
5-Aminolevrinic Acid Exhibits Dual Effects on Stemness in Human Sarcoma Cell Lines under Dark Conditions
 , , , ,
, , , ,
Abstract
:1. Introduction
2. Results
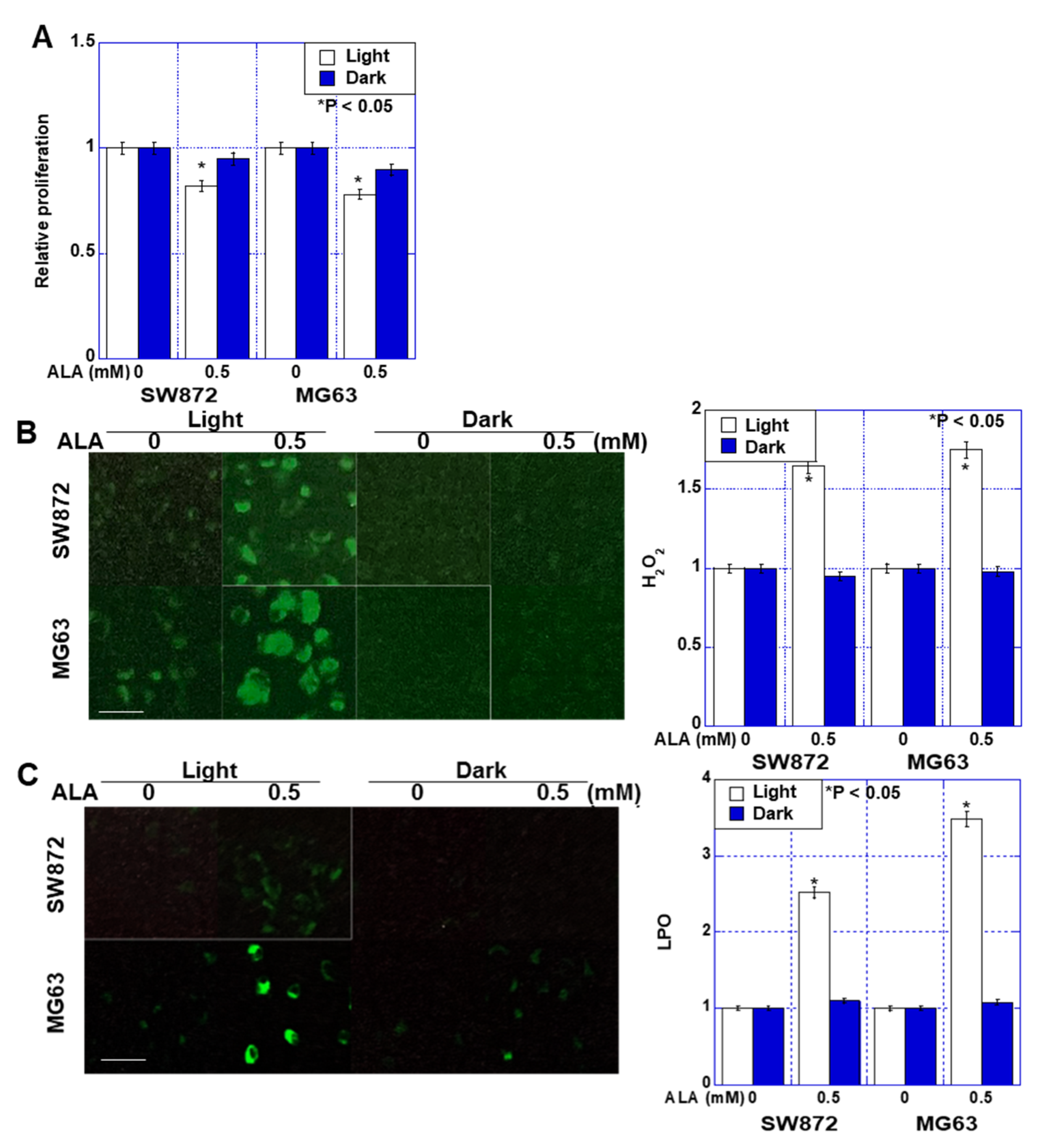
2.1. Effect of ALA without Light on Proliferation and Oxidative Stress in Sarcoma Cells
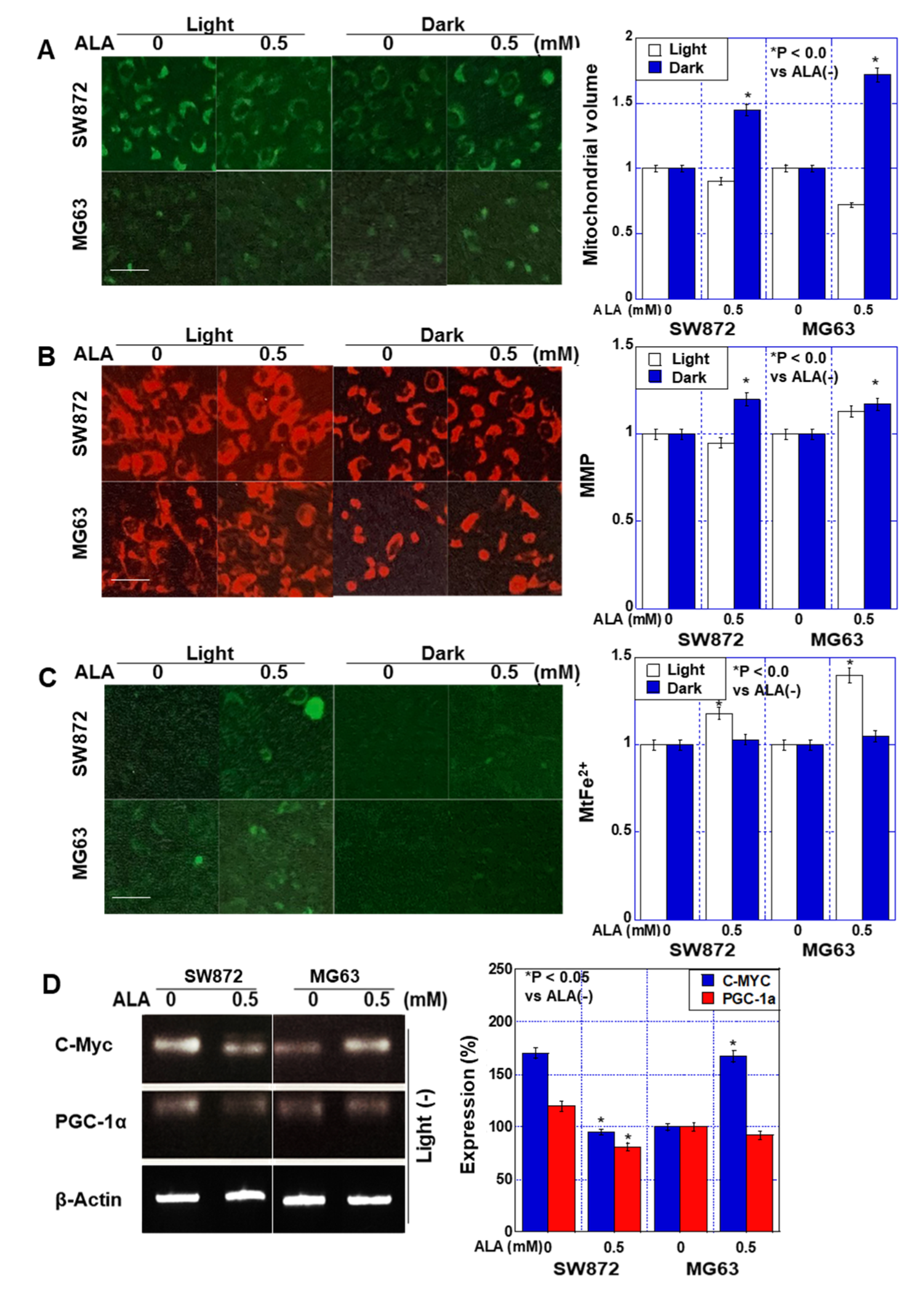
2.2. Effect of ALA without Light on Mitochondria in Sarcoma Cells
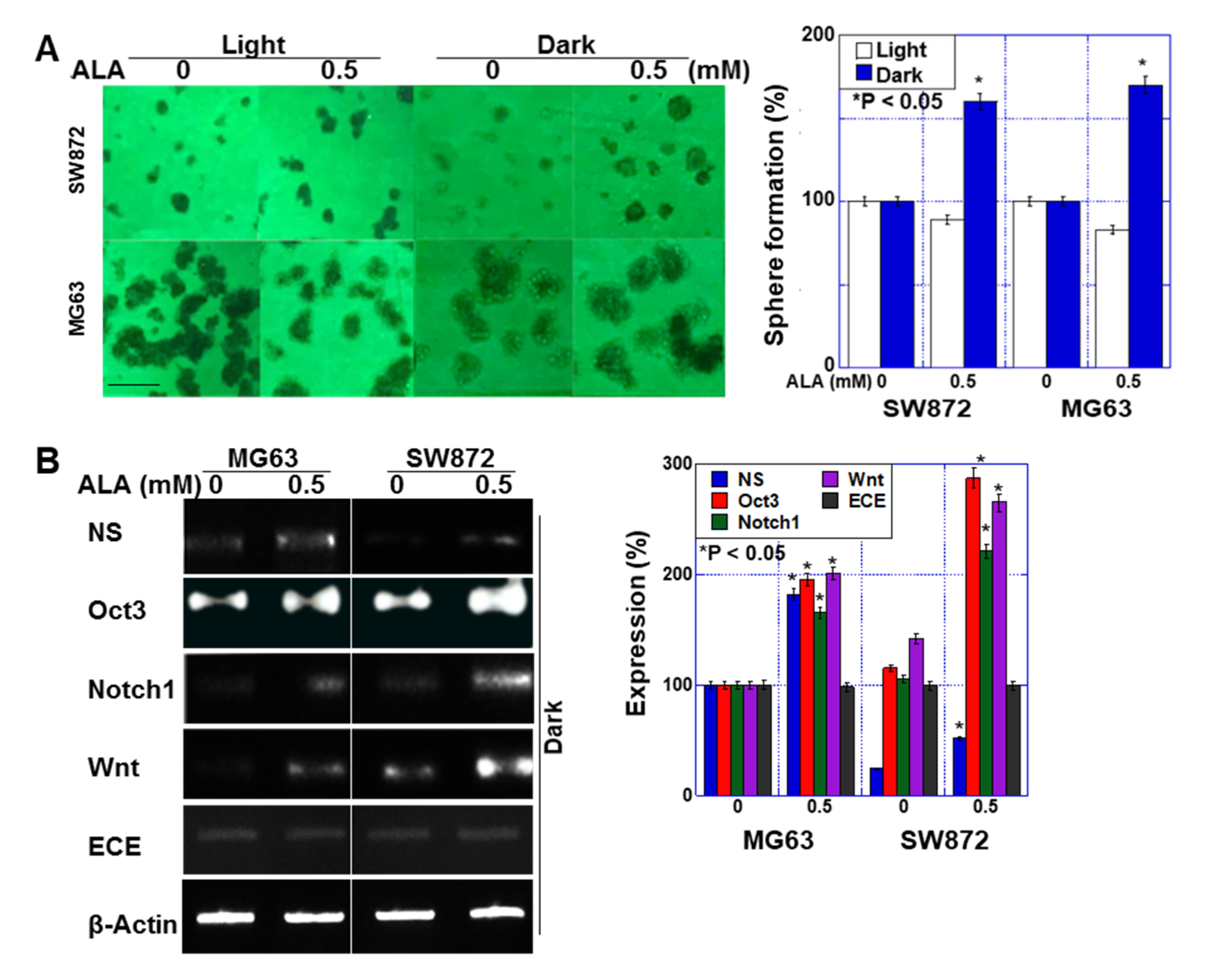
2.3. Effect of ALA without Light on Stemness in Sarcoma Cells
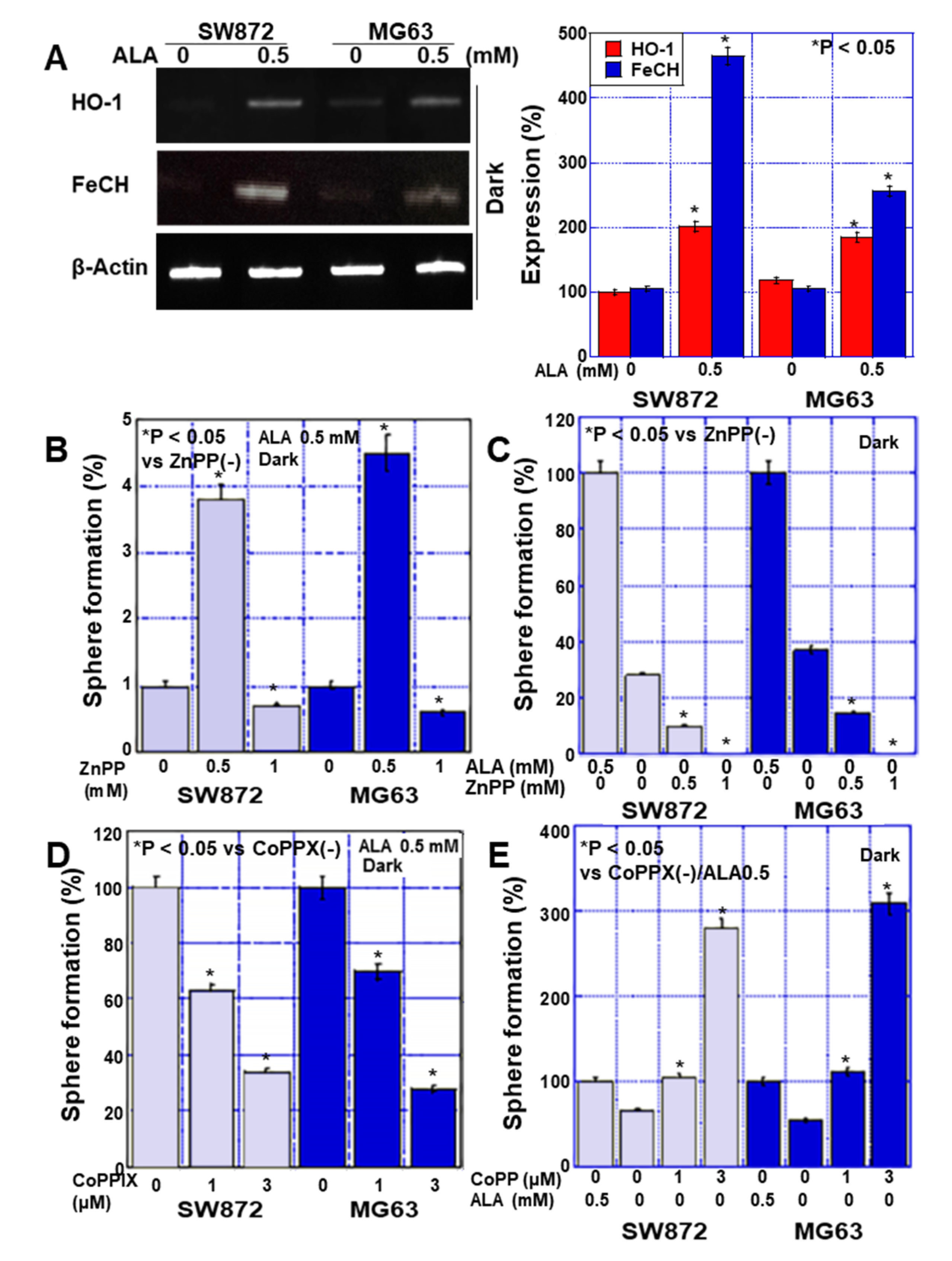
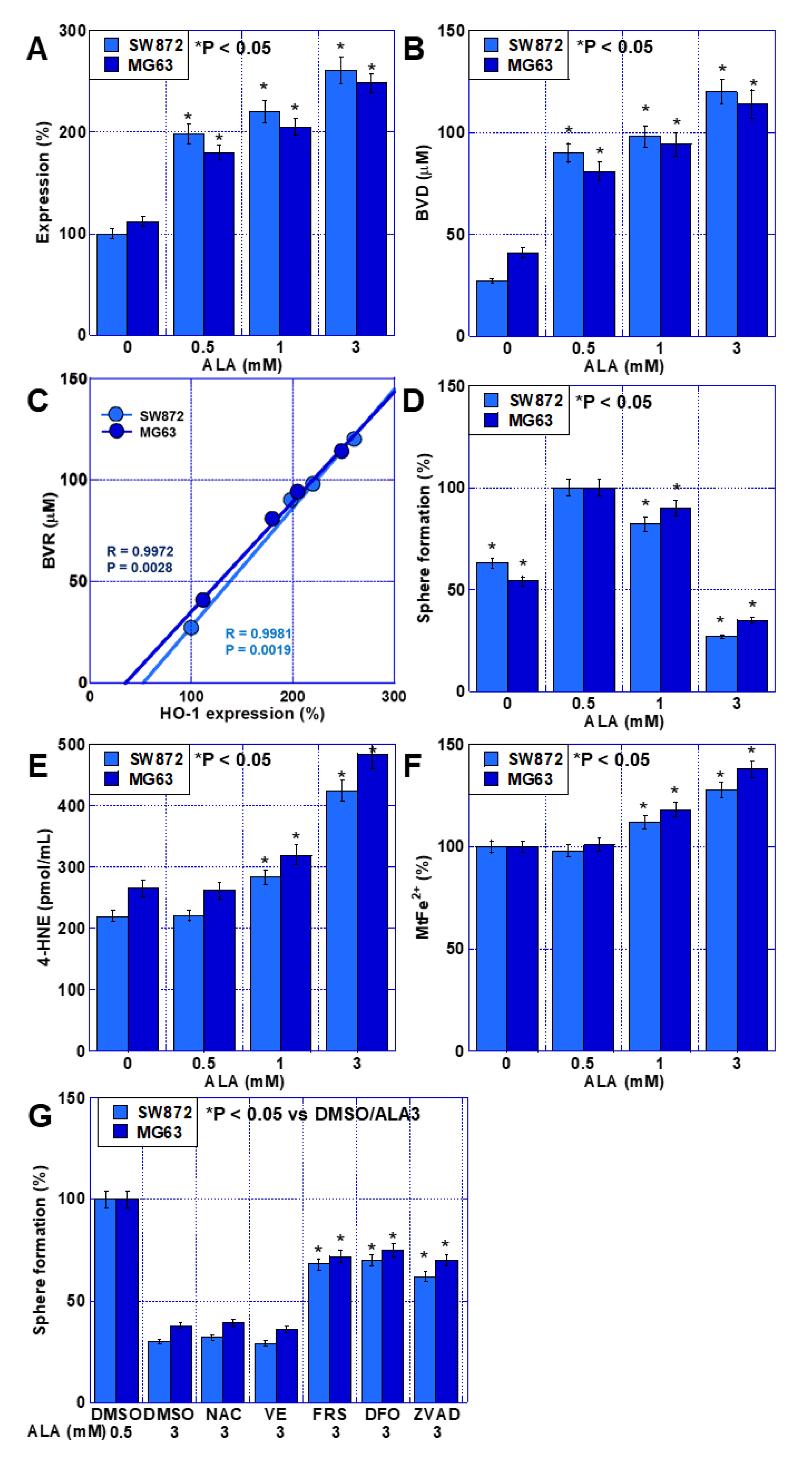
2.4. Effects of HO-1 on Stemness in Sarcoma Cells
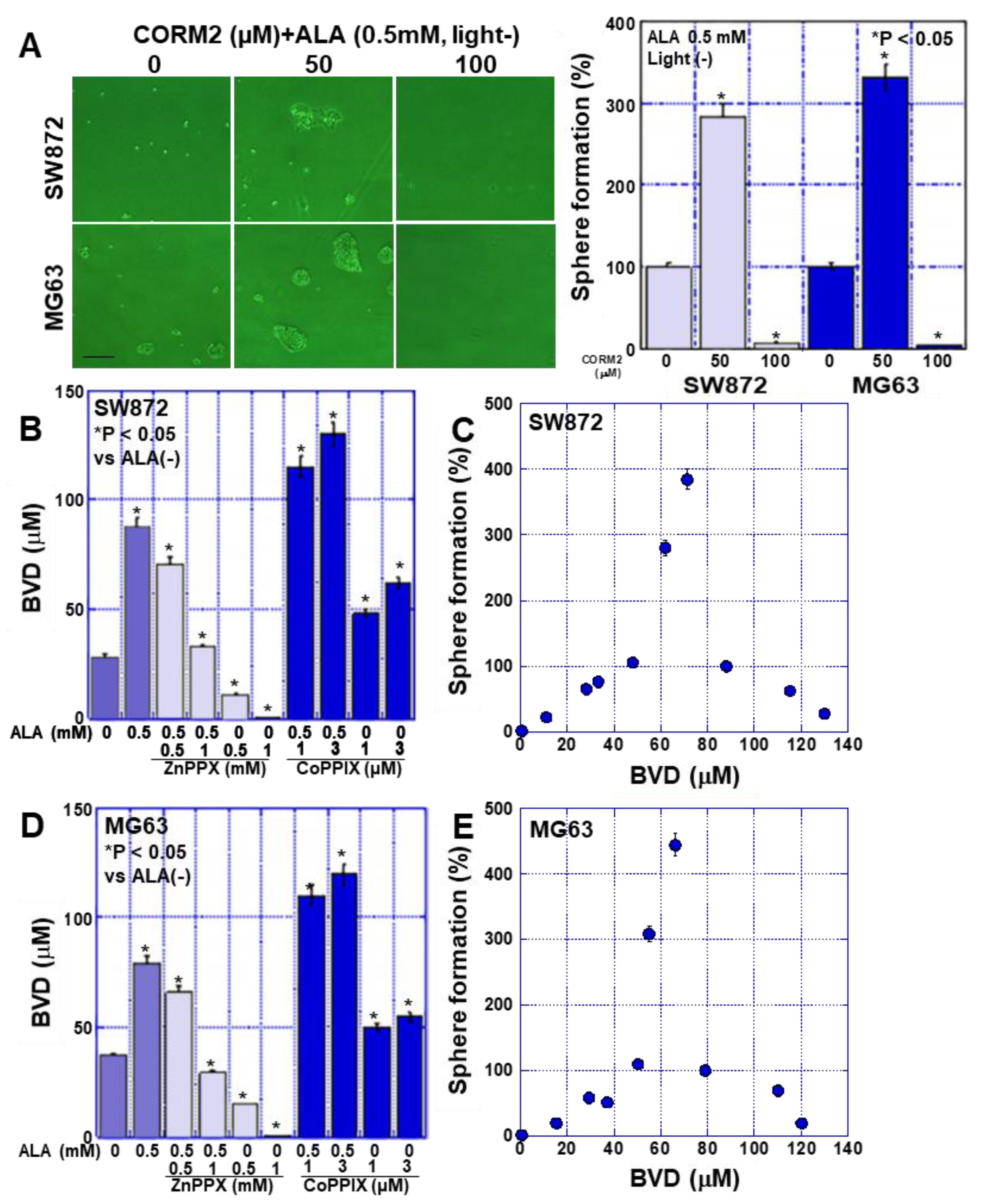
2.5. Effects of CO on Stemness in Sarcoma Cells
2.6. Effects of High ALA Concentration on Stemness in Sarcoma Cells
3. Discussion
4. Materials and Methods
4.1. Cell Culture
4.2. Reagents
4.3. Cell Proliferation
4.4. Mitochondrial Imaging
4.5. Sphere Formation Assay
4.6. RNA Isolation
4.7. Reverse-Transcription-Polymerase Chain Reaction (RT-PCR)
4.8. Enzyme-Linked Immunosorbent Assay (ELISA)
4.9. Quantification of BVD
4.10. Statistical Analysis
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Abbreviations
References
- Fuchs, J.; Weber, S.; Kaufmann, R. Genotoxic potential of porphyrin type photosensitizers with particular emphasis on 5-aminolevulinic acid: Implications for clinical photodynamic therapy. Free Radic. Biol. Med. 2000, 28, 537–548. [Google Scholar]
- Shinoda, Y.; Kato, D.; Ando, R.; Endo, H.; Takahashi, T.; Tsuneoka, Y.; Fujiwara, Y. Systematic Review and Meta-Analysis of In Vitro Anti-Human Cancer Experiments Investigating the Use of 5-Aminolevulinic Acid (5-ALA) for Photodynamic Therapy. Pharmaceuticals 2021, 14, 229. [Google Scholar]
- Owari, T.; Tanaka, N.; Nakai, Y.; Miyake, M.; Anai, S.; Kishi, S.; Mori, S.; Fujiwara-Tani, R.; Hojo, Y.; Mori, T.; et al. 5-Aminolevulinic acid overcomes hypoxia-induced radiation resistance by enhancing mitochondrial reactive oxygen species production in prostate cancer cells. Br. J. Cancer 2022, 127, 350–363. [Google Scholar]
- Jozkowicz, A.; Was, H.; Dulak, J. Heme oxygenase-1 in tumors: Is it a false friend? Antioxid. Redox Signal. 2007, 9, 2099–2117. [Google Scholar]
- Hjortsø, M.D.; Andersen, M.H. The expression, function and targeting of haem oxygenase-1 in cancer. Curr. Cancer Drug Targets 2014, 14, 337–347. [Google Scholar]
- Sasaki, T.; Fujii, K.; Yoshida, K.; Shimura, H.; Sasahira, T.; Ohmori, H.; Kuniyasu, H. Peritoneal metastasis inhibition by linoleic acid with activation of PPARgamma in human gastrointestinal cancer cells. Virchows Arch. 2006, 448, 422–427. [Google Scholar]
- Nitti, M.; Piras, S.; Marinari, U.M.; Moretta, L.; Pronzato, M.A.; Furfaro, A.L. HO-1 Induction in Cancer Progression: A Matter of Cell Adaptation. Antioxidants 2017, 6, 29. [Google Scholar]
- Chiang, S.K.; Chen, S.E.; Chang, L.C. A Dual Role of Heme Oxygenase-1 in Cancer Cells. Int. J. Mol. Sci. 2018, 20, 39. [Google Scholar]
- Hirai, K.; Sasahira, T.; Ohmori, H.; Fujii, K.; Kuniyasu, H. Inhibition of heme oxygenase-1 by zinc protoporphyrin IX reduces tumor growth of LL/2 lung cancer in C57BL mice. Int. J. Cancer 2007, 120, 500–505. [Google Scholar]
- Salerno, L.; Romeo, G.; Modica, M.N.; Amata, E.; Sorrenti, V.; Barbagallo, I.; Pittalà, V. Heme oxygenase-1: A new druggable target in the management of chronic and acute myeloid leukemia. Eur. J. Med. Chem. 2017, 142, 163–178. [Google Scholar]
- Kozakowska, M.; Szade, K.; Dulak, J.; Jozkowicz, A. Role of heme oxygenase-1 in postnatal differentiation of stem cells: A possible cross-talk with microRNAs. Antioxid. Redox Signal. 2014, 20, 1827–1850. [Google Scholar]
- Luu Hoang, K.N.; Anstee, J.E.; Arnold, J.N. The Diverse Roles of Heme Oxygenase-1 in Tumor Progression. Front. Immunol. 2021, 12, 658315. [Google Scholar]
- Ferrari, A.; Dirksen, U.; Bielack, S. Sarcomas of Soft Tissue and Bone. Prog. Tumor. Res. 2016, 43, 128–141. [Google Scholar]
- Shoaib, Z.; Fan, T.M.; Irudayaraj, J.M.K. Osteosarcoma mechanobiology and therapeutic targets. Br. J. Pharmacol. 2022, 179, 201–217. [Google Scholar]
- Fisher, S.B.; Roland, C.L. ASO Author Reflections: Location-Specific Staging in Sarcoma-A Step in the Right Direction. Ann. Surg. Oncol. 2018, 25 (Suppl. S3), 930–931. [Google Scholar]
- Honoki, K. Do stem-like cells play a role in drug resistance of sarcomas? Expert Rev. Anticancer. Ther. 2010, 10, 261–270. [Google Scholar]
- Hatina, J.; Kripnerova, M.; Houfkova, K.; Pesta, M.; Kuncova, J.; Sana, J.; Slaby, O.; Rodríguez, R. Sarcoma Stem Cell Heterogeneity. Adv. Exp. Med. Biol. 2019, 1123, 95–118. [Google Scholar]
- Sun, J.; Xing, F.; Braun, J.; Traub, F.; Rommens, P.M.; Xiang, Z.; Ritz, U. Progress of Phototherapy Applications in the Treatment of Bone Cancer. Int. J. Mol. Sci. 2021, 22, 11354. [Google Scholar]
- Yoon, Y.E.; Lee, K.S.; Lee, Y.J.; Lee, H.H.; Han, W.K. Renoprotective Effects of Carbon Monoxide-Releasing Molecule 3 in Ischemia-Reperfusion Injury and Cisplatin-Induced Toxicity. Transplant. Proc. 2017, 49, 1175–1182. [Google Scholar]
- Kim, D.H.; Yoon, H.J.; Cha, Y.N.; Surh, Y.J. Role of heme oxygenase-1 and its reaction product, carbon monoxide, in manifestation of breast cancer stem cell-like properties: Notch-1 as a putative target. Free Radic. Res. 2018, 52, 1336–1347. [Google Scholar]
- Tulsulkar, J.; Ward, A.; Shah, Z.A. HO1 and Wnt expression is independently regulated in female mice brains following permanent ischemic brain injury. Brain Res. 2017, 1662, 1–6. [Google Scholar]
- Ríos-Arrabal, S.; Puentes-Pardo, J.D.; Moreno-SanJuan, S.; Szuba, Á.; Casado, J.; García-Costela, M.; Escudero-Feliu, J.; Verbeni, M.; Cano, C.; González-Puga, C.; et al. Endothelin-1 as a Mediator of Heme Oxygenase-1-Induced Stemness in Colorectal Cancer: Influence of p53. J. Pers. Med. 2021, 11, 509. [Google Scholar]
- Ryter, S.W. Heme Oxgenase-1, a Cardinal Modulator of Regulated Cell Death and Inflammation. Cells 2021, 10, 515. [Google Scholar]
- Kwon, M.Y.; Park, E.; Lee, S.J.; Chung, S.W. Heme oxygenase-1 accelerates erastin-induced ferroptotic cell death. Oncotarget 2015, 6, 24393–24403. [Google Scholar]
- Cosialls, E.; El Hage, R.; Dos Santos, L.; Gong, C.; Mehrpour, M.; Hamaï, A. Ferroptosis: Cancer Stem Cells Rely on Iron until “to Die for” It. Cells 2021, 10, 2981. [Google Scholar]
- Ryter, S.W.; Alam, J.; Choi, A.M. Heme oxygenase-1/carbon monoxide: From basic science to therapeutic applications. Physiol. Rev. 2006, 86, 583–650. [Google Scholar]
- Vítek, L.; Schwertner, H.A. The heme catabolic pathway and its protective effects on oxidative stress-mediated diseases. Adv. Clin. Chem. 2007, 43, 1–57. [Google Scholar]
- Papanikolaou, G.; Pantopoulos, K. Iron metabolism and toxicity. Toxicol. Appl. Pharmacol. 2005, 202, 199–211. [Google Scholar]
- Dixon, S.J.; Lemberg, K.M.; Lamprecht, M.R.; Skouta, R.; Zaitsev, E.M.; Gleason, C.E.; Patel, D.N.; Bauer, A.J.; Cantley, A.M.; Yang, W.S.; et al. Ferroptosis: An iron-dependent form of nonapoptotic cell death. Cell 2012, 149, 1060–1072. [Google Scholar]
- Yang, W.S.; SriRamaratnam, R.; Welsch, M.E.; Shimada, K.; Skouta, R.; Viswanathan, V.S.; Cheah, J.H.; Clemons, P.A.; Shamji, A.F.; Clish, C.B.; et al. Regulation of ferroptotic cancer cell death by GPX4. Cell 2014, 156, 317–331. [Google Scholar]
- Xie, Y.; Hou, W.; Song, X.; Yu, Y.; Huang, J.; Sun, X.; Kang, R.; Tang, D. Ferroptosis: Process and function. Cell Death Differ. 2016, 23, 369–379. [Google Scholar]
- Stockwell, B.R.; Friedmann Angeli, J.P.; Bayir, H.; Bush, A.I.; Conrad, M.; Dixon, S.J.; Fulda, S.; Gascón, S.; Hatzios, S.K.; Kagan, V.E.; et al. Ferroptosis: A Regulated Cell Death Nexus Linking Metabolism, Redox Biology, and Disease. Cell 2017, 171, 273–285. [Google Scholar]
- Lanceta, L.; Li, C.; Choi, A.M.; Eaton, J.W. Haem oxygenase-1 overexpression alters intracellular iron distribution. Biochem. J. 2013, 449, 189–194. [Google Scholar]
- Gonzales, S.; Erario, M.A.; Tomaro, M.L. Heme oxygenase-1 induction and dependent increase in ferritin. A protective antioxidant stratagem in hemin-treated rat brain. Dev. Neurosci. 2002, 24, 161–168. [Google Scholar]
- Ryter, S.W.; Otterbein, L.E. Carbon monoxide in biology and medicine. Bioessays 2004, 26, 270–280. [Google Scholar]
- Tofighi, R.; Tillmark, N.; Daré, E.; Aberg, A.M.; Larsson, J.E.; Ceccatelli, S. Hypoxia-independent apoptosis in neural cells exposed to carbon monoxide in vitro. Brain Res. 2006, 1098, 1–8. [Google Scholar]
- Xu, X.; Zhang, H.; Wang, K.; Tu, T.; Jiang, Y. Protective Effect of Edaravone against Carbon Monoxide Induced Apoptosis in Rat Primary Cultured Astrocytes. Biochem. Res. Int. 2017, 2017, 5839762. [Google Scholar]
- Tian, X.; Guan, T.; Guo, Y.; Zhang, G.; Kong, J. Selective Susceptibility of Oligodendrocytes to Carbon Monoxide Poisoning: Implication for Delayed Neurologic Sequelae (DNS). Front. Psychiatry 2020, 11, 815. [Google Scholar]
- Loboda, A.; Jozkowicz, A.; Dulak, J. HO-1/CO system in tumor growth, angiogenesis and metabolism-Targeting HO-1 as an anti-tumor therapy. Vascul. Pharmacol. 2015, 74, 11–22. [Google Scholar]
- Shahrokhi, S.Z.; Karami Tehrani, F.S.; Salami, S. Induction of cell apoptosis by biliverdin reductase inhibitor in MCF-7 and MDA-MB-468 breast cancer cell lines: Experimental and in silico studies. Excli. J. 2021, 20, 1502–1516. [Google Scholar]
- Chang, L.C.; Chiang, S.K.; Chen, S.E.; Yu, Y.L.; Chou, R.H.; Chang, W.C. Heme oxygenase-1 mediates BAY 11-7085 induced ferroptosis. Cancer Lett. 2018, 416, 124–137. [Google Scholar]
- Adedoyin, O.; Boddu, R.; Traylor, A.; Lever, J.M.; Bolisetty, S.; George, J.F.; Agarwal, A. Heme oxygenase-1 mitigates ferroptosis in renal proximal tubule cells. Am. J. Physiol. Renal. Physiol. 2018, 314, F702–F714. [Google Scholar]
- Sun, X.; Ou, Z.; Chen, R.; Niu, X.; Chen, D.; Kang, R.; Tang, D. Activation of the p62-Keap1-NRF2 pathway protects against ferroptosis in hepatocellular carcinoma cells. Hepatology 2016, 63, 173–184. [Google Scholar]
- Gorrini, C.; Harris, I.S.; Mak, T.W. Modulation of oxidative stress as an anticancer strategy. Nat. Rev. Drug Discov. 2013, 12, 931–947. [Google Scholar]
- Hassannia, B.; Wiernicki, B.; Ingold, I.; Qu, F.; Van Herck, S.; Tyurina, Y.Y.; Bayır, H.; Abhari, B.A.; Angeli, J.P.F.; Choi, S.M.; et al. Nano-targeted induction of dual ferroptotic mechanisms eradicates high-risk neuroblastoma. J. Clin. Investig. 2018, 128, 3341–3355. [Google Scholar]
- Fujino, M.; Nishio, Y.; Ito, H.; Tanaka, T.; Li, X.K. 5-Aminolevulinic acid regulates the inflammatory response and alloimmune reaction. Int. Immunopharmacol. 2016, 37, 71–78. [Google Scholar]
- Liu, C.; Fujino, M.; Zhu, S.; Isaka, Y.; Ito, H.; Takahashi, K.; Nakajima, M.; Tanaka, T.; Zhu, P.; Li, X.K. 5-ALA/SFC enhances HO-1 expression through the MAPK/Nrf2 antioxidant pathway and attenuates murine tubular epithelial cell apoptosis. FEBS Open Bio. 2019, 9, 1928–1938. [Google Scholar]
- Uchida, A.; Kidokoro, K.; Sogawa, Y.; Itano, S.; Nagasu, H.; Satoh, M.; Sasaki, T.; Kashihara, N. 5-Aminolevulinic acid exerts renoprotective effect via Nrf2 activation in murine rhabdomyolysis-induced acute kidney injury. Nephrology 2019, 24, 28–38. [Google Scholar]
- Han, L.; Batistel, F.; Ma, Y.; Alharthi, A.S.M.; Parys, C.; Loor, J.J. Methionine supply alters mammary gland antioxidant gene networks via phosphorylation of nuclear factor erythroid 2-like 2 (NFE2L2) protein in dairy cows during the periparturient period. Dairy Sci. 2018, 101, 8505–8512. [Google Scholar]
- Marra, K.; LaRochelle, E.P.; Chapman, M.S.; Hoopes, P.J.; Lukovits, K.; Maytin, E.V.; Hasan, T.; Pogue, B.W. Comparison of Blue and White Lamp Light with Sunlight for Daylight-Mediated, 5-ALA Photodynamic Therapy, in vivo. Photochem. Photobiol. 2018, 94, 1049–1057. [Google Scholar]
- Aalders, M.C.; vd Vange, N.; Stewart, F.A.; Klein, M.G.; vd Vijver, M.J.; Sterenborg, H.J. White-light toxicity, resulting from systemically administered 5-aminolevulinic acid, under normal operating conditions. J. Photochem. Photobiol. B 1999, 50, 88–93. [Google Scholar]
- Pittalà, V.; Salerno, L.; Romeo, G.; Modica, M.N.; Siracusa, M.A. A focus on heme oxygenase-1 (HO-1) inhibitors. Curr. Med. Chem. 2013, 20, 3711–3732. [Google Scholar]
- Mori, S.; Kishi, S.; Honoki, K.; Fujiwara-Tani, R.; Moriguchi, T.; Sasaki, T.; Fujii, K.; Tsukamoto, S.; Fujii, H.; Kido, A.; et al. Anti-Stem Cell Property of Pterostilbene in Gastrointestinal Cancer Cells. Int. J. Mol. Sci. 2020, 21, 9347. [Google Scholar]
- Kuniyasu, H.; Luo, Y.; Fujii, K.; Sasahira, T.; Moriwaka, Y.; Tatsumoto, N.; Sasaki, T.; Yamashita, Y.; Ohmori, H. CD10 enhances metastasis of colorectal cancer by abrogating the anti-tumoural effect of methionine-enkephalin in the liver. Gut 2010, 59, 348–356. [Google Scholar]
- Suzuki, Y.; Sakagishi, Y. Diazo coupling reaction for the determination of biliverdin. Bunseki Kagaku 1991, 40, 377–381. [Google Scholar]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Gene | ID | Sequence | |
|---|---|---|---|
| ACTB | NM_001101.3 | Upper | GGACTTCGAGCAAGAGATGG |
| Lower | AGCACTGTGTTGGCGTACAG | ||
| HO-1 | NM_002133.2 | Upper | TAAGCTGGTGATGGCTTCCT |
| Lower | ATGATTTCCTGCCAGTGAGG | ||
| FeCH | KR712044.1 | Upper | GATGAATTGTCCCCCAACAC |
| Lower | GCTTCCGTCCCACTTGATTA | ||
| NS | BC001024.2 | Upper | ATTGCCAACAGTGGTGTTCA |
| Lower | AATGGCTTTGCTGCAAGTTT | ||
| Oct3 | BC117437.1 | Upper | GAAGGATGTGGTCCGAGTGT |
| Lower | GTGAAGTGAGGGCTCCCATA | ||
| Wnt1 | NM_005430.4 | Upper | CGGCGTTTATCTTCGCTATC |
| Lower | GCCTCGTTGTTGTGAAGGTT | ||
| Notch1 | CR457221.1 | Upper | GATGTGTGGACTGTGGCACT |
| Lower | TGTGTTGCTGGAGCATCTTC | ||
| ECE | NM_001397.3 | Upper | GACAGATGCCTGCTCAACAA |
| Lower | GCCCAGGTTGTTTTCTGTGT | ||
| c-MYC | NM_002467.4 | Upper | TTCGGGTAGTGGAAAACCAG |
| Lower | CAGCAGCTCGAATTTCTTCC | ||
| PGC-1α | BC156323.1 | Upper | GTGAAGACCAGCCTCTTTGC |
| Lower | AATCCGTCTTCATCCACAGG |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Horii, S.; Mori, S.; Ogata, R.; Nukaga, S.; Nishida, R.; Kishi, S.; Sasaki, R.; Ikemoto, A.; Owari, T.; Maesaka, F.; et al. 5-Aminolevrinic Acid Exhibits Dual Effects on Stemness in Human Sarcoma Cell Lines under Dark Conditions. Int. J. Mol. Sci. 2023, 24, 6189. https://doi.org/10.3390/ijms24076189
Horii S, Mori S, Ogata R, Nukaga S, Nishida R, Kishi S, Sasaki R, Ikemoto A, Owari T, Maesaka F, et al. 5-Aminolevrinic Acid Exhibits Dual Effects on Stemness in Human Sarcoma Cell Lines under Dark Conditions. International Journal of Molecular Sciences. 2023; 24(7):6189. https://doi.org/10.3390/ijms24076189
Chicago/Turabian StyleHorii, Shohei, Shiori Mori, Ruiko Ogata, Shota Nukaga, Ryoichi Nishida, Shingo Kishi, Rika Sasaki, Ayaka Ikemoto, Takuya Owari, Fumisato Maesaka, and et al. 2023. "5-Aminolevrinic Acid Exhibits Dual Effects on Stemness in Human Sarcoma Cell Lines under Dark Conditions" International Journal of Molecular Sciences 24, no. 7: 6189. https://doi.org/10.3390/ijms24076189