
Gut Region-Specific Interleukin 1β Induction in Different Myenteric Neuronal Subpopulations of Type 1 Diabetic Rats
 , , and
, , and
Abstract
:1. Introduction
2. Results
2.1. Weight and Glycemic Characteristics of Experimental Rats
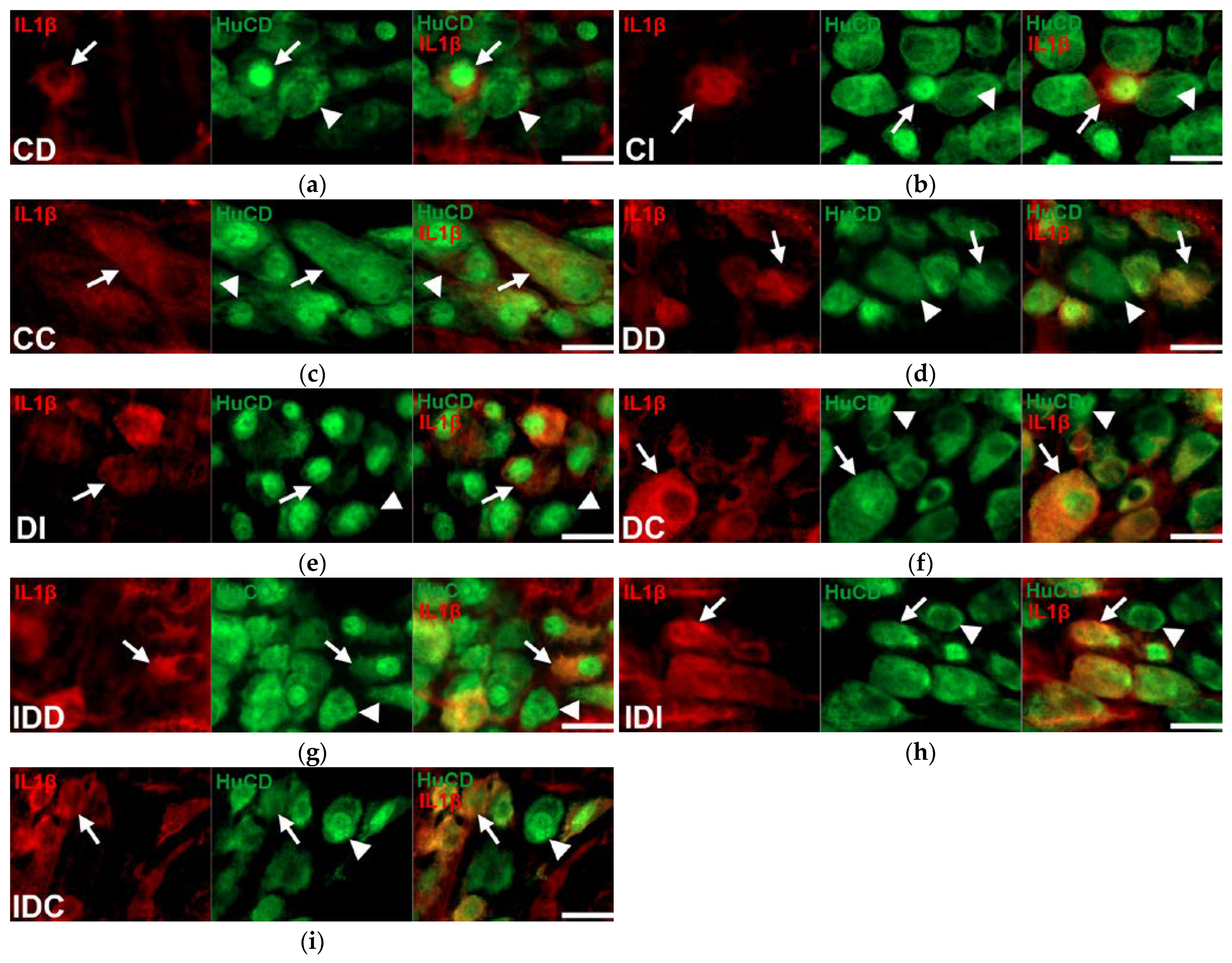
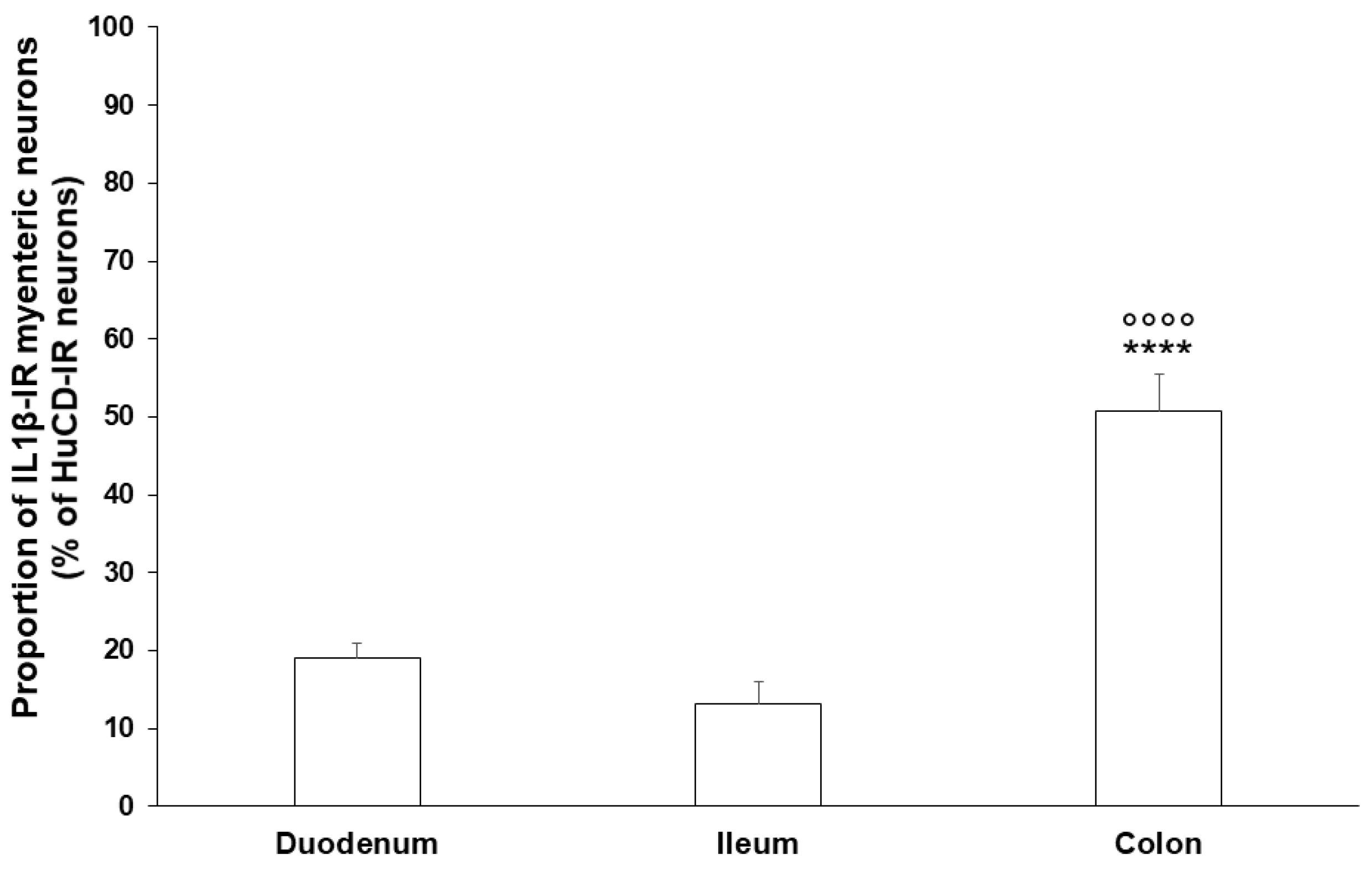
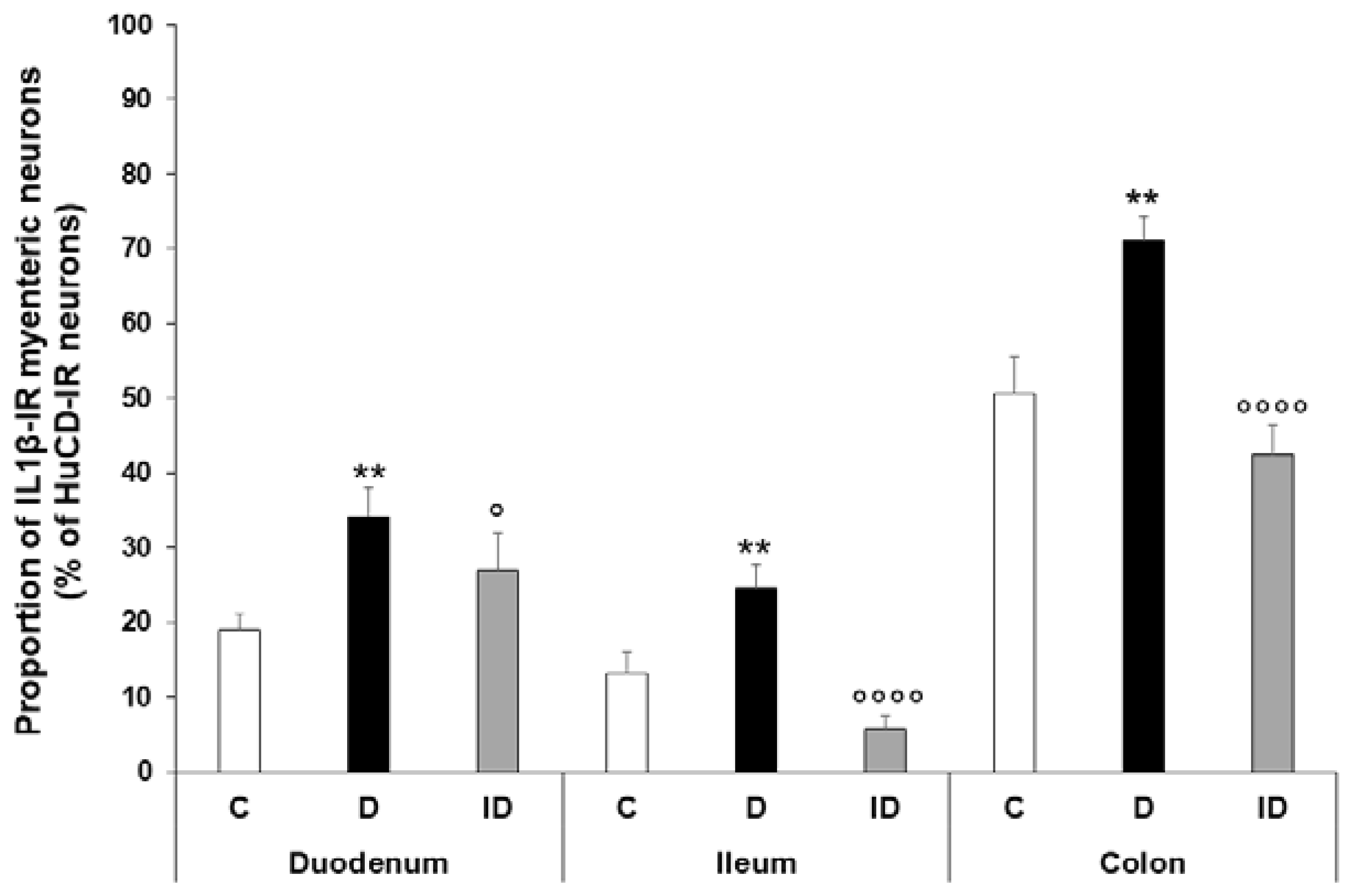
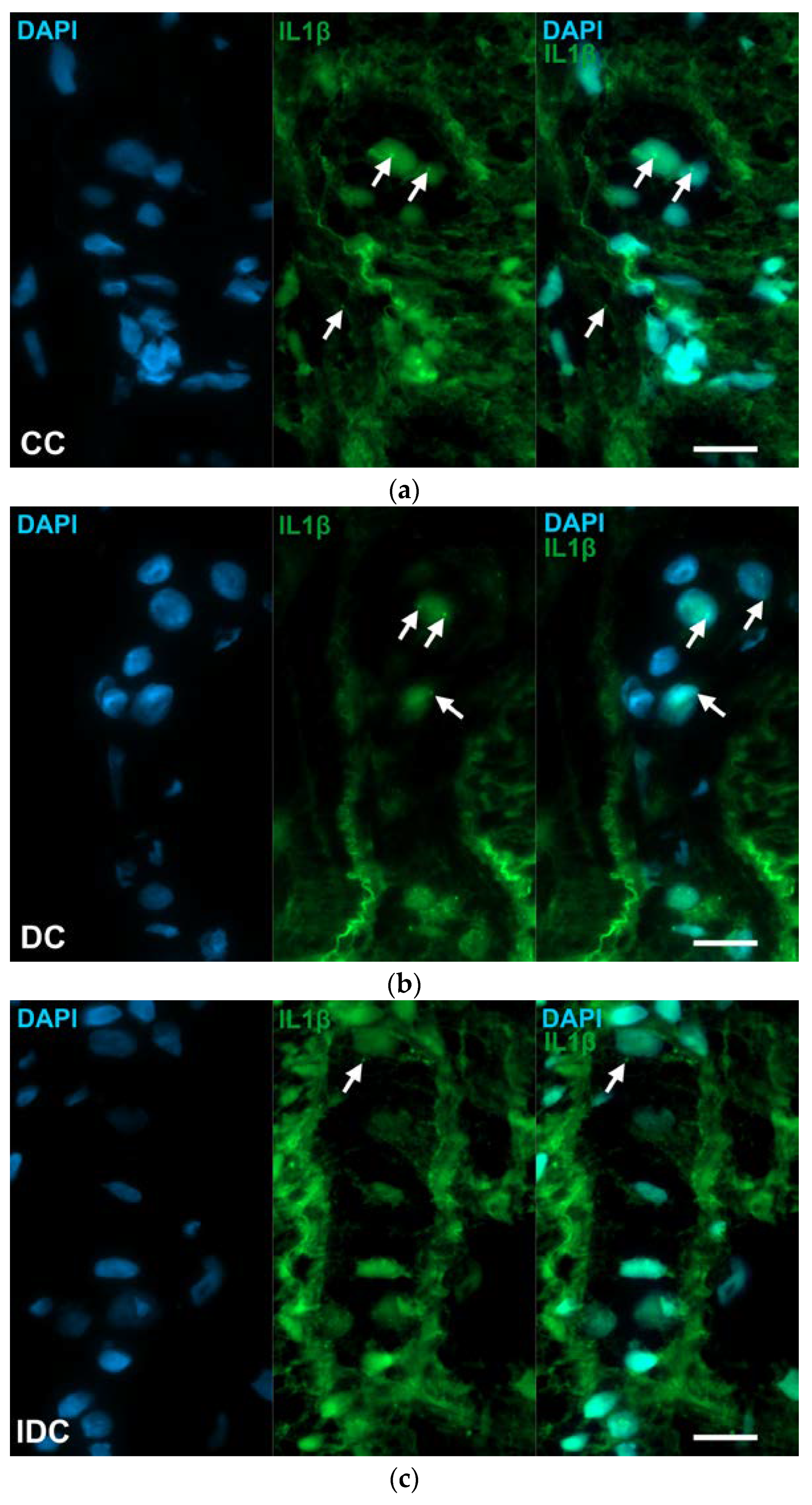
2.2. Gut Region-Specific Presence and Diabetic Induction of IL1β-Immunoreactive Myenteric Neurons
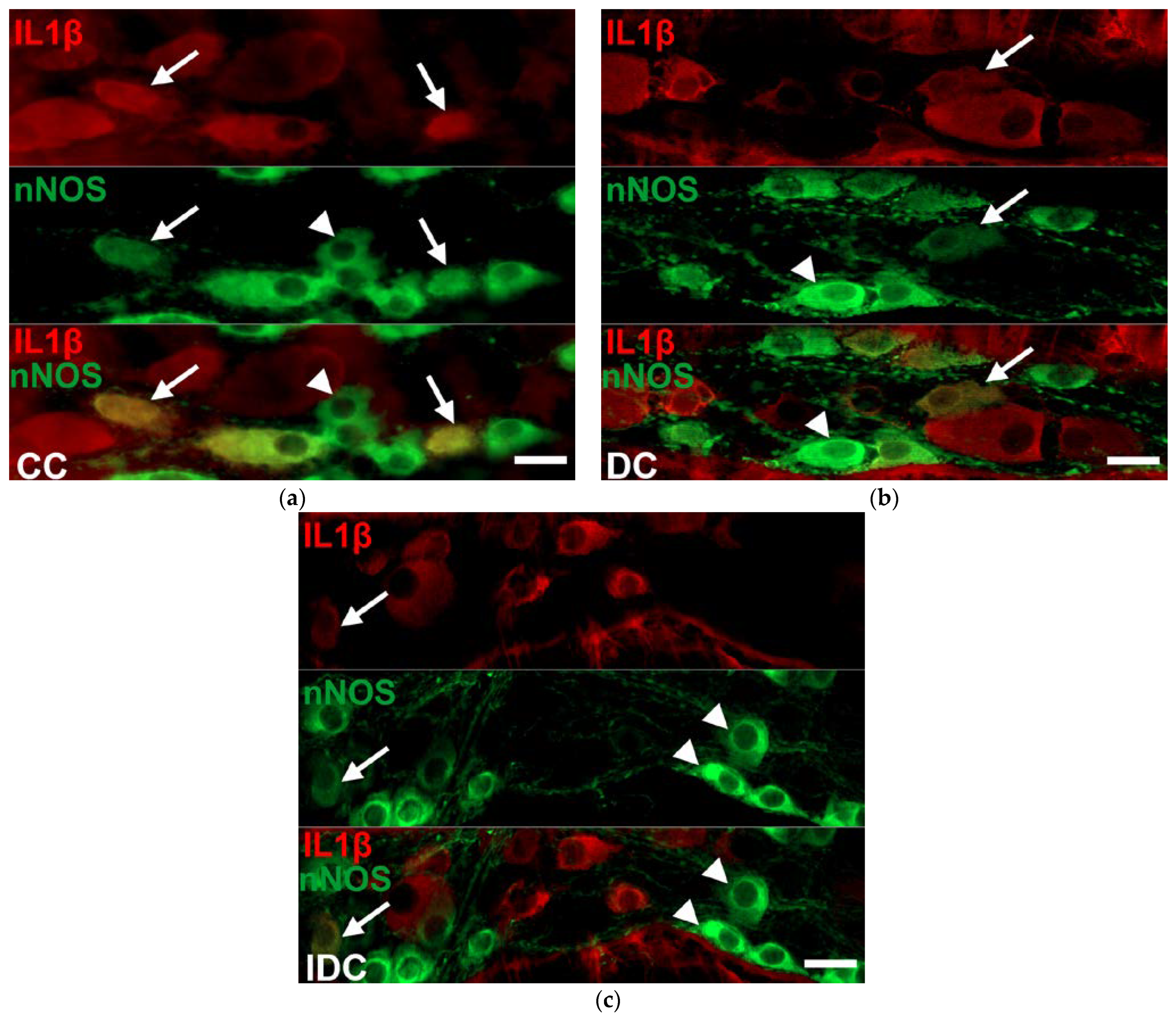
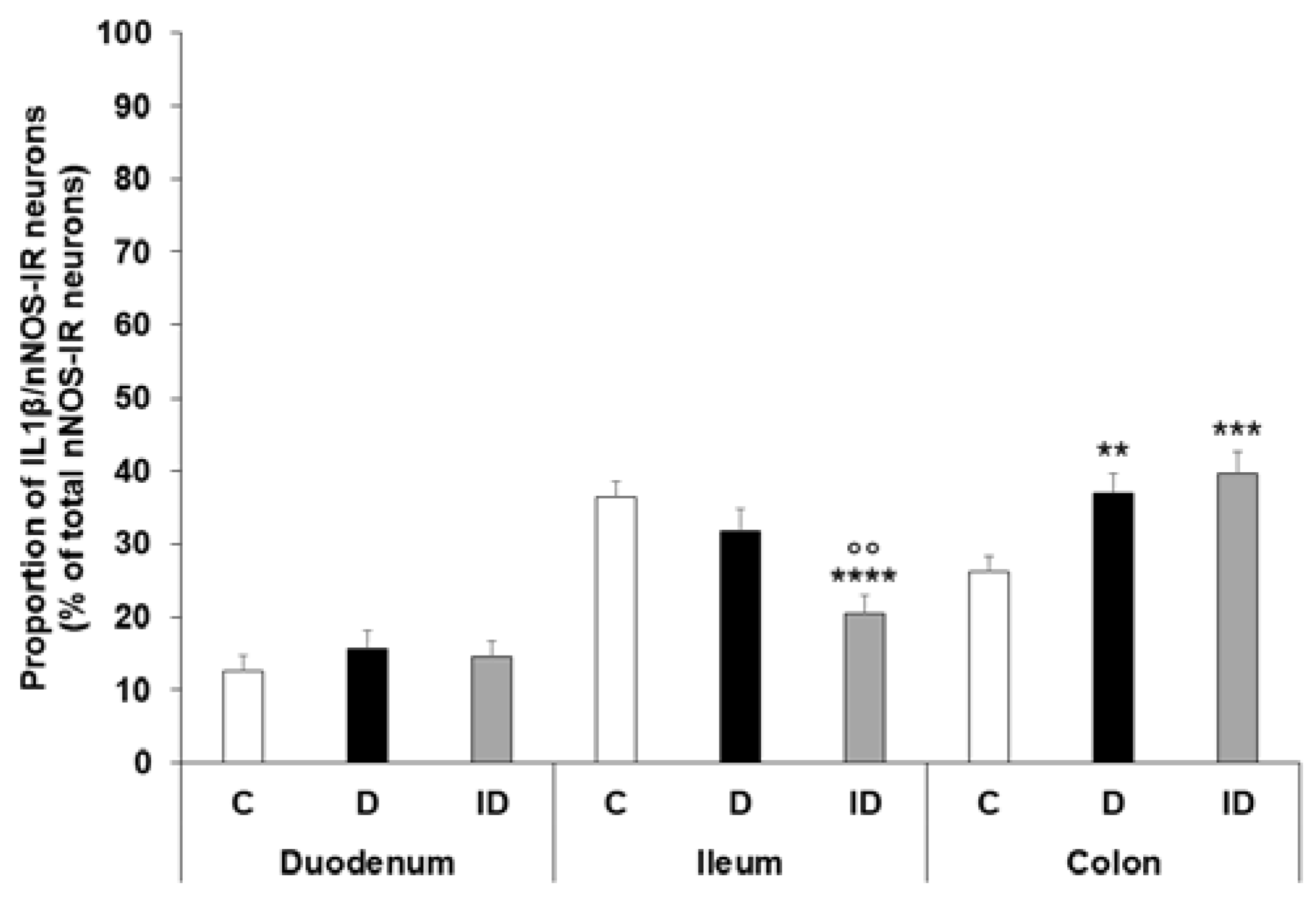
2.3. Region-Dependent Increase of IL1β Immunoreactivity in nNOS-Immunoreactive Myenteric Neurons of Diabetic Rats
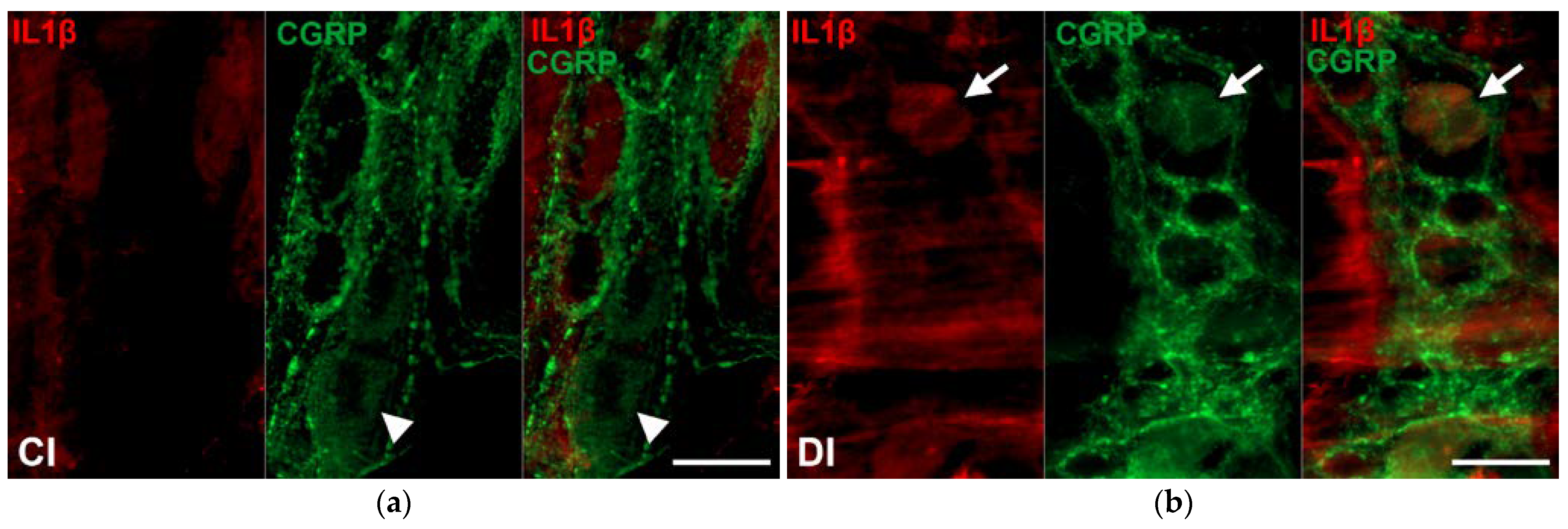
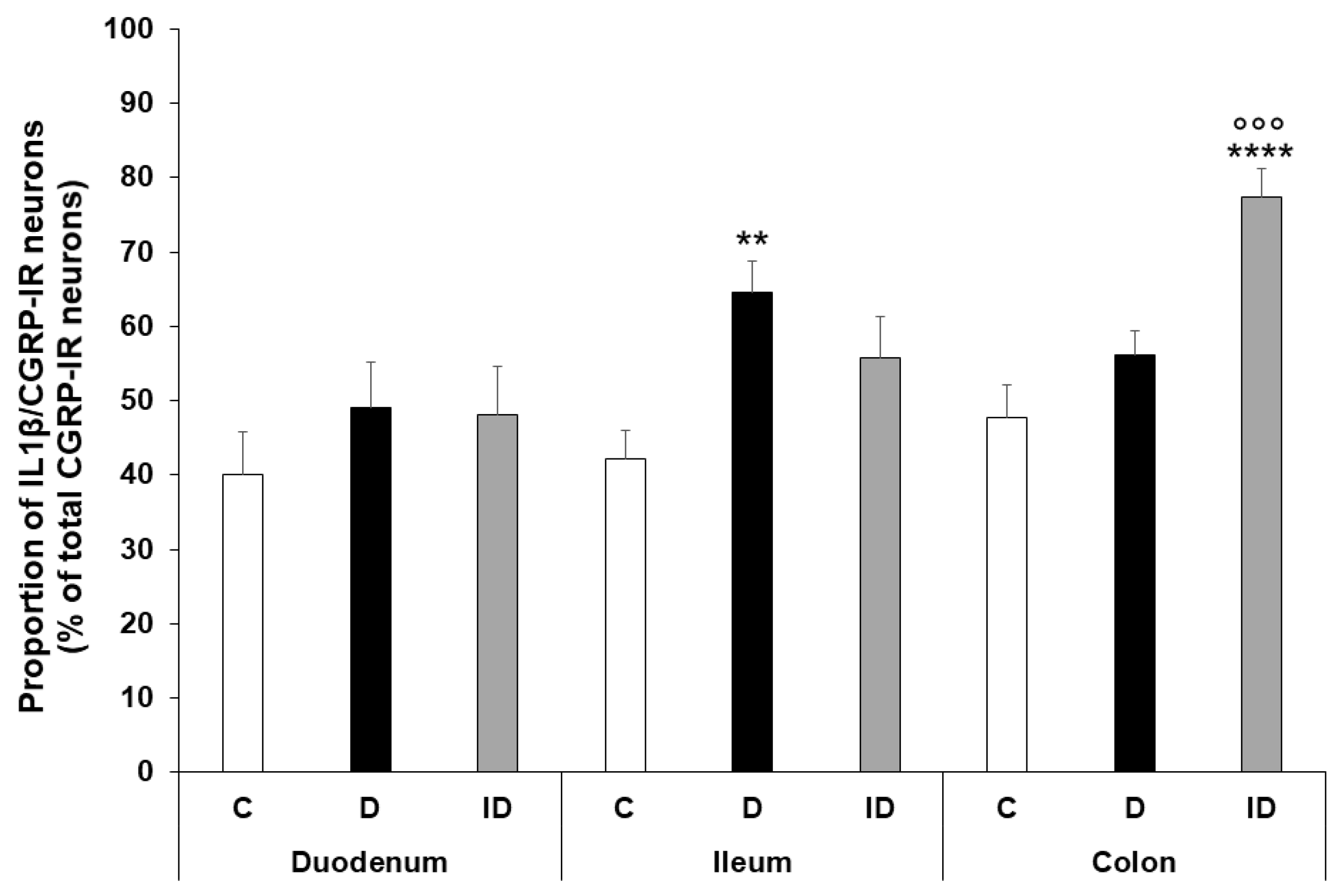
2.4. Region-Dependent Increase of IL1β Immunoreactivity in CGRP-IR Myenteric Neurons of Diabetic Rats
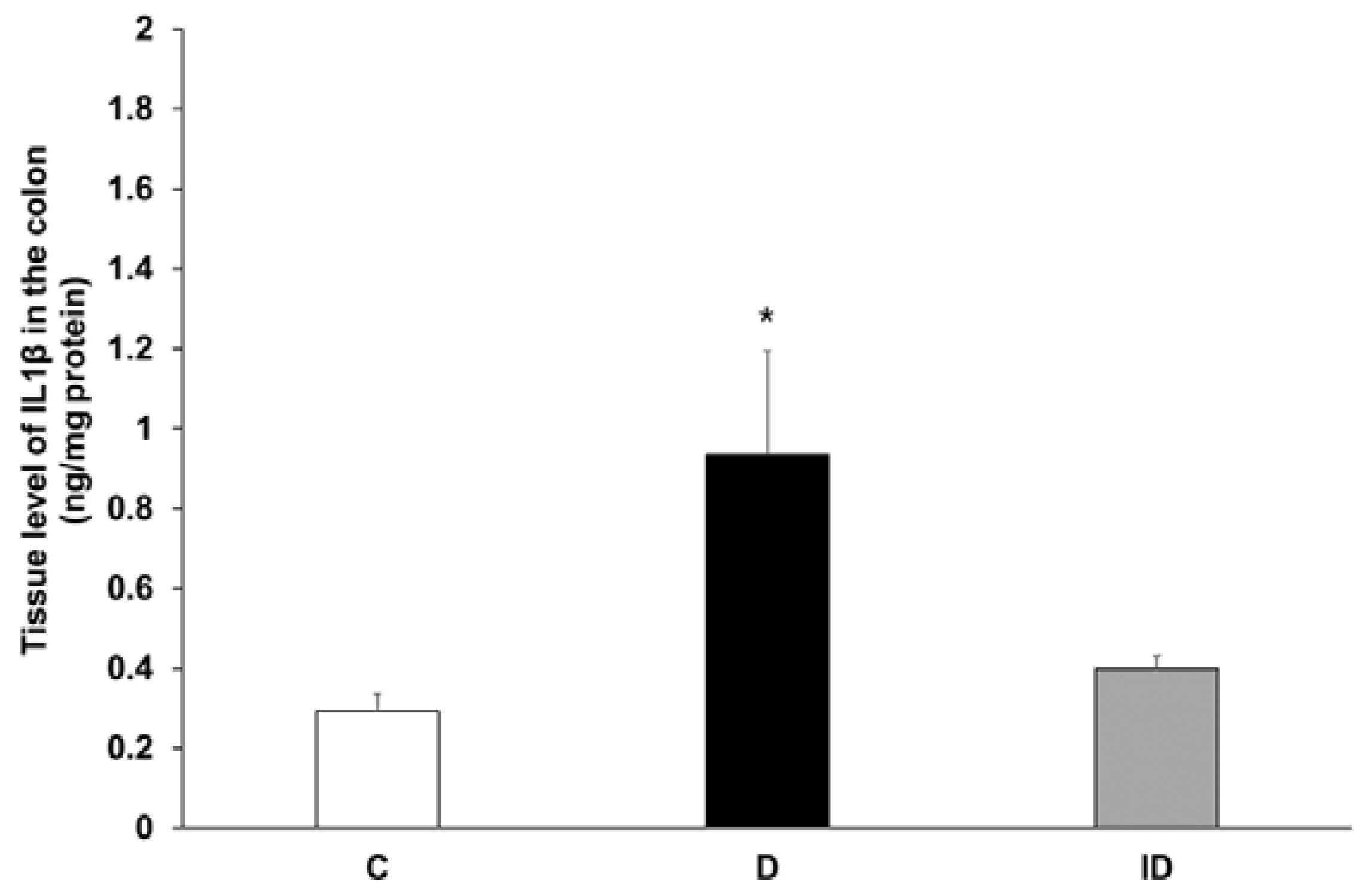
2.5. Increased Tissue Level of IL1β in Colonic Muscle/Myenteric Plexus Homogenates of Diabetics
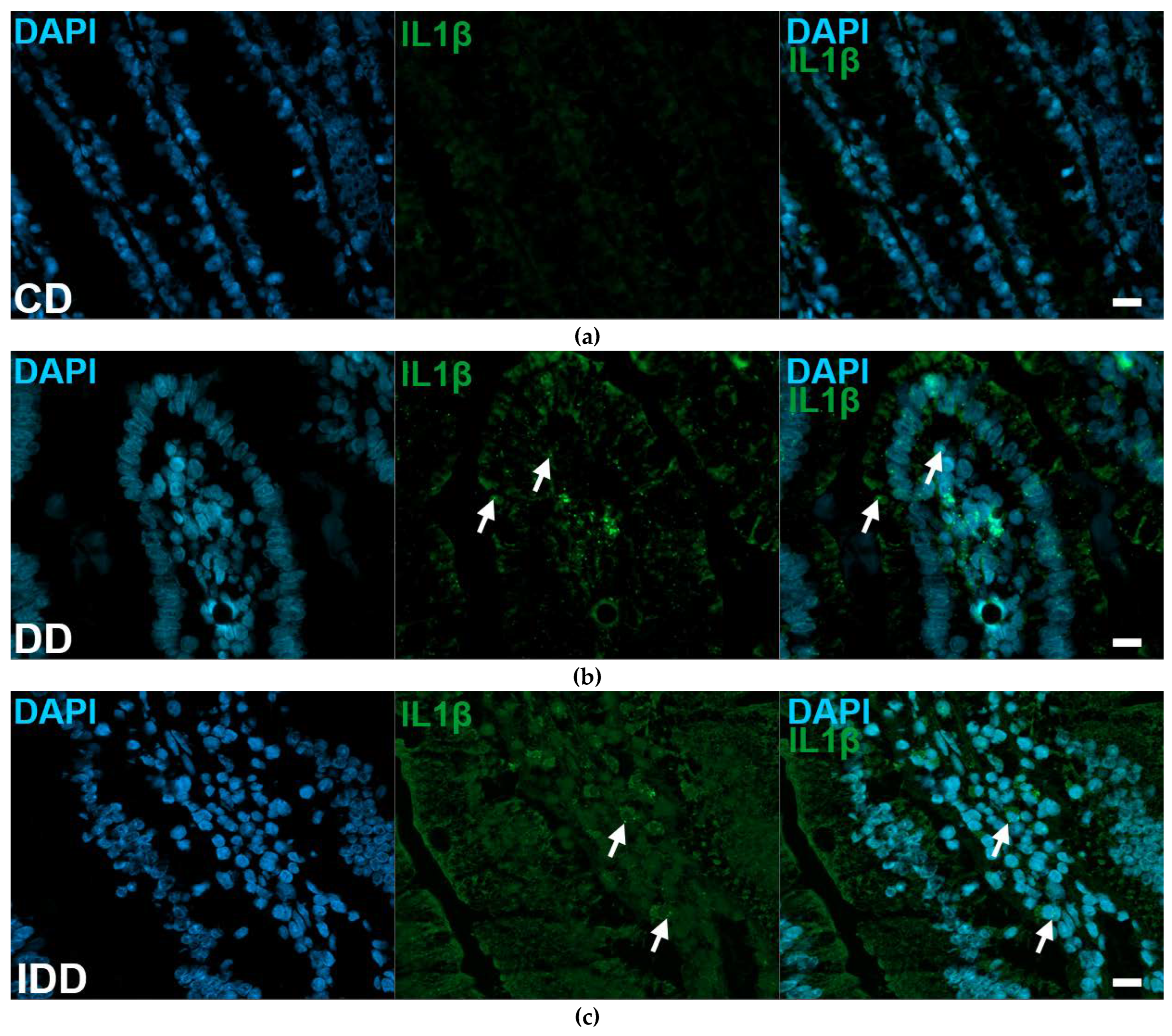
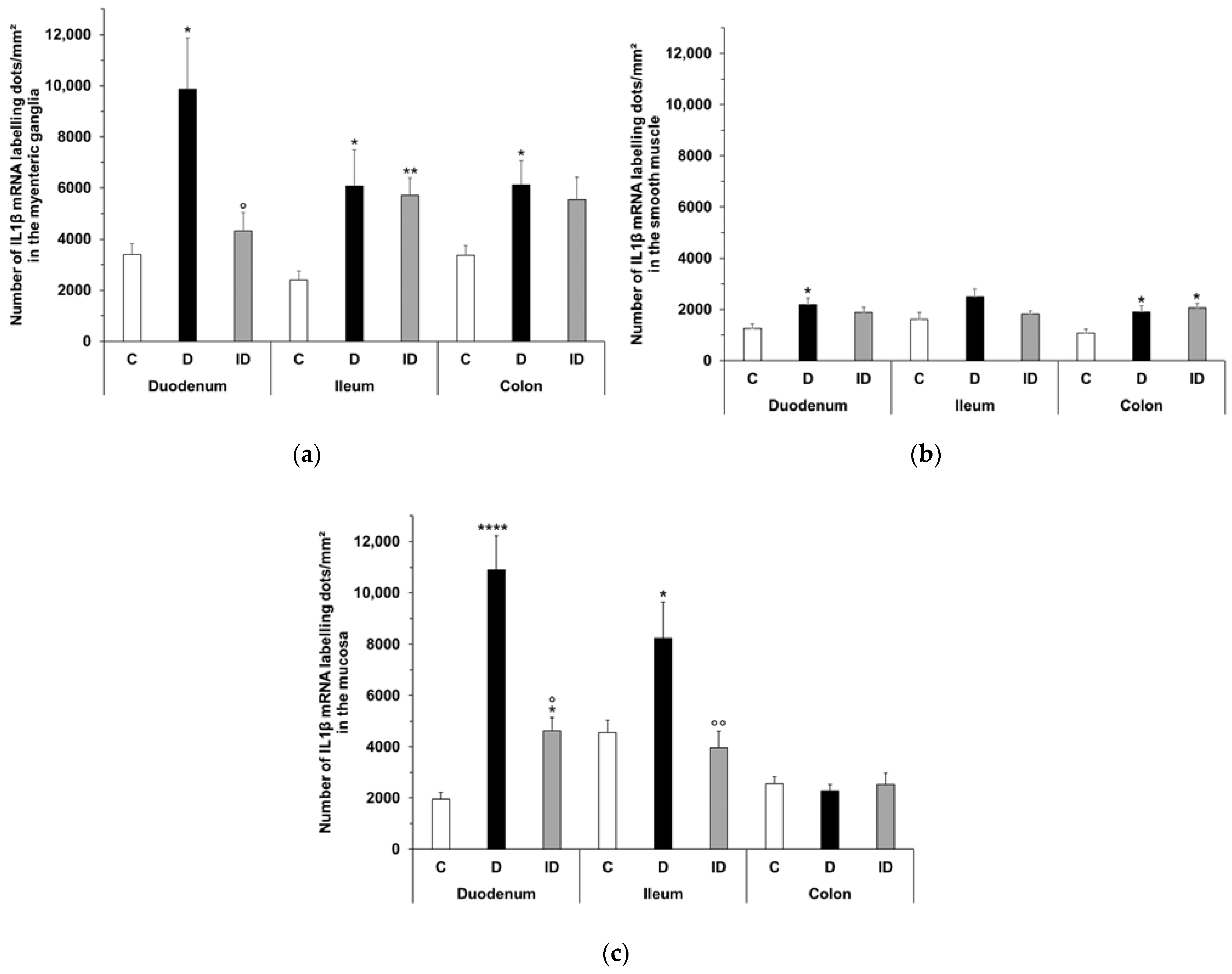
2.6. Quantitative Evaluation of IL1β mRNA Level
3. Discussion
4. Materials and Methods
4.1. Animal Model
4.2. Tissue Handling
4.3. Fluorescent Immunohistochemistry
4.4. Measurement of Tissue IL1β Concentration
4.5. Bradford Protein Micromethod for the Determination of Tissue Protein Content
4.6. RNAscope Multiplex Fluorescent V2 Assay
4.7. Statistical Analysis
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Alexandraki, K.I.; Piperi, C.; Ziakas, P.D.; Apostolopoulos, N.V.; Makrilakis, K.; Syriou, V.; Diamanti-Kandarakis, E.; Kaltsas, G.; Kalofoutis, A. Cytokine secretion in long-standing diabetes mellitus type 1 and 2: Associations with low-grade systemic inflammation. J. Clin. Immunol. 2008, 28, 314–321. [Google Scholar] [CrossRef] [PubMed]
- Chandrakumar, L.; Bagyanszki, M.; Szalai, Z.; Mezei, D.; Bodi, N. Diabetes-Related Induction of the Heme Oxygenase System and Enhanced Colocalization of Heme Oxygenase 1 and 2 with Neuronal Nitric Oxide Synthase in Myenteric Neurons of Different Intestinal Segments. Oxid. Med. Cell. Longev. 2017, 2017, 1890512. [Google Scholar] [CrossRef] [PubMed]
- Kaiko, G.E.; Stappenbeck, T.S. Host-microbe interactions shaping the gastrointestinal environment. Trends Immunol. 2014, 35, 538–548. [Google Scholar] [CrossRef] [Green Version]
- Wirth, R.; Bodi, N.; Maroti, G.; Bagyanszki, M.; Talapka, P.; Fekete, E.; Bagi, Z.; Kovacs, K.L. Regionally distinct alterations in the composition of the gut microbiota in rats with streptozotocin-induced diabetes. PLoS ONE 2014, 9, e110440. [Google Scholar] [CrossRef] [Green Version]
- Izbeki, F.; Wittman, T.; Rosztoczy, A.; Linke, N.; Bodi, N.; Fekete, E.; Bagyanszki, M. Immediate insulin treatment prevents gut motility alterations and loss of nitrergic neurons in the ileum and colon of rats with streptozotocin-induced diabetes. Diabetes Res. Clin. Pract. 2008, 80, 192–198. [Google Scholar] [CrossRef] [Green Version]
- Klinge, M.W.; Sutter, N.; Mark, E.B.; Haase, A.M.; Borghammer, P.; Schlageter, V.; Lund, S.; Fleischer, J.; Knudsen, K.; Drewes, A.M.; et al. Gastric Emptying Time and Volume of the Small Intestine as Objective Markers in Patients with Symptoms of Diabetic Enteropathy. J. Neurogastroenterol. Motil. 2021, 27, 390–399. [Google Scholar] [CrossRef]
- Yarandi, S.S.; Srinivasan, S. Diabetic gastrointestinal motility disorders and the role of enteric nervous system: Current status and future directions. Neurogastroenterol. Motil. 2014, 26, 611–624. [Google Scholar] [CrossRef] [Green Version]
- Caputi, V.; Popov, J.; Giron, M.C.; O’Mahony, S. Gut Microbiota as a Mediator of Host Neuro-Immune Interactions: Implications in Neuroinflammatory Disorders. Mod. Trends Psychiatry 2021, 32, 40–57. [Google Scholar] [CrossRef]
- Al-Sadi, R.M.; Ma, T.Y. IL-1beta causes an increase in intestinal epithelial tight junction permeability. J. Immunol. 2007, 178, 4641–4649. [Google Scholar] [CrossRef] [Green Version]
- Jarret, A.; Jackson, R.; Duizer, C.; Healy, M.E.; Zhao, J.; Rone, J.M.; Bielecki, P.; Sefik, E.; Roulis, M.; Rice, T.; et al. Enteric Nervous System-Derived IL-18 Orchestrates Mucosal Barrier Immunity. Cell 2020, 180, 50–63 e12. [Google Scholar] [CrossRef]
- Ruder, B.; Atreya, R.; Becker, C. Tumour Necrosis Factor Alpha in Intestinal Homeostasis and Gut Related Diseases. Int. J. Mol. Sci. 2019, 20, 1887. [Google Scholar] [CrossRef] [Green Version]
- Wei, H.X.; Wang, B.; Li, B. IL-10 and IL-22 in Mucosal Immunity: Driving Protection and Pathology. Front. Immunol. 2020, 11, 1315. [Google Scholar] [CrossRef] [PubMed]
- Bodi, N.; Chandrakumar, L.; Al Doghmi, A.; Mezei, D.; Szalai, Z.; Barta, B.P.; Balazs, J.; Bagyanszki, M. Intestinal Region-Specific and Layer-Dependent Induction of TNFalpha in Rats with Streptozotocin-Induced Diabetes and after Insulin Replacement. Cells 2021, 10, 2410. [Google Scholar] [CrossRef] [PubMed]
- Barta, B.P.; Onhausz, B.; Al Doghmi, A.; Szalai, Z.; Balazs, J.; Bagyanszki, M.; Bodi, N. Gut region-specific TNFR expression: TNFR2 is more affected than TNFR1 in duodenal myenteric ganglia of diabetic rats. World J. Diabetes 2023, 14, 48–61. [Google Scholar] [CrossRef]
- McEntee, C.P.; Finlay, C.M.; Lavelle, E.C. Divergent Roles for the IL-1 Family in Gastrointestinal Homeostasis and Inflammation. Front. Immunol. 2019, 10, 1266. [Google Scholar] [CrossRef] [Green Version]
- Lopez-Castejon, G.; Brough, D. Understanding the mechanism of IL-1beta secretion. Cytokine Growth Factor Rev. 2011, 22, 189–195. [Google Scholar] [CrossRef]
- Mantovani, A.; Dinarello, C.A.; Molgora, M.; Garlanda, C. Interleukin-1 and Related Cytokines in the Regulation of Inflammation and Immunity. Immunity 2019, 50, 778–795. [Google Scholar] [CrossRef] [Green Version]
- Limbert, C. Type 1 diabetes—An auto-inflammatory disease: A new concept, new therapeutical strategies. In Proceedings of the 7th European Workshop on Immune-Mediated Inflammatory Diseases, Noordwijk aan Zee, The Netherlands, 28 November 2012. [Google Scholar]
- Broderick, L.; Hoffman, H.M. IL-1 and autoinflammatory disease: Biology, pathogenesis and therapeutic targeting. Nat. Rev. Rheumatol. 2022, 18, 448–463. [Google Scholar] [CrossRef]
- Kaneko, N.; Kurata, M.; Yamamoto, T.; Morikawa, S.; Masumoto, J. The role of interleukin-1 in general pathology. Inflamm. Regen. 2019, 39, 12. [Google Scholar] [CrossRef] [Green Version]
- Mandrup-Poulsen, T.; Pickersgill, L.; Donath, M.Y. Blockade of interleukin 1 in type 1 diabetes mellitus. Nat. Rev. Endocrinol. 2010, 6, 158–166. [Google Scholar] [CrossRef]
- Peiro, C.; Lorenzo, O.; Carraro, R.; Sanchez-Ferrer, C.F. IL-1beta Inhibition in Cardiovascular Complications Associated to Diabetes Mellitus. Front. Pharmacol. 2017, 8, 363. [Google Scholar] [CrossRef] [Green Version]
- Kaminsky, L.W.; Al-Sadi, R.; Ma, T.Y. IL-1beta and the Intestinal Epithelial Tight Junction Barrier. Front. Immunol. 2021, 12, 767456. [Google Scholar] [CrossRef] [PubMed]
- Ren, K.; Torres, R. Role of interleukin-1beta during pain and inflammation. Brain Res. Rev. 2009, 60, 57–64. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gougeon, P.Y.; Lourenssen, S.; Han, T.Y.; Nair, D.G.; Ropeleski, M.J.; Blennerhassett, M.G. The pro-inflammatory cytokines IL-1beta and TNFalpha are neurotrophic for enteric neurons. J. Neurosci. 2013, 33, 3339–3351. [Google Scholar] [CrossRef] [Green Version]
- Tjwa, E.T.; Bradley, J.M.; Keenan, C.M.; Kroese, A.B.; Sharkey, K.A. Interleukin-1beta activates specific populations of enteric neurons and enteric glia in the guinea pig ileum and colon. Am. J. Physiol. Gastrointest. Liver Physiol. 2003, 285, G1268-1276. [Google Scholar] [CrossRef]
- Xia, Y.; Hu, H.Z.; Liu, S.; Ren, J.; Zafirov, D.H.; Wood, J.D. IL-1beta and IL-6 excite neurons and suppress nicotinic and noradrenergic neurotransmission in guinea pig enteric nervous system. J. Clin. Investig. 1999, 103, 1309–1316. [Google Scholar] [CrossRef]
- Azpiroz, F.; Malagelada, C. Diabetic neuropathy in the gut: Pathogenesis and diagnosis. Diabetologia 2016, 59, 404–408. [Google Scholar] [CrossRef]
- Uranga-Ocio, J.A.; Bastus-Diez, S.; Delkader-Palacios, D.; Garcia-Cristobal, N.; Leal-Garcia, M.A.; Abalo-Delgado, R. Enteric neuropathy associated to diabetes mellitus. Rev. Esp. Enferm. Dig. 2015, 107, 366–373. [Google Scholar]
- Cellek, S. Point of NO return for nitrergic nerves in diabetes: A new insight into diabetic complications. Curr. Pharm. Des. 2004, 10, 3683–3695. [Google Scholar] [CrossRef]
- Demedts, I.; Masaoka, T.; Kindt, S.; De Hertogh, G.; Geboes, K.; Farre, R.; Vanden Berghe, P.; Tack, J. Gastrointestinal motility changes and myenteric plexus alterations in spontaneously diabetic biobreeding rats. J. Neurogastroenterol. Motil. 2013, 19, 161–170. [Google Scholar] [CrossRef]
- Noorian, A.R.; Taylor, G.M.; Annerino, D.M.; Greene, J.G. Neurochemical phenotypes of myenteric neurons in the rhesus monkey. J. Comp. Neurol. 2011, 519, 3387–3401. [Google Scholar] [CrossRef] [Green Version]
- Qu, Z.D.; Thacker, M.; Castelucci, P.; Bagyanszki, M.; Epstein, M.L.; Furness, J.B. Immunohistochemical analysis of neuron types in the mouse small intestine. Cell Tissue Res. 2008, 334, 147–161. [Google Scholar] [CrossRef]
- Bodi, N.; Szalai, Z.; Bagyanszki, M. Nitrergic Enteric Neurons in Health and Disease-Focus on Animal Models. Int. J. Mol. Sci. 2019, 20, 2003. [Google Scholar] [CrossRef] [Green Version]
- Boyer, L.; Sidpra, D.; Jevon, G.; Buchan, A.M.; Jacobson, K. Differential responses of VIPergic and nitrergic neurons in paediatric patients with Crohn’s disease. Auton. Neurosci. 2007, 134, 106–114. [Google Scholar] [CrossRef] [PubMed]
- Burlinski, P.J.; Rychlik, A.; Calka, J. Effects of inflammation and axotomy on expression of acetylcholine transferase and nitric oxide synthetase within the cocaine- and amphetamine-regulated transcript-immunoreactive neurons of the porcine descending colon. J. Comp. Pathol. 2014, 150, 287–296. [Google Scholar] [CrossRef]
- Furness, J.B.; Jones, C.; Nurgali, K.; Clerc, N. Intrinsic primary afferent neurons and nerve circuits within the intestine. Prog. Neurobiol. 2004, 72, 143–164. [Google Scholar] [CrossRef]
- Hibberd, T.J.; Yew, W.P.; Dodds, K.N.; Xie, Z.; Travis, L.; Brookes, S.J.; Costa, M.; Hu, H.; Spencer, N.J. Quantification of CGRP-immunoreactive myenteric neurons in mouse colon. J. Comp. Neurol. 2022, 530, 3209–3225. [Google Scholar] [CrossRef] [PubMed]
- Chandrasekharan, B.; Srinivasan, S. Diabetes and the enteric nervous system. Neurogastroenterol. Motil. 2007, 19, 951–960. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Domenech, A.; Pasquinelli, G.; De Giorgio, R.; Gori, A.; Bosch, F.; Pumarola, M.; Jimenez, M. Morphofunctional changes underlying intestinal dysmotility in diabetic RIP-I/hIFNbeta transgenic mice. Int. J. Exp. Pathol. 2011, 92, 400–412. [Google Scholar] [CrossRef]
- Zandecki, M.; Vanden Berghe, P.; Depoortere, I.; Geboes, K.; Peeters, T.; Janssens, J.; Tack, J. Characterization of myenteric neuropathy in the jejunum of spontaneously diabetic BB-rats. Neurogastroenterol. Motil. 2008, 20, 818–828. [Google Scholar] [CrossRef]
- Copray, J.C.; Mantingh, I.; Brouwer, N.; Biber, K.; Kust, B.M.; Liem, R.S.; Huitinga, I.; Tilders, F.J.; Van Dam, A.M.; Boddeke, H.W. Expression of interleukin-1 beta in rat dorsal root ganglia. J. Neuroimmunol. 2001, 118, 203–211. [Google Scholar] [CrossRef]
- Guo, W.; Wang, H.; Watanabe, M.; Shimizu, K.; Zou, S.; LaGraize, S.C.; Wei, F.; Dubner, R.; Ren, K. Glial-cytokine-neuronal interactions underlying the mechanisms of persistent pain. J. Neurosci. 2007, 27, 6006–6018. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Brown, H.; Esterhazy, D. Intestinal immune compartmentalization: Implications of tissue specific determinants in health and disease. Mucosal Immunol. 2021, 14, 1259–1270. [Google Scholar] [CrossRef]
- Sanders, L.M.; Henderson, C.E.; Hong, M.Y.; Barhoumi, R.; Burghardt, R.C.; Carroll, R.J.; Turner, N.D.; Chapkin, R.S.; Lupton, J.R. Pro-oxidant environment of the colon compared to the small intestine may contribute to greater cancer susceptibility. Cancer Lett. 2004, 208, 155–161. [Google Scholar] [CrossRef] [PubMed]
- Bodi, N.; Egyed-Kolumban, A.; Onhausz, B.; Barta, B.P.; Doghmi, A.A.; Balazs, J.; Szalai, Z.; Bagyanszki, M. Intestinal Region-Dependent Alterations of Toll-Like Receptor 4 Expression in Myenteric Neurons of Type 1 Diabetic Rats. Biomedicines 2023, 11, 129. [Google Scholar] [CrossRef]
- Price, A.E.; Shamardani, K.; Lugo, K.A.; Deguine, J.; Roberts, A.W.; Lee, B.L.; Barton, G.M. A Map of Toll-like Receptor Expression in the Intestinal Epithelium Reveals Distinct Spatial, Cell Type-Specific, and Temporal Patterns. Immunity 2018, 49, 560–575 e566. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Honma, K.; Machida, C.; Mochizuki, K.; Goda, T. Glucose and TNF enhance expression of TNF and IL1B, and histone H3 acetylation and K4/K36 methylation, in juvenile macrophage cells. Gene 2020, 763S, 100034. [Google Scholar] [CrossRef] [PubMed]
- Maedler, K.; Sergeev, P.; Ris, F.; Oberholzer, J.; Joller-Jemelka, H.I.; Spinas, G.A.; Kaiser, N.; Halban, P.A.; Donath, M.Y. Glucose-induced beta cell production of IL-1beta contributes to glucotoxicity in human pancreatic islets. J. Clin. Investig. 2002, 110, 851–860. [Google Scholar] [CrossRef]
- Flodstrom, M.; Welsh, N.; Eizirik, D.L. Cytokines activate the nuclear factor kappa B (NF-kappa B) and induce nitric oxide production in human pancreatic islets. FEBS Lett. 1996, 385, 4–6. [Google Scholar] [CrossRef] [Green Version]
- Gruber, H.E.; Hoelscher, G.L.; Bethea, S.; Hanley, E.N., Jr. Interleukin 1-beta upregulates brain-derived neurotrophic factor, neurotrophin 3 and neuropilin 2 gene expression and NGF production in annulus cells. Biotech. Histochem. 2012, 87, 506–511. [Google Scholar] [CrossRef]
- Gruber, H.E.; Jones, B.; Marrero, E.; Hanley, E.N., Jr. Proinflammatory Cytokines IL-1beta and TNF-alpha Influence Human Annulus Cell Signaling Cues for Neurite Growth: In Vitro Coculture Studies. Spine 2017, 42, 1529–1537. [Google Scholar] [CrossRef]
- Park, S.Y.; Kang, M.J.; Han, J.S. Interleukin-1 beta promotes neuronal differentiation through the Wnt5a/RhoA/JNK pathway in cortical neural precursor cells. Mol. Brain 2018, 11, 39. [Google Scholar] [CrossRef] [PubMed]
- Cho, J.M.; Chang, S.Y.; Kim, D.B.; Needs, P.W.; Jo, Y.H.; Kim, M.J. Effects of physiological quercetin metabolites on interleukin-1beta-induced inducible NOS expression. J. Nutr. Biochem. 2012, 23, 1394–1402. [Google Scholar] [CrossRef]
- Kim, M.J.; Ryu, G.R.; Kang, J.H.; Sim, S.S.; Min, D.S.; Rhie, D.J.; Yoon, S.H.; Hahn, S.J.; Jeong, I.K.; Hong, K.J.; et al. Inhibitory effects of epicatechin on interleukin-1beta-induced inducible nitric oxide synthase expression in RINm5F cells and rat pancreatic islets by down-regulation of NF-kappaB activation. Biochem. Pharmacol. 2004, 68, 1775–1785. [Google Scholar] [CrossRef]
- Shotton, H.R.; Broadbent, S.; Lincoln, J. Prevention and partial reversal of diabetes-induced changes in enteric nerves of the rat ileum by combined treatment with alpha-lipoic acid and evening primrose oil. Auton. Neurosci. 2004, 111, 57–65. [Google Scholar] [CrossRef]
- Spangeus, A.; Suhr, O.; El-Salhy, M. Diabetic state affects the innervation of gut in an animal model of human type 1 diabetes. Histol. Histopathol. 2000, 15, 739–744. [Google Scholar] [CrossRef] [PubMed]
- Bulc, M.; Calka, J.; Palus, K. Effect of Streptozotocin-Inducted Diabetes on the Pathophysiology of Enteric Neurons in the Small Intestine Based on the Porcine Diabetes Model. Int. J. Mol. Sci. 2020, 21, 2047. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hou, L.; Li, W.; Wang, X. Mechanism of interleukin-1 beta-induced calcitonin gene-related peptide production from dorsal root ganglion neurons of neonatal rats. J. Neurosci. Res. 2003, 73, 188–197. [Google Scholar] [CrossRef] [PubMed]
- Khan, I.; al-Awadi, F.M. Colonic muscle enhances the production of interleukin-1 beta messenger RNA in experimental colitis. Gut 1997, 40, 307–312. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Khan, I.; Blennerhassett, M.G.; Kataeva, G.V.; Collins, S.M. Interleukin 1 beta induces the expression of interleukin 6 in rat intestinal smooth muscle cells. Gastroenterology 1995, 108, 1720–1728. [Google Scholar] [CrossRef]
- Wirth, R.; Bódi, N.; Szalai, Z.; Chandrakumar, L.; Maróti, G.; Kovács, L.K.; Bagi, Z.; Mezei, D.; Balázs, J.; Bagyánszki, M. Perturbation of the mucosa-associated anaerobic gut microbiota in streptozotocin-induced diabetic rats. Acta Biol. Szeged. 2021, 65, 75–84. [Google Scholar] [CrossRef]
- Benedetti, F.; Curreli, S.; Zella, D. Mycoplasmas-Host Interaction: Mechanisms of Inflammation and Association with Cellular Transformation. Microorganisms 2020, 8, 1351. [Google Scholar] [CrossRef]
- Dinarello, C.A. Blocking interleukin-1beta in acute and chronic autoinflammatory diseases. J. Intern. Med. 2011, 269, 16–28. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jancso, Z.; Bodi, N.; Borsos, B.; Fekete, E.; Hermesz, E. Gut region-specific accumulation of reactive oxygen species leads to regionally distinct activation of antioxidant and apoptotic marker molecules in rats with STZ-induced diabetes. Int. J. Biochem. Cell Biol. 2015, 62, 125–131. [Google Scholar] [CrossRef] [PubMed] [Green Version]












{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Weight (g) | Blood Glucose Level (mmol/L) | |||
|---|---|---|---|---|
| Initial | Final | Initial | Final (Average) | |
| Controls (n = 16) | 206 ± 2.48 | 445.6 ± 13.58 c | 4.82 ± 0.29 | 5.81 ± 0.1 |
| Diabetics (n = 15) | 206.1 ± 2.57 | 317.7 ± 13.34 a,e | 5.47 ± 0.42 | 27.05 ± 1.12 c,e |
| Insulin-treated diabetics (n = 14) | 216.1 ± 5.37 | 432.1 ± 15.12 c,g | 5.21 ± 0.42 | 12.57 ± 1.19 b,d,f |
| Duodenum (dots/mm2) | Ileum (dots/mm2) | Colon (dots/mm2) | |
|---|---|---|---|
| Myenteric ganglia | 3400 ± 429 | 2420 ± 486.7 | 3361 ± 279.5 |
| Smooth muscle | 1251 ± 181.5 b | 1629 ± 257.7 | 1080 ± 146 c |
| Mucosa | 1948 ± 281.4 | 4558 ± 486.7 a,e,f | 2549 ± 279.5 d,g |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
AL Doghmi, A.; Barta, B.P.; Egyed-Kolumbán, A.; Onhausz, B.; Kiss, S.; Balázs, J.; Szalai, Z.; Bagyánszki, M.; Bódi, N. Gut Region-Specific Interleukin 1β Induction in Different Myenteric Neuronal Subpopulations of Type 1 Diabetic Rats. Int. J. Mol. Sci. 2023, 24, 5804. https://doi.org/10.3390/ijms24065804
AL Doghmi A, Barta BP, Egyed-Kolumbán A, Onhausz B, Kiss S, Balázs J, Szalai Z, Bagyánszki M, Bódi N. Gut Region-Specific Interleukin 1β Induction in Different Myenteric Neuronal Subpopulations of Type 1 Diabetic Rats. International Journal of Molecular Sciences. 2023; 24(6):5804. https://doi.org/10.3390/ijms24065804
Chicago/Turabian StyleAL Doghmi, Afnan, Bence Pál Barta, Abigél Egyed-Kolumbán, Benita Onhausz, Szilvia Kiss, János Balázs, Zita Szalai, Mária Bagyánszki, and Nikolett Bódi. 2023. "Gut Region-Specific Interleukin 1β Induction in Different Myenteric Neuronal Subpopulations of Type 1 Diabetic Rats" International Journal of Molecular Sciences 24, no. 6: 5804. https://doi.org/10.3390/ijms24065804