

Triazine Herbicides Risk Management Strategies on Environmental and Human Health Aspects Using In-Silico Methods
Abstract
:1. Introduction
2. Results
2.1. Construction and Evaluation of the Single-Effect and Comprehensive-Effect 3D-QSAR Models of Herbicidal Functionality Properties, Microbial Degradability, and Human Lung Cytotoxicity of S-THs
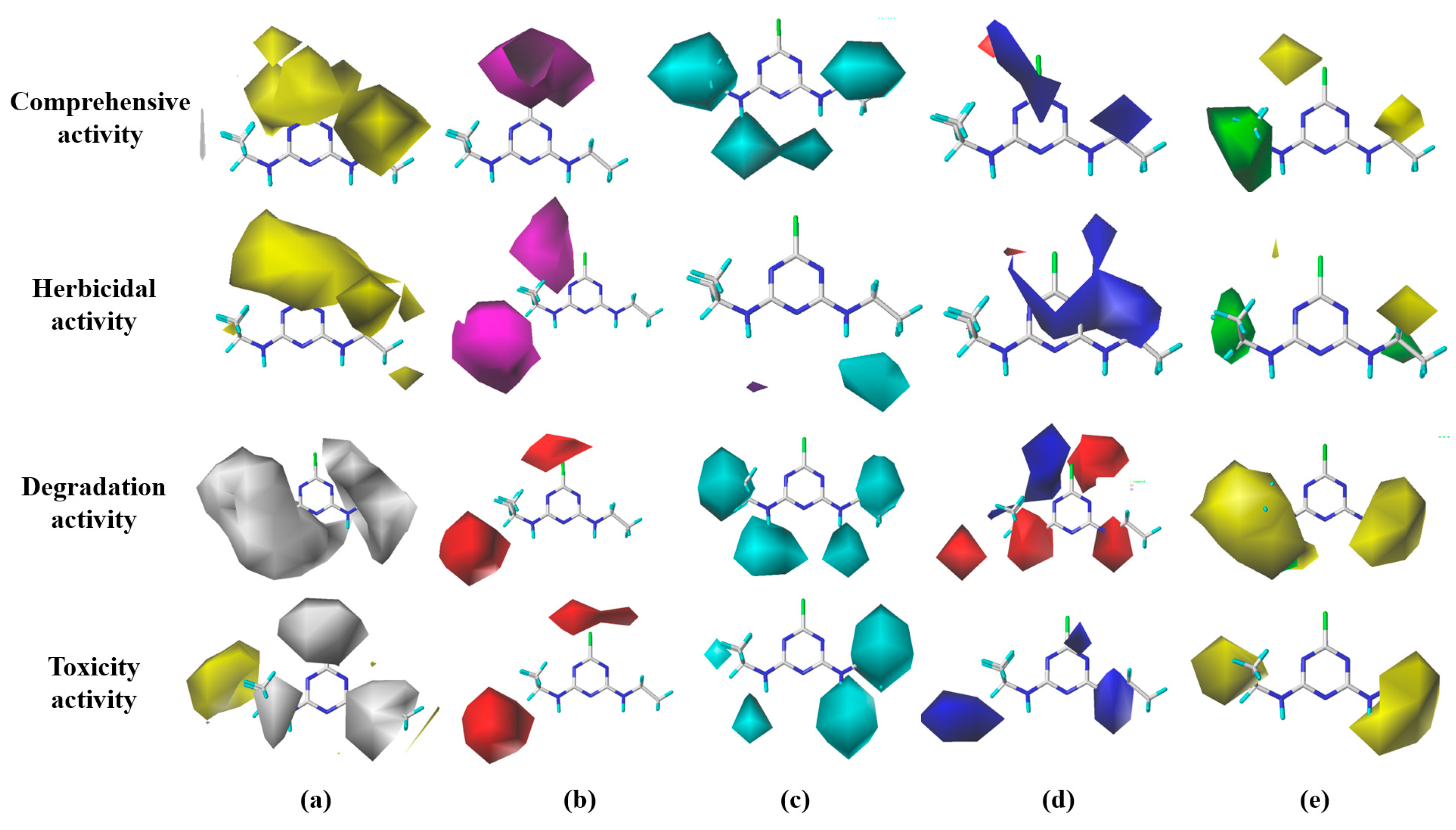
2.2. Design of S-TH Substitutes Based on the 3D Isopotential Diagrams of the CoMSIA Model
2.3. Prediction and Evaluation of the Single-Effect and Comprehensive-Effect 3D-QSAR Models of Herbicidal Properties, Microbial Degradability, and Human Lung Cytotoxicity of S-THs
2.4. Evaluation of the Microbial Degradability Universality and Toxicity of Antioxidant Systems in Fish of S-TH Substitutes
2.4.1. Evaluation of the Microbial Degradability Universality of S-TH Substitutes
2.4.2. Evaluation of the Toxicity of Antioxidant Systems in Fish of S-TH Substitutes
2.5. Screen of Optimal Field Application Schemes to Promote the Microbial Degradation of S-TH Substitutes in Maize Cropping Fields
2.6. Simulation of Microbial Degradation Pathways of S-TH Substitutes
2.7. Evaluation of Aquatic Biotoxicity and Human Health Risks of Microbial Degradation Products of S-TH Substitutes
3. Discussion
4. Materials and Methods
4.1. Characterization of Herbicidal Functionality Properties, Microbial Degradability, and Human Lung Cytotoxicity of S-THs—Molecular Docking Method
4.2. Characterization of the Comprehensive Effects of Herbicidal Functionality Properties, Microbial Degradability, and Human Lung Cytotoxicity of S-THs—AHP-TOPSIS Method
4.3. Construction of a Model for the Comprehensive Effects of Herbicidal Functionality Properties, Microbial Degradability, and Human Lung Cytotoxicity of S-THs—3D-QSAR Model
4.4. Design of S-TH Substitutes—SYBYL-X2.0 Software
4.5. Evaluation of the Microbial Degradability Universality and Toxicity of Antioxidant Systems in Fish of S-THs Substitutes—Molecular Docking Method
4.5.1. Evaluation of the Microbial Degradability Universality of S-THs Substitutes in the Soil Environment
4.5.2. Evaluation of the Toxicity of Antioxidant Systems in Fish of S-TH Substitutes in the Aquatic Environment
4.6. Screen of Optimal Field Application Schemes to Promote the Microbial Degradation of S-TH Substitutes in Maize Cropping Fields—Taguchi Orthogonal Experiments, Full Factorial Design of Experiments, and Molecular Dynamics Methods
4.7. Simulation of Microbial Degradation Pathways of S-TH Substitutes—DFT and Microbial Degradation Pathway Simulation
4.8. Evaluation of Aquatic Biotoxicity and Human Health Risks of Microbial Degradation Products of S-TH Substitutes—EPI Software Method, Pharmacokinetic and Toxicokinetic Methods
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Manousi, N.; Kabir, A.; Zachariadis, G.A. Recent advances in the extraction of triazine herbicides from water samples. J. Sep. Sci. 2021, 45, 113–133. [Google Scholar] [CrossRef] [PubMed]
- Yu, Q.Q.; Lu, F.F.; Ma, L.Y.; Yang, H.; Song, N.H. Residues of Reduced Herbicides Terbuthylazine, Ametryn, and Atrazine and Toxicology to Maize and the Environment through Salicylic Acid. ACS Omega 2021, 6, 27396–27404. [Google Scholar] [CrossRef] [PubMed]
- Funar-Timofei, S.; Borota, A.; Crisan, L. Combined molecular docking and QSAR study of fused heterocyclic herbicide inhibitors of D1 protein in photosystem II of plants. Mol. Divers. 2017, 21, 437–454. [Google Scholar] [CrossRef] [PubMed]
- Li, D.J.; Daler, D. Ocean pollution from land-based sources: East China sea, China. Ambio 2004, 33, 107–113. [Google Scholar] [CrossRef] [PubMed]
- EPA Releases Draft Biological Evaluations for Atrazine, Simazine and Propazine. Available online: https://www.epa.gov/pesticides/epa-releases-draft-biological-evaluations-atrazine-simazine-and-propazine (accessed on 15 May 2022).
- Gu, L.L. New progress of research and development of triazine herbicides. World Pestic. 2021, 43, 12–23. [Google Scholar] [CrossRef]
- Galvín, R.M.; Rodríguez Mellado, J.M.; Higuera, M.J. Reductive deactivation of some s-triazine herbicides: Prometryne, desmetryne and terbutryne. J. Serb. Chem. Soc. 2002, 67, 381–392. [Google Scholar] [CrossRef]
- Zhao, Y.H. Biodegradation and Regulation of the s-Triazine Herbicides. Ph.D. Thesis, Zhejiang University, Hangzhou, China, 2002. [Google Scholar]
- Velisek, J.; Stara, A.; Koutnik, D.; Machova, J. Effects of prometryne on early life stages of common carp (Cyprinus carpio L.). Pestic. Biochem. Physiol. 2015, 118, 58–63. [Google Scholar] [CrossRef]
- Hao, M.J.; Wang, J.H.; Dong, W.; Duan, L.M. Study on the toxicity of herbicide atrazine to zebrafish embryos. Sci. Technol. Vis. 2018, 22, 68–69. [Google Scholar] [CrossRef]
- Khoshnood, Z.; Jamili, S.; Khodabandeh, S. Histopathological effects of atrazine on gills of caspian kutum rutilus frisii kutum fingerlings. Dis. Aquat. Organ. 2015, 113, 227–234. [Google Scholar] [CrossRef]
- Mela, M.; Guiloski, I.C.; Doria, H.B.; Randi, M.A.F.; Ribeiro, C.A.D.; Pereira, L.; Maraschi, A.C.; Prodocimo, V.; Freire, C.A.; de Assis, H.C.S. Effects of the herbicide atrazine in neotropical catfish (Rhamdia quelen). Ecotoxicol. Environ. Saf. 2013, 93, 13–21. [Google Scholar] [CrossRef]
- Tillitt, D.E.; Papoulias, D.M.; Whyte, J.J.; Richter, C.A. Atrazine reduces reproduction in fathead minnow (Pimephales promelas). Aquat. Toxicol. 2010, 99, 149–159. [Google Scholar] [CrossRef] [Green Version]
- Liu, Q.Y. Research on the Cytotoxicity and Related Mechanisms of Herbicide Prometrvn In Vitro. Master’s Thesis, Zhejiang University, Hangzhou, China, 2017. [Google Scholar]
- Joo, H.; Choi, K.; Hodgson, E. Human metabolism of atrazine. Pestic. Biochem. Physiol. 2010, 98, 73–79. [Google Scholar] [CrossRef]
- Zeljezic, D.; Garaj-Vrhovac, V.; Perkovic, P. Evaluation of DNA damage induced by atrazine and atrazine-based herbicide in human lymphocytes in vitro using a comet and DNA diffusion assay. Toxicol. Vitro 2006, 20, 923–935. [Google Scholar] [CrossRef] [PubMed]
- Wang, Q.F.; Xie, S.G. Isolation and characterization of a high-efficiency soil atrazine-degrading Arthrobacter sp. strain. Int. Biodeterior. Biodegrad. 2012, 71, 61–66. [Google Scholar] [CrossRef]
- Li, Q.Y.; Li, Y.; Zhu, X.K.; Cai, B.L. Isolation and characterization of atrazine-degrading Arthrobacter sp. AD26 and use of this strain in bioremediation of contaminated soil. J. Environ. Sci. 2008, 20, 1226–1230. [Google Scholar] [CrossRef]
- Vargha, M.; Takats, Z.; Marialigeti, K. Degradation of atrazine in a laboratory scale model system with Danube River sediment. Water Res. 2005, 39, 1560–1568. [Google Scholar] [CrossRef]
- Singh, P.; Suri, C.R.; Cameotra, S.S. Isolation of a member of Acinetobacter species involved in atrazine degradation. Biochem. Biophys. Res. Commun. 2004, 317, 697–702. [Google Scholar] [CrossRef]
- Strong, L.C.; Rosendahl, C.; Johnson, G.; Sadowsky, M.J.; Wackett, L.P. Arthrobacter aurescens TC1 metabolizes diverse s-triazine ring compounds. Appl. Environ. Microbiol. 2002, 68, 5973–5980. [Google Scholar] [CrossRef] [Green Version]
- Katz, I.; Dosoretz, C.G.; Mandelbaum, R.T.; Green, M. Atrazine degradation under denitrifying conditions in continuous culture of Pseudomonas ADP. Water Res. 2001, 35, 3272–3275. [Google Scholar] [CrossRef]
- Jones, L.R.; Owen, S.A.; Horrell, P. Bacterial inoculation of granular activated carbon filters for the removal of atrazine from surface water. Water Res. 1998, 32, 2542–2549. [Google Scholar] [CrossRef]
- Rousseaux, S.; Hartmann, A.; Lagacherie, B.; Piutti, S.; Andreux, F.; Soulas, G. Inoculation of an atrazine-degrading strain, Chelatobacter heintzii Cit1, in four different soils: Effects of different inoculum densities. Chemosphere 2003, 51, 569–576. [Google Scholar] [CrossRef] [PubMed]
- Mougin, C.; Laugero, C.; Asther, M.; Dubroca, J.; Frasse, P.; Asther, M. Biotransformation of the herbicide atrazine by the white rot rungus Phanerochaete chrysosporium. Appl. Environ. Microbiol. 1994, 60, 705–708. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Masaphy, S.; Levanon, D.; Vaya, J.; Henis, Y. Isolation and characterization of a novel atrazine metabolite produced by the fungus Pleurotus pulmonarius, 2-chloro-4-ethylamino-6-(1-hydroxyisopropyl) amino-1,3,5-triazine. Appl. Environ. Microbiol. 1993, 59, 4342–4346. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Vonberg, D.; Vanderborght, J.; Cremer, N.; Pütz, T.; Herbst, M.; Vereecken, H. 20 years of long-term atrazine monitoring in a shallow aquifer in western Germany. Water Res. 2014, 50, 294–306. [Google Scholar] [CrossRef]
- Qu, R.J.; Liu, H.X.; Feng, M.B.; Yang, X.; Wang, Z.Y. Investigation on intramolecular hydrogen bond and some thermodynamic properties of polyhydroxylated anthraquinones. J. Chem. Eng. Data 2012, 57, 2442–2455. [Google Scholar] [CrossRef]
- Wang, X.L.; Gu, W.E.; Guo, E.; Cui, C.Y.; Li, Y. Assessment of long-range transport potential of polychlorinated Naphthalenes based on three-dimensional QSAR models. Environ. Sci. Pollut. Res. Int. 2017, 24, 14802–14818. [Google Scholar] [CrossRef]
- Veerasamy, R.; Rajak, H.; Jain, A.; Sivadasan, S.; Varghese, C.P.; Agrawal, R.K. Validation of QSAR models—Strategies and importance. Int. J. Drug Des. Discov. 2011, 2, 511–519. [Google Scholar]
- Karns, J.S. Gene sequence and properties of an s-triazine ring-cleavage enzyme from Pseudomonas sp. strain NRRLB-12227. Appl. Environ. Microbiol. 1999, 65, 3512–3517. [Google Scholar] [CrossRef] [Green Version]
- Asgher, M.; Bhatti, H.N.; Ashraf, M.; Legge, R.L. Recent developments in biodegradation of industrial pollutants by white rot fungi and their enzyme system. Biodegradation 2008, 19, 771–783. [Google Scholar] [CrossRef]
- Protein Data Bank. Available online: https://www.rcsb.org (accessed on 15 July 2022).
- UniProt. Available online: https://www.uniprot.org (accessed on 6 August 2022).
- Sun, Y.H. Researches on the Oxidation Mechanisms of Typical Organic Pollutants in the Atmosphere. Ph.D. Thesis, Shandong University, Jinan, China, 2015. [Google Scholar]
- Sun, P.X.; Zhao, W.J. Strategies to control human health risks arising from antibiotics in the environment: Molecular modification of QNs for enhanced plant-microbial synergistic degradation. Int. J. Environ. Res. Public Health 2021, 18, 10610. [Google Scholar] [CrossRef]
- Shen, Y.Y. Study on the Behavior Characteristics of Petroleum Containment in the Soil and Rhizoremediation. Ph.D. Thesis, Chang’an University, Xi’an, China, 2013. [Google Scholar]
- Lipson, D.A.; Raab, T.K.; Schmidt, S.K.; Monson, R.K. An empirical model of amino acid transformations in an alpine soil. Soil. Biol. Biochem. 2001, 33, 189–198. [Google Scholar] [CrossRef]
- Chapin, F.S.; Matson, P.A.; Mooney, H.A. Terrestrial nutrient cycling. In Principles of Terrestrial Ecosystem Ecology, 1st ed.; Springer: New York, NY, USA, 2002; pp. 97–122. [Google Scholar] [CrossRef]
- Li, J.B.; Luo, C.L.; Zhang, D.Y.; Cai, X.X.; Jiang, L.F.; Zhao, X.; Zhang, G. Diversity of the active phenanthrene degraders in PAH-polluted soil is shaped by ryegrass rhizosphere and root exudates. Soil. Biol. Biochem. 2019, 128, 100–110. [Google Scholar] [CrossRef]
- Fu, R.; Li, X.; Zhao, Y.; Pu, Q.; Li, Y.; Gu, W. Efficient and synergistic degradation of fluoroquinolones by bacteria and microalgae: Design of environmentally friendly substitutes, risk regulation and mechanism analysis. J. Hazard. Mater. 2022, 437, 129384. [Google Scholar] [CrossRef]
- Li, X.; Hou, Y.; Li, Q.; Gu, W.; Li, Y. Molecular design of high-efficacy and high drug safety Fluoroquinolones suitable for a variety of aerobic biodegradation bacteria. J. Environ. Manag. 2021, 299, 113628. [Google Scholar] [CrossRef] [PubMed]
- Xue, J.Q.; Chen, X.Y.; Zhao, Y.Y.; Li, Y. Exposure to high-performance benzotriazole ultraviolet stabilizers: Advance in toxicological effects, environmental behaviors and remediation mechanism using in-silica methods. Chemosphere 2023, 315, 137699. [Google Scholar] [CrossRef] [PubMed]
- Gu, W.W.; Zhao, Y.Y.; Li, Q.; Li, Y. Environmentally friendly polychlorinated naphthalenes (PCNs) derivatives designed using 3D-QSAR and screened using molecular docking, density functional theory and health-based risk assessment. J. Hazard. Mater. 2019, 363, 316–327. [Google Scholar] [CrossRef] [PubMed]
- Liao, D.; Qian, J.; Chisholm, D.A.; Jordan, D.B.; Diner, B.A. Crystal structures of the photosystem II D1 C-terminal processing protease. Nat. Struct. Mol. Biol. 2000, 7, 749–753. [Google Scholar] [CrossRef]
- Jackson, C.J.; Coppin, C.W.; Carr, P.D.; Aleksandrov, A.; Wilding, M.; Sugrue, E.; Ubels, J.; Paks, M.; Newman, J.; Peat, T.S.; et al. 300-fold increase in production of the Zn2+-dependent dechlorinase TrzN in soluble form via apoenzyme stabilization. Appl. Environ. Microbiol. 2014, 80, 4003–4011. [Google Scholar] [CrossRef] [Green Version]
- Sharma, D.; De Falco, L.; Padavattan, S.; Rao, C.; Geifman-Shochat, S.; Liu, C.F.; Davey, C.A. PARP1 exhibits enhanced association and catalytic efficiency with γH2A.X-nucleosome. Nat. Commun. 2019, 10, 5751. [Google Scholar] [CrossRef] [Green Version]
- Pan, J.L.; Xu, Q.J.; Liu, H.X. Typhoon disaster risk assessment based on AHP-TOPSIS optimal combination empowerment. J. Nanning Norm. Univ. (Nat. Sci. Ed.) 2021, 38, 60–67. [Google Scholar] [CrossRef]
- Zeng, X.L.; Qu, R.J.; Feng, M.B.; Chen, J.; Wang, L.S.; Wang, Z.Y. Photodegradation of Polyfluorinated Dibenzo-p-Dioxins in Organic Solvents: Experimental and Theoretical Studies. Environ. Sci. Technol. 2016, 50, 8128–8134. [Google Scholar] [CrossRef] [PubMed]
- Hou, Y.L.; Zhao, Y.Y.; Li, Y. Environmentally friendly fluoroquinolone derivatives with lower plasma protein binding rate designed using 3D-QSAR, molecular docking and molecular dynamics simulation. Int. J. Environ. Res. Public Health 2020, 17, 6626. [Google Scholar] [CrossRef]
- Yang, L.Z.; Liu, M. A double-activity (green algae toxicity and bacterial genotoxicity) 3D-QSAR model based on the comprehensive index method and its application in fluoroquinolones’ modification. Int. J. Environ. Res. Public Health 2020, 17, 942. [Google Scholar] [CrossRef] [Green Version]
- Morris, T.F.; Murrell, T.S.; Beegle, D.B.; Camberato, J.J.; Ferguson, R.B.; Grove, J.; Ketterings, Q.; Kyveryga, P.M.; Laboski, C.A.M.; McGrath, J.M.; et al. Strengths and limitations of nitrogen rate recommendations for corn and opportunities for improvement. Agron. J. 2018, 110, 1–37. [Google Scholar] [CrossRef] [Green Version]
- Liu, X.W.; Wang, K.Q.; Zhao, Y.Y.; Duan, X.; Zhang, Y. Characteristics of maize root exudates at seedling stage and their response to rhizosphere soil enzyme activities under contour reverse-slope terrace. J. Sichuan Agric. Univ. 2021, 39, 477–485. [Google Scholar] [CrossRef]
- Zhu, H.F.; Zhang, L.; Li, S.S.; Wang, Y.J.; Sun, S.R.; Chen, J.; Kou, S.M.; Jin, Q.; Xiao, M. The rhizosphere and root exudates of maize seedlings drive plasmid mobilization in soil. Appl. Soil Ecol. 2018, 124, 194–202. [Google Scholar] [CrossRef]
- Majumdar, A.; Ghosh, D. Genetic algorithm parameter optimization using Taguchi robust design for multi-response optimization of experimental and historical data. Int. J. Comput. Appl. 2015, 127, 26–32. [Google Scholar] [CrossRef]
- Childers, M.C.; Daggett, V. Insights from molecular dynamics simulations for computational protein design. Mol. Syst. Des. Eng. 2017, 2, 9–33. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kendrick, M.R.; Benstead, J.P. Temperature and nutrient availability interact to mediate growth and body stoichiometry in a detritivorous stream insect. Freshwater Biol. 2013, 58, 1820–1830. [Google Scholar] [CrossRef]
- Shapir, N.; Cheng, G.; Sadowsky, M.J.; Wackett, L.P. Purification and characterization of TrzF: Biuret hydrolysis by allophanate hydrolase supports growth. Appl. Environ. Microbiol. 2006, 72, 2491–2495. [Google Scholar] [CrossRef] [Green Version]
- Salahinejad, M.; Ghasemi, J.B. 3D-QSAR studies on the toxicity of substituted benzenes to Tetrahymena pyriformis: CoMFA, CoMSIA and VolSurf approaches. Ecotoxicol. Environ. Saf. 2014, 105, 128–134. [Google Scholar] [CrossRef] [PubMed]
- US EPA. Estimation Programs Interface Suite™ for Microsoft® Windows, v 4.11 or Insert Version Used; United States Environmental Protection Agency: Washington, DC, USA, 2012.







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| CoMSIA Models | Enzymes | q2 | n | R2 | SEE | F | r2pred | (R2 − q2)/R2 (%) |
|---|---|---|---|---|---|---|---|---|
| Comprehensive | ALL | 0.789 | 8 | 0.993 | 0.007 | 132.292 | 0.615 | 20.54 |
| Herbicide | 1FC9 | 0.751 | 9 | 0.997 | 0.393 | 207.333 | 0.785 | 24.67 |
| Degradation | 4L9X | 0.757 | 5 | 0.986 | 1.529 | 157.196 | 0.713 | 23.23 |
| Toxicity | 6K1J | 0.706 | 10 | 1.000 | 0.285 | 1660.695 | 0.792 | 29.40 |
| Fields | Proportion of Fields (%) | |||
|---|---|---|---|---|
| CoMSIA Models | ||||
| For Comprehensive Activity | For Herbicidal Activity | For Degradation Activity | For Toxicity Activity | |
| Hydrophobic (H) | 37.4 | 50 | 29.3 | 32.5 |
| Hydrogen-bond acceptor (A) | 4.7 | 6.9 | 7.1 | 6.9 |
| Hydrogen-bond donor (D) | 26.2 | 6.8 | 27.6 | 22.8 |
| Electrostatic (E) | 16.8 | 18.8 | 17.8 | 19.4 |
| Steric (S) | 14.9 | 16.5 | 18.1 | 18.4 |
| Compounds | Steps | Reactants | Products | ΔE | ΔE (Total) | Change Rate (%) |
|---|---|---|---|---|---|---|
| (kJ/mol) | (kJ/mol) | |||||
| ATZ | 1 | ATZ | ATZ-1 | 160.893 | 200.806 | - |
| 2 | ATZ-1 | Com-1 | 39.913 | |||
| D-5 | 1 | D-5 | D-5-1 | 60.147 | 94.533 | −62.62 |
| 2 | D-5-1 | Com-1 | 34.386 | −13.85 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Yao, T.; Sun, P.; Zhao, W. Triazine Herbicides Risk Management Strategies on Environmental and Human Health Aspects Using In-Silico Methods. Int. J. Mol. Sci. 2023, 24, 5691. https://doi.org/10.3390/ijms24065691
Yao T, Sun P, Zhao W. Triazine Herbicides Risk Management Strategies on Environmental and Human Health Aspects Using In-Silico Methods. International Journal of Molecular Sciences. 2023; 24(6):5691. https://doi.org/10.3390/ijms24065691
Chicago/Turabian StyleYao, Tianfu, Peixuan Sun, and Wenjin Zhao. 2023. "Triazine Herbicides Risk Management Strategies on Environmental and Human Health Aspects Using In-Silico Methods" International Journal of Molecular Sciences 24, no. 6: 5691. https://doi.org/10.3390/ijms24065691