
Molecular Aspects Implicated in Dantrolene Selectivity with Respect to Ryanodine Receptor Isoforms

Abstract
:1. Introduction
2. RyR-Linked Channelopathies
3. The Dantrolene-Mediated Inhibitory Effect on the RyR1 Channel
3.1. Sensitivity of the RyR1 Channel to the Activation Effect of Volatile Anesthetics
3.2. Structural Determinants of Halothane Binding to the RyR1 Channel
3.3. Essential Conditions for Observing the Dantrolene-Mediated Inhibitory Effect on the RyR1 Channel
4. The RyR2 Channel as an Endogenous Target of Dantrolene
4.1. Activation of the RyR2 Channel by Volatile Anesthetics
4.2. RyR2 Responsiveness to Dantrolene in the Normal Heart
4.3. RyR2 Responsiveness to Dantrolene under Pathological Conditions
5. The Inhibitory Effect of Dantrolene on the RyR3 Channel
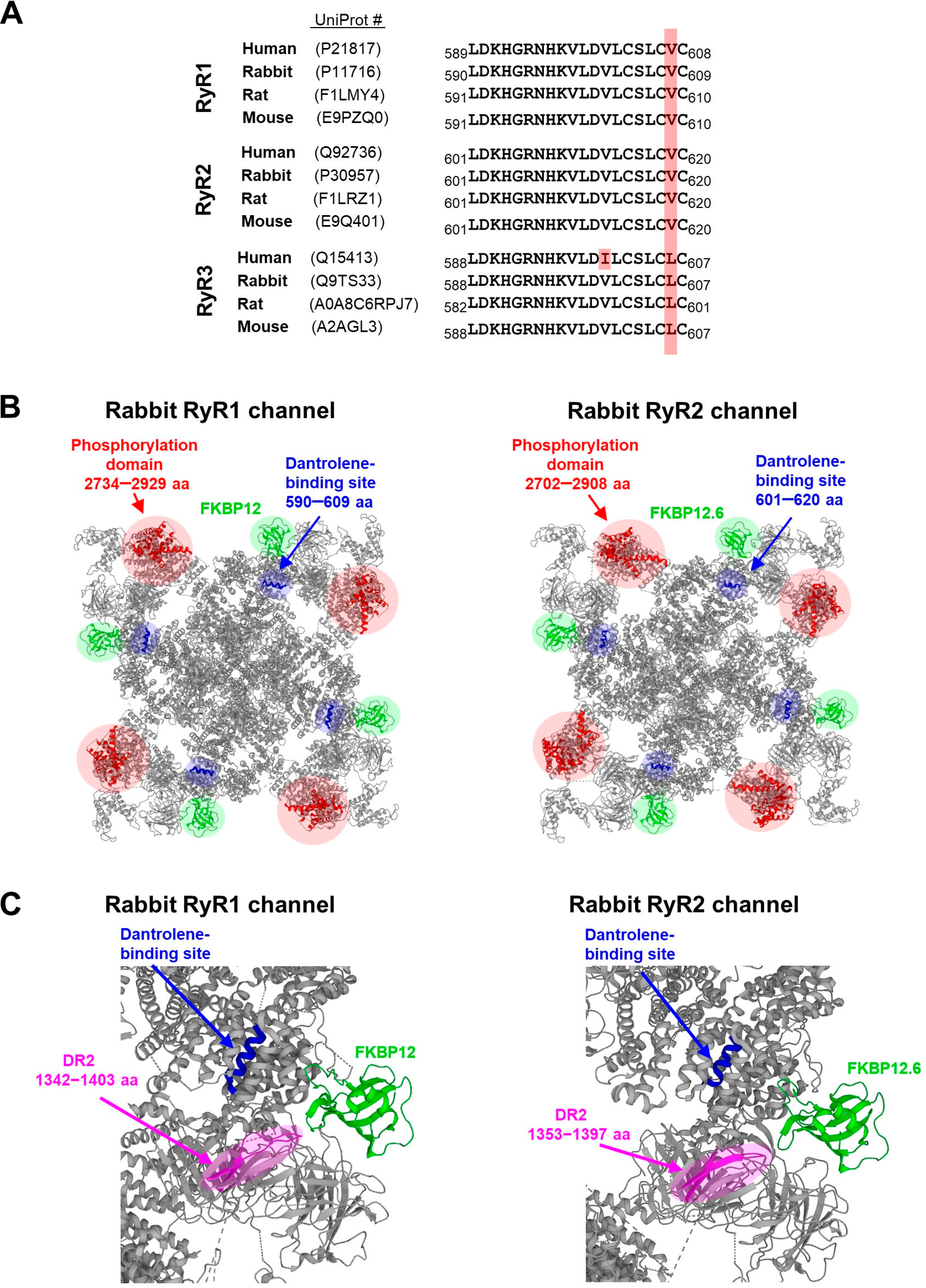
6. Localization of the Binding Site for Dantrolene in the Three-Dimensional (3D) Structure of RyR Isoforms
7. Dantrolene Suitability for Novel Clinical Applications
8. Conclusions and Future Outlook
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Carafoli, E. Calcium signaling: A tale for all seasons. Proc. Natl. Acad. Sci. USA 2002, 99, 1115–1122. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Berridge, M.J.; Bootman, M.D.; Roderick, H.L. Calcium signalling: Dynamics, homeostasis and remodelling. Nat. Rev. Mol. Cell Biol. 2003, 4, 517–529. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bers, D.M. Cardiac sarcoplasmic reticulum calcium leak: Basis and roles in cardiac dysfunction. Annu. Rev. Physiol. 2014, 76, 107–127. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Landstrom, A.P.; Dobrev, D.; Wehrens, X.H.T. Calcium signaling and cardiac arrhythmias. Circ. Res. 2017, 120, 1969–1993. [Google Scholar] [CrossRef]
- Supnet, C.; Bezprozvanny, I. Neuronal calcium signaling, mitochondrial dysfunction and Alzheimer’s disease. J. Alzheimer’s Dis. 2010, 20, S487–S498. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zaichick, S.V.; McGrath, K.M.; Caraveo, G. The role of Ca2+ signaling in Parkinson’s disease. Dis. Model. Mech. 2017, 10, 519–535. [Google Scholar] [CrossRef] [Green Version]
- Emptage, N.J.; Reid, C.A.; Fine, A. Calcium stores in hippocampal synaptic boutons mediate short-term plasticity, store-operated Ca2+ entry, and spontaneous transmitter release. Neuron 2001, 29, 197–208. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Holbro, N.; Grunditz, A.; Oertner, T.G. Differential distribution of endoplasmic reticulum controls metabotropic signaling and plasticity at hippocampal synapses. Proc. Natl. Acad. Sci. USA 2009, 106, 15055–15060. [Google Scholar] [CrossRef] [Green Version]
- Lanner, J.T. Ryanodine receptor physiology and its role in disease. Adv. Exp. Med. Biol. 2012, 740, 217–234. [Google Scholar] [CrossRef]
- Bers, D.M. Stabilizing ryanodine receptor gating quiets arrhythmogenic events in human heart failure and atrial fibrillation. Heart Rhythm. 2017, 14, 420–421. [Google Scholar] [CrossRef]
- Kushnir, A.; Wajsberg, B.; Marksa, A.R. Ryanodine receptor dysfunction in human disorders. Biochim. Biophys. Acta 2018, 1865, 1687–1697. [Google Scholar] [CrossRef] [PubMed]
- Connell, P.; Word, T.A.; Wehrens, X.H.T. Targeting pathological leak of ryanodine receptors: Preclinical progress and the potential impact on treatments for cardiac arrhythmias and heart failure. Expert Opin. Ther. Targets 2020, 24, 25–36. [Google Scholar] [CrossRef] [PubMed]
- Avila, G.; Dirksen, R.T. Functional effects of central core disease mutations in the cytoplasmic region of the skeletal muscle ryanodine receptor. J. Gen. Physiol. 2001, 118, 277–290. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Szentandrássy, N.; Magyar, Z.É.; Hevesi, J.; Bányász, T.; Nánási, P.P.; Almássy, J. Therapeutic approaches of ryanodine receptor-associated heart diseases. Int. J. Mol. Sci. 2022, 23, 4435. [Google Scholar] [CrossRef] [PubMed]
- Saljic, A.; Heijman, J.; Dobrev, D. Emerging antiarrhythmic drugs for atrial fibrillation. Int. J. Mol. Sci. 2022, 23, 4096. [Google Scholar] [CrossRef]
- Krause, T.; Gerbershagen, M.U.; Fiege, M.; Weisshorn, R.; Wappler, F. Dantrolene--a review of its pharmacology, therapeutic use and new developments. Anaesthesia 2004, 59, 364–373. [Google Scholar] [CrossRef]
- Denborough, M.A.; Forster, J.F.; Lovell, R.R.; Maplestone, P.A.; Villiers, J.D. Anaesthetic deaths in a family. Br. J. Anaesth. 1962, 34, 395–396. [Google Scholar] [CrossRef]
- Paul-Pletzer, K.; Palnitkar, S.S.; Jimenez, L.S.; Morimoto, H.; Parness, J. The skeletal muscle ryanodine receptor identified as a molecular target of [3H]azidodantrolene by photoaffinity labeling. Biochemistry 2001, 40, 531–542. [Google Scholar] [CrossRef]
- Paul-Pletzer, K.; Yamamoto, T.; Ikemoto, N.; Jimenez, L.S.; Morimoto, H.; Williams, P.G.; Ma, J.; Parness, J. Probing a putative dantrolene-binding site on the cardiac ryanodine receptor. Biochem. J. 2005, 387, 905–909. [Google Scholar] [CrossRef]
- Ellis, R.H.; Simpson, P.; Tatham, P.; Leighton, M.; Williams, J. The cardiovascular effects of dantrolene sodium in dogs. Anaesthesia 1975, 30, 318–322. [Google Scholar] [CrossRef]
- Ellis, K.O.; Butterfield, J.L.; Wessels, F.L.; Carpenter, J.F. A comparison of skeletal, cardiac, and smooth muscle actions of dantrolene sodium--a skeletal muscle relaxant. Arch. Int. Pharmacodyn. Ther. 1976, 224, 118–132. [Google Scholar]
- Van Winkle, W.B. Calcium release from skeletal muscle sarcoplasmic reticulum: Site of action of dantrolene sodium? Science 1976, 193, 1130–1131. [Google Scholar] [CrossRef] [PubMed]
- Fratea, S.; Langeron, O.; Lecarpentier, Y.; Coria, P.; Riou, B. In vitro effects of dantrolene on rat myocardium. Anesthesiology 1997, 86, 205–215. [Google Scholar] [CrossRef]
- Bowman, W.C.; Khan, H.H. The effects of dantrolene sodium on isolated skeletal, smooth and cardiac muscle of the guinea-pig. J. Pharm. Pharmacol. 1977, 29, 628–630. [Google Scholar] [CrossRef]
- Putney, J.W.; Biancri, C.P. Site of action of dantrolene in frog sartorius muscle. J. Pharmacol. Exp. Ther. 1974, 189, 202–212. [Google Scholar]
- Hainaut, K.; Desmedt, J.E. Effect of dantrolene sodium on calcium movements in single muscle fibres. Nature 1974, 252, 728–730. [Google Scholar] [CrossRef] [PubMed]
- Flewellen, E.H.; Nelson, T.E.; Jones, W.P.; Arens, J.F.; Wagner, D.L. Dantrolene dose response in awake man: Implications for management of malignant hyperthermia. Anesthesiology 1983, 59, 275–280. [Google Scholar] [CrossRef] [PubMed]
- Szentesi, P.; Collet, C.; Sárközi, S.; Szegedi, C.; Jona, I.; Jacquemond, V.; Kovács, L.; Csernoch, L. Effects of dantrolene on steps of excitation-contraction coupling in mammalian skeletal muscle fibers. J. Gen. Physiol. 2001, 118, 355–375. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fruen, B.R.; Mickelson, J.R.; Louis, C.F. Dantrolene inhibition of sarcoplasmic reticulum Ca2+ release by direct and specific action at skeletal muscle ryanodine receptors. J. Biol. Chem. 1997, 272, 26965–26971. [Google Scholar] [CrossRef] [Green Version]
- Zhao, F.; Li, P.; Chen, S.R.; Louis, C.F.; Fruen, B.R. Dantrolene inhibition of ryanodine receptor Ca2+ release channels. Molecular mechanism and isoform selectivity. J. Biol. Chem. 2001, 276, 13810–13816. [Google Scholar] [CrossRef] [Green Version]
- Kobayashi, S.; Yano, M.; Suetomi, T.; Ono, M.; Tateishi, H.; Mochizuki, M.; Xu, X.; Uchinoumi, H.; Okuda, S.; Yamamoto, T.; et al. Dantrolene, a therapeutic agent for malignant hyperthermia, markedly improves the function of failing cardiomyocytes by stabilizing interdomain interactions within the ryanodine receptor. J. Am. Coll. Cardiol. 2009, 53, 1993–2005. [Google Scholar] [CrossRef] [Green Version]
- Kobayashi, S.; Yano, M.; Uchinoumi, H.; Suetomi, T.; Susa, T.; Ono, M.; Xu, X.; Tateishi, H.; Oda, T.; Okuda, S.; et al. Dantrolene, a therapeutic agent for malignant hyperthermia, inhibits catecholaminergic polymorphic ventricular tachycardia in a RyR2R2474S/+ knock-in mouse model. Circ. J. 2010, 74, 2579–2584. [Google Scholar] [CrossRef] [Green Version]
- Jung, C.B.; Moretti, A.; Schnitzler, M.M.; Iop, L.; Storch, U.; Bellin, M.; Dorn, T.; Ruppenthal, S.; Pfeiffer, S.; Goedel, A.; et al. Dantrolene rescues arrhythmogenic RYR2 defect in a patient-specific stem cell model of catecholaminergic polymorphic ventricular tachycardia. EMBO Mol. Med. 2012, 4, 180–190. [Google Scholar] [CrossRef]
- Maxwell, J.T.; Domeier, T.L.; Blatter, L.A. Dantrolene prevents arrhythmogenic Ca2+ release in heart failure. Am. J. Physiol. 2012, 302, H953–H963. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Azam, M.A.; Chakraborty, P.; Bokhari, M.M.; Dadson, K.; Du, B.; Massé, S.; Si, D.; Niri, A.; Aggarwal, A.K.; Lai, P.F.H.; et al. Cardioprotective effects of dantrolene in doxorubicin-induced cardiomyopathy in mice. Heart Rhythm. O2 2021, 2, 733–741. [Google Scholar] [CrossRef]
- Litman, R.S.; Rosenberg, H. Malignant hyperthermia: Update on susceptibility testing. JAMA J. Am. Med. Assoc. 2005, 293, 2918–2924. [Google Scholar] [CrossRef]
- Rosero, E.B.; Adesanya, A.O.; Timaran, C.H.; Joshi, G.P. Trends and outcomes of malignant hyperthermia in the United States, 2000 to 2005. Anesthesiology 2009, 110, 89–94. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shi, Y.; Wang, Y.; Wei, H. Dantrolene: From malignant hyperthermia to Alzheimer’s disease. CNS Neurol. Disord. Drug Targets 2019, 18, 668–676. [Google Scholar] [CrossRef] [PubMed]
- Stenson, P.D.; Mort, M.; Ball, E.V.; Chapman, M.; Evans, K.; Azevedo, L.; Hayden, M.; Heywood, S.; Millar, D.S.; Phillips, A.D.; et al. The Human Gene Mutation Database (HGMD®): Optimizing its use in a clinical diagnostic or research setting. Hum. Genet. 2020, 139, 1197–1207. [Google Scholar] [CrossRef]
- Tunwell, R.E.; Wickenden, C.; Bertrand, B.M.; Shevchenko, V.I.; Walsh, M.B.; Allen, P.D.; Lai, F.A. The human cardiac muscle ryanodine receptor-calcium release channel: Identification, primary structure and topological analysis. Biochem. J. 1996, 318, 477–487. [Google Scholar] [CrossRef] [Green Version]
- Marks, A.R.; Tempst, P.; Hwang, K.S.; Taubman, M.B.; Inui, M.; Chadwick, C.; Fleischer, S.; Nadal-Ginard, B. Molecular cloning and characterization of the ryanodine receptor/junctional channel complex cDNA from skeletal muscle sarcoplasmic reticulum. Proc. Natl. Acad. Sci. USA 1989, 86, 8683–8687. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Froemming, G.R.; Murray, B.E.; Harmon, S.; Pette, D.; Ohlendieck, K. Comparative analysis of the isoform expression pattern of Ca2+-regulatory membrane proteins in fast-twitch, slow-twitch, cardiac, neonatal and chronic low-frequency stimulated muscle fibers. Biochim. Biophys. Acta 2000, 1466, 151–168. [Google Scholar] [CrossRef] [Green Version]
- Nakai, J.; Imagawa, T.; Hakamata, Y.; Shigekawa, M.; Takeshima, H.; Numa, S. Primary structure and functional expression from cDNA of the cardiac ryanodine receptor/calcium release channel. FEBS Lett. 1990, 271, 169–177. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Otsu, K.; Willard, H.F.; Khanna, V.K.; Zorzato, F.; Green, N.M.; MacLennan, D.H. Molecular cloning of cDNA encoding the Ca2+ release channel (ryanodine receptor) of rabbit cardiac muscle sarcoplasmic reticulum. J. Biol. Chem. 1990, 265, 13472–13483. [Google Scholar] [CrossRef]
- Hakamata, Y.; Nakai, J.; Takeshima, H.; Imoto, K. Primary structure and distribution of a novel ryanodine receptor/calcium release channel from rabbit brain. FEBS Lett. 1992, 312, 229–235. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Giannini, G.; Clementi, E.; Ceci, R.; Marziali, G.; Sorrentino, V. Expression of a ryanodine receptor-Ca2+ channel that is regulated by TGF-β. Science 1992, 257, 91–94. [Google Scholar] [CrossRef]
- Giannini, G.; Conti, A.; Mammarella, S.; Scrobogna, M.; Sorrentino, V. The ryanodine receptor/calcium channel genes are widely and differentially expressed in murine brain and peripheral tissues. J. Cell Biol. 1995, 128, 893–904. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Furuichi, T.; Furutama, D.; Hakamata, Y.; Nakai, J.; Takeshima, H.; Mikoshiba, K. Multiple types of ryanodine receptor/Ca2+ release channels are differentially expressed in rabbit brain. J. Neurosci. 1994, 14, 4794–4805. [Google Scholar] [CrossRef] [Green Version]
- Robinson, R.; Carpenter, D.; Shaw, M.A.; Halsall, J.; Hopkins, P. Mutations in RYR1 in malignant hyperthermia and central core disease. Hum. Mutat. 2006, 27, 977–989. [Google Scholar] [CrossRef] [PubMed]
- Tammaro, A.; Di Martino, A.; Bracco, A.; Cozzolino, S.; Savoia, G.; Andria, B.; Cannavo, A.; Spagnuolo, M.; Piluso, G.; Aurino, S.; et al. Novel missense mutations and unexpected multiple changes of RYR1 gene in 75 malignant hyperthermia families. Clin. Genet. 2011, 79, 438–447. [Google Scholar] [CrossRef]
- Zhang, Y.; Chen, S.; Khanna, V.K.; De Leon, S.; Phillips, M.S.; Schappert, K.; Britt, B.A.; Brownell, K.W.; MacLennan, D.H. A mutation in the human ryanodine receptor gene associated with central core disease. Nat. Genet. 1993, 5, 46–50. [Google Scholar] [CrossRef] [PubMed]
- Romero, N.B.; Monnier, N.; Viollet, L.; Cortey, A.; Chevallay, M.; Leroy, J.P.; Lunardi, J.; Fardeau, M. Dominant and recessive central core disease associated with RYR1 mutations and fetal akinesia. Brain 2003, 126, 2341–2349. [Google Scholar] [CrossRef] [PubMed]
- Shepherd, S.; Ellis, F.; Halsall, J.; Hopkins, P.; Robinson, R. RYR1 mutations in UK central core disease patients: More than just the C-terminal transmembrane region of the RYR1 gene. J. Med. Genet. 2004, 41, e33. [Google Scholar] [CrossRef] [Green Version]
- Jungbluth, H.; Zhou, H.; Hartley, L.; Halliger-Keller, B.; Messina, S.; Longman, C.; Brockington, M.; Robb, S.A.; Straub, V.; Voit, T.; et al. Minicore myopathy with ophthalmoplegia caused by mutations in the ryanodine receptor type 1 gene. Neurology 2005, 65, 1930–1935. [Google Scholar] [CrossRef] [PubMed]
- Zhou, H.; Yamaguchi, N.; Xu, L.; Wang, Y.; Sewry, C.; Jungbluth, H.; Zorzato, F.; Bertini, E.; Muntoni, F.; Meissner, G.; et al. Characterization of recessive RYR1 mutations in core myopathies. Hum. Mol. Genet. 2006, 15, 2791–2803. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Snoeck, M.; van Engelen, B.G.M.; Küsters, B.; Lammens, M.; Meijer, R.; Molenaar, J.P.F.; Raaphorst, J.; Verschuuren-Bemelmans, C.C.; Straathof, C.S.M.; Sie, L.T.L.; et al. RYR1-related myopathies: A wide spectrum of phenotypes throughout life. Eur. J. Neurol. 2015, 22, 1094–1112. [Google Scholar] [CrossRef]
- Lanner, J.T.; Georgiou, D.K.; Joshi, A.D.; Hamilton, S.L. Ryanodine receptors: Structure, expression, molecular details, and function in calcium release. Cold Spring Harb. Perspect. Biol. 2010, 2, a003996. [Google Scholar] [CrossRef] [Green Version]
- Bauerová-Hlinková, V.; Hajdúchová, D.; Bauer, J.A. Structure and function of the human ryanodine receptors and their association with myopathies—Present state, challenges, and perspectives. Molecules 2020, 23, 4040. [Google Scholar] [CrossRef]
- Avila, G. Intracellular Ca2+ dynamics in malignant hyperthermia and central core disease: Established concepts, new cellular mechanisms involved. Cell Calcium 2005, 37, 121–127. [Google Scholar] [CrossRef] [PubMed]
- Zhou, H.; Jungbluth, H.; Sewry, C.A.; Feng, L.; Bertini, E.; Bushby, K.; Straub, V.; Roper, H.; Rose, M.R.; Brockington, M.; et al. Molecular mechanisms and phenotypic variation in RYR1-related congenital myopathies. Brain Res. 2007, 130, 2024–2036. [Google Scholar] [CrossRef] [Green Version]
- Jiang, D.; Chen, W.; Xiao, J.; Wang, R.; Kong, H.; Jones, P.P.; Zhang, L.; Fruen, B.; Wayne Chen, S.R. Reduced threshold for luminal Ca2+ activation of RyR1 underlies a causal mechanism of porcine malignant hyperthermia. J. Biol. Chem. 2008, 283, 20813–20820. [Google Scholar] [CrossRef] [Green Version]
- Feng, W.; Barrientos, G.C.; Cherednichenko, G.C.; Yang, T.; Padilla, I.T.; Truong, K.; Allen, P.D.; Lopez, J.R.; Pessah, I.N. Functional and biochemical properties of ryanodine receptor type 1 channels from heterozygous R163C malignant hyperthermia-susceptible mice. Mol. Pharmacol. 2011, 79, 420–431. [Google Scholar] [CrossRef] [Green Version]
- Lawal, T.A.; Todd, J.J.; Meilleur, K.G. Ryanodine receptor 1-related myopathies: Diagnostic and therapeutic approaches. Neurotherapeutics 2018, 15, 885–899. [Google Scholar] [CrossRef] [Green Version]
- Riazi, S.; Kraeva, N.; Muldoon, S.M.; Dowling, J.; Ho, C.; Petre, M.-A.; Parness, J.; Dirksen, R.T.; Rosenberg, H. Clinical significance of ryanodine receptor 1 gene (RYR1) variants: Proceedings of the 2013 MHAUS scientific conference. Can. J. Anaesth. 2014, 61, 1040–1049. [Google Scholar] [CrossRef] [Green Version]
- Olubando, D.; Hopton, C.; Eden, J.; Caswell, R.; Thomas, N.L.; Roberts, S.A.; Morris-Rosendahl, D.; Venetucci, L.; Newman, W.G. Classification and correlation of RYR2 missense variants in individuals with catecholaminergic polymorphic ventricular tachycardia reveals phenotypic relationships. J. Hum. Genet. 2020, 65, 531–539. [Google Scholar] [CrossRef] [PubMed]
- Hadiatullah, H.; He, Z.; Yuchi, Z. Structural insight into ryanodine receptor channelopathies. Front. Pharmacol. 2022, 13, 897494. [Google Scholar] [CrossRef] [PubMed]
- Priori, S.G.; Napolitano, C.; Tiso, N. Mutations in the cardiac ryanodine receptor gene (hRyR2) underlie catecholaminergic polymorphic ventricular tachycardia. Circulation 2001, 103, 196–200. [Google Scholar] [CrossRef] [Green Version]
- Laitinen, P.J.; Swan, H.; Piippo, K.; Viitasalo, M.; Toivonen, L.; Kontula, K. Genes, exercise and sudden death: Molecular basis of familial catecholaminergic polymorphic ventricular tachycardia. Ann. Med. 2004, 36, 81–86. [Google Scholar] [CrossRef] [PubMed]
- Tiso, N.; Stephan, D.A.; Nava, A.; Bagattin, A.; Devaney, J.M.; Stanchi, F.; Larderet, G.; Brahmbhatt, B.; Brown, K.; Bauce, B.; et al. Identification of mutations in the cardiac ryanodine receptor gene in families affected with arrhythmogenic right ventricular cardiomyopathy type 2 (ARVD2). Hum. Mol. Genet. 2001, 10, 189–194. [Google Scholar] [CrossRef] [Green Version]
- Milting, H.; Lukas, N.; Klauke, B.; Körfer, R.; Perrot, A.; Osterziel, K.J.; Vogt, J.; Peters, S.; Thieleczek, R.; Varsányi, M. Composite polymorphisms in the ryanodine receptor 2 gene associated with arrhythmogenic right ventricular cardiomyopathy. Cardiovasc. Res. 2006, 71, 496–505. [Google Scholar] [CrossRef]
- Choi, G.; Kopplin, L.J.; Tester, D.J.; Will, M.L.; Haglund, C.M.; Ackerman, M.J. Spectrum and frequency of cardiac channel defects in swimming-triggered arrhythmia syndromes. Circulation 2004, 110, 2119–2124. [Google Scholar] [CrossRef] [Green Version]
- Cheung, J.W.; Meli, A.C.; Xie, W.; Mittal, S.; Reiken, S.; Wronska, A.; Xu, L.; Steinberg, J.S.; Markowitz, S.M.; Iwai, S.; et al. Short-coupled polymorphic ventricular tachycardia at rest linked to a novel ryanodine receptor (RyR2) mutation: Leaky RyR2 channels under non-stress conditions. Int. J. Cardiol. 2015, 180, 228–236. [Google Scholar] [CrossRef] [Green Version]
- George, C.H.; Jundi, H.; Thomas, N.L.; Fry, D.L.; Lai, F.A. Ryanodine receptors and ventricular arrhythmias: Emerging trends in mutations, mechanisms and therapies. J. Mol. Cell. Cardiol. 2007, 42, 34–50. [Google Scholar] [CrossRef] [PubMed]
- Zhao, Y.T.; Valdivia, C.R.; Gurrola, G.B.; Hernández, J.J.; Valdivia, H.H. Arrhythmogenic mechanisms in ryanodine receptor channelopathies. Sci. China Life Sci. 2015, 58, 54–558. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wleklinski, M.J.; Kannankeril, P.J.; Knollmann, B.C. Molecular and tissue mechanisms of catecholaminergic polymorphic ventricular tachycardia. J. Physiol. 2020, 598, 2817–2834. [Google Scholar] [CrossRef] [PubMed]
- Basso, C.; Thiene, G.; Corrado, D.; Angelini, A.; Nava, A.; Valente, M. Arrhythmogenic right ventricular cardiomyopathy. Dysplasia, dystrophy, or myocarditis? Circulation 1996, 94, 983–991. [Google Scholar] [CrossRef] [PubMed]
- Thiene, G.; Corrado, D.; Basso, C. Arrhythmogenic right ventricular cardiomyopathy/dysplasia. Orphanet J. Rare Dis. 2007, 2, 45. [Google Scholar] [CrossRef]
- Mallat, Z.; Tedgui, A.; Fontaliran, F.; Frank, R.; Durigon, M.; Fontaine, G. Evidence of apoptosis in arrhythmogenic right ventricular dysplasia. N. Engl. J. Med. 1996, 335, 1190–1196. [Google Scholar] [CrossRef]
- Valente, M.; Calabrese, F.; Thiene, G.; Angelini, A.; Basso, C.; Nava, A.; Rossi, L. In vivo evidence of apoptosis in arrhythmogenic right ventricular cardiomyopathy. Am. J. Pathol. 1998, 152, 479–484. [Google Scholar]
- Yang, F.; Zhu, X.H.; Zhang, Q.; Sun, N.X.; Ji, Y.X.; Ma, J.Z.; Xiao, B.; Ding, H.X.; Sun, S.H.; Li, W. Genomic characteristics of gender dysphoria patients and identification of rare mutations in RYR3 gene. Sci. Rep. 2017, 7, 8339. [Google Scholar] [CrossRef]
- Gong, S.; Su, B.B.; Tovar, H.; Mao, C.; Gonzalez, V.; Liu, Y.; Lu, Y.; Wang, K.S.; Xu, C. Polymorphisms within RYR3 gene are associated with risk and age at onset of hypertension, diabetes, and Alzheimer’s disease. Am. J. Hypertens. 2018, 31, 818–826. [Google Scholar] [CrossRef] [Green Version]
- Nelson, T.E. Abnormality in calcium release from skeletal sarcoplasmic reticulum of pigs susceptible to malignant hyperthermia. J. Clin. Investig. 1983, 72, 862–870. [Google Scholar] [CrossRef] [Green Version]
- Otsu, K.; Nishida, K.; Kimura, Y.; Kuzuya, T.; Hori, M.; Kamada, T.; Tada, M. The point mutation Arg615→Cys in the Ca2+ release channel of skeletal sarcoplasmic reticulum is responsible for hypersensitivity to caffeine and halothane in malignant hyperthermia. J. Biol. Chem. 1994, 269, 9413–9415. [Google Scholar] [CrossRef] [PubMed]
- Richter, M.; Schleithoff, L.; Deufel, T.; Lehmann-Horn, F.; Herrmann-Frank, A. Functional characterization of a distinct ryanodine receptor mutation in human malignant hyperthermia susceptible muscle. J. Biol. Chem. 1997, 272, 5256–5260. [Google Scholar] [CrossRef] [Green Version]
- Nelson, T.E. Halothane effects on human malignant hyperthermia skeletal muscle single calcium-release channels in planar lipid bilayers. Anesthesiology 1992, 76, 588–595. [Google Scholar] [CrossRef] [PubMed]
- Connelly, T.J.; El Hayek, R.; Rusy, B.F.; Coronado, R. Volatile anesthetics selectively alter [3H]ryanodine binding to skeletal and cardiac ryanodine receptors. Biochem. Biophys. Res. Commun. 1992, 186, 595–600. [Google Scholar] [CrossRef] [PubMed]
- Ohnishi, S.T.; Taylor, S.; Gronert, G.A. Calcium-induced Ca2+ release from sarcoplasmic reticulum of pigs susceptible to malignant hyperthermia. The effects of halothane and dantrolene. FEBS Lett. 1983, 161, 103–107. [Google Scholar] [CrossRef] [Green Version]
- Beeler, T.; Gable, K. Effect of halothane on Ca2+-induced Ca2+ release from sarcoplasmic reticulum vesicles isolated from rat skeletal muscle. Biochim. Biophys. Acta 1985, 821, 142–152. [Google Scholar] [CrossRef]
- Diaz-Sylvester, P.L.; Porta, M.; Copello, J.A. Halothane modulation of skeletal muscle ryanodine receptors: Dependence on Ca2+, Mg2+ and ATP. Am. J. Physiol. 2008, 294, C1103–C1112. [Google Scholar] [CrossRef] [Green Version]
- Davies, D.N.; Steward, A.; Allott, P.R.; Mapleson, W.W. A comparison of arterial and arterialized venous concentrations of halothane. Br. J. Anaesth. 1972, 44, 548–550. [Google Scholar] [CrossRef] [Green Version]
- Eckenhoff, R.G.; Johansson, J.S. On the relevance of ‘clinically relevant concentrations’ of inhaled anesthetics in in vitro experiments. Anesthesiology 1999, 91, 856–860. [Google Scholar] [CrossRef] [PubMed]
- Laver, D.R.; Attia, J.; Oldmeadow, C.; Quail, A.W. Cardiac calcium release channel (ryanodine receptor 2) regulation by halogenated anesthetics. Anesthesiology 2017, 126, 495–506. [Google Scholar] [CrossRef] [PubMed]
- Duke, A.M.; Hopkins, P.M.; Steele, D.S. Mg2+ dependence of halothane-induced Ca2+ release from the sarcoplasmic reticulum in rat skeletal muscle. J. Physiol. 2003, 551, 447–454. [Google Scholar] [CrossRef] [PubMed]
- Duke, A.M.; Hopkins, P.M.; Halsal, P.J.; Steele, D.S. Mg2+ dependence of Ca2+ release from the sarcoplasmic reticulum induced by sevoflurane or halothane in skeletal muscle from humans susceptible to malignant hyperthermia. Br. J. Anaesth. 2006, 97, 320–328. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Duke, A.M.; Hopkins, P.M.; Halsal, J.P.; Steele, D.S. Mg2+ dependence of halothane-induced Ca2+ release from the sarcoplasmic reticulum in skeletal muscle from humans susceptible to malignant hyperthermia. Anesthesiology 2004, 101, 1339–1346. [Google Scholar] [CrossRef] [PubMed]
- Launikonis, B.S.; Stephenson, D.G. Effects of Mg2+ on Ca2+ release from sarcoplasmic reticulum of skeletal muscle fibres from yabby (crustacean) and rat. J. Physiol. 2000, 526, 299312. [Google Scholar] [CrossRef]
- Louis, C.F.; Zualkernan, K.; Roghair, T.; Mickelson, J.R. The effects of volatile anesthetics on calcium regulation by malignant hyperthermia-susceptible sarcoplasmic reticulum. Anesthesiology 1992, 77, 114–125. [Google Scholar] [CrossRef] [PubMed]
- Ayuk, J.; Gittoes, N.J. How should hypomagnesaemia be investigated and treated? Clin. Endocrinol. 2011, 75, 743–746. [Google Scholar] [CrossRef]
- Owen, V.J.; Taske, N.L.; Lamb, G.D. Reduced Mg2+ inhibition of Ca2+ release in muscle fibers of pigs susceptible to malignant hyperthermia. Am. J. Physiol. 1997, 272, C203–C211. [Google Scholar] [CrossRef] [PubMed]
- Mickelson, J.R.; Ross, J.A.; Reed, B.K.; Louis, C.F. Enhanced Ca2+-induced calcium release by isolated sarcoplasmic reticulum vesicles from malignant hyperthermia susceptible pig muscle. Biochim. Biophys. Acta 1986, 862, 318–328. [Google Scholar] [CrossRef]
- Laver, D.R.; Owen, V.J.; Junankar, P.R.; Taske, N.L.; Dulhunty, A.F.; Lamb, G.D. Reduced inhibitory effect of Mg2+ on ryanodine receptor-Ca2+ release channels in malignant hyperthermia. Biophys. J. 1997, 73, 1913–1924. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nelson, T.E.; Flewellen, E.H.; Belt, M.W.; Kennamer, D.L.; Winsett, O.E.; Bee, D.E. Calcium and magnesium content of skeletal muscle. Studies in subjects undergoing diagnostic testing for malignant hyperthermia. Br. J. Anaesth. 1987, 59, 730–734. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yost, C.S. Tandem pore domain K channels: An important site of volatile anesthetic action. Curr. Drug Targets 2000, 1, 207–217. [Google Scholar] [CrossRef]
- Gray, A.T.; Zhao, B.B.; Kindler, C.H.; Winegar, B.D.; Mazurek, M.J.; Xu, J.; Chavez, R.A.; Forsayeth, J.R.; Yost, C.S. Volatile anesthetics activate the human tandem pore domain baseline K+ channel KCNK5. Anesthesiology 2000, 92, 1722–1730. [Google Scholar] [CrossRef] [PubMed]
- Xu, Y.; Seto, T.; Tang, P.; Firestone, L. NMR study of volatile anesthetic binding to nicotinic acetylcholine receptors. Biophys. J. 2000, 78, 746–751. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bondarenko, V.; Yushmanov, V.E.; Xu, Y.; Tang, P. NMR study of general anesthetic interaction with nAChR beta(2) subunit. Biophys. J. 2008, 94, 1681–1688. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yamamoto, T.; El-Hayek, R.; Ikemoto, N. Postulated role of interdomain interaction within the ryanodine receptor in Ca(2+) channel regulation. J. Biol. Chem. 2000, 275, 11618–11625. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yamamoto, T.; Ikemoto, N. Spectroscopic monitoring of local conformational changes during the intramolecular domain-domain interaction of the ryanodine receptor. Biochemistry 2002, 41, 1492–1501. [Google Scholar] [CrossRef] [PubMed]
- Kobayashi, S.; Yamamoto, T.; Parness, J.; Ikemoto, N. Antibody probe study of Ca2+ channel regulation by interdomain interaction within the ryanodine receptor. Biochem. J. 2004, 380, 561–569. [Google Scholar] [CrossRef] [Green Version]
- Valdivia, H.H.; Hogan, K.; Coronado, R. Altered binding site for Ca2+ in the ryanodine receptor of human malignant hyperthermia. Am. J. Physiol. 1991, 261, C237–C245. [Google Scholar] [CrossRef]
- Durham, W.J.; Aracena-Parks, P.; Long, C.; Rossi, A.E.; Goonasekera, S.A.; Boncompagni, S.; Galvan, D.L.; Gilman, C.P.; Baker, M.R.; Shirokova, N.; et al. RyR1 S-nitrosylation underlies environmental heat stroke and sudden death in Y522S RyR1 knockin mice. Cell 2008, 133, 53–65. [Google Scholar] [CrossRef] [Green Version]
- Iyer, K.A.; Hu, Y.; Nayak, A.R.; Kurebayashi, N.; Murayama, T.; Samsó, M. Structural mechanism of two gain-of-function cardiac and skeletal RyR mutations at an equivalent site by cryo-EM. Sci. Adv. 2020, 6, eabb2964. [Google Scholar] [CrossRef] [PubMed]
- Woll, K.A.; Haji-Ghassemi, O.; Van Petegem, F. Pathological conformations of disease mutant Ryanodine Receptors revealed by cryo-EM. Nat. Commun. 2021, 12, 807. [Google Scholar] [CrossRef]
- Fill, M.; Stefani, E.; Nelson, T.E. Abnormal human sarcoplasmic reticulum Ca2+ release channels in malignant hyperthermic skeletal muscle. Biophys. J. 1991, 59, 1085–1090. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Britt, B.A.; Scott, E.; Frodis, W.; Clements, M.-J.; Endrenyi, L. Dantrolene- In vitro studies in malignant hyperthermia susceptible (MHS) and normal skeletal muscle. Can. Anaesth. Soc. J. 1984, 3, 130–154. [Google Scholar] [CrossRef] [Green Version]
- Lopez, J.R.; Allen, P.D.; Alamo, L.; Jones, D.; Sreter, F.A. Myoplasmic free [Ca2+] during a malignant hyperthermia episode in swine. Muscle Nerve 1988, 11, 82–88. [Google Scholar] [CrossRef] [PubMed]
- Foster, P.S.; Denborough, M.A. Effect of diltiazem and dantrolene on the contractility of isolated malignant hyperpyrexia-susceptible porcine skeletal muscle. Br. J. Anaesth. 1989, 62, 566–572. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cherednichenko, G.; Ward, C.W.; Feng, W.; Cabrales, E.; Michaelson, L.; Samso, M.; López, J.R.; Allen, P.D.; Pessah, I.N. Enhanced excitation-coupled calcium entry in myotubes expressing malignant hyperthermia mutation R163C is attenuated by dantrolene. Mol. Pharmacol. 2008, 73, 1203–1212. [Google Scholar] [CrossRef] [Green Version]
- Kobayashi, S.; Bannister, M.L.; Gangopadhyay, J.P.; Hamada, T.; Parness, J.; Ikemoto, N. Dantrolene stabilizes domain interactions within the ryanodine receptor. J. Biol. Chem. 2005, 280, 6580–6587. [Google Scholar] [CrossRef] [Green Version]
- Wagner, L.E.I.; Groom, L.A.; Dirksen, R.T.; Yule, D.I. Characterization of ryanodine receptor type 1 single channel activity using “on-nucleus” patch clamp. Cell Calcium 2014, 56, 96–107. [Google Scholar] [CrossRef] [Green Version]
- Oo, Y.W.; Gomez-Hurtado, N.; Walweel, K.; van Helden, D.F.; Imtiaz, M.S.; Knollmann, B.C.; Laver, D.R. Essential role of calmodulin in RyR inhibition by dantrolene. Mol. Pharmacol. 2015, 88, 57–63. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Diszházi, G.; Magyar, Z.É.; Mótyán, J.A.; Csernoch, L.; Jóna, I.; Nánási, P.P.; Almássy, J. Dantrolene requires Mg(2+) and ATP to inhibit the ryanodine receptor. Mol. Pharmacol. 2019, 96, 401–407. [Google Scholar] [CrossRef]
- Khan, S.R.; Tawakkul, M.; Sayeed, V.A.; Faustino, P.; Khan, M.A. Stability characterization, kinetics and mechanism of degradation of dantrolene in aqueous solution: Effect of pH and temperature. Pharm. Pharmacol. 2012, 3, 281–290. [Google Scholar] [CrossRef] [Green Version]
- Palnitkar, S.S.; Mickelson, J.R.; Louis, C.F.; Parness, J. Pharmacological distinction between dantrolene and ryanodine binding sites: Evidence from normal and malignant hyperthermia-susceptible porcine skeletal muscle. Biochem. J. 1997, 326, 847–852. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sengupta, C.; Meyer, U.A.; Carafoli, E. Binding of Dantrolene sodium to muscle intracellular membranes. FEBS Lett. 1980, 117, 37–38. [Google Scholar] [CrossRef] [Green Version]
- Podranski, T.; Bouillon, T.; Schumacher, P.M.; Taguchi, A.; Sessler, D.I.; Kurz, A. Compartmental pharmacokinetics of dantrolene in adults: Do malignant hyperthermia association dosing guidelines work? Anesth. Analg. 2005, 101, 1695–1699. [Google Scholar] [CrossRef] [PubMed]
- Meissner, G.; Darling, E.; Eveleth, J. Kinetics of rapid Ca2+ release by sarcoplasmic reticulum. Effects of Ca2+, Mg2+, and adenine nucleotides. Biochemistry 1986, 25, 236–244. [Google Scholar] [CrossRef] [PubMed]
- Meissner, G.; Rousseau, E.; Lai, F.A.; Liu, Q.Y.; Anderson, K.A. Biochemical characterisation of the Ca2+ release channel of skeletal and cardiac sarcoplasmic reticulum. Mol. Cell. Biochem. 1988, 82, 59–65. [Google Scholar] [CrossRef]
- Laver, D.R.; Lenz, G.K.; Lamb, G.D. Regulation of the calcium release channel from rabbit skeletal muscle by the nucleotides ATP, AMP, IMP and adenosine. J. Physiol. 2001, 537, 763–778. [Google Scholar] [CrossRef] [PubMed]
- Lamb, G.D.; Stephenson, D. Effect of Mg2+ on the control of Ca2+ release in skeletal muscle fibres of the toad. J. Physiol. 1991, 434, 507–528. [Google Scholar] [CrossRef]
- Lamb, G.D.; Stephenson, D.G. Importance of Mg2+ in excitation–contraction coupling in skeletal muscle. News Physiol. Sci. 1992, 7, 270–274. [Google Scholar] [CrossRef]
- Lamb, G.D.; Stephenson, D.G. Effects of intracellular pH and [Mg2+] on excitation–contraction coupling in skeletal muscle fibres of the rat. J. Physiol. 1994, 478, 331–339. [Google Scholar] [CrossRef] [Green Version]
- Tripathy, A.; Xu, L.; Mann, G.; Meissner, G. Calmodulin activation and inhibition of skeletal muscle Ca2+ release channel (ryanodine receptor). Biophys. J. 1995, 69, 106–111. [Google Scholar] [CrossRef] [Green Version]
- Rodney, G.G.; Williams, B.Y.; Strasburg, G.M.; Beckingham, K.; Hamilton, S.L. Regulation of RYR1 activity by Ca2+ and calmodulin. Biochemistry 2000, 39, 7807–7812. [Google Scholar] [CrossRef] [PubMed]
- Haeseleer, F.; Palczewski, K. Calmodulin and Ca2+-binding proteins (CaBPs): Variations on a theme. Adv. Exp. Med. Biol. 2002, 514, 303–317. [Google Scholar] [CrossRef]
- Eger, E.I.; Smith, N.T.; Stoeliting, R.K.; Cullen, D.J.; Kadis, L.B.; Whitcher, C.E. Cardiovascular effects of halothane in man. Anesthesiology 1970, 32, 396–409. [Google Scholar] [CrossRef]
- Brown, B.R.; Crout, J.R. A comparative study of the effects of five general anesthetics on myocardial contractility. Anesthesiology 1971, 34, 236–245. [Google Scholar] [CrossRef] [PubMed]
- Calverley, R.K.; Smith, N.T.; Prys-Roberts, C.; Eger, E.I.; Jones, C.W. Cardiovascular effects of enflurane anesthesia during controlled ventilation in man. Anesth. Analg. 1978, 57, 619–628. [Google Scholar] [CrossRef] [PubMed]
- Housmans, P.R.; Murat, I. Comparative effects of halothane, enflurane, and isoflurane at equipotent anesthetic concentrations on isolated ventricular myocardium of the ferret: I. Contractility. Anesthesiology 1988, 69, 451–463. [Google Scholar] [CrossRef] [PubMed]
- Lynch, C.; Frazer, M.J. Depressant effects of the volatile anesthetics upon rat and amphibian ventricular myocardium: Insights into anesthetic mechanisms of action. Anesthesiology 1989, 70, 511–522. [Google Scholar] [CrossRef]
- Komai, H.; Rusy, B.F. Direct effect of halothane and isoflurane on the function of the sarcoplasmic reticulum. Anesthesiology 1990, 72, 694–698. [Google Scholar] [CrossRef] [PubMed]
- Wheeler, D.M.; Rice, R.T.; Hansford, R.G.; Lakatta, E.G. The effect of halothane on the free intracellular calcium concentration of isolated rat heart cells. Anesthesiology 1988, 69, 578–583. [Google Scholar] [CrossRef]
- Wilde, D.W.; Knight, P.R.; Sheth, N.; Williams, B.S. Halothane alters control of intracellular Ca2+ mobilization in single rat ventricular myocytes. Anesthesiology 1991, 75, 1075–1086. [Google Scholar] [CrossRef]
- Herland, J.S.; Julian, F.J.; Stephenson, D.G. Halothane increases Ca2+ efflux via Ca2+ channels of sarcoplasmic reticulum in chemically skinned rat myocardium. J. Physiol. 1990, 426, 1–18. [Google Scholar] [CrossRef] [PubMed]
- Wheeler, D.M.; Katz, A.; Rice, T.; Hansford, R.G. Volatile anesthetic effects on sarcoplasmic reticulum Ca content and sarcolemmal Ca flux in isolated rat cardiac cell suspensions. Anesthesiology 1994, 80, 372–382. [Google Scholar] [CrossRef]
- Davies, L.A.; Hamilton, D.L.; Hopkins, P.M.; Boyett, M.R.; Harrison, S.M. Concentration-dependent inotropic effects of halothane, isoflurane and sevoflurane on rat ventricular myocytes. Br. J. Anaesth. 1999, 82, 723–730. [Google Scholar] [CrossRef] [PubMed]
- Hannon, J.D.; Cody, M.J. Effects of volatile anesthetics on sarcolemmal calcium transport and sarcoplasmic reticulum calcium content in isolated myocytes. Anesthesiology 2002, 96, 1457–1464. [Google Scholar] [CrossRef] [PubMed]
- Connelly, T.J.; Coronado, R. Activation of the Ca2+ release channel of cardiac sarcoplasmic reticulum by volatile anesthetics. Anesthesiology 1994, 81, 459–469. [Google Scholar] [CrossRef]
- Ogawa, Y.; Kurebayashi, N. The Ca-releasing action of halothane on fragmented sarcoplasmic reticulum. J. Biochem. 1982, 92, 899–905. [Google Scholar] [CrossRef] [PubMed]
- Parida, S.; Thangaswamy, C.R. Cardiac tachyarrhythmias and anaesthesia: General principles and focus on atrial fibrillation. Indian J. Anaesth. 2017, 61, 712–720. [Google Scholar] [CrossRef] [PubMed]
- Schmidt, U.; Schwinger, R.H.; Bohm, S.; Uberfuhr, P.; Kreuzer, E.; Reichart, B.; Meyer, L.; Erdmann, E.; Bohm, M. Evidence for an interaction of halothane with the L-type Ca2+ channel in human myocardium. Anesthesiology 1993, 79, 332–339. [Google Scholar] [CrossRef]
- Kojima, A.; Kitagawa, H.; Omatsu-Kanbe, M.; Matsuura, H.; Nosaka, S. Sevoflurane protects ventricular myocytes from Ca2+ paradox-mediated Ca2+ overload by blocking the activation of transient receptor potential canonical channels. Anesthesiology 2011, 115, 509–522. [Google Scholar] [CrossRef] [Green Version]
- Kojima, A.; Kitagawa, H.; Omatsu-Kanbe, M.; Matsuura, H.; Nosaka, S. Sevoflurane protects ventricular myocytes against oxidative stress-induced cellular Ca2+ overload and hypercontracture. Anesthesiology 2013, 119, 606–620. [Google Scholar] [CrossRef] [PubMed]
- Haworth, R.A.; Goknur, A.B. Inhibition of sodium/calcium exchange and calcium channels of heart cells by volatile anesthestics. Anesthesiology 1995, 82, 1255–1265. [Google Scholar] [CrossRef] [PubMed]
- Kojima, A.; Kitagawa, H.; Omatsu-Kanbe, M.; Matsuura, H.; Nosaka, S. Presence of store-operated Ca2+ entry in C57BL/6J mouse ventricular myocytes and its suppression by sevoflurane. Br. J. Anaesth. 2012, 109, 352–360. [Google Scholar] [CrossRef] [Green Version]
- Ellis, K.O.; Carpenter, J.F. Mechanisms of control of skeletal-muscle contraction by dantrolene sodium. Arch. Phys. Med. Rehabil. 1974, 55, 362–369. [Google Scholar] [CrossRef] [PubMed]
- Hatae, J.; Ohba, M.; Kawata, H. Effects of dantrolene sodium on the excitation-contraction coupling of the mammalian and amphibian cardiac muscle. J. Mol. Cell. Cardiol. 1980, 12, 857–867. [Google Scholar] [CrossRef]
- Hiraoka, M.; Kinoshita, H.; Kawano, S. Action of dantrolene sodium on electrical and mechanical activity of guinea-pig ventricular muscles. Jpn. J. Physiol. 1985, 35, 123–138. [Google Scholar] [CrossRef]
- Seidel, M.; Thomas, N.L.; Williams, A.J.; Lai, F.A.; Zissimopoulos, S. Dantrolene rescues aberrant N-terminus intersubunit interactions in mutant pro-arrhythmic cardiac ryanodine receptors. Cardiovasc. Res. 2015, 105, 118–128. [Google Scholar] [CrossRef] [Green Version]
- Paul-Pletzer, K.; Yamamoto, T.; Bhat, M.B.; Ma, J.; Ikemoto, N.; Jimenez, L.S.; Morimoto, H.; Williams, P.G.; Parness, J. Identification of a dantrolene-binding sequence on the skeletal muscle ryanodine receptor. J. Biol. Chem. 2002, 277, 34918–34923. [Google Scholar] [CrossRef] [Green Version]
- Kobayashi, S.; Wakeyama, T.; Ono, S.; Ikeda, Y.; Omura, M.; Oda, T.; Hisamatsu, Y.; Seki, K.; Satoh, A.; Hiromoto, M.; et al. A multicenter, randomized, double-blind, controlled study to evaluate the efficacy and safety of dantrolene on ventricular arrhythmia as well as mortality and morbidity in patients with chronic heart failure (SHO-IN trial): Rationale and design. J. Cardiol. 2020, 75, 454–461. [Google Scholar] [CrossRef]
- Meissner, A.; Min, J.Y.; Haake, N.; Hirt, S.; Simon, R. Dantrolene sodium improves the force-frequency relationship and beta-adregenic responsiveness in failing human myocardium. Eur. J. Heart Fail. 1999, 1, 177–186. [Google Scholar] [CrossRef] [PubMed]
- Min, J.Y.; Meissner, A.; Feng, X.; Wang, J.; Malek, S.; Wang, J.F.; Simon, R.; Morgan, J.P. Dantrolene: Effects on abnormal intracellular Ca2+ handling and inotropy in postinfarcted rat myocardium. Eur. J. Pharmacol. 2003, 471, 41–47. [Google Scholar] [CrossRef] [PubMed]
- Zamiri, N.; Massé, S.; Ramadeen, A.; Kusha, M.; Hu, X.; Azam, M.A.; Liu, J.; Lai, P.F.H.; Vigmond, E.J.; Boyle, P.M.; et al. Dantrolene improves survival after ventricular fibrillation by mitigating impaired calcium handling in animal models. Circulation 2014, 129, 875–885. [Google Scholar] [CrossRef] [Green Version]
- Nofi, C.; Zhang, K.; Tang, Y.D.; Li, Y.; Migirov, A.; Ojamaa, K.; Gerdes, A.M.; Zhang, Y. Chronic dantrolene treatment attenuates cardiac dysfunction and reduces atrial fibrillation inducibility in a rat myocardial infarction heart failure model. Heart Rhythm. O2 2020, 1, 126–135. [Google Scholar] [CrossRef] [PubMed]
- Penttinen, K.; Swan, H.; Vanninen, S.; Paavola, J.; Lahtinen, A.M.; Kontula, K.; Aalto-Setälä, K. Antiarrhythmic effects of dantrolene in patients with catecholaminergic polymorphic ventricular tachycardia and replication of the responses using iPSC models. PLoS ONE 2015, 10, e0125366. [Google Scholar] [CrossRef]
- Suetomi, T.; Yano, M.; Uchinoumi, H.; Fukuda, M.; Hino, A.; Ono, M.; Xu, X.; Tateishi, H.; Okuda, S.; Doi, M.; et al. Mutation-linked defective inter-domain interactions within ryanodine receptor cause aberrant Ca2+ release leading to catecholaminergic polymorphic ventricular tachycardia. Circulation 2011, 124, 682–694. [Google Scholar] [CrossRef] [Green Version]
- Domeier, T.L.; Roberts, C.J.; Gibson, A.K.; Hanft, L.M.; McDonald, K.S.; Segal, S.S. Dantrolene suppresses spontaneous Ca2+ release without altering excitation-contraction coupling in cardiomyocytes of aged mice. Am. J. Physiol. 2014, 307, H818–H829. [Google Scholar] [CrossRef] [Green Version]
- Frommeyer, G.; Krawczyk, J.; Ellermann, C.; Bögeholz, N.; Kochhäuser, S.; Dechering, D.G.; Fehr, M.; Eckardt, L. Ryanodine-receptor inhibition by dantrolene effectively suppresses ventricular arrhythmias in an ex vivo model of long-QT syndrome. J. Cardiovasc. Electrophysiol. 2018, 29, 471–476. [Google Scholar] [CrossRef] [PubMed]
- Robinson, V.M.; Alsalahat, I.; Freeman, S.; Antzelevitch, C.; Barajas-Martinez, H.; Venetucci, L. A carvedilol analogue, VK-II-86, prevents hypokalaemia-induced ventricular arrhythmia through novel multi-channel effects. Br. J. Pharmacol. 2022, 179, 2713–2732. [Google Scholar] [CrossRef]
- Grimm, M.; Ling, H.; Brown, J.H. Crossing signals: Relationships between β-adrenergic stimulation and CaMKII activation. Heart Rhythm. 2011, 8, 1296–1298. [Google Scholar] [CrossRef] [Green Version]
- Dobrev, D.; Wehrens, X.H. Role of RyR2 phosphorylation in heart failure and arrhythmias: Controversies around ryanodine receptor phosphorylation in cardiac disease. Circ. Res. 2014, 114, 1311–1319. [Google Scholar] [CrossRef] [Green Version]
- Gaburjakova, J.; Krejciova, E.; Gaburjakova, M. Multisite phosphorylation of the cardiac ryanodine receptor: A random or coordinated event? Pflug. Arch. 2020, 472, 1793–1807. [Google Scholar] [CrossRef]
- Yuchi, Z.; Lau, K.; Van Petegem, F. Disease mutations in the ryanodine receptor central region: Crystal structures of a phosphorylation hot spot domain. Structure 2012, 20, 1201–1211. [Google Scholar] [CrossRef] [Green Version]
- Lohse, M.J.; Engelhardt, S.; Eschenhagen, T. What is the role of β-adrenergic signaling in heart failure? Circ. Res. 2003, 93, 896–906. [Google Scholar] [CrossRef] [Green Version]
- Yang, J.; Liu, Y.; Fan, X.; Li, Z.; Cheng, Y. A pathway and network review on beta-adrenoceptor signaling and beta blockers in cardiac remodeling. Heart Fail. Rev. 2014, 19, 799–814. [Google Scholar] [CrossRef] [PubMed]
- Szentmiklosi, A.J.; Szentandrássy, N.; Hegyi, B.; Horvath, B.; Magyar, J.; Bányász, T.; Nanasi, P.P. Chemistry, physiology, and pharmacology of β-adrenergic mechanisms in the heart. Why are β-blocker antiarrhythmics superior? Curr. Pharm. Des. 2015, 21, 1030–1041. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Meyler, W.J.; Wesseling, H.; Agoston, S. The effects of dantrolene sodium on cardiac and skeletal muscle in rats. Eur. J. Pharmacol. 1976, 39, 127–131. [Google Scholar] [CrossRef] [PubMed]
- Sufu-Shimizu, Y.; Okuda, S.; Kato, T.; Nishimura, S.; Uchinoumi, H.; Oda, T.; Kobayashi, S.; Yamamoto, T.; Yano, M. Stabilizing cardiac ryanodine receptor prevents the development of cardiac dysfunction and lethal arrhythmia in Ca2+/calmodulin-dependent protein kinase IIδc transgenic mice. Biochem. Biophys. Res. Commun. 2020, 524, 431–438. [Google Scholar] [CrossRef]
- Si, D.; Azam, M.A.; Lai, P.F.H.; Zamiri, N.; Kichigina, G.; Asta, J.; Massé, S.; Bokhari, M.M.; Porta-Sánchez, A.; Labos, C.; et al. Essential role of ryanodine receptor 2 phosphorylation in the effect of azumolene on ventricular arrhythmia vulnerability in a rabbit heart model. J. Cardiovasc. Electrophysiol. 2018, 29, 1707–1715. [Google Scholar] [CrossRef] [PubMed]
- Jiang, D.; Xiao, B.; Yang, D.; Wang, R.; Choi, P.; Zhang, L.; Cheng, H.; Chen, S.R.W. RyR2 mutations linked to ventricular tachycardia and sudden death reduce the threshold for store-overload-induced Ca2+ release (SOICR). Proc. Natl. Acad. Sci. USA 2004, 101, 13062–13067. [Google Scholar] [CrossRef] [Green Version]
- Jiang, D.; Wang, R.; Xiao, B.; Kong, H.; Hunt, D.J.; Choi, P.; Zhang, L.; Chen, S.R.W. Enhanced store overload-induced Ca2+ release and channel sensitivity to luminal Ca2+ activation are common defects of RyR2 mutations linked to ventricular tachycardia and sudden death. Circ. Res. 2005, 97, 1173–1181. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jones, P.P.; Jiang, D.; Bolstad, J.; Hunt, D.J.; Zhang, L.; Demaurex, N.; Chen, S.R. Endoplasmic reticulum Ca2+ measurements reveal that the cardiac ryanodine receptor mutations linked to cardiac arrhythmia and sudden death alter the threshold for store-overload-induced Ca2+ release. Biochem. J. 2008, 412, 171–178. [Google Scholar] [CrossRef] [PubMed]
- Liu, Y.; Kimlicka, L.; Hiess, F.; Tian, X.; Wang, R.; Zhang, L.; Jones, P.P.; Van Petegem, F.; Chen, S.R. The CPVT-associated RyR2 mutation G230C enhances store overload-induced Ca2+ release and destabilizes the N-terminal domains. Biochem. J. 2013, 454, 123–131. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lehnart, S.E.; Wehrens, X.H.T.; Laitinen, P.J.; Reiken, S.R.; Deng, S.-X.; Cheng, Z.; Landry, D.W.; Kontula, K.; Swan, H.; Marks, A.R. Sudden death in familial polymorphic ventricular tachycardia associated with calcium release channel (ryanodine receptor) leak. Circulation 2004, 109, 3208–3214. [Google Scholar] [CrossRef] [Green Version]
- Jiang, D.; Jones, P.P.; Davis, D.R.; Gow, R.; Green, M.S.; Birnie, D.H.; Wayne Chen, S.R.; Gollob, M.H. Characterization of a novel mutation in the cardiac ryanodine receptor that results in catecholaminergic polymorphic ventricular tachycardia. Channels 2010, 4, 302–310. [Google Scholar] [CrossRef] [Green Version]
- Meli, A.C.; Refaat, M.M.; Dura, M.; Reiken, S.; Wronska, A.; Wojciak, J.; Carroll, J.; Scheinman, M.M.; Marks, A.R. A novel ryanodine receptor mutation linked to sudden death increases sensitivity to cytosolic calcium. Circ. Res. 2011, 109, 281–290. [Google Scholar] [CrossRef] [Green Version]
- Li, S.; Lv, T.; Yang, J.; Li, K.; Yang, Y.; Zhang, P. A gain of function ryanodine receptor 2 mutation (R1760W-RyR2) in catecholaminergic polymorphic ventricular tachycardia. Clin. Exp. Pharmacol. Physiol. 2023, 59, 39–49. [Google Scholar] [CrossRef]
- Dulhunty, A.F. Molecular changes in the cardiac RyR2 with catecholaminergic polymorphic ventricular tachycardia (CPVT). Front. Physiol. 2022, 13, 830367. [Google Scholar] [CrossRef]
- Fowler, E.D.; Zissimopoulos, S. Molecular, subcellular, and arrhythmogenic mechanisms in genetic RyR2 disease. Biomolecules 2022, 12, 1030. [Google Scholar] [CrossRef]
- Wehrens, X.H.T.; Lehnart, S.E.; Huang, F.; Vest, J.A.; Reiken, S.R.; Mohler, P.J.; Sun, J.; Guatimosim, S.; Song, L.S.; Rosemblit, N.; et al. FKBP12.6 deficiency and defective calcium release channel (ryanodine receptor) function linked to exercise-induced sudden cardiac death. Cell 2003, 113, 829–840. [Google Scholar] [CrossRef] [Green Version]
- Ikemoto, N.; Yamamoto, T. Regulation of calcium release by interdomain interaction within ryanodine receptors. Front. Biosci. 2002, 7, d671–d683. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Iyer, K.A.; Barnakov, V.; Samsó, M. Three-dimensional perspective on ryanodine receptor mutations causing skeletal and cardiac muscle-related diseases. Curr. Opin. Pharmacol. 2023, 68, 102327. [Google Scholar] [CrossRef] [PubMed]
- Miotto, M.C.; Weninger, G.; Dridi, H.; Yuan, Q.; Liu, Y.; Wronska, A.; Melville, Z.; Sittenfeld, L.; Reiken, S.; Marks, A.R. Structural analyses of human ryanodine receptor type 2 channels reveal the mechanisms for sudden cardiac death and treatment. Sci. Adv. 2022, 8, eabo1272. [Google Scholar] [CrossRef] [PubMed]
- Shan, J.; Kushnir, A.; Betzenhauser, M.J.; Reiken, S.; Li, J.; Lehnart, S.E.; Lindegger, N.; Mongillo, M.; Mohler, P.J.; Marks, A.R. Phosphorylation of the ryanodine receptor mediates the cardiac fight or flight response in mice. J. Clin. Investig. 2010, 120, 4388–4398. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, H.; Makarewich, C.A.; Kubo, H.; Wang, W.E.; Duran, J.M.; Li, Y.; Berretta, R.M.; Koch, W.J.; Chen, X.; Gao, E.; et al. Hyperphosphorylation of the cardiac ryanodine receptor at Serine 2808 is not involved in cardiac dysfunction after myocardial infarction. Circ. Res. 2012, 110, 831–840. [Google Scholar] [CrossRef] [Green Version]
- Dhindwal, S.; Lobo, J.; Cabra, V.; Santiago, D.J.; Nayak, A.R.; Dryden, K.; Samsó, M. A cryo-EM–based model of phosphorylation- and FKBP12.6-mediated allosterism of the cardiac ryanodine receptor. Sci. Signal. 2017, 10, eaai8842. [Google Scholar] [CrossRef] [PubMed]
- Yan, Z.; Bai, X.; Yan, C.; Wu, J.; Li, Z.; Xie, T.; Peng, W.; Yin, C.; Li, X.; Scheres, S.H.W.; et al. Structure of the rabbit ryanodine receptor RyR1 at near-atomic resolution. Nature 2014, 517, 50–55. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sorrentino, V.; Volpe, P. Ryanodine receptors: How many, where and why? Trends Pharmacol. Sci. 1993, 141, 98–103. [Google Scholar] [CrossRef]
- Berman, H.M.; Westbrook, J.; Feng, Z.; Gilliland, G.; Bhat, T.N.; Weissig, H.; Shindyalov, I.N.; Bourne, P.E. The Protein Data Bank. Nucleic Acids Res. 2000, 28, 235–242. [Google Scholar] [CrossRef] [Green Version]
- Wang, R.; Zhong, X.; Meng, X.; Koop, A.; Tian, X.; Jones, P.P.; Fruen, B.R.; Wagenknecht, T.; Liu, Z.; Chen, S.R. Localization of the dantrolene-binding sequence near the FK506-binding protein-binding site in the three-dimensional structure of the ryanodine receptor. J. Biol. Chem. 2011, 286, 12202–12212. [Google Scholar] [CrossRef] [Green Version]
- Doi, M.; Yano, M.; Kobayashi, S.; Kohno, M.; Tokuhisa, T.; Okuda, S.; Suetsugu, M.; Hisamatsu, Y.; Ohkusa, T.; Kohno, M.; et al. Propranolol prevents the development of heart failure by restoring FKBP12.6-mediated stabilization of ryanodine receptor. Circulation 2002, 105, 1374–1379. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yano, M.; Kobayashi, S.; Kohno, M.; Doi, M.; Tokuhisa, T.; Okuda, S.; Suetsugu, M.; Hisaoka, T.; Obayashi, M.; Ohkusa, T.; et al. FKBP12.6-mediated stabilization of calcium-release channel (ryanodine receptor) as a novel therapeutic strategy against heart failure. Circulation 2003, 107, 477–484. [Google Scholar] [CrossRef] [Green Version]
- Galfré, E.; Pitt, S.J.; Venturi, E.; Sitsapesan, M.; Zaccai, N.R.; Tsaneva-Atanasova, K.; O’Neill, S.; Sitsapesan, R. FKBP12 activates the cardiac ryanodine receptor Ca2+-release channel and is antagonised by FKBP12.6. PLoS ONE 2012, 7, e31956. [Google Scholar] [CrossRef] [Green Version]
- Lehnart, S.E.; Wehrens, X.H.; Marks, A.R. Calstabin deficiency, ryanodine receptors, and sudden cardiac death. Biochem. Biophys. Res. Commun. 2004, 322, 1267–1279. [Google Scholar] [CrossRef] [PubMed]
- Pogwizd, S.M.; Bers, D.M. Cellular basis of triggered arrhythmias in heart failure. Trends Cardiovasc. Med. 2004, 14, 61–66. [Google Scholar] [CrossRef]
- Eisner, D.A.; Kashimura, T.; O’Neill, S.C.; Venetucci, L.A.; Trafford, A.W. What role does modulation of the ryanodine receptor play in cardiac inotropy and arrhythmogenesis? J. Mol. Cell Cardiol. 2009, 46, 474–481. [Google Scholar] [CrossRef]
- Brillantes, A.B.; Ondrias, K.; Scott, A.; Kobrinsky, E.; Ondriasova, E.; Moschella, M.C.; Jayaraman, T.; Landers, M.; Ehrlich, B.E.; Marks, A.R. Stabilization of calcium release channel (ryanodine receptor) function by FK506-binding protein. Cell 1994, 77, 513–523. [Google Scholar] [CrossRef]
- Gaburjakova, M.; Gaburjakova, J.; Reiken, S.; Huang, F.; Marx, S.O.; Rosemblit, N.; Marks, A.R. FKBP12 binding modulates ryanodine receptor channel gating. J. Biol. Chem. 2001, 276, 16931–16935. [Google Scholar] [CrossRef] [Green Version]
- Avila, G.; Lee, E.H.; Perez, C.F.; Allen, P.D.; Dirksen, R.T. FKBP12 binding to RyR1 modulates excitation-contraction coupling in mouse skeletal myotubes. J. Biol. Chem. 2003, 278, 22600–22608. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Reiken, S.; Lacampagne, A.; Zhou, H.; Kherani, A.; Lehnart, S.E.; Ward, C.; Huang, F.; Gaburjakova, M.; Gaburjakova, J.; Rosemblit, N.; et al. PKA phosphorylation activates the calcium release channel (ryanodine receptor) in skeletal muscle: Defective regulation in heart failure. J. Cell Biol. 2003, 160, 919–928. [Google Scholar] [CrossRef] [PubMed]
- Samsó, M.; Shen, X.; Allen, P.D. Structural characterization of the RyR1–FKBP12 interaction. J. Mol. Biol. 2006, 356, 917–927. [Google Scholar] [CrossRef] [PubMed]
- Acimovic, I.; Refaat, M.M.; Moreau, A.; Salykin, A.; Reiken, S.; Sleiman, Y.; Souidi, M.; Přibyl, J.; Kajava, A.V.; Richard, S.; et al. Post-translational modifications and diastolic calcium leak associated to the novel RyR2-D3638A mutation lead to CPVT in patient-specific hiPSC-derived cardiomyocytes. J. Clin. Med. 2018, 7, 423. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Marx, S.O.; Reiken, S.; Hisamatsu, Y.; Jayaraman, T.; Burkhoff, D.; Rosemblit, N.; Marks, A.R. PKA phosphorylation dissociates FKBP12.6 from the calcium release channel (ryanodine receptor): Defective regulation in failing hearts. Cell 2000, 101, 365–376. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shan, J.; Betzenhauser, M.J.; Kushnir, A.; Reiken, S.; Meli, A.C.; Wronska, A.; Dura, M.; Chen, B.X.; Marks, A.R. Role of chronic ryanodine receptor phosphorylation in heart failure and β-adrenergic receptor blockade in mice. J. Clin. Investig. 2010, 120, 4375–4387. [Google Scholar] [CrossRef] [Green Version]
- Suko, J.; Maurer-Fogy, I.; Plank, B.; Bertel, O.; Wyskovsky, W.; Hohenegger, M.; Hellmann, G. Phosphorylation of serine 2843 in ryanodine receptor-calcium release channel of skeletal muscle by cAMP-, cGMP and CaM-dependent protein kinase. Biochim. Biophys. Acta 1993, 1175, 193–206. [Google Scholar] [CrossRef] [PubMed]
- Kugler, Y.; Russell, W. Speeding dantrolene preparation for treating malignant hyperthermia. Anaesth. Intensive Care 2011, 39, 84–88. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Schütte, J.; Becker, S.; Burmester, S.; Starosse, A.; Lenz, D.; Kröner, L.; Wappler, F.; Gerbershagen, M.U. Comparison of the therapeutic effectiveness of a dantrolene sodium solution and a novel nanocrystalline suspension of dantrolene sodium in malignant hyperthermia normal and susceptible pigs. Eur. J. Anaesthesiol. 2011, 28, 256–264. [Google Scholar] [CrossRef]
- Riazi, S.; Larach, M.G.; Hu, C.; Wijeysundera, D.; Massey, C.; Kraeva, N. Malignant hyperthermia in Canada: Characteristics of index anesthetics in 129 malignant hyperthermia susceptible probands. Anesth. Analg. 2014, 118, 381–387. [Google Scholar] [CrossRef] [Green Version]
- White, R.L.; Wessels, F.L.; Schawan, T.J.; Ellis, K.O. 1-[[[5-(substitutedphenyl)-2-oxazolyl]-methylene]amino]-2,4-imidazolidinediones, a new class of skeletal muscle relaxants. J. Med. Chem. 1987, 30, 263–266. [Google Scholar] [CrossRef]
- Dershwitz, M.; Sreter, F.A. Azumolene reverses episodes ofmalignant hyperthermia in susceptible swine. Anesth. Analg. 1990, 70, 253–255. [Google Scholar] [CrossRef] [PubMed]
- do Carmo, P.L.; Zapata-Sudo, G.; Trachez, M.M.; Antunes, F.; Guimarães, S.E.F.; Debom, R.; Rizzi, M.D.R.; Sudo, R.T. Intravenous administration of azumolene to reverse malignant hyperthermia in swine. J. Vet. Intern. Med. 2010, 24, 1224–1228. [Google Scholar] [CrossRef] [PubMed]
- McAvoy, J.C.; Brodsky, J.B.; Brock-Utne, J. Pennywise and a pound foolish: The advantage of dantrolene nanosuspension (Ryanodex) in the treatment of malignant hyperthermia. Anesth. Analg. 2019, 129, e201–e202. [Google Scholar] [CrossRef] [PubMed]
- Abou, M.B.; Sun, L.; Wei, H. Approaches to optimizing dantrolene neuroprotection for the treatment of Alzheimer’s disease. Curr. Alzheimer Res. 2020, 17, 324–328. [Google Scholar] [CrossRef] [PubMed]
- Wang, J.; Shi, Y.; Yu, S.; Wang, Y.; Meng, Q.; Liang, G.; Eckenhoff, M.F.; Wei, H. Intranasal administration of dantrolene increased brain concentration and duration. PLoS ONE 2020, 15, e0229156. [Google Scholar] [CrossRef] [Green Version]


{kind=link}
| Effect of Dantrolene | Presence | Temperature | RyR1 Type | References | ||
|---|---|---|---|---|---|---|
| ATP or Analog | Mg2+ | CaM | ||||
| Yes | ▪ | ▪ | ND | RT | MH | [87] |
| No | ▫ | ▫ | ND | RT | MH | [97] |
| No | ▫ | ▫ | ND | 37 °C | ||
| No | ▫ | ▫ | ND | RT | WT | [28] |
| Yes | ▪ | ▪ | ND | RT | ||
| No | ▪ | ▫ | ND | RT | WT | [120] |
| No | ▫ | ▫ | ND | RT | WT | [89] |
| No | ▪ | ▪ | ND | RT | ||
| No | ▪ | ▫ | ▪ | RT | WT | [118] |
| No | ▪ | ▫ | ▪ | 35 °C | ||
| No | ▫ | ▫ | ND | RT | WT | [121] |
| Yes | ▫ | ▫ | ▪ | RT | ||
| No | ▪ | ▫ | ▫ | RT | WT | [122] |
| No | ▪ | ▫ | ▪ | RT | ||
| Yes | ▪ | ▪ | ▫ | RT | ||
| No | ▪ | ▫ | ND | RT | WT | [119] |
| Yes | ▪ | ▫ | ND | 37 °C | ||
| Yes | ▪ | ▫ | ▪ | RT | ||
| No | ▪ | ▪ | ND | 36 °C | MH, WT | [29] |
| No | ▪ | ▪ | ▪ | 19 °C | MH | |
| Yes | ▪ | ▪ | ▪ | 36 °C | MH, WT | |
| No | ▫ | ▫ | ▪ | 37 °C | MH | [30] |
| Yes | ▪ | ▫ | ▪ | 37 °C | MH, WT | |
| Yes | ▪ | ▪ | ▪ | 37 °C | MH |
| RyR Isoform | Kd (nM) | Experimental Approach | References |
|---|---|---|---|
| RyR1 | 365 ± 50 | [3H]dantrolene binding | [124] |
| 277 ± 25 | [3H]dantrolene binding | [18] | |
| 5 | [14C]dantrolene binding | [125] | |
| RyR2 | 2000 | [14C]dantrolene binding | [125] |
| RyR3 | ~Kd of RyR1 | Inhibition of [3H]ryanodine binding | [30] |
| Effect of Dantrolene | Presence | Temperature | RyR2 Type | References | ||
|---|---|---|---|---|---|---|
| ATP or Analog | Mg2+ | CaM | ||||
| No | ▪ | ▪ | ▫ | 36 °C | WT | [29] |
| No | ▪ | ▪ | ▪ | 36 °C | ||
| No | ▪ | ▫ | ▪ | 37 °C | WT | [30] |
| No | ▫ | ▫ | ▫ | RT | WT | [121] |
| Yes | ▫ | ▫ | ▪ | RT | ||
| Yes | ▫ | ▫ | ▪ | RT | WT | [92] |
| No | ▪ | ▪ | ND | 35 °C | WT | [31] |
| Yes | ▪ | ▪ | ND | 35 °C | WT-P | |
| Yes | ▪ | ▪ | ND | 35 °C | WT-FH | [31] |
| Effect of Dantrolene | Presence | Temperature | RyR3 Type | References | ||
|---|---|---|---|---|---|---|
| ATP or Analog | Mg2+ | CaM | ||||
| No | ▪ | ▫ | ▪ | RT | WT | [30] |
| No | ▫ | ▫ | ▪ | 37 °C | ||
| Yes | ▪ | ▫ | ▪ | 37 °C |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Gaburjakova, J.; Gaburjakova, M. Molecular Aspects Implicated in Dantrolene Selectivity with Respect to Ryanodine Receptor Isoforms. Int. J. Mol. Sci. 2023, 24, 5409. https://doi.org/10.3390/ijms24065409
Gaburjakova J, Gaburjakova M. Molecular Aspects Implicated in Dantrolene Selectivity with Respect to Ryanodine Receptor Isoforms. International Journal of Molecular Sciences. 2023; 24(6):5409. https://doi.org/10.3390/ijms24065409
Chicago/Turabian StyleGaburjakova, Jana, and Marta Gaburjakova. 2023. "Molecular Aspects Implicated in Dantrolene Selectivity with Respect to Ryanodine Receptor Isoforms" International Journal of Molecular Sciences 24, no. 6: 5409. https://doi.org/10.3390/ijms24065409