
A Proposed New Model to Explain the Role of Low Dose Non-DNA Targeted Radiation Exposure in Chronic Fatigue and Immune Dysfunction Syndrome
{kind=link}
Abstract
:1. Introduction
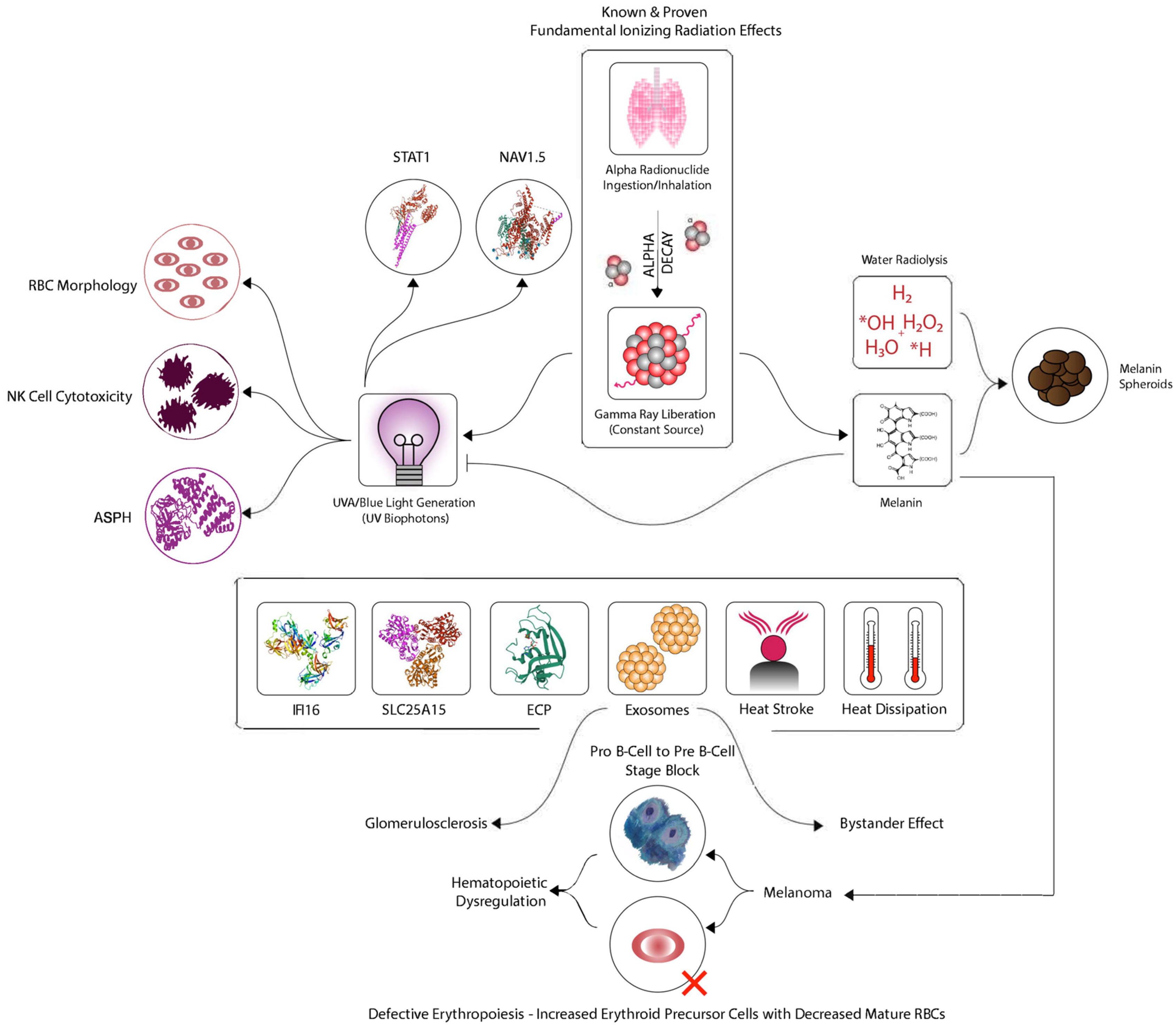
1.1. Background to the Model
1.2. Background to the Proposed Mechanism
1.3. The Bridge from Bystander Signaling Stress to CFIDS
1.4. The Connection to Melanoma
1.5. CFIDS and Cancers Other Than Melanoma
2. Various Biomarkers Tied to UVA Exposure and to Melanoma
2.1. STAT1
2.2. NaV1.5
2.3. ASPH
2.4. NK Cell Cytotoxicity
2.5. RBC Morphology
2.6. IFI16
2.7. SLC25A15
2.8. ECP
2.9. Heat Stroke and Heat Dissipation
2.10. Exosomes
3. Discussion
4. Conclusions
Author Contributions
Funding
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Bell, D.S. The Disease of a Thousand Names: CFIDS—CHRONIC Fatigue/Immune Dysfunction Syndrome; Pollard Publications: La Jolla, CA, USA, 1991. [Google Scholar]
- Loganovsky, K. Chronic Fatigue Syndrome as a Characteristic Aftermath of Radioecological Catastrophe. Int. J. Psychophysiol. 2000, 35, 69. [Google Scholar]
- Loganovsky, K.N. Chronic Fatigue Syndrome in the Chernobyl Accident Consequences Liquidators. Int. J. Radiat. Med. 2001, 3, 76. [Google Scholar]
- Loganovsky, K. Do Low Doses of Ionizing Radiation Affect the Human Brain? Data Sci. J. 2009, 8, 13–35. [Google Scholar] [CrossRef]
- Tang, F.R.; Loganovsky, K. Low Dose or Low Dose Rate Ionizing Radiation-Induced Health Effect in the Human. J. Environ. Radioact. 2018, 192, 32–47. [Google Scholar] [CrossRef] [PubMed]
- Bazyka, D.; Loganovsky, K.; Ilyenko, I.; Volovyk, S.; Perchuk, I.; Pleskach, O.; Nechayev, S. Psychophysiological, Neuroimmune and Gene Expression Changes in Chronic Fatigue Syndrome after Low-Dose Radiation Exposure. Int. J. Psychophysiol. 2010, 77, 340. [Google Scholar] [CrossRef]
- Loganovsky, K.N. Vegetative-Vascular Dystonia and Osteoalgetic Syndrome or Chronic Fatigue Syndrome as a Characteristic After-Effect of Radioecological Disaster. J. Chronic Fatigue Syndr. 2000, 7, 3–16. [Google Scholar] [CrossRef]
- National CFIDS Foundation. National CFIDS Foundation (NCF) Announces Link between Chronic Fatigue Syndrome and Low Level Radiation Exposure; PR Newswire: Chicago, IL, USA, 2010. [Google Scholar]
- National CFIDS Foundation. National CFIDS Foundation’s Research Finds Chromosome Damage in Patients Diagnosed with Chronic Fatigue Syndrome and Myalgic Encephalomyelitis; PR Newswire: Chicago, IL, USA, 2014. [Google Scholar]
- Heng, H.H.Q.; Liu, G.; Stevens, J.B.; Abdallah, B.Y.; Horne, S.D.; Ye, K.J.; Bremer, S.W.; Chowdhury, S.K.; Ye, C.J. Karyotype Heterogeneity and Unclassified Chromosomal Abnormalities. Cytogenet. Genome Res. 2013, 139, 144–157. [Google Scholar] [CrossRef]
- Boubriak, I.; Akimkina, T.; Polischuk, V.; Dmitriev, A.; Mccready, S.; Grodzinsky, D. Long term effects of chernobyl contamination on dna repair function and plant resistance to biotic and abiotic stress factors. TSitologiia Genet. 2016, 50, 34–59. [Google Scholar] [CrossRef] [Green Version]
- Yushkova, E. Genetic mechanisms of formation of radiation-induced instability of the genome and its transgenerational effects in the descendants of chronically irradiated individuals of Drosophila melanogaster. Radiat. Environ. Biophys. 2020, 59, 221–236. [Google Scholar] [CrossRef]
- Ahmad, S.B.; McNeill, F.E.; Byun, S.H.; Prestwich, W.V.; Seymour, C.; Mothersill, C.E. Ion Beam Induced Luminescence; Relevance to Radiation Induced Bystander Effects. Nucl. Instrum. Methods Phys. Res. Sect. B Beam Interact. Mater. At. 2012, 288, 81–88. [Google Scholar] [CrossRef]
- Ahmad, S.B.; McNeill, F.E.; Byun, S.H.; Prestwich, W.V.; Mothersill, C.; Seymour, C.; Armstrong, A.; Fernandez, C. Ultra-Violet Light Emission from HPV-G Cells Irradiated with Low Let Radiation From (90)Y; Consequences for Radiation Induced Bystander Effects. Dose Response 2013, 11, 498–516. [Google Scholar] [CrossRef] [Green Version]
- Mothersill, C.; Le, M.; Rusin, A.; Seymour, C. Biophotons in Radiobiology: Inhibitors, Communicators and Reactors. Radiat. Prot. Dosim. 2019, 183, 136–141. [Google Scholar] [CrossRef]
- Le, M.; McNeill, F.E.; Seymour, C.; Rainbow, A.J.; Mothersill, C.E. An observed effect of ultraviolet radiation emitted from beta-irradiated HaCaT cells upon non-beta-irradiated bystander cells. Radiat. Res. 2015, 183, 279–290. [Google Scholar] [CrossRef] [PubMed]
- Le, M.; Mothersill, C.E.; Seymour, C.B.; Ahmad, S.B.; Armstrong, A.; Rainbow, A.J.; McNeill, F.E. Factors affecting ultraviolet-A photon emission from β-irradiated human keratinocyte cells. Phys. Med. Biol. 2015, 60, 6371–6389. [Google Scholar] [CrossRef] [PubMed]
- Le, M.; Mothersill, C.E.; Seymour, C.B.; Rainbow, A.J.; McNeill, F.E. An Observed Effect of p53 Status on the Bystander Response to Radiation-Induced Cellular Photon Emission. Radiat. Res. 2017, 9187, 169–185. [Google Scholar] [CrossRef] [PubMed]
- Cohen, J.; Vo, N.T.K.; Chettle, D.R.; McNeill, F.E.; Seymour, C.B.; Mothersill, C.E. Quantifying Biophoton Emissions From Human Cells Directly Exposed to Low-Dose Gamma Radiation. Dose Response 2020, 18, 1559325820926763. [Google Scholar] [CrossRef] [PubMed]
- Le, M.; McNeill, F.E.; Seymour, C.B.; Rusin, A.; Diamond, K.; Rainbow, A.J.; Murphy, J.; Mothersill, C.E. Modulation of oxidative phosphorylation (OXPHOS) by radiation- induced biophotons. Environ. Res. 2018, 163, 80–87. [Google Scholar] [CrossRef]
- The Water in You: Water and the Human Body. Water Science School. 2019. Available online: https://www.usgs.gov/special-topics/water-science-school/science/water-you-water-and-human-body (accessed on 13 March 2023).
- Le Caer, S. Radiolysis: Influence of Oxide Surfaces on H2 Production under Ionizing Radiation. Water 2011, 3, 235–253. [Google Scholar] [CrossRef] [Green Version]
- Richards, R.S.; Roberts, T.K.; McGregor, N.R.; Dunstan, R.H.; Butt, H.L. Blood parameters indicative of oxidative stress are associated with symptom expression in chronic fatigue syndrome. Redox Rep. 2000, 5, 35–41. [Google Scholar] [CrossRef] [Green Version]
- Clutton, S.M.; Townsend, K.M.; Walker, C.; Ansell, J.D.; Wright, E.G. Radiation-induced genomic instability and persisting oxidative stress in primary bone marrow cultures. Carcinogenesis 1996, 17, 1633–1639. [Google Scholar] [CrossRef]
- O’Reilly, J.P.; Mothersill, C. Comparative effects of UV A and UV B on clonogenic survival and delayed cell death in skin cell lines from humans and fish. Int. J. Radiat. Biol. 1997, 72, 111–119. [Google Scholar] [PubMed]
- Ridley, A.J.; Whiteside, J.R.; McMillan, T.J.; Allinson, S.L. Cellular and sub-cellular responses to UVA in relation to carcinogenesis. Int. J. Radiat. Biol. 2009, 85, 177–195. [Google Scholar] [CrossRef] [PubMed]
- Whiteside, J.R.; McMillan, T.J. A bystander effect is induced in human cells treated with UVA radiation but not UVB radiation. Radiat. Res. 2009, 171, 204–211. [Google Scholar] [CrossRef] [PubMed]
- Jella, K.K.; Moriarty, R.; McClean, B.; Byrne, H.J.; Lyng, F.M. Reactive oxygen species and nitric oxide signaling in bystander cells. PLoS ONE 2018, 13, e0195371. [Google Scholar] [CrossRef] [PubMed]
- NIH News in Health. Sun and Skin: The Dark Side of Sun Exposure; NIH News in Health: Bethesda, MD, USA, 2014. [Google Scholar]
- Jin, S.G.; Padron, F.; Pfeifer, G.P. UVA Radiation, DNA Damage, and Melanoma. ACS Omega 2022, 7, 32936–32948. [Google Scholar] [CrossRef]
- Bernerd, F.; Passeron, T.; Castiel, I.; Marionnet, C. The Damaging Effects of Long UVA (UVA1) Rays: A Major Challenge to Preserve Skin Health and Integrity. Int. J. Mol. Sci. 2022, 23, 8243. [Google Scholar] [CrossRef]
- Khan, A.Q.; Travers, J.B.; Kemp, M.G. Roles of UVA Radiation and DNA Damage Responses in Melanoma Pathogenesis. Environ. Mol. Mutagen. 2018, 59, 438–460. [Google Scholar] [CrossRef]
- Kapp, F.G.; Perlin, J.R.; Hagedorn, E.J.; Gansner, J.M.; Schwarz, D.E.; O’Connell, L.A.; Johnson, N.S.; Amemiya, C.; Fisher, D.E.; Wolfle, U.; et al. Protection from UV light is an evolutionarily conserved feature of the hematopoietic niche. Nature 2018, 558, 445–448. [Google Scholar] [CrossRef]
- Kamran, N.; Li, Y.; Sierra, M.; Alghamri, M.S.; Kadiyala, P.; Appelman, H.D.; Edwards, M.; Lowenstein, P.R.; Castro, M.G. Melanoma induced immunosuppression is mediated by hematopoietic dysregulation. Oncoimmunology 2017, 7, e1408750. [Google Scholar] [CrossRef] [Green Version]
- Decker, J.M. Immunology: B-Cell Development. Department of Veterinary Science and Microbiology, University of Arizona. Available online: http://microvet.arizona.edu/Courses/MIC419/Tutorials/Bcelldevelopment.html (accessed on 13 March 2023).
- Chang, C.M.; Warren, J.L.; Engels, E.A. Chronic fatigue syndrome and subsequent risk of cancer among elderly US adults. Cancer 2012, 118, 5929–5936. [Google Scholar] [CrossRef] [Green Version]
- Johnson, H. Osler’s Web: Inside the Labyrinth of the Chronic Fatigue Syndrome Epidemic; Crown Publishers: New York, NY, USA, 1996. [Google Scholar]
- Lundell, K.; Qazi, S.; Eddy, L.; Uckun, F.M. Clinical activity of folinic acid in patients with chronic fatigue syndrome. Arzneimittelforschung 2006, 56, 399–404. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Turick, C.E.; Ekechukwu, A.A.; Milliken, C.E.; Casadevall, A.; Dadachova, E. Gamma radiation interacts with melanin to alter its oxidation-reduction potential and results in electric current production. Bioelectrochemistry 2011, 82, 69–73. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Vienneau, D.; de Hoogh, K.; Hauri, D.; Vicedo-Cabrera, A.M.; Schindler, C.; Huss, A.; Roosli, M.; SNC Study Group. Effects of Radon and UV Exposure on Skin Cancer Mortality in Switzerland. Environ. Health Perspect. 2017, 125, 067009. [Google Scholar]
- Knox, K.K.; Carrigan, D.R. Potential Role of STAT1 in the Pathogenesis of Chronic Fatigue Syndrome; Institute for Viral Pathogenesis: Madison, WI, USA, 2003. [Google Scholar]
- Knox, K.K.; Cocchetto, A.; Jordan, E.; Leech, D.; Carrigan, D.R. Deficiency in the Expression of STAT1 Protein in a Subpopulation of Patients with Chronic Fatigue Syndrome (CFS). In Proceedings of the Seventh International AACFS Conference on Chronic Fatigue Syndrome, Fibromyalgia and other Related Illnesses, Madison, WI, USA, 8–10 October 2004. [Google Scholar]
- Maziere, C.; Dantin, F.; Dubois, F.; Santus, R.; Maziere, J. Biphasic Effect of UVA Radiation on Stat1 Activity and Tyrosine Phosphorylation in Cultured Human Keratinocytes. Free Radic. Bio. Med. 2000, 28, 1430–1437. [Google Scholar] [CrossRef]
- Aragane, Y.; Kulms, D.; Luger, T.A.; Schwarz, T. Down-regulation of interferon gamma-activated STAT1 by UV light. Proc. Natl. Acad. Sci. USA 1997, 94, 11490–11495. [Google Scholar] [CrossRef] [Green Version]
- Kwon, T.R.; Oh, C.T.; Choi, E.J.; Kim, S.R.; Jang, Y.J.; Ko, E.J.; Suh, D.; Yoo, K.H.; Kim, B.J. Ultraviolet light-emitting-diode irradiation inhibits TNF-α and IFN-γ-induced expression of ICAM-1 and STAT1 phosphorylation in human keratinocytes. Lasers Surg. Med. 2015, 47, 824–832. [Google Scholar] [CrossRef]
- Eschrich, S.; Zhang, H.; Zhao, H.; Boulware, D.; Lee, J.H.; Bloom, G.; Torres-Roca, J.F. Systems biology modeling of the radiation sensitivity network: A biomarker discovery platform. Int. J. Radiat. Oncol. Biol. Phys. 2009, 75, 497–505. [Google Scholar] [CrossRef] [Green Version]
- Huang, L.; Chen, J.; Zhao, Y.; Gu, L.; Shao, X.; Li, J.; Xu, Y.; Liu, Z.; Xu, Q. Key candidate genes of STAT1 and CXCL10 in melanoma identified by integrated bioinformatical analysis. IUBMB Life 2019, 71, 1634–1644. [Google Scholar] [CrossRef]
- Friedman, M.A.; Fernandez, M.; Backer, L.C.; Dickey, R.W.; Bernstein, J.; Schrank, K.; Kibler, S.; Wendy, S.; Gribble, M.O.; Bienfang, P.; et al. An Updated Review of Ciguatera Fish Poisoning: Clinical, Epidemiological, Environmental, and Public Health Management. Mar. Drugs 2017, 15, 72. [Google Scholar] [CrossRef] [Green Version]
- Pearn, J.H. Chronic fatigue syndrome: Chronic ciguatera poisoning as a differential diagnosis. Med. J. Aust. 1997, 166, 309–310. [Google Scholar] [CrossRef]
- Lombet, A.; Bidard, J.N.; Lazdunski, M. Ciguatoxin and brevetoxins share a common receptor site on the neuronal voltage-dependent Na+ channel. FEBS Lett. 1987, 219, 355–359. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rook, M.B.; Evers, M.M.; Vos, M.A.; Bierhuizen, M.F.A. Biology of cardiac sodium channel Nav1. 5 expression. Cardiovasc. Res. 2012, 93, 12–23. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hokama, Y.; John A Burns School of Medicine, University of Hawaii, Honolulu, HI, USA. Personal communication, 2008.
- Hokama, Y.; Uto, G.A.; Palafox, N.A.; Enlander, D.; Jordan, E.; Cocchetto, A. Chronic phase lipids in sera of chronic fatigue syndrome (CFS), chronic ciguatera fish poisoning (CCFP), hepatitis B, and cancer with antigenic epitope resembling ciguatoxin, as assessed with Mab-CTX. J. Clin. Lab. Anal. 2003, 17, 132–139. [Google Scholar] [CrossRef] [PubMed]
- Hokama, Y.; Empey-Campora, C.; Hara, C.; Higa, N.; Siu, N.; Lau, R.; Kuribayashi, T.; Yabusaki, K. Acute phase phospholipids related to the cardiolipin of mitochondria in the sera of patients with chronic fatigue syndrome (CFS), chronic Ciguatera fish poisoning (CCFP), and other diseases attributed to chemicals, Gulf War, and marine toxins. J. Clin. Lab. Anal. 2008, 22, 99–105. [Google Scholar] [CrossRef] [PubMed]
- Wang, G.K.; Wang, S.Y. Modifications of human cardiac sodium channel gating by UVA light. J. Membr. Biol. 2002, 189, 153–165. [Google Scholar] [CrossRef]
- Xie, A.; Gallant, B.; Guo, H.; Gonzalez, A.; Clark, M.; Madigan, A.; Feng, F.; Chen, H.D.; Cui, Y.; Dudley, S.C., Jr.; et al. Functional cardiac Na + channels are expressed in human melanoma cells. Onco. Lett. 2018, 16, 1689–1695. [Google Scholar] [CrossRef] [PubMed]
- Djamgoz, M.B.A.; Fraser, S.P.; Brackenbury, W.J. In Vivo Evidence for Voltage-Gated Sodium Channel Expression in Carcinomas and Potentiation of Metastasis. Cancers 2019, 11, 1675. [Google Scholar] [CrossRef] [Green Version]
- Roelant, C.H.S.; DeMeirleir, K.L. Methods and means for diagnosing and and/or treating a fatiguing illness. World Patent WO2019012159, 16 July 2018. [Google Scholar]
- Hou, G.; Xu, B.; Bi, Y.; Wu, C.; Ru, B.; Sun, B.; Bai, X. Recent advances in research on aspartate β-hydroxylase (ASPH) in pancreatic cancer: A brief update. Bosn. J. Basic Med. Sci. 2018, 18, 297–304. [Google Scholar] [CrossRef]
- Zheng, W.; Wang, X.; Hu, J.; Bai, B.; Zhu, H. Diverse molecular functions of aspartate β-hydroxylase in cancer (Review). Oncol. Rep. 2020, 44, 2364–2372. [Google Scholar] [CrossRef]
- Wands, J.R.; De La Monte, S.; Aihara, A.; Olsen, M.J.; Thomas, J.M. Inhibitors of beta-hydroxylase for treatment of cancer. U.S. Patent Application 20200361925, 2 June 2020. [Google Scholar]
- Radrezza, S.; Carini, M.; Baron, G.; Aldini, G.; Negre-Salvayre, A.; D’Amat, A. Study of Carnosine’s effect on nude mice skin to prevent UV-A damage. Free Radic. Biol. Med. 2021, 173, 97–103. [Google Scholar] [CrossRef]
- Miller, J.S.; McCullar, V.; Punzel, M.; Lemischka, I.R.; Moore, K.A. Single adult human CD34(+)/Lin-/CD38(-) progenitors give rise to natural killer cells, B-lineage cells, dendritic cells, and myeloid cells. Blood 1999, 93, 96–106. [Google Scholar] [CrossRef]
- Brenu, E.W.; Hardcastle, S.L.; Atkinson, G.M.; van Driel, M.L.; Kreijkamp-Kaspers, S.; Ashton, K.J.; Staines, D.R.; Marshall-Gradisnik, S.M. Natural killer cells in patients with severe chronic fatigue syndrome. Auto Immun. Highlights 2013, 4, 69–80. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Vokurkova, D.; Vavrova, J.; Sinkora, J.; Stoklasova, A.; Blaha, V.; Rezacova, M. Radiosensitivity of CD3-CD8+CD56+ NK cells. Radiat. Meas. 2010, 45, 1020–1023. [Google Scholar] [CrossRef]
- Brenu, E.W.; Staines, D.R.; Baskurt, O.K.; Ashton, K.J.; Ramos, S.B.; Christy, R.M.; Marshall-Gradisnik, S.M. Immune and hemorheological changes in chronic fatigue syndrome. J. Transl. Med. 2010, 8, 1. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Brenu, E.W.; van Driel, M.L.; Staines, D.R.; Ashton, K.J.; Ramos, S.B.; Keane, J.; Klimas, N.G.; Marshall-Gradisnik, S.M. Immunological abnormalities as potential biomarkers in Chronic Fatigue Syndrome/Myalgic Encephalomyelitis. J. Transl. Med. 2011, 9, 81. [Google Scholar] [CrossRef] [Green Version]
- Brenu, E.W.; van Driel, M.L.; Staines, D.R.; Ashton, K.J.; Hardcastle, S.L.; Keane, J.; Tajouri, L.; Peterson, D.; Ramos, S.B.; Marshall-Gradisnik, S.M. Longitudinal investigation of natural killer cells and cytokines in chronic fatigue syndrome/myalgic encephalomyelitis. J. Transl. Med. 2012, 10, 88. [Google Scholar] [CrossRef] [Green Version]
- Hersey, P.; MacDonald, M.; Henderson, C.; Schibeci, S.; D’Alessandro, G.; Pryor, M.; Wilkinson, F.J. Suppression of natural killer cell activity in humans by radiation from solarium lamps depleted of UVB. J. Investig. Dermatol. 1988, 90, 305–310. [Google Scholar] [CrossRef]
- Hersey, P.; Magrath, H.; Wilkinson, F. Development of an in vitro system for the analysis of ultraviolet radiation-induced suppression of natural killer cell activity. Photochem. Photobiol. 1993, 57, 279–284. [Google Scholar] [CrossRef]
- de Jonge, K.; Ebering, A.; Nassiri, S.; Maby-El Hajjami, H.; Ouertatani-Sakouhi, H.; Baumgaertner, P.; Speiser, D.E. Circulating CD56 bright NK cells inversely correlate with survival of melanoma patients. Sci. Rep. 2019, 9, 4487. [Google Scholar] [CrossRef] [Green Version]
- Simpson, L.O. The role of nondiscocytic erythrocytes in the pathogenesis of myalgic encephalomyelitis/chronic fatigue syndrome. In The Clinical and Scientific Basis of Myalgic Encephalomyelitis/Chronic Fatigue Syndrome; Hyde, B.M., Ed.; The Nightingale Research Foundation: Street Ottawa, ON, Canada, 1992; Volume 65, pp. 597–605. [Google Scholar]
- Richards, R.S.; Wang, L.; Jelinek, H. Erythrocyte oxidative damage in chronic fatigue syndrome. Arch Med. Res. 2007, 38, 94–98. [Google Scholar] [CrossRef]
- Saha, A.K.; Schmidt, B.R.; Wilhelmy, J.; Nguyen, V.; Abugherir, A.; Do, J.K.; Nemat-Gorgani, M.; Davis, R.W.; Ramasubramanian, A.K. Red blood cell deformability is diminished in patients with Chronic Fatigue Syndrome. Clin. Hemorheol. Microcirc. 2019, 71, 113–116. [Google Scholar] [CrossRef]
- Kozlova, E.; Chernysh, A.; Sergunova, V.; Gudkova, O.; Manchenko, E.; Kozlov, A. Atomic force microscopy study of red blood cell membrane nanostructure during oxidation-reduction processes. J. Mol. Recognit. 2018, 31, e2724. [Google Scholar] [CrossRef] [PubMed]
- Nguyen, C.B.; Alsøe, L.; Lindvall, J.M.; Sulheim, D.; Fagermoen, E.; Winger, A.; Kaarbø, M.; Nilsen, H.; Wyller, V.B. Whole blood gene expression in adolescent chronic fatigue syndrome: An exploratory cross-sectional study suggesting altered B cell differentiation and survival. J. Transl. Med. 2017, 15, 102. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sato, W.; Ono, H.; Matsutani, T.; Nakamura, M.; Shin, I.; Amano, K.; Suzuki, R.; Yamamura, T. Skewing of the B cell receptor repertoire in myalgic encephalomyelitis/chronic fatigue syndrome. Brain Behav. Immun. 2021, 95, 245–255. [Google Scholar] [CrossRef]
- Costa, S.; Borgogna, C.; Mondini, M.; De Andrea, M.; Meroni, P.L.; Berti, E.; Gariglio, M.; Landolfo, S. Redistribution of the nuclear protein IFI16 into the cytoplasm of ultraviolet B-exposed keratinocytes as a mechanism of autoantigen processing. Br. J. Dermatol. 2011, 164, 282–290. [Google Scholar] [CrossRef] [PubMed]
- Wang, H.; Xie, X.; Zhu, J.; Qi, S.; Xie, J. Comprehensive analysis identifies IFI16 as a novel signature associated with overall survival and immune infiltration of skin cutaneous melanoma. Cancer Cell Int. 2021, 21, 694. [Google Scholar] [CrossRef]
- Dibble, J.J.; McGrath, S.J.; Ponting, C.P. Genetic risk factors of ME/CFS: A critical review. Hum. Mol. Genet. 2020, 29, R117–R124. [Google Scholar] [CrossRef]
- Ji, S.M. Overexpression of SLC25A15 is involved in the proliferation of cutaneous melanoma and leads to poor prognosis. Med. Sci. (Paris) 2018, 34, 74–80. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Boix, E.; Carreras, E.; Nikolovski, Z.; Cuchillo, C.M.; Nogues, M.V. Identification and characterization of human eosinophil cationic protein by an epitope-specific antibody. J. Leukoc. Biol. 2001, 69, 1027–1035. [Google Scholar] [CrossRef] [PubMed]
- Conti, F.; Magrini, L.; Priori, R.; Valesini, G.; Bonini, S. Eosinophil cationic protein serum levels and allergy in chronic fatigue syndrome. Allergy 1996, 51, 124–127. [Google Scholar] [CrossRef]
- Sorensen, B.; Streib, J.E.; Strand, M.; Make, B.; Giclas, P.C.; Fleshner, M.; Jones, J.F. Complement activation in a model of chronic fatigue syndrome. J. Allergy Clin. Immunol. 2003, 112, 397–403. [Google Scholar] [CrossRef] [PubMed]
- Kruckel, A.; Moreira, A.; Frohlich, W.; Schuler, G.; Heinzerling, L. Eosinophil-cationic protein—A novel liquid prognostic biomarker in melanoma. BMC Cancer 2019, 19, 207. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mueller, C.; Lin, J.C.; Sheriff, S.; Maudsley, A.A.; Younger, J.W. Evidence of widespread metabolite abnormalities in Myalgic encephalomyelitis/chronic fatigue syndrome: Assessment with whole-brain magnetic resonance spectroscopy. Brain Imaging Behav. 2020, 14, 562–572. [Google Scholar] [CrossRef]
- Stanculescu, D.; Sepulveda, N.; Lim, C.L.; Bergquist, J. Lessons From Heat Stroke for Understanding Myalgic Encephalomyelitis/Chronic Fatigue Syndrome. Front. Neurol. 2021, 12, 789784. [Google Scholar] [CrossRef] [PubMed]
- Sasso, E.M.; Muraki, K.; Eaton-Fitch, N.; Smith, P.; Lesslar, O.L.; Deed, G.; Marshall-Gradisnik, S. Transient Receptor Potential Melastatin 3 Dysfunction in Post COVID-19 Condition and Myalgic Encephalomyelitis/Chronic Fatigue Syndrome Patients. Mol. Med. 2022, 28, 98. [Google Scholar] [CrossRef] [PubMed]
- Vriens, J.; Owsianik, G.; Hofmann, T.; Philipp, S.E.; Stab, J.; Chen, X.; Benoit, M.; Xue, F.; Janssens, A.; Kerselaers, S.; et al. TRPM3 is a Nociceptor Channel Involved in the Detection of Noxious Heat. Neuron 2011, 70, 482–494. [Google Scholar] [CrossRef] [Green Version]
- Caterina, M.J. Transient Receptor Potential Ion Channels as Participants in Thermosensation and Thermoregulation. Am. J. Physiol. Regul. Integr. Comp. Physiol. 2007, 292, R64–R76. [Google Scholar] [CrossRef] [PubMed]
- Okabe, T.; Fujimura, T.; Okajima, J.; Kambayashi, Y.; Aiba, S.; Maruyama, S. First-in-human clinical study of novel technique to diagnose malignant melanoma via thermal conductivity measurements. Sci. Rep. 2019, 9, 3853. [Google Scholar] [CrossRef] [Green Version]
- Wrotek, S.; Brycht, L.; Wrotek, W.; Kozak, W. Fever as a factor contributing to long-term survival in a patient with metastatic melanoma: A case report. Complement. Ther. Med. 2018, 38, 7–10. [Google Scholar] [CrossRef]
- Le, M.; Fernandez-Palomo, C.; McNeill, F.E.; Seymour, C.B.; Rainbow, A.J.; Mothersill, C.E. Exosomes are released by bystander cells exposed to radiation-induced biophoton signals: Reconciling the mechanisms mediating the bystander effect. PLoS ONE 2017, 12, e0173685. [Google Scholar] [CrossRef] [Green Version]
- Blauensteiner, J.; Bertinat, R.; Leon, L.E.; Riederer, M.; Sepulveda, N.; Westermeier, F. Altered endothelial dysfunction-related miRs in plasma from ME/CFS patients. Sci. Rep. 2021, 11, 10604. [Google Scholar] [CrossRef] [PubMed]
- Brenu, E.W.; Ashton, K.J.; van Driel, M.; Staines, D.R.; Peterson, D.; Atkinson, G.M.; Marshall-Gradisnik, S.M. Cytotoxic lymphocyte microRNAs as prospective biomarkers for Chronic Fatigue Syndrome/Myalgic Encephalomyelitis. J. Affect Discord 2012, 141, 261–269. [Google Scholar] [CrossRef] [PubMed]
- Xu, S.; Ding, N.; Pei, H.; Hu, W.; Wei, W.; Zhang, X.; Zhou, G.; Wang, J. MiR-21 is involved in radiation-induced bystander effects. RNA Biol. 2014, 11, 1161–1170. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Xu, S.; Wang, J.; Ding, N.; Hu, W.; Zhang, X.; Wang, B.; Hua, J.; Wei, W.; Zhu, Q. Exosome-mediated microRNA transfer plays a role in radiation-induced bystander effect. RNA Biol. 2015, 12, 1355–1363. [Google Scholar] [CrossRef] [Green Version]
- Melnik, B.C.; John, S.M.; Carrera-Bastos, P.; Schmitz, G. MicroRNA-21-Enriched Exosomes as Epigenetic Regulators in Melanomagenesis and Melanoma Progression: The Impact of Western Lifestyle Factors. Cancers 2020, 12, 2111. [Google Scholar] [CrossRef]
- Lai, J.Y.; Luo, J.; O’Connor, C.; Jing, X.; Nair, V.; Ju, W.; Randolph, A.; Ben-Dov, I.Z.; Matar, R.N.; Briskin, D.; et al. MicroRNA-21 in glomerular injury. J. Am. Soc. 2015, 26, 805–816. [Google Scholar] [CrossRef] [Green Version]
- Zhang, L.; He, S.; Yang, F.; Yu, H.; Xie, W.; Dai, Q.; Zhang, D.; Liu, X.; Zhou, S.; Zhang, K. Hyperoside ameliorates glomerulosclerosis in diabetic nephropathy by downregulating miR-21. Can. J. Physiol. Pharmacol. 2016, 94, 1249–1256. [Google Scholar] [CrossRef]
- Hyde, B.M.; Goldstein, J.A.; Levine, P.H. The Nightingale Research Foundation Review of the Clinical and Scientific Basis of Myalgic Encephalomyelitis/Chronic Fatigue Syndrome; Nightingale Research Foundation: Ottawa, ON, Canada, 1992. [Google Scholar]
- Englebienne, P.; DeMeirleir, K. Chronic Fatigue Syndrome: A Biological Approach; CRC Press: Boca Raton, FL, USA, 2002. [Google Scholar]
- Journal of Chronic Fatigue Syndrome. 1995:1(1)–2007:14(4). Taylor & Francis Publ. Available online: https://www.tandfonline.com/journals/icfs20 (accessed on 17 March 2023).
- Fitzpatrick, T.B. Color Atlas and Synopsis of Clinical Dermatology: Common and Serious Diseases; McGraw-Hill: New York, NY, USA, 2001. [Google Scholar]
- National Research Council, Committee on the Biological Effects of Ionizing Radiation. Health Effects of Exposure to Low Levels of Ionizing Radiation (BEIR V); National Academy Press: Washington, DC, USA, 1990. [Google Scholar]
- Fink, C.A.; Bates, M.N. Melanoma and ionizing radiation: Is there a causal relationship? Radiat. Res. 2005, 164, 701–710. [Google Scholar] [CrossRef]
- Telle-Lamberton, M.; Samson, E.; Caër, S.; Bergot, D.; Bard, D.; Bermann, F.; Gélas, J.M.; Giraud, J.M.; Hubert, P.; Metz-Flamant, C.; et al. External radiation exposure and mortality in a cohort of French nuclear workers. Occup. Environ. Med. 2007, 64, 694–700. [Google Scholar] [CrossRef] [Green Version]
- Rajeevan, M.S.; Murray, J.; Oakley, L.; Lin, J.S.; Unger, E.R. Association of chronic fatigue syndrome with premature telomere attrition. J. Transl. Med. 2018, 16, 44. [Google Scholar] [CrossRef]
- Rochette, P.J.; Brash, D.E. Human telomeres are hypersensitive to UV-induced DNA Damage and refractory to repair. PLoS Genet. 2010, 6, e1000926. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Oikawa, S.; Tada-Oikawa, S.; Kawanishi, S. Site-specific DNA damage at the GGG sequence by UVA involves acceleration of telomere shortening. Biochemistry 2001, 40, 4763–4768. [Google Scholar] [CrossRef] [PubMed]
- Oikawa, S.; Kawanishi, S. Site-specific DNA damage at GGG sequence by oxidative stress may accelerate telomere shortening. FEBS Lett. 1999, 453, 365–368. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shim, G.; Ricoul, M.; Hempel, W.M.; Azzam, E.I.; Sabatier, L. Crosstalk between telomere maintenance and radiation effects: A key player in the process of radiation-induced carcinogenesis. Mutat. Res. Rev. Mutat. Res. 2014, 760, 1–17. [Google Scholar] [CrossRef] [PubMed]
- Samuel, P.; Tsapekos, M.; de Pedro, N.; Liu, A.G.; Lippmeier, J.C.; Chen, S. Ergothioneine mitigates telomere shortening under oxidative stress conditions. J. Diet. Suppl. 2020, 19, 212–225. [Google Scholar] [CrossRef]
- Marozik, P.; Mothersill, C.; Seymour, C.B.; Mosse, I.; Melnov, S. Bystander effects induced by serum from survivors of the Chernobyl accident. Exp. Hematol. 2007, 35, 55–63. [Google Scholar] [CrossRef]
- Mosse, I.; Marozik, P.; Seymour, C.; Mothersill, C. The effect of melanin on the bystander effect in human keratinocytes. Mutat. Res. 2006, 597, 133–137. [Google Scholar] [CrossRef]
- Lad, J.; Rusin, A.; Seymour, C.; Mothersill, C. An investigation into neutron-induced bystander effects: How low can you go? Environ. Res. 2019, 175, 84–99. [Google Scholar] [CrossRef]
- Rusin, A.; Li, M.; Cocchetto, A.; Seymour, C.; Mothersill, C. Radiation exposure and mitochondrial insufficiency in chronic fatigue and immune dysfunction syndrome. Med. Hypotheses 2021, 154, 110647. [Google Scholar] [CrossRef]
- Rusin, A.; Seymour, C.; Cocchetto, A.; Mothersill, C. Commonalities in the Features of Cancer and Chronic Fatigue Syndrome (CFS): Evidence for Stress-Induced Phenotype Instability? Int. J. Mol. Sci. 2022, 23, 691. [Google Scholar] [CrossRef]

Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Cocchetto, A.; Seymour, C.; Mothersill, C. A Proposed New Model to Explain the Role of Low Dose Non-DNA Targeted Radiation Exposure in Chronic Fatigue and Immune Dysfunction Syndrome. Int. J. Mol. Sci. 2023, 24, 6022. https://doi.org/10.3390/ijms24076022
Cocchetto A, Seymour C, Mothersill C. A Proposed New Model to Explain the Role of Low Dose Non-DNA Targeted Radiation Exposure in Chronic Fatigue and Immune Dysfunction Syndrome. International Journal of Molecular Sciences. 2023; 24(7):6022. https://doi.org/10.3390/ijms24076022
Chicago/Turabian StyleCocchetto, Alan, Colin Seymour, and Carmel Mothersill. 2023. "A Proposed New Model to Explain the Role of Low Dose Non-DNA Targeted Radiation Exposure in Chronic Fatigue and Immune Dysfunction Syndrome" International Journal of Molecular Sciences 24, no. 7: 6022. https://doi.org/10.3390/ijms24076022