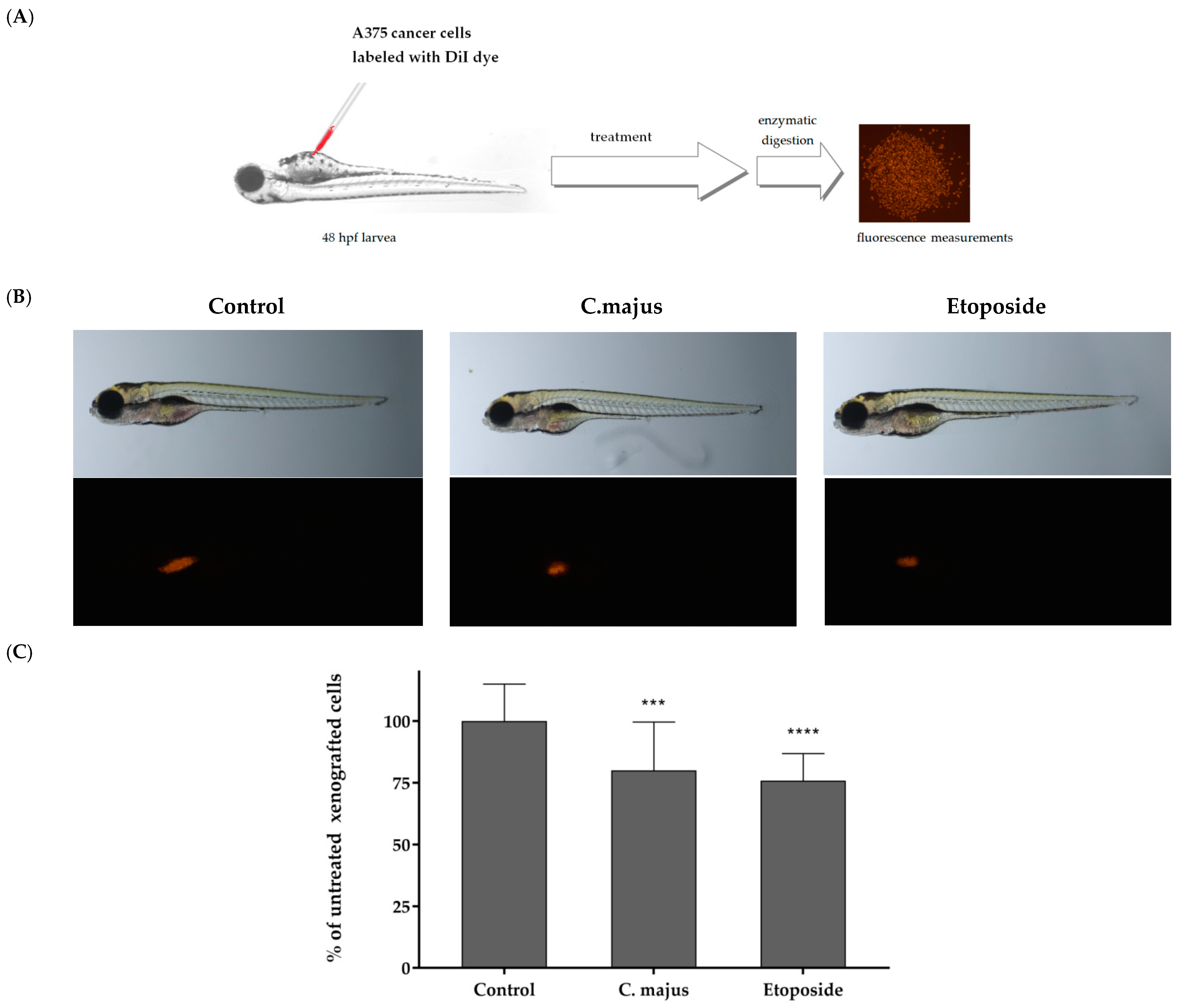
2.9. In Vivo Investigations of Antitumor Activity of the Chelidonium majus Root Extract
3. Materials and Methods
3.1. Experimental Procedure
3.1.1. Chemicals and Plant Materials
3.1.2. Apparatus and HPLC-DAD Conditions
3.1.3. HPLC-MS/MS
3.1.4. Extraction Procedure
3.1.5. Investigation of Cytotoxic Activity
Cell Cultivation
MTT Assay
BrdU Assay
3.1.6. Danio Rerio Culture and Fish Embryo Toxicity Test (FET)
3.1.7. Danio Rerio Human Tumor Cell Xenograft
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Hammerova, J.; Uldrijan, S.; Taborska, E.; Slaninova, I. Benzo[c]phenanthridine alkaloids exhibit strong anti-proliferative activity in malignant melanoma cells regardless of their p53 status. J. Dermatol. Sci. 2011, 62, 22–35. [Google Scholar] [CrossRef] [PubMed]
- Yun, D.; Yoon, S.Y.; Park, S.J.; Park, Y.J. The Anticancer Effect of Natural Plant Alkaloid Isoquinolines. Int. J. Mol. Sci. 2021, 22, 1653. [Google Scholar] [CrossRef] [PubMed]
- Warowicka, A.; Popenda, Ł.; Bartkowiak, G.; Musidlak, O.; Litowczenko-Cybulska, J.; Kuźma, D.; Nawrot, R.; Jurga, S.; Goździcka-Józefiak, A. Protoberberine compounds extracted from Chelidonium majus L. as novel natural photosensitizers for cancer therapy. Phytomedicine 2019, 64, 152919. [Google Scholar] [CrossRef] [PubMed]
- Kulp, M.; Bragina, O. Capillary electrophoretic study of the synergistic biological effects of alkaloids from Chelidonium majus L. in normal and cancer cells. Anal. Bioanal. Chem. 2013, 405, 3391–3397. [Google Scholar] [CrossRef] [PubMed]
- El-Readi, M.Z.; Eid, S.Y.; Ashour, M.L.; Tahrani, A.; Wink, M. Modulation of multidrug resistance in cancer cells by chelidonine and Chelidonium majus alkaloids. Phytomedicine 2013, 20, 282–294. [Google Scholar] [CrossRef]
- Wu, C.; Wang, X.; Xu, M.; Liu, Y.; Di, X. Intracellular Accumulation as an Indicator of Cytotoxicity to Screen Hepatotoxic Components of Chelidonium majus L. by LC–MS/MS. Molecules 2019, 24, 2410. [Google Scholar] [CrossRef] [Green Version]
- Capistrano, I.R.; Wouters, A.; Lardon, F.; Gravekamp, C.; Apers, S.; Pieters, L. In Vitro and in vivo investigations on the antitumour activity of Chelidonium majus. Phytomedicine 2015, 22, 1279–1287. [Google Scholar] [CrossRef]
- Gu, Y.; Qian, D.; Duan, J.-a.; Wang, Z.; Guo, J.; Tang, Y.; Guo, S. Simultaneous determination of seven main alkaloids of Chelidonium majus L. by ultraperformance LC with photodiode-array detection. J. Sep. Sci. 2010, 33, 1004–1009. [Google Scholar] [CrossRef]
- Huang, Y.; Wang, T.; Yin, G.; Wang, J.; Jiang, K.; Tu, J. High-performance liquid chromatography–based fingerprint analysis with chemical pattern recognition for evaluation of Mahonia bealei (Fort.). Carr. J. Sep. Sci. 2020, 43, 3625–3635. [Google Scholar] [CrossRef]
- Wang, W.; Ma, X.; Guo, X.; Zhao, M.; Tu, P.; Jiang, Y. A series of strategies for solving the shortage of reference standardsfor multi-components determination of traditional Chinese medicine, Mahoniae Caulis as a case. J. Chromatogr. A 2015, 1412, 100–111. [Google Scholar] [CrossRef]
- Singh, A.; Bajpai, V.; Kumar, S.; Rawat, A.K.S.; Kumar, B. Analysis of isoquinoline alkaloids from Mahonia leschenaultia and Mahonia napaulensis roots using UHPLC-Orbitrap-MSn and UHPLC-QqQLIT-MS/MS. J. Pharm. Anal. 2017, 7, 77–86. [Google Scholar] [CrossRef]
- Park, S.-W.; Kim, S.R.; Kim, Y.; Lee, J.-H.; Woo, H.-J.; Yoon, Y.-K.; Kim, Y.I. Chelidonium majus L. extract induces apoptosis through caspase activity via MAPK-independent NF-κB signalling in human epidermoid carcinoma A431 cells. Oncol. Rep. 2015, 33, 419–424. [Google Scholar] [CrossRef] [Green Version]
- Havelek, R.; Seifrtova, M.; Kralovec, K.; Krocova, E.; Tejkalova, V.; Novotny, I.; Cahlikova, L.; Safratova, M.; Opletal, L.; Bilkova, Z.; et al. Comparative cytotoxicity of chelidonine and homochelidonine, the dimethoxy analogues isolated from Chelidonium majus L. (Papaveraceae), against human leukemic and lung carcinoma cells. Phytomedicine 2016, 23, 253–266. [Google Scholar] [CrossRef]
- Zhang, Z.H.; Mi, C.; Wang, K.S.; Wang, Z.; Li, M.Y.; Zuo, H.X.; Xu, G.H.; Li, X.; Piao, L.X.; Ma, J.; et al. Chelidonine inhibits TNF-α-induced inflammation by suppressing the NF-κB pathways in HCT116 cells. Phytother. Res. 2018, 32, 65–75. [Google Scholar] [CrossRef]
- Singh, T.; Vaid, M.; Katiyar, N.; Sharma, S.; Katiyar, S.K. Berberine, an isoquinoline alkaloid, inhibits melanoma cancer cell migration by reducing the expressions of cyclooxygenase-2, prostaglandin E2 and prostaglandin E2 receptors. Carcinogenesis 2011, 32, 86–92. [Google Scholar] [CrossRef] [Green Version]
- Liu, J.-F.; Lai, K.C.; Peng, S.-F.; Maraming, P.; Huang, Y.-P.; Huang, A.-C.; Chueh, F.-S.; Huang, W.-W.; Chung, J.-G. Berberine Inhibits Human Melanoma A375.S2 Cell Migration and Invasion via Affecting the FAK, uPA, and NF-B Signaling Pathways and Inhibits PLX4032 Resistant A375.S2 Cell Migration In Vitro. Molecules 2018, 23, 2019. [Google Scholar] [CrossRef] [Green Version]
- Ren, M.; Yang, L.; Li, D.; Yang, L.; Su, Y.; Su, X. Cell Cycle Regulation by Berberine in Human Melanoma A375 Cells. Bull. Exp. Biol. Med. 2020, 169, 491–496. [Google Scholar] [CrossRef]
- Kim, J.-H.; Ryu, A.-R.; Kang, M.-J.; Lee, M.-Y. Berberine-induced changes in protein expression and antioxidant enzymes in melanoma cells. Mol. Cell. Toxicol. 2016, 12, 53–61. [Google Scholar] [CrossRef]
- Kou, Y.; Li, L.; Li, H.; Tan, Y.; Li, B.; Wang, K.; Du, B. Berberine suppressed epithelial mesenchymal transition through cross-talk regulation of PI3K/AKT and RARa/RARb in melanoma cells. Biochem. Biophys. Res. Commun. 2016, 479, 290–296. [Google Scholar] [CrossRef] [Green Version]
- He, J.M.; Mu, Q. The medicinal uses of the genus Mahonia in traditional Chinese medicine: An ethnopharmacological, phytochemical and pharmacological review. J. Ethnopharmacol. 2015, 175, 668–683. [Google Scholar] [CrossRef]
- Godevac, D.; Damjanovic, A.; Stanojkovic, T.P.; Anđelkovic, B.; Zdunic, G. Identification of cytotoxic metabolites from Mahonia aquifolium using1H NMR-based metabolomics approach. J. Pharm. Biomed. Anal. 2018, 150, 9–14. [Google Scholar] [CrossRef] [PubMed]
- Rezadoost, M.H.; Kumleh, H.H.; Ghasempour, A. Cytotoxicity and apoptosis induction in breast cancer, skin cancer and glioblastoma cells by plant extracts. Mol. Biol. Rep. 2019, 46, 5131–5142. [Google Scholar] [CrossRef] [PubMed]
- Damjanovic, A.; Kolundžija, B.; Matic, I.Z.; Krivokuca, A.; Zdunic, G.; Šavikin, K.; Jankovic, R.; Stankovic, J.A.; Stanojkovic, T.P. Mahonia aquifolium Extracts Promote Doxorubicin Effects against Lung Adenocarcinoma Cells In Vitro. Molecules 2020, 25, 5233. [Google Scholar] [CrossRef] [PubMed]
- Andreicuț, A.D.; Fischer-Fodor, E.; APârvu, E.; Ţigu, A.B.; Cenariu, M.; Pârvu, M.; Cătoi, F.A.; Irimie, A. Antitumoral and Immunomodulatory Effect of Mahonia aquifolium Extracts. Oxid. Med. Cell. Longev. 2019, 2019, 6439021. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ma, W.-K.; Li, H.; Dong, C.-L.; He, X.; Guo, C.-R.; Zhang, C.-F.; Yu, C.-H.; Wang, C.-Z.; Yuan, C.-S. Palmatine from Mahonia bealei attenuates gut tumorigenesis in ApcMin/+ mice via inhibition of inflammatory cytokines. Mol. Med. Rep. 2016, 14, 491–498. [Google Scholar] [CrossRef] [Green Version]
- Wong, B.-S.; Hsiao, Y.-C.; Lin, T.-W.; Chen, K.-S.; Chen, P.-N.; Kuo, W.-H.; Chu, S.-C.; Hsieh, Y.-S. The in vitro and in vivo apoptotic effects of Mahonia oiwakensis on human lung cancer cells. Chem. Biol. Interact. 2009, 180, 165–174. [Google Scholar] [CrossRef]
- Senchina, D.S.; Flinn, G.N.; McCann, D.A.; Kohut, M.L.; Shearn, C.T. Bloodroot (Sanguinaria canadensis L., Papaveraceae) Enhances Proliferation and Cytokine Production by Human Peripheral Blood Mononuclear Cells in an In Vitro Model. J. Herbs Spices Med. Plants 2009, 15, 45–65. [Google Scholar] [CrossRef] [Green Version]
- Tuzimski, T.; Petruczynik, A.; Plech, T.; Kaproń, B.; Makuch-Kocka, A.; Szultka-Młyńska, M.; Misiurek, J.; Buszewski, B. Determination of Cytotoxic Activity of Sanguinaria Canadensis Extracts against Human Melanoma Cells and Comparison of Their Cytotoxicity with Cytotoxicity of Some Anticancer Drugs. Molecules 2021, 26, 1738. [Google Scholar] [CrossRef]
- Petruczynik, A.; Plech, T.; Tuzimski, T.; Misiurek, J.; Kaproń, B.; Misiurek, D.; Szultka-Młyńska, M.; Buszewski, B.; Waksmundzka-Hajnos, M. Determination of Selected Isoquinoline Alkaloids from Mahonia aquifolia; Meconopsis cambrica; Corydalis lutea; Dicentra spectabilis; Fumaria offcinalis; Macleaya cordata Extracts by HPLC-DAD and Comparison of Their Cytotoxic Activity. Toxins 2019, 11, 575. [Google Scholar] [CrossRef] [Green Version]
- Kulp, M.; Bragina, O.; Kogerman, P.; Kaljurand, M. Capillary electrophoresis with led-induced native fluorescence detection for determination of isoquinoline alkaloids and their cytotoxicity in extracts of Chelidonium majus L. J. Chromatogr. A 2011, 1218, 5298–5304. [Google Scholar] [CrossRef]
- Campbell, S.; Affolter, J.; Randle, W. Spatial and temporal distribution of the alkaloid sanguinarine in Sanguinaria canadensis L. (Bloodroot). Econ. Bot. 2007, 61, 223–234. [Google Scholar] [CrossRef]
- Petruczynik, A.; Tuzimski, T.; Plech, T.; Misiurek, J.; Szalast, K.; Szymczak, G. Comparison of Anticancer Activity and HPLC-DAD Determination of Selected Isoquinoline Alkaloids from Thalictrum foetidum, Berberis sp. and Chelidonium majus Extracts. Molecules 2019, 24, 3417. [Google Scholar] [CrossRef] [Green Version]
- Bresciani, E.; Broadbridge, E.; Liu, P.P. An efficient dissociation protocol for generation of single cell suspension from zebrafish embryos and larvae. Methods X 2018, 5, 1287–1290. [Google Scholar] [CrossRef]



{kind=link}
{kind=link}
| Plant Extract | Berberine | Chelerythrine | Chelidonine | Magnoflorine | Palmatine | Protopine | Sanguinarine |
|---|---|---|---|---|---|---|---|
| Chelidonium majus herb before flowering | 0.168 | 0.024 | 0.891 | - | - | 0.984 | 0.075 |
| Chelidonium majus root before flowering | 0.184 | 1.105 | 1.341 | - | - | 0.358 | 0.344 |
| Chelidonium majus herb during flowering | 0.120 | 0.037 | 0.317 | - | - | 0.690 | 0.081 |
| Chelidonium majus root during flowering | 0.023 | 1.182 | 0.967 | - | - | 0.559 | 0.638 |
| Chelidonium majus herb after flowering | 0.148 | 0.130 | 0.479 | - | - | 1.227 | 0.128 |
| Chelidonium majus root after flowering | 0.011 | 1.744 | 1.490 | - | - | 0.308 | 0.821 |
| Chelidonium majus seeds after flowering | 0.017 | 0.002 | 0.069 | - | - | 0.008 | 0.003 |
| Chelidonium majus pods after flowering | 0.174 | 0.245 | 0.545 | - | - | 0.133 | 0.545 |
| Mahonia aquifolium cortex before flowering | 0.135 | - | - | 0.087 | 0.039 | - | - |
| Mahonia aquifolium cortex during flowering | 3.316 | - | - | - | 0.794 | - | - |
| Mahonia aquifolium cortex after flowering | 0.067 | - | - | 0.102 | 0.793 | - | - |
| Mahonia aquifolium leaves before flowering | - | - | - | 0.324 | - | - | - |
| Mahonia aquifolium leaves during flowering | 0.0220 | - | - | 0.238 | <LOQ | - | - |
| Mahonia aquifolium leaves after flowering | <LOQ | - | - | 0.319 | - | - | - |
| Mahonia aquifolium barked stalk during flowering | 0.438 | - | - | - | 0.045 | - | - |
| Mahonia aquifolium barked stalk after flowering | 0.044 | - | - | 0.191 | 0.342 | - | - |
| Mahonia aquifolium root during flowering | 7.046 | - | - | - | 0.629 | - | - |
| Mahonia aquifolium root after flowering | 0.008 | - | - | - | 0.305 | - | - |
| Sanguinaria canadensis before flowering | 0.006 | 2.738 | - | - | <LOQ | - | 4.874 |
| Sanguinaria canadensis during flowering | 0.014 | 5.362 | <LOQ | - | - | 0.015 | 9.598 |
| Sanguinaria canadensis after flowering | 0.010 | 6.878 | - | - | - | 0.110 | 6.949 |
| Compound | HPLC-DAD | LC-MS | ||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|
| tR | As | Elemental Composition | tR (min) | Polarity | Theoretical (m/z) | Measured (m/z) | Major Fragment Ions | Error (ppm) | ID Score [%] | Collision Energy (eV) | Fragmentor Voltage | |
| Berberine | 19.70 | 1.05 | C20H18NO4 [M+H]+ | 3.12 | ESI+ | 335.7426 | 335.7429 | 319.7029 305.6823 304.1893 291.6987 277.6827 | −0.41 | 99.18 | 20 | 195 |
| Chelerythrine | 23.32 | 1.06 | C21H18NO4 [M+H]+ | 3.98 | ESI+ | 347.7491 | 347.7489 | 331.7071 303.6990 274.6920 231.6968 | 1.19 | 99.56 | 20 | 195 |
| Chelidonine | 10.91 | 1.07 | C20H20NO5 [M+H]+ | 1.94 | ESI+ | 354.3922 | 354.3920 | 336.6503 303.6990 274.6920 189.4355 | −1.17 | 99.90 | 20 | 195 |
| Magnoflorine | 3.94 | 1.08 | C20H24NO4 [M+H]+ | 3.27 | ESI+ | 341.7915 | 341.7917 | 296.7147 264.6899 236.7089 206.7327 | −0.39 | 99.48 | 20 | 195 |
| Palmatine | 16.96 | 1.18 | C21H22NO4 [M+H]+ | 2.53 | ESI+ | 351.7872 | 351.7853 | 335.7442 307.7351 277.6835 249.6977 | 1.07 | 99.62 | 20 | 195 |
| Protopine | 8.49 | 1.03 | C20H20NO5 [M+H]+ | 1.68 | ESI+ | 353.7653 | 353.7655 | 336.1209 274.6716 205.7380 188.7681 148.8711 | −0.93 | 99.15 | 20 | 195 |
| Sanguinarine | 20.37 | 1.06 | C20H14NO4 [M+H]+ | 2.75 | ESI+ | 331.7068 | 331.7065 | 316.6761 303.6993 288.6723 273.6853 245.7012 | 1.26 | 99.34 | 20 | 195 |
| Plant Extract | IC50 [μg/mL] ± SD | |||
|---|---|---|---|---|
| MCF-7 | MDA-MB-231 | SCC-25 | FaDu | |
| Chelidonium majus herb after flowering | 17.20 ± 2.07 | 25.31 ± 2.41 | 1.76 ± 0.29 | 3.68 ± 0.63 |
| Chelidonium majus root after flowering | 7.72 ± 0.28 | 17.01 ± 1.68 | 0.95 ± 0.02 | 2.06 ± 0.17 |
| Chelidonium majus seeds after flowering | >100 | >100 | 13.74 ± 1.17 | 18.38 ± 1.78 |
| Chelidonium majus pods after flowering | 8.26 ± 1.07 | >100 | 5.05 ± 0.36 | 3.94 ± 0.33 |
| Mahonia aquifolium root during flowering | 34.79 ± 3.29 | 24.00 ± 2.63 | 36.98 ± 4.28 | 10.85 ± 1.86 |
| Mahonia aquifolium barked stalk during flowering | 29.48 ± 2.38 | 89.70 ± 6.03 | 28.44 ± 1.36 | 5.73 ± 1.07 |
| Mahonia aquifalium leaves during flowering * | 89.14 ± 2.73 | 90.71 ± 7.29 | 97.25 ± 8.07 | 46.77 ± 7.84 |
| Mahonia aquifalium cortex during flowering * | 15.71 ± 1.92 | 31.87 ± 4.35 | 31.37 ± 2.29 | 7.67 ± 0.82 |
| Sanguinaria candensis before flowering | 1.48 ± 0.17 | 4.15 ± 0.39 | 0.90 ± 0.03 | 0.21 ± 0.02 |
| Sanguinaria candensis during flowering | 1.36 ± 0.11 | 4.26 ± 0.43 | 2.99 ± 0.23 | 0.27 ± 0.04 |
| Sanguinaria candensis after flowering | 3.14 ± 0.62 | 11.37 ± 0.77 | 1.05 ± 0.08 | 0.41 ± 0.09 |
| Etoposide | 136.48 ± 8.95 | 219.31 ± 24.47 | 223.94 ± 24.81 | 38.73 ± 1.56 |
| Plant Extract | IC50 [μg/mL] ± SD | ||||
|---|---|---|---|---|---|
| MCF-7 | MDA-MB-231 | SCC-25 | FaDu | Fibroblasts | |
| Chelidonium majus root 10.04.2018 | 5.46 ± 0.01 | 5.77 ± 0.37 | 1.26 ± 0.23 | 3.67 ± 0.21 | 0.62 ± 0.12 (0.96 ± 0.12) |
| Chelidonium majus root 10.05.2018 | 0.20 ± 0.03 (0.18 ± 0.02) | 0.32 ± 0.04 (0.24 ± 0.04) | 0.62 ± 0.10 (0.41 ± 0.03) | 1.96 ± 0.17 | 0.67 ± 0.06 (0.85 ± 0.05) |
| Chelidonium majus root 10.06.2018 | 0.17 ± 0.02 (0.11 ± 0.01) | 1.49 ± 0.36 | 0.83 ± 0.09 | 2.56 ± 0.47 | 0.76 ± 0.10 (0.68 ± 0.06) |
| Chelidonium majus herb 10.04.2018 | 28.23 ± 3.67 | 43.70 ± 4.79 | 75.62 ± 4.28 | 13.10 ± 1.50 (8.94 ± 0.47) | 16.58 ± 1.27 (17.52 ± 0.71) |
| Chelidonium majus herb 10.05.2018 | 28.03 ± 2.04 | 9.36 ± 0.77 | 32.52 ± 5.01 | 0.14 ± 0.05 (0.10 ± 0.01) | 0.56 ± 0.03 (0.85 ± 0.10) |
| Chelidonium Majus herb 10.06.2018 | 11.97 ± 0.30 (8.03 ± 0.37) | 24.68 ± 2.45 | 26.93 ± 3.12 | 18.34 ± 2.17 (14.28 ± 0.77) | 18.78 ± 0.58 (20.79 ± 1.04) |
| Plant Extract | IC50 [μg/mL] ± SD | ||
|---|---|---|---|
| A375 | G361 | SK-MEL-3 | |
| Chelidonium majus herb | 27.91 ± 2.76 (18.08 ± 1.32) | 2.27 ± 0.32 (4.16 ± 0.51) | 7.89 ± 0.57 (3.62 ± 0.13) |
| Chelidonium majus root | 12.65 ± 1.85 (7.24 ± 0.21) | 4.17 ± 0.29 (3.98 ± 0.14) | 1.93 ± 0.08 (1.17 ± 0.07) |
| Chelidonium majus seeds | >200 (142.65 ± 4.65) | 6.97 ± 0.75 (5.17 ± 0.24) | 33.33 ± 4.70 (19.73 ± 0.45) |
| Chelidonium majus pods with seeds | 180.16 ± 12.51 (120.47 ± 4.83) | 5.77 ± 0.43 (4.19 ± 0.18) | 19.85 ± 2.07 (11.05 ± 0.81) |
| Etoposide | 92.34 ± 4.58 (32.8 ± 2.38) | 52.32 ± 3.86 (18.01 ± 0.57) | >200 (>100) |
| Cisplatin | 10.62 ± 1.04 (6.02 ± 0.41) | 11.53 ± 1.46 (4.85 ±0.33) | 14.42 ± 1.61 (7.58 ± 0.73) |
| Hydroxyurea | >200 | >200 | >200 |
| Cell Line | Chelerythrine | Sanguinarine | Chelerythrine and sanguinarine | |||
|---|---|---|---|---|---|---|
| Equation | r | Equation | r | Equation | r | |
| MCF-7 | y = −15.72x + 23.26 | 0.8845 | y = −35.25x + 24.60 | 0.8608 | y = −11.02x + 23.83 | 0.8835 |
| MDA-MB-231 | y = −17.11x + 26.10 | 0.7391 | y = −37.98x + 27.43 | 0.7120 | y = −11.97x + 26.69 | 0.7360 |
| SCC-25 | y = −32.13x + 45.28 | 0.7964 | y = −69.04x + 46.98 | 0.7426 | y = −22.26x + 46.16 | 0.7856 |
| FaDu | y = −5.357x + 10.35 | 0.5340 | y = −11.71x + 10.70 | 0.5065 | y = −3.729x + 10.52 | 0.5293 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Tuzimski, T.; Petruczynik, A.; Plech, T.; Kaproń, B.; Makuch-Kocka, A.; Szultka-Młyńska, M.; Misiurek, J.; Buszewski, B.; Waksmundzka-Hajnos, M. Determination of Selected Isoquinoline Alkaloids from Chelidonium majus, Mahonia aquifolium and Sanguinaria canadensis Extracts by Liquid Chromatography and Their In Vitro and In Vivo Cytotoxic Activity against Human Cancer Cells. Int. J. Mol. Sci. 2023, 24, 6360. https://doi.org/10.3390/ijms24076360
Tuzimski T, Petruczynik A, Plech T, Kaproń B, Makuch-Kocka A, Szultka-Młyńska M, Misiurek J, Buszewski B, Waksmundzka-Hajnos M. Determination of Selected Isoquinoline Alkaloids from Chelidonium majus, Mahonia aquifolium and Sanguinaria canadensis Extracts by Liquid Chromatography and Their In Vitro and In Vivo Cytotoxic Activity against Human Cancer Cells. International Journal of Molecular Sciences. 2023; 24(7):6360. https://doi.org/10.3390/ijms24076360
Chicago/Turabian StyleTuzimski, Tomasz, Anna Petruczynik, Tomasz Plech, Barbara Kaproń, Anna Makuch-Kocka, Małgorzata Szultka-Młyńska, Justyna Misiurek, Bogusław Buszewski, and Monika Waksmundzka-Hajnos. 2023. "Determination of Selected Isoquinoline Alkaloids from Chelidonium majus, Mahonia aquifolium and Sanguinaria canadensis Extracts by Liquid Chromatography and Their In Vitro and In Vivo Cytotoxic Activity against Human Cancer Cells" International Journal of Molecular Sciences 24, no. 7: 6360. https://doi.org/10.3390/ijms24076360