
Prognostic Value of Monocarboxylate Transporter 1 Overexpression in Cancer: A Systematic Review

 , , , and
, , , and
Abstract
:
1. Introduction
1.1. Glucose Metabolism in Cancer
1.2. Monocarboxylate Transporters: Key Players in Cancer Aggressiveness
1.3. MCT1 as a Target for Cancer Therapy
2. Materials and Methods
2.1. Aim, Study Design and Eligibility Criteria
2.2. Search Strategy
2.3. Study Selection Strategy
2.4. Quality Assessment and Data Synthesis
3. Results
3.1. Literature Search
3.2. Characterization and Qualitative Assessment of the Studies
3.3. Global Quality
3.4. Association of MCT1 Expression with Clinicopathological Parameters
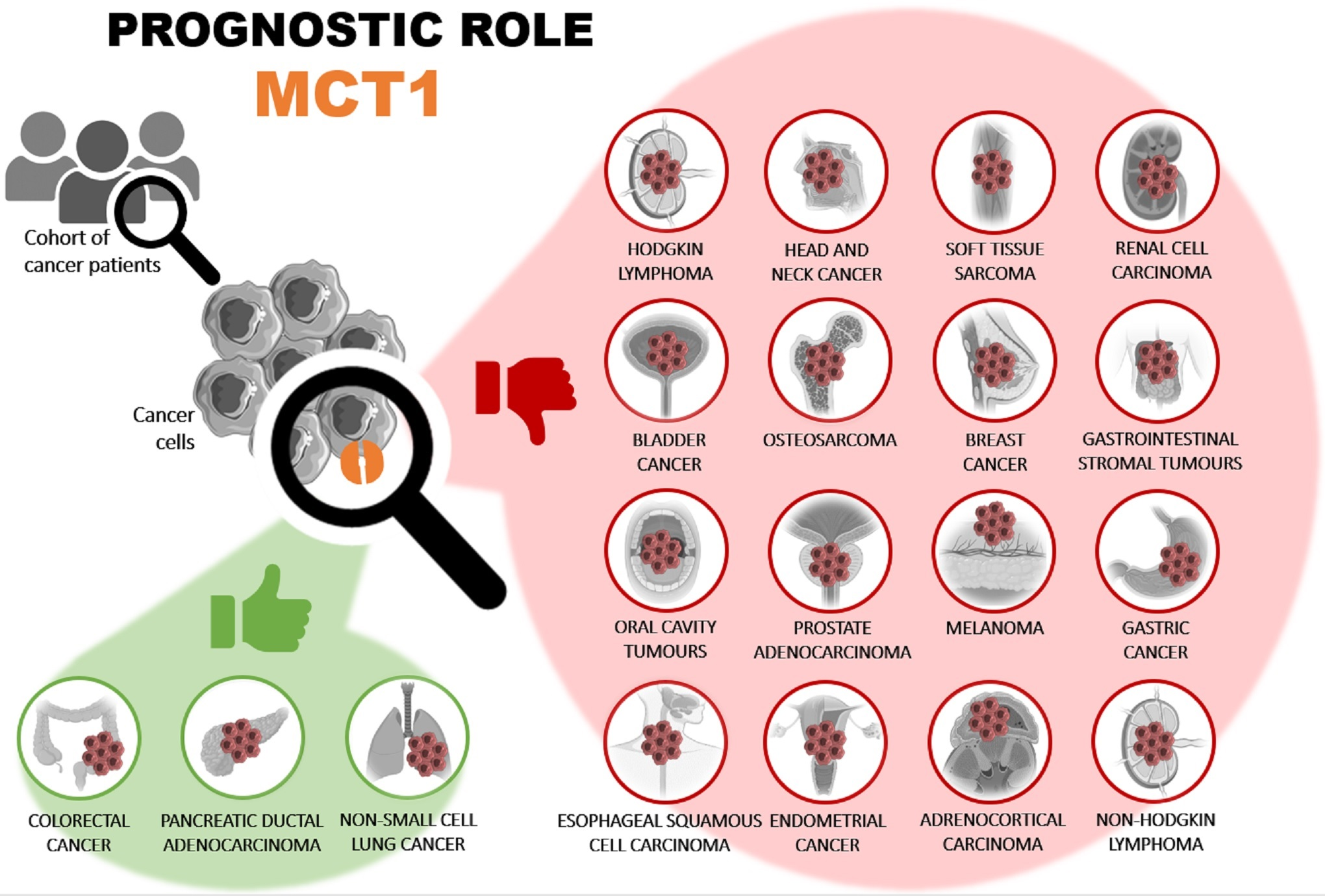
3.5. Prognostic Value of MCT1 Expression
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Hsu, P.P.; Sabatini, D.M. Cancer cell metabolism: Warburg and beyond. Cell 2008, 134, 703–707. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bose, S.; Zhang, C.; Le, A. Glucose Metabolism in Cancer: The Warburg Effect and Beyond. Adv. Exp. Med. Biol. 2021, 1311, 3–15. [Google Scholar] [CrossRef] [PubMed]
- de la Cruz-Lopez, K.G.; Castro-Munoz, L.J.; Reyes-Hernandez, D.O.; Garcia-Carranca, A.; Manzo-Merino, J. Lactate in the Regulation of Tumor Microenvironment and Therapeutic Approaches. Front. Oncol. 2019, 9, 1143. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Miranda-Goncalves, V.; Goncalves, C.S.; Granja, S.; Vieira de Castro, J.; Reis, R.M.; Costa, B.M.; Baltazar, F. MCT1 Is a New Prognostic Biomarker and Its Therapeutic Inhibition Boosts Response to Temozolomide in Human Glioblastoma. Cancers 2021, 13, 3468. [Google Scholar] [CrossRef]
- Frezza, C. Metabolism and cancer: The future is now. Br. J. Cancer 2020, 122, 133–135. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Payen, V.L.; Mina, E.; Van Hée, V.F.; Porporato, P.E.; Sonveaux, P. Monocarboxylate transporters in cancer. Mol. Metab. 2020, 33, 48–66. [Google Scholar] [CrossRef] [PubMed]
- Warburg, O.; Wind, F.; Negelein, E. The Metabolism of Tumors in the Body. J. Gen. Physiol. 1927, 8, 519–530. [Google Scholar] [CrossRef] [Green Version]
- Baltazar, F.; Afonso, J.; Costa, M.; Granja, S. Lactate Beyond a Waste Metabolite: Metabolic Affairs and Signaling in Malignancy. Front. Oncol. 2020, 10, 231. [Google Scholar] [CrossRef] [Green Version]
- Pavlova, N.N.; Thompson, C.B. The Emerging Hallmarks of Cancer Metabolism. Cell Metab. 2016, 23, 27–47. [Google Scholar] [CrossRef] [Green Version]
- Xiao, C.; Tian, H.; Zheng, Y.; Yang, Z.; Li, S.; Fan, T.; Xu, J.; Bai, G.; Liu, J.; Deng, Z.; et al. Glycolysis in tumor microenvironment as a target to improve cancer immunotherapy. Front. Cell Dev. Biol. 2022, 10, 1013885. [Google Scholar] [CrossRef]
- San-Millán, I.; Brooks, G.A. Reexamining cancer metabolism: Lactate production for carcinogenesis could be the purpose and explanation of the Warburg Effect. Carcinogenesis 2017, 38, 119–133. [Google Scholar] [CrossRef] [Green Version]
- Vander Heiden, M.G.; Cantley, L.C.; Thompson, C.B. Understanding the Warburg effect: The metabolic requirements of cell proliferation. Science 2009, 324, 1029–1033. [Google Scholar] [CrossRef] [Green Version]
- Pereira-Nunes, A.; Afonso, J.; Granja, S.; Baltazar, F. Lactate and Lactate Transporters as Key Players in the Maintenance of the Warburg Effect. Adv. Exp. Med. Biol. 2020, 1219, 51–74. [Google Scholar] [CrossRef]
- Schiliro, C.; Firestein, B.L. Mechanisms of Metabolic Reprogramming in Cancer Cells Supporting Enhanced Growth and Proliferation. Cells 2021, 10, 1056. [Google Scholar] [CrossRef]
- Abbaszadeh, Z.; Çeşmeli, S.; Avcı, B. Crucial players in glycolysis: Cancer progress. Gene 2020, 726, 144158. [Google Scholar] [CrossRef]
- Danhier, P.; Bański, P.; Payen, V.L.; Grasso, D.; Ippolito, L.; Sonveaux, P.; Porporato, P.E. Cancer metabolism in space and time: Beyond the Warburg effect. Biochim. Biophys. Acta Bioenerg. 2017, 1858, 556–572. [Google Scholar] [CrossRef]
- Hirschhaeuser, F.; Sattler, U.G.; Mueller-Klieser, W. Lactate: A metabolic key player in cancer. Cancer Res. 2011, 71, 6921–6925. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Courtnay, R.; Ngo, D.C.; Malik, N.; Ververis, K.; Tortorella, S.M.; Karagiannis, T.C. Cancer metabolism and the Warburg effect: The role of HIF-1 and PI3K. Mol. Biol. Rep. 2015, 42, 841–851. [Google Scholar] [CrossRef]
- De Saedeleer, C.J.; Copetti, T.; Porporato, P.E.; Verrax, J.; Feron, O.; Sonveaux, P. Lactate activates HIF-1 in oxidative but not in Warburg-phenotype human tumor cells. PLoS ONE 2012, 7, e46571. [Google Scholar] [CrossRef] [Green Version]
- Wanka, C.; Brucker, D.P.; Bähr, O.; Ronellenfitsch, M.; Weller, M.; Steinbach, J.P.; Rieger, J. Synthesis of cytochrome C oxidase 2: A p53-dependent metabolic regulator that promotes respiratory function and protects glioma and colon cancer cells from hypoxia-induced cell death. Oncogene 2012, 31, 3764–3776. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Baltazar, F.; Valente, D.; Afonso, J.; Queirós, O.; Granja, S. New horizons on pH regulators as cancer biomarkers and targets for pharmacological intervention. In An Innovative Approach to Understanding and Treating Cancer: Targeting pH from Etiopathogenesis to New Therapeutic Avenues; Academic Press: Cambridge, MA, USA, 2020. [Google Scholar]
- Halestrap, A.P. The SLC16 gene family—structure, role and regulation in health and disease. Mol. Asp. Med. 2013, 34, 337–349. [Google Scholar] [CrossRef] [PubMed]
- Bovenzi, C.D.; Hamilton, J.; Tassone, P.; Johnson, J.; Cognetti, D.M.; Luginbuhl, A.; Keane, W.M.; Zhan, T.; Tuluc, M.; Bar-Ad, V.; et al. Prognostic Indications of Elevated MCT4 and CD147 across Cancer Types: A Meta-Analysis. Biomed. Res. Int. 2015, 2015, 242437. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Puri, S.; Juvale, K. Monocarboxylate transporter 1 and 4 inhibitors as potential therapeutics for treating solid tumours: A review with structure-activity relationship insights. Eur. J. Med. Chem. 2020, 199, 112393. [Google Scholar] [CrossRef]
- Sun, X.; Wang, M.; Wang, M.; Yao, L.; Li, X.; Dong, H.; Li, M.; Sun, T.; Liu, X.; Liu, Y.; et al. Role of Proton-Coupled Monocarboxylate Transporters in Cancer: From Metabolic Crosstalk to Therapeutic Potential. Front. Cell Dev. Biol. 2020, 8, 651. [Google Scholar] [CrossRef]
- Pértega-Gomes, N.; Vizcaíno, J.R.; Gouveia, C.; Jerónimo, C.; Henrique, R.M.; Lopes, C.; Baltazar, F. Monocarboxylate transporter 2 (MCT2) as putative biomarker in prostate cancer. Prostate 2013, 73, 763–769. [Google Scholar] [CrossRef] [Green Version]
- Afonso, J.; Santos, L.L.; Miranda-Gonçalves, V.; Morais, A.; Amaro, T.; Longatto-Filho, A.; Baltazar, F. CD147 and MCT1-potential partners in bladder cancer aggressiveness and cisplatin resistance. Mol. Carcinog. 2015, 54, 1451–1466. [Google Scholar] [CrossRef] [Green Version]
- Yan, C.; Yang, F.; Zhou, C.; Chen, X.; Han, X.; Liu, X.; Ma, H.; Zheng, W. MCT1 promotes the cisplatin-resistance by antagonizing Fas in epithelial ovarian cancer. Int. J. Clin. Exp. Pathol. 2015, 8, 2710–2718. [Google Scholar]
- Hu, K.Y.; Wang, D.G.; Liu, P.F.; Cao, Y.W.; Wang, Y.H.; Yang, X.C.; Hu, C.X.; Sun, L.J.; Niu, H.T. Targeting of MCT1 and PFKFB3 influences cell proliferation and apoptosis in bladder cancer by altering the tumor microenvironment. Oncol. Rep. 2016, 36, 945–951. [Google Scholar] [CrossRef] [Green Version]
- Zhao, Z.; Wu, M.S.; Zou, C.; Tang, Q.; Lu, J.; Liu, D.; Wu, Y.; Yin, J.; Xie, X.; Shen, J.; et al. Downregulation of MCT1 inhibits tumor growth, metastasis and enhances chemotherapeutic efficacy in osteosarcoma through regulation of the NF-kappaB pathway. Cancer Lett. 2014, 342, 150–158. [Google Scholar] [CrossRef]
- Sonveaux, P.; Vegran, F.; Schroeder, T.; Wergin, M.C.; Verrax, J.; Rabbani, Z.N.; De Saedeleer, C.J.; Kennedy, K.M.; Diepart, C.; Jordan, B.F.; et al. Targeting lactate-fueled respiration selectively kills hypoxic tumor cells in mice. J. Clin. Investig. 2008, 118, 3930–3942. [Google Scholar] [CrossRef] [Green Version]
- Guan, X.; Morris, M.E. In Vitro and In Vivo Efficacy of AZD3965 and Alpha-Cyano-4-Hydroxycinnamic Acid in the Murine 4T1 Breast Tumor Model. AAPS J. 2020, 22, 84. [Google Scholar] [CrossRef]
- Guan, X.; Bryniarski, M.A.; Morris, M.E. In Vitro and In Vivo Efficacy of the Monocarboxylate Transporter 1 Inhibitor AR-C155858 in the Murine 4T1 Breast Cancer Tumor Model. AAPS J. 2018, 21, 3. [Google Scholar] [CrossRef] [PubMed]
- Hanson, D.J.; Nakamura, S.; Amachi, R.; Hiasa, M.; Oda, A.; Tsuji, D.; Itoh, K.; Harada, T.; Horikawa, K.; Teramachi, J.; et al. Effective impairment of myeloma cells and their progenitors by blockade of monocarboxylate transportation. Oncotarget 2015, 6, 33568–33586. [Google Scholar] [CrossRef] [Green Version]
- Polański, R.; Hodgkinson, C.L.; Fusi, A.; Nonaka, D.; Priest, L.; Kelly, P.; Trapani, F.; Bishop, P.W.; White, A.; Critchlow, S.E.; et al. Activity of the monocarboxylate transporter 1 inhibitor AZD3965 in small cell lung cancer. Clin. Cancer Res. 2014, 20, 926–937. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- US National Library of Medicine. ClinicalTrials.gov. Available online: https://www.clinicaltrials.gov/ct2/show/NCT01791595?term=AZD3965&cond=cancer&draw=2&rank=1 (accessed on 30 May 2021).
- Silva, A.; Antunes, B.; Batista, A.; Pinto-Ribeiro, F.; Baltazar, F.; Afonso, J. In Vivo Anticancer Activity of AZD3965: A Systematic Review. Molecules 2021, 27, 181. [Google Scholar] [CrossRef]
- Javaeed, A.; Ghauri, S.K. MCT4 has a potential to be used as a prognostic biomarker—a systematic review and meta-analysis. Oncol. Rev. 2019, 13, 403. [Google Scholar] [CrossRef] [Green Version]
- Page, M.J.; McKenzie, J.E.; Bossuyt, P.M.; Boutron, I.; Hoffmann, T.C.; Mulrow, C.D.; Shamseer, L.; Tetzlaff, J.M.; Akl, E.A.; Brennan, S.E.; et al. The PRISMA 2020 statement: An updated guideline for reporting systematic reviews. BMJ 2021, 372, n71. [Google Scholar] [CrossRef]
- Critical Appraisal Skills Programme (2018). CASP Cohort Study Checklist. Available online: https://casp-uk.net/images/checklist/documents/CASP-Cohort-Study-Checklist/CASP-Cohort-Study-Checklist_2018.pdf (accessed on 15 October 2022).
- Pinheiro, C.; Miranda-Gonçalves, V.; Longatto-Filho, A.; Vicente, A.L.; Berardinelli, G.N.; Scapulatempo-Neto, C.; Costa, R.F.; Viana, C.R.; Reis, R.M.; Baltazar, F.; et al. The metabolic microenvironment of melanomas: Prognostic value of MCT1 and MCT4. Cell Cycle 2016, 15, 1462–1470. [Google Scholar] [CrossRef] [Green Version]
- Simões-Sousa, S.; Granja, S.; Pinheiro, C.; Fernandes, D.; Longatto-Filho, A.; Laus, A.C.; Alves, C.D.; Suárez-Peñaranda, J.M.; Pérez-Sayáns, M.; Lopes Carvalho, A.; et al. Prognostic significance of monocarboxylate transporter expression in oral cavity tumors. Cell Cycle 2016, 15, 1865–1873. [Google Scholar] [CrossRef] [Green Version]
- Latif, A.; Chadwick, A.L.; Kitson, S.J.; Gregson, H.J.; Sivalingam, V.N.; Bolton, J.; McVey, R.J.; Roberts, S.A.; Marshall, K.M.; Williams, K.J.; et al. Monocarboxylate Transporter 1 (MCT1) is an independent prognostic biomarker in endometrial cancer. BMC Clin. Pathol. 2017, 17, 27. [Google Scholar] [CrossRef]
- Silva, E.C.A.; Cárcano, F.M.; Bonatelli, M.; Zaia, M.G.; Morais-Santos, F.; Baltazar, F.; Lopes, L.F.; Scapulatempo-Neto, C.; Pinheiro, C. The clinicopathological significance of monocarboxylate transporters in testicular germ cell tumors. Oncotarget 2018, 9, 20386–20398. [Google Scholar] [CrossRef] [Green Version]
- Pinheiro, C.; Penna, V.; Morais-Santos, F.; Abrahão-Machado, L.F.; Ribeiro, G.; Curcelli, E.C.; Olivieri, M.V.; Morini, S.; Valença, I.; Ribeiro, D.; et al. Characterization of monocarboxylate transporters (MCTs) expression in soft tissue sarcomas: Distinct prognostic impact of MCT1 sub-cellular localization. J. Transl. Med. 2014, 12, 118. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Leu, M.; Kitz, J.; Pilavakis, Y.; Hakroush, S.; Wolff, H.A.; Canis, M.; Rieken, S.; Schirmer, M.A. Monocarboxylate transporter-1 (MCT1) protein expression in head and neck cancer affects clinical outcome. Sci. Rep. 2021, 11, 4578. [Google Scholar] [CrossRef] [PubMed]
- Sukeda, A.; Nakamura, Y.; Nishida, Y.; Kojima, M.; Gotohda, N.; Akimoto, T.; Ochiai, A. Expression of Monocarboxylate Transporter 1 Is Associated With Better Prognosis and Reduced Nodal Metastasis in Pancreatic Ductal Adenocarcinoma. Pancreas 2019, 48, 1102–1110. [Google Scholar] [CrossRef]
- Hiltunen, N.; Rintala, J.; Väyrynen, J.P.; Böhm, J.; Karttunen, T.J.; Huhta, H.; Helminen, O. Monocarboxylate Transporters 1 and 4 and Prognosis in Small Bowel Neuroendocrine Tumors. Cancers 2022, 14, 2552. [Google Scholar] [CrossRef]
- Yokoo, S.; Fujiwara, T.; Yoshida, A.; Uotani, K.; Morita, T.; Kiyono, M.; Hasei, J.; Nakata, E.; Kunisada, T.; Iwata, S.; et al. Liquid Biopsy Targeting Monocarboxylate Transporter 1 on the Surface Membrane of Tumor-Derived Extracellular Vesicles from Synovial Sarcoma. Cancers 2021, 13, 1823. [Google Scholar] [CrossRef]
- Pinheiro, C.; Granja, S.; Longatto-Filho, A.; Faria, A.M.; Fragoso, M.C.; Lovisolo, S.M.; Lerário, A.M.; Almeida, M.Q.; Baltazar, F.; Zerbini, M.C. Metabolic reprogramming: A new relevant pathway in adult adrenocortical tumors. Oncotarget 2015, 6, 44403–44421. [Google Scholar] [CrossRef] [Green Version]
- de Oliveira, A.T.; Pinheiro, C.; Longatto-Filho, A.; Brito, M.J.; Martinho, O.; Matos, D.; Carvalho, A.L.; Vazquez, V.L.; Silva, T.B.; Scapulatempo, C.; et al. Co-expression of monocarboxylate transporter 1 (MCT1) and its chaperone (CD147) is associated with low survival in patients with gastrointestinal stromal tumors (GISTs). J. Bioenerg. Biomembr. 2012, 44, 171–178. [Google Scholar] [CrossRef]
- Dell'Anno, I.; Barone, E.; Mutti, L.; Rassl, D.M.; Marciniak, S.J.; Silvestri, R.; Landi, S.; Gemignani, F. Tissue expression of lactate transporters (MCT1 and MCT4) and prognosis of malignant pleural mesothelioma (brief report). J. Transl. Med. 2020, 18, 341. [Google Scholar] [CrossRef]
- Mikkilineni, L.; Whitaker-Menezes, D.; Domingo-Vidal, M.; Sprandio, J.; Avena, P.; Cotzia, P.; Dulau-Florea, A.; Gong, J.; Uppal, G.; Zhan, T.; et al. Hodgkin lymphoma: A complex metabolic ecosystem with glycolytic reprogramming of the tumor microenvironment. Semin. Oncol. 2017, 44, 218–225. [Google Scholar] [CrossRef]
- Sáenz-de-Santa-Maria, I.; Bernardo-Castiñeira, C.; Secades, P.; Bernaldo-de-Quirós, S.; Rodrigo, J.P.; Astudillo, A.; Chiara, M.D. Clinically relevant HIF-1alpha-dependent metabolic reprogramming in oropharyngeal squamous cell carcinomas includes coordinated activation of CAIX and the miR-210/ISCU signaling axis, but not MCT1 and MCT4 upregulation. Oncotarget 2017, 8, 13730–13746. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bonatelli, M.; Fornari, I.F.; Bernécule, P.N.; Pinheiro, L.E.; Costa, R.F.A.; Longatto-Filho, A.; Junior, J.N.A.; Silva, E.C.A.; Cárcano, F.M.; Pinheiro, C. Expression of Glycolysis-Related Proteins in Cancer of Unknown Primary Origin. Front. Oncol. 2021, 11, 682665. [Google Scholar] [CrossRef] [PubMed]
- Andersen, S.; Solstad, O.; Moi, L.; Donnem, T.; Eilertsen, M.; Nordby, Y.; Ness, N.; Richardsen, E.; Busund, L.T.; Bremnes, R.M. Organized metabolic crime in prostate cancer: The coexpression of MCT1 in tumor and MCT4 in stroma is an independent prognosticator for biochemical failure. Urol. Oncol. Semin. Orig. Investig. 2015, 33, 338.e9–338.e17. [Google Scholar] [CrossRef] [PubMed]
- Pértega-Gomes, N.; Vizcaíno, J.R.; Attig, J.; Jurmeister, S.; Lopes, C.; Baltazar, F. A lactate shuttle system between tumour and stromal cells is associated with poor prognosis in prostate cancer. BMC Cancer 2014, 14, 352. [Google Scholar] [CrossRef]
- Roseweir, A.K.; Clark, J.; McSorley, S.T.; van Wyk, H.C.; Quinn, J.A.; Horgan, P.G.; McMillan, D.C.; Park, J.H.; Edwards, J. The association between markers of tumour cell metabolism, the tumour microenvironment and outcomes in patients with colorectal cancer. Int. J. Cancer 2019, 144, 2320–2329. [Google Scholar] [CrossRef]
- Martins, S.F.; Amorim, R.; Viana-Pereira, M.; Pinheiro, C.; Costa, R.F.; Silva, P.; Couto, C.; Alves, S.; Fernandes, S.; Vilaça, S.; et al. Significance of glycolytic metabolism-related protein expression in colorectal cancer, lymph node and hepatic metastasis. BMC Cancer 2016, 16, 535. [Google Scholar] [CrossRef] [Green Version]
- Kim, Y.; Choi, J.W.; Lee, J.H.; Kim, Y.S. Expression of lactate/H+ symporters MCT1 and MCT4 and their chaperone CD147 predicts tumor progression in clear cell renal cell carcinoma: Immunohistochemical and The Cancer Genome Atlas data analyses. Hum. Pathol. 2015, 46, 104–112. [Google Scholar] [CrossRef]
- Cao, Y.W.; Liu, Y.; Dong, Z.; Guo, L.; Kang, E.H.; Wang, Y.H.; Zhang, W.; Niu, H.T. Monocarboxylate transporters MCT1 and MCT4 are independent prognostic biomarkers for the survival of patients with clear cell renal cell carcinoma and those receiving therapy targeting angiogenesis. Urol. Oncol. Semin. Orig. Investig. 2018, 36, 311.e15–311.e25. [Google Scholar] [CrossRef]
- Chen, X.; Chen, X.; Liu, F.; Yuan, Q.; Zhang, K.; Zhou, W.; Guan, S.; Wang, Y.; Mi, S.; Cheng, Y. Monocarboxylate transporter 1 is an independent prognostic factor in esophageal squamous cell carcinoma. Oncol. Rep. 2019, 41, 2529–2539. [Google Scholar] [CrossRef]
- Zheng, S.; Liu, Q.; Tan, D.; Shen, T.; Zhang, X.; Han, X.; Lu, X. Clinicopathological and prognostic involvements of MCT1, MCT4 and IL7R in esophageal squamous cell carcinoma. Res. Sq. 2019. [Google Scholar] [CrossRef]
- Zhao, H.; Chen, Y.; Liao, Y.P.; Chen, H.M.; Yang, Q.H.; Xiao, Y.; Luo, J.; Chen, Z.Z.; Yi, L.; Hu, G.Y. Immunohistochemical evaluation and prognostic value of monocarboxylate transporter 1 (MCT1) and 4 (MCT4) in T-cell non-Hodgkin lymphoma. Clin. Exp. Med. 2023, 23, 55–64. [Google Scholar] [CrossRef]
- Afonso, J.; Pinto, T.; Simões-Sousa, S.; Schmitt, F.; Longatto-Filho, A.; Pinheiro, C.; Marques, H.; Baltazar, F. Clinical significance of metabolism-related biomarkers in non-Hodgkin lymphoma—MCT1 as potential target in diffuse large B cell lymphoma. Cell. Oncol. 2019, 42, 303–318. [Google Scholar] [CrossRef] [Green Version]
- Eskuri, M.; Kemi, N.; Kauppila, J.H. Monocarboxylate Transporters 1 and 4 and MTCO1 in Gastric Cancer. Cancers 2021, 13, 2142. [Google Scholar] [CrossRef]
- Wang, C.; Wen, Z.; Xie, J.; Zhao, Y.; Zhao, L.; Zhang, S.; Liu, Y.; Xue, Y.; Shi, M. MACC1 mediates chemotherapy sensitivity of 5-FU and cisplatin via regulating MCT1 expression in gastric cancer. Biochem. Biophys. Res. Commun. 2017, 485, 665–671. [Google Scholar] [CrossRef]
- Tong, Y.H.; Hu, X.P.; Xiang, X.P.; Fang, L. High expression of monocarboxylate transporter 4 (MCT 4), but not MCT 1, predicts poor prognosis in patients with non-small cell lung cancer. Transl. Cancer Res. 2021, 10, 1336–1345. [Google Scholar] [CrossRef]
- Eilertsen, M.; Andersen, S.; Al-Saad, S.; Kiselev, Y.; Donnem, T.; Stenvold, H.; Pettersen, I.; Al-Shibli, K.; Richardsen, E.; Busund, L.T.; et al. Monocarboxylate transporters 1–4 in NSCLC: MCT1 is an independent prognostic marker for survival. PLoS ONE 2014, 9, e105038. [Google Scholar] [CrossRef] [Green Version]
- Giatromanolaki, A.; Sivridis, E.; Arelaki, S.; Koukourakis, M.I. Expression of enzymes related to glucose metabolism in non-small cell lung cancer and prognosis. Exp. Lung Res. 2017, 43, 167–174. [Google Scholar] [CrossRef]
- Johnson, J.M.; Cotzia, P.; Fratamico, R.; Mikkilineni, L.; Chen, J.; Colombo, D.; Mollaee, M.; Whitaker-Menezes, D.; Domingo-Vidal, M.; Lin, Z.; et al. MCT1 in Invasive Ductal Carcinoma: Monocarboxylate Metabolism and Aggressive Breast Cancer. Front. Cell Dev. Biol. 2017, 5, 27. [Google Scholar] [CrossRef] [Green Version]
- Li, Z.; Wu, Q.; Sun, S.; Wu, J.; Li, J.; Zhang, Y.; Wang, C.; Yuan, J.; Sun, S. Monocarboxylate transporters in breast cancer and adipose tissue are novel biomarkers and potential therapeutic targets. Biochem. Biophys. Res. Commun. 2018, 501, 962–967. [Google Scholar] [CrossRef]
- Sun, Y.; Liang, S.; Li, T.; Peng, C.; Yang, Y.; Lin, Y.; Ma, Y.; Dong, C. Prognostic implications of combined high expression of CD47 and MCT1 in breast cancer: A retrospective study during a 10-year period. Transl. Cancer Res. 2022, 11, 29–42. [Google Scholar] [CrossRef]
- Choi, J.W.; Kim, Y.; Lee, J.H.; Kim, Y.S. Prognostic significance of lactate/proton symporters MCT1, MCT4, and their chaperone CD147 expressions in urothelial carcinoma of the bladder. Urology 2014, 84, 245.e9–245.e15. [Google Scholar] [CrossRef] [PubMed]
- Zhang, G.; Zhang, Y.; Dong, D.; Wang, F.; Ma, X.; Guan, F.; Sun, L. MCT1 regulates aggressive and metabolic phenotypes in bladder cancer. J. Cancer 2018, 9, 2492–2501. [Google Scholar] [CrossRef] [PubMed]
- Afonso, J.; Santos, L.L.; Morais, A.; Amaro, T.; Longatto-Filho, A.; Baltazar, F. Metabolic coupling in urothelial bladder cancer compartments and its correlation to tumor aggressiveness. Cell Cycle 2016, 15, 368–380. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ferlay, J.; Colombet, M.; Soerjomataram, I.; Parkin, D.M.; Piñeros, M.; Znaor, A.; Bray, F. Cancer statistics for the year 2020: An overview. Int. J. Cancer 2021, 149, 778–789. [Google Scholar] [CrossRef]
- Guo, X.; Chen, C.; Liu, B.; Wu, Y.; Chen, Y.; Zhou, X.; Huang, X.; Li, X.; Yang, H.; Chen, Z.; et al. Genetic variations in monocarboxylate transporter genes as predictors of clinical outcomes in non-small cell lung cancer. Tumor Biol. 2015, 36, 3931–3939. [Google Scholar] [CrossRef]
- Jin, P.; Jiang, J.; Xie, N.; Zhou, L.; Huang, Z.; Zhang, L.; Qin, S.; Fu, S.; Peng, L.; Gao, W.; et al. MCT1 relieves osimertinib-induced CRC suppression by promoting autophagy through the LKB1/AMPK signaling. Cell Death Dis. 2019, 10, 615. [Google Scholar] [CrossRef] [Green Version]
- Nakayama, Y.; Torigoe, T.; Inoue, Y.; Minagawa, N.; Izumi, H.; Kohno, K.; Yamaguchi, K. Prognostic significance of monocarboxylate transporter 4 expression in patients with colorectal cancer. Exp. Ther. Med. 2012, 3, 25–30. [Google Scholar] [CrossRef] [Green Version]
- Kong, S.C.; Nohr-Nielsen, A.; Zeeberg, K.; Reshkin, S.J.; Hoffmann, E.K.; Novak, I.; Pedersen, S.F. Monocarboxylate Transporters MCT1 and MCT4 Regulate Migration and Invasion of Pancreatic Ductal Adenocarcinoma Cells. Pancreas 2016, 45, 1036–1047. [Google Scholar] [CrossRef]
- Baek, G.; Tse, Y.F.; Hu, Z.; Cox, D.; Buboltz, N.; McCue, P.; Yeo, C.J.; White, M.A.; DeBerardinis, R.J.; Knudsen, E.S.; et al. MCT4 defines a glycolytic subtype of pancreatic cancer with poor prognosis and unique metabolic dependencies. Cell Rep. 2014, 9, 2233–2249. [Google Scholar] [CrossRef] [Green Version]
- Landras, A.; Reger de Moura, C.; Jouenne, F.; Lebbe, C.; Menashi, S.; Mourah, S. CD147 Is a Promising Target of Tumor Progression and a Prognostic Biomarker. Cancers 2019, 11, 1803. [Google Scholar] [CrossRef] [Green Version]
- Massari, F.; Ciccarese, C.; Santoni, M.; Iacovelli, R.; Mazzucchelli, R.; Piva, F.; Scarpelli, M.; Berardi, R.; Tortora, G.; Lopez-Beltran, A.; et al. Metabolic phenotype of bladder cancer. Cancer Treat. Rev. 2016, 45, 46–57. [Google Scholar] [CrossRef]
- Huhta, H.; Helminen, O.; Palomäki, S.; Kauppila, J.H.; Saarnio, J.; Lehenkari, P.P.; Karttunen, T.J. Intratumoral lactate metabolism in Barrett’s esophagus and adenocarcinoma. Oncotarget 2017, 8, 22894–22902. [Google Scholar] [CrossRef] [Green Version]
- Alves, V.A.; Pinheiro, C.; Morais-Santos, F.; Felipe-Silva, A.; Longatto-Filho, A.; Baltazar, F. Characterization of monocarboxylate transporter activity in hepatocellular carcinoma. World J. Gastroenterol. 2014, 20, 11780–11787. [Google Scholar] [CrossRef] [Green Version]
- Halford, S.E.R.; Jones, P.; Wedge, S.; Hirschberg, S.; Katugampola, S.; Veal, G.; Payne, G.; Bacon, C.; Potter, S.; Griffin, M.; et al. A first-in-human first-in-class (FIC) trial of the monocarboxylate transporter 1 (MCT1) inhibitor AZD3965 in patients with advanced solid tumours. J. Clin. Oncol. 2017, 35. [Google Scholar] [CrossRef]
- Halford, S.E.R.; Walter, H.; McKay, P.; Townsend, W.; Linton, K.; Heinzmann, K.; Dragoni, I.; Brotherton, L.; Veal, G.; Siskos, A.; et al. Phase I expansion study of the first-in-class monocarboxylate transporter 1 (MCT1) inhibitor AZD3965 in patients with diffuse large B-cell lymphoma (DLBCL) and Burkitt lymphoma (BL). J. Clin. Oncol. 2021, 39, 3115. [Google Scholar] [CrossRef]
- Pavlides, S.; Whitaker-Menezes, D.; Castello-Cros, R.; Flomenberg, N.; Witkiewicz, A.K.; Frank, P.G.; Casimiro, M.C.; Wang, C.; Fortina, P.; Addya, S.; et al. The reverse Warburg effect: Aerobic glycolysis in cancer associated fibroblasts and the tumor stroma. Cell Cycle 2009, 8, 3984–4001. [Google Scholar] [CrossRef] [Green Version]
- Whitaker-Menezes, D.; Martinez-Outschoorn, U.E.; Lin, Z.; Ertel, A.; Flomenberg, N.; Witkiewicz, A.K.; Birbe, R.C.; Howell, A.; Pavlides, S.; Gandara, R.; et al. Evidence for a stromal-epithelial “lactate shuttle” in human tumors: MCT4 is a marker of oxidative stress in cancer-associated fibroblasts. Cell Cycle 2011, 10, 1772–1783. [Google Scholar] [CrossRef] [Green Version]
- Wang, Z.H.; Peng, W.B.; Zhang, P.; Yang, X.P.; Zhou, Q. Lactate in the tumour microenvironment: From immune modulation to therapy. eBioMedicine 2021, 73, 103627. [Google Scholar] [CrossRef]
- Liu, X.; Zhang, Y.; Li, W.; Zhou, X. Lactylation, an emerging hallmark of metabolic reprogramming: Current progress and open challenges. Front. Cell Dev. Biol. 2022, 10, 972020. [Google Scholar] [CrossRef]



{kind=link}
{kind=link}
{kind=link}
| Reference | Collected Samples (Country) | Cancer Type | n | Age | Sex | Association of MCT1 Expression (Cancer Cells) with Prognosis |
|---|---|---|---|---|---|---|
| de Oliveira et al., 2012 [51] | Brazil and Portugal | Gastrointestinal stromal tumours | Brazil: 51 Portugal: 13 | NR | NR | ↓DFS when ↑MCT1 + ↑CD147 |
| Choi et al., 2014 [74] | South Korea | Bladder cancer | 360 | Median: 69 y (range 23–97) | M: 311 (86,4%) F: 49 (13,6%) | ↓OS ↓RFS |
| Eilertsen et al., 2014 [69] | Norway | Non-small cell lung cancer | 335 | ≤65 y: 156 (47%) >65 y: 179 (53%) | M: 253 (76%) F: 82 (24%) | ↑OS ↑DSS |
| Pértega-Gomes et al., 2014 [57] | Portugal | Prostate cancer | 480 | NR | M: 480 (100%) | No association |
| Pinheiro et al., 2014 [45] | Brazil | Soft tissue sarcoma | 85 | ≤51 y: 37 (43.5%) >51 y: 48 (56.5%) | M: 52 (61.2%) F: 33 (38.8%) | ↓OS; MCT1 nuclear expression: ↑OS |
| Polanski et al., 2014 [35] | United Kingdom | Small cell lung cancer | 58 | Median: 61 y (range 35–83) | M: 33 (57%) F: 25 (43%) | ↓OS |
| Zhao et al., 2014 [30] | China | Osteosarcoma | 61 | NR | NR | ↓OS |
| Afonso et al., 2015 [27] | Portugal | Bladder cancer | 114 | Median: 70 y (range 41–86) | M: 94 (82.5%) F: 20 (17.5%) | ↓DFS + ↓OS when ↑MCT1 + ↑CD147 |
| Andersen et al., 2015 [56] | Norway | Prostate adenocarcinoma | 535 | ≤65 y: 357 (67%) >65 y: 178 (33.0%) | M: 535 (100%) | ↓BFFS when stromal ↑MCT4 |
| Kim et al., 2015 [60] | South Korea | Renal cell carcinoma | 180 | Median: 58 y (range 25–83) | M: 127 (70.6%) F: 53 (29.4%) | ↓PFS |
| Afonso et al., 2016 [76] | Portugal | Bladder cancer | 111 | Median: 70 y (range: 41–86) | M: 91 (82%) F:20 (18%) | ↓OS in cisplatin treated-patients when ↑MCT1 CN + ↑MCT4 CH + ↑MCT4 S *1 |
| Martins et al., 2016 [59] | Portugal | Colorectal cancer | 500 | ≤45 y: 23 (4.6%) >45 y: 477 (95.4%) | M: 314 (62.8%) F: 186 (37.2%) | ↑OS |
| Pinheiro et al., 2016 [41] | Brazil | Melanoma | 357 | Median: 58.3 y (range 25–83) | M: 174 (48.9%) F: 183 (51.1%) | ↓OS |
| Pinheiro et al., 2016 [50] | Brazil | Adrenocortical carcinoma | 78 | <40.6 y: 34 (43.6%) ≥40.6 y: 44 (56.4%) | M: 17 (21.8%) F:61 (78.2%) | ↓OS |
| Sousa-Simões et al., 2016 [42] | Brazil and Spain | Oral cavity cancer | Brazil: 90 Spain: 45 | ≤60 y: 69 (50.4%) >60 y: 66 (48.2%) | M: 103 (75.2%) F: 32 (23.4%) | ↓OS when ↑MCT1 + ↑MCT4 + ↓MCT2 |
| Giatromanolaki et al., 2017 [70] | Greece | Non-small cell lung cancer | 98 | Median: 68 y (range 32–81) | M: 86 (88%) F: 12 (12%) | No association |
| Johnson et al., 2017 [71] | United States | Breast cancer | 257 | Mean: 57.2 y (range 26.9–97.8) | F: 257 (100%) | ↑RR, ↓PFS |
| Latif et al., 2017 [43] | United Kingdom | Endometrial cancer | 90 | Median: 67 y (range 57.7–74) | F: 90 (100%) | ↓RFS, ↓CSS, ↓OS |
| Mikkilineni et al., 2017 [53] | United States | Hodgkin lymphoma | 22 | ≤40 y: 14 (64%) >40 y: 8 (36%) | M: 13 (59%) F: 9 (41%) | ↓PFS for high metabolic heterogeneity group *2 |
| Sáenz-de-Santa-María et al., 2017 [54] | Spain | Oropharyngeal squamous cell carcinoma | 249 | ≤58 y: 134 (53.8%) >58 y: 115 (46.2%) | M: 240 (96.4%) F: 9 (3.6%) | No association |
| Wang et al., 2017 [67] | China | Gastric cancer | 85 | ≤50 y: 60 (70.6%) >50 y: 25 (24.7%) | M: 52 (61.2%) F: 33 (38.8%) | ↓OS, ↓PFS |
| Cao et al., 2018 [61] | China | Renal cell carcinoma | 150 | Mean: 56.9 y (TT)/54.6 y (no TT) | M: 84 (56%) F: 66 (44%) | ↓OS, ↓PFS |
| Li et al., 2018 [72] | China | Breast cancer | 146 | ≤64 y: 80.8 (81.6%) >65 y: 28 (19.2%) | F: 146 (100%) | ↓RFS |
| Roseweir et al., 2018 [58] | Scotland | Colorectal cancer | 150 | ≤65 y: 58 (39%) >65: 92 (61%) | M: 83 (55%) F: 67 (45%) | No association |
| Silva et al., 2018 [44] | Brazil | Testicular germ cell cancer | 149 | Mean: 32.3 y (range 18–73) | M: 149 (100%) | No association |
| Zhang et al., 2018 [75] | China | Bladder cancer | 124 | Median: 65 y (range 30–88) | M: 100 (80.6%) F: 24 (19.4%) | ↓OS |
| Afonso et al., 2019 [65] | Portugal | Non-Hodgkin lymphoma | 104 | Median: 67 y (range 19–97) | M: 54 (51.9%) F: 50 (48.1%) | No association |
| Chen et al., 2019 [62] | China | Esophageal squamous cell carcinoma | 103 | ≤60 y: 46 (45%) >60 y: 57 (55%) | M: 68 (66%) F: 35 (34%) | ↓OS, ↓PFS |
| Sukeda et al., 2019 [47] | China | Pancreatic ductal adenocarcinoma | 240 | <65 y: 86 (36%) ≥65 y: 154 (64%) | M: 154 (64%) F: 86 (36%) | ↑OS, ↑PFS |
| Zheng et al., 2019 [63] | China | Esophageal squamous cell carcinoma | 86 | ≤60 y: 55 (64%) >60 y: 31 (36%) | M: 60 (67.5%) F: 26 (32.5%) | ↓OS |
| Dell’Anno et al., 2020 [52] | United Kingdom | Malignant pleural mesothelioma | 135 | NR | M: 109 (80.7%) F: 26 (19.3%) | No association |
| Bonatelli et al., 2021 [55] | Brazil | Cancer of unknown primary origin | 118 | <59.5 y: 57 (48.3%) ≥59.5 y: 61 (51.7%) | M: 55 (47%) F: 63 (53%) | ↓OS when ↑MCT1 + ↑CD147 |
| Eskuri et al., 2021 [66] | Finland | Gastric cancer | 560 | ≤69 y: 283 (49.5%) >69 y: 277 (50.5% | M: 341 (60.9%) F:219 (39.1%) | No association |
| Leu et al., 2021 [46] | Germany | Head and neck cancer | 82 | Median: 56.4 y (range 20–88) | M: 67 (81.7%) F: 15 (18.3%) | ↓OS , ↓PFS |
| Tong et al., 2021 [68] | Zhejiang | Non-small cell lung cancer | 100 | Median: 59 y (range 40–79) | M: 74 (74%) F: 26 (26%) | No association |
| Yokoo et al., 2021 [49] | Japan | Synovial sarcoma | 29 | ≤40 y: 17 (56.7%) >41 y: 12 (40%) | M: 14 (46.7%) F: 15 (53.3%) | ↓OS |
| Hiltunen et al., 2022 [48] | Finland | Small bowel neuroendocrine cancer | 109 | Median: 66 y (range 56–72) | M: 60 (55%) F: 49 (45%) | No association |
| Sun et al., 2022 [73] | China | Breast cancer | 137 | <50 y: 43 (31.4%) ≥50 y: 94 (68.6%) | F: 137 (100%) | ↓OS, ↓DFS |
| Zhao et al., 2022 [64] | China | Non-Hodgkin lymphoma | 38 | ≤60 y: 22 (57.9%) >60 y: 16 (42.1%) | M: 28 (73.7%) F: 10 (26.3%) | ↓OS, ↓PFS |
| Reference | Primary Antibody | Immunohistochemistry Kit/System | Measurement of MCT1 Expression | Positive Cut-Off |
|---|---|---|---|---|
| de Oliveira et al., 2012 [51] | AB3538P, rabbit polyclonal, Chemicon International | R.T.U. VECTASTAIN Elite ABC Kit, Vector Laboratories | I + E | ≥3 |
| Choi et al., 2014 [74] | AB3538P, rabbit polyclonal, Chemicon International | ChemMate EnVision Kit (Dako) | I + MC | >5 |
| Eilertsen et al., 2014 [69] | AB3538P, rabbit polyclonal, Chemicon International | Ventana BenchMark XT (Ventana Medical Systems Inc.) | E | >1.5 |
| Pértega-Gomes et al., 2014 [57] | sc-50329, mouse monoclonal, Santa Cruz Biotechnology | R.T.U. VECTASTAIN Elite ABC Kit, Vector Laboratories | I + E | ≥4 |
| Pinheiro et al., 2014 [45] | AB3538P, rabbit polyclonal, Chemicon International | R.T.U. VECTASTAIN Elite ABC Kit, Vector Laboratories | I + E | ≥3 |
| Polanski et al., 2014 [35] | Produced in house | Envision Kit (Dako) | I × E | Mean |
| Zhao et al., 2014 [30] | NR, Millipore | NR | I + E | ≥3 |
| Afonso et al., 2015 [27] | AB3538P, rabbit polyclonal, Chemicon International | R.T.U. VECTASTAIN Elite ABC Kit, Vector Laboratories | I + E | ≥4 |
| Andersen et al., 2015 [56] | AB3538P, rabbit polyclonal, Chemicon International | Ventana BenchMark XT (Ventana Medical Systems Inc.) | E | >2 |
| Kim et al., 2015 [60] | AB3538P, rabbit polyclonal, Chemicon International | BondTM Polymer Refine Detection kit | I + MC | >15 |
| Afonso et al., 2016 [76] | AB3538P, rabbit polyclonal, Chemicon International | R.T.U. VECTASTAIN Elite ABC Kit, Vector Laboratories | I + E | ≥3 |
| Martins et al., 2016 [59] | AB3538P, rabbit polyclonal, Chemicon International | R.T.U. VECTASTAIN Elite ABC Kit, Vector Laboratories | I + E | ≥3 |
| Pinheiro et al., 2016 [41] | AB3538P, rabbit polyclonal, Chemicon International | R.T.U. VECTASTAIN Elite ABC Kit, Vector Laboratories | I + E | ≥3 |
| Pinheiro et al., 2016 [50] | AB3538P, rabbit polyclonal, Chemicon International | R.T.U. VECTASTAIN Elite ABC Kit, Vector Laboratories | I + E | ≥3 |
| Sousa-Simões et al., 2016 [42] | AB3538P, rabbit polyclonal, Chemicon International | R.T.U. VECTASTAIN Elite ABC Kit, Vector Laboratories | I + E | ≥4 |
| Giatromanolaki et al., 2017 [70] | AB85021, rabbit polyclonal, Chemicon International | Thermo-kit Ultravision, Quanto-HRP kit | E | >50% |
| Johnson et al., 2017 [71] | NR | Ventana Discovery ULTRA | I | ≥2 |
| Latif et al., 2017 [43] | sc-365501, mouse monoclonal, Santa Cruz Biotechnology | Leica BOND_MAX and Bond Polymer Refine Detection Kit | I | ≥200 |
| Mikkilineni et al., 2017 [53] | NR | NR | I + E | 0–2 |
| Sáenz-de-Santa-María et al., 2017 [54] | NR, Abcam | Dako Autostainer Plus | I × E | Median |
| Wang et al., 2017 [67] | NR | Dako Envision System | I × E | 3–12 |
| Cao et al., 2018 [61] | NR, Abcam | NR | I | >2 |
| Li et al., 2018 [72] | NR | NR | I × E | NR |
| Roseweir et al., 2018 [58] | Produced in house | NR | I × %E | 0–300 |
| Silva et al., 2018 [44] | AB3538P, rabbit polyclonal, Chemicon International | NR | I + E | ≥6 |
| Zhang et al., 2018 [75] | NR, Abcam | NR | I + E | ≥2 |
| Afonso et al., 2019 [65] | AB3538P, rabbit polyclonal, Chemicon International | R.T.U. VECTASTAIN Elite ABC Kit, Vector Laboratories | I + E | ≥3 |
| Chen et al., 2019 [62] | 20139-1-AP, rabbit polyclonal, ProteinTech Group Inc. | NR | I + E | ≥2 |
| Sukeda et al., 2019 [47] | sc-365501 Mouse monoclonal, Santa Cruz Biotechnology | Ventana BenchMark XT (Ventana Medical Systems Inc.) | I + E | ≥1 |
| Zheng et al., 2019 [63] | 20139-1-AP, rabbit polyclonal, ProteinTech Group Inc. | NR | I × E | ≥9 |
| Dell’Anno et al., 2020 [52] | AB3538P, rabbit polyclonal, Chemicon International | Dako EnVisionTM FLEX Target Retrieval | I | NR |
| Bonatelli et al., 2021 [55] | AB3538P, rabbit polyclonal, Chemicon International | Ultra Vision ONE Detection System: HRP Polymer, Lab Vision Corp. | I + E | ≥3 |
| Eskuri et al., 2021 [66] | NR, Santa Cruz Biotechnology | Dako EnVisionTM FLEX Target Retrieval | I + E | >150 |
| Leu et al., 2021 [46] | sc-50324, rabbit polyclonal, Santa Cruz Biotechnology | Ventana BenchMark XT (Ventana Medical Systems Inc.) | I × E | 0–300 |
| Tong et al., 2021 [68] | AB238825, rabbit polyclonal, Chemicon International | Dako EnVisionTM FLEX Target Retrieval | I × E | ≥6 |
| Yokoo et al., 2021 [49] | sc-365501, mouse monoclonal, Santa Cruz Biotechnology | NR, Nichirei Biosciences | I + E | ≥4 |
| Hiltunen et al., 2022 [48] | sc-365501, mouse monoclonal, Santa Cruz Biotechnology | Dako EnVisionTM FLEX Target Retrieval | I + E | 0–2 |
| Sun et al., 2022 [73] | AB90582, rabbit polyclonal, Abcam | Dako EnVisionTM FLEX Target Retrieval | I | ≥0.8 |
| Zhao et al., 2022 [64] | 20139-1-AP, rabbit polyclonal, ProteinTech Group Inc. | NR | IOD × E | >120 |
| Reference | Critical Appraisal Skills Programme (CASP) Checklist | Score | Classification | ||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| 1 | 2 | 4 | 5a | 5b | 6a | 6b | 7 | 8 | 9 | 10 | 11 | 12 | |||
| de Oliveira et al., 2012 [51] | ● | ● | ● | ● | ● | ● | ● | ● | ● | ● | ● | ● | ● | 17 | Moderate |
| Choi et al., 2014 [74] | ● | ● | ● | ● | ● | ● | ● | ● | ● | ● | ● | ● | ● | 22 | High |
| Eilertsen et al., 2014 [69] | ● | ● | ● | ● | ● | ● | ● | ● | ● | ● | ● | ● | ● | 22 | High |
| Pértega-Gomes et al., 2014 [57] | ● | ● | ● | ● | ● | ● | ● | ● | ● | ● | ● | ● | ● | 17 | Moderate |
| Pinheiro et al., 2014 [45] | ● | ● | ● | ● | ● | ● | ● | ● | ● | ● | ● | ● | ● | 19 | Moderate |
| Polanski et al., 2014 [35] | ● | ● | ● | ● | ● | ● | ● | ● | ● | ● | ● | ● | ● | 21 | High |
| Zhao et al., 2014 [30] | ● | ● | ● | ● | ● | ● | ● | ● | ● | ● | ● | ● | ● | 12 | Low |
| Afonso et al., 2015 [27] | ● | ● | ● | ● | ● | ● | ● | ● | ● | ● | ● | ● | ● | 21 | High |
| Andersen et al., 2015 [56] | ● | ● | ● | ● | ● | ● | ● | ● | ● | ● | ● | ● | ● | 19 | Moderate |
| Kim et al., 2015 [60] | ● | ● | ● | ● | ● | ● | ● | ● | ● | ● | ● | ● | ● | 22 | High |
| Afonso et al., 2016 [76] | ● | ● | ● | ● | ● | ● | ● | ● | ● | ● | ● | ● | ● | 22 | High |
| Martins et al., 2016 [59] | ● | ● | ● | ● | ● | ● | ● | ● | ● | ● | ● | ● | ● | 23 | High |
| Pinheiro et al., 2016 [41] | ● | ● | ● | ● | ● | ● | ● | ● | ● | ● | ● | ● | ● | 21 | High |
| Pinheiro et al., 2016 [50] | ● | ● | ● | ● | ● | ● | ● | ● | ● | ● | ● | ● | ● | 21 | High |
| Sousa-Simões et al., 2016 [42] | ● | ● | ● | ● | ● | ● | ● | ● | ● | ● | ● | ● | ● | 19 | Moderate |
| Giatromanolaki et al., 2017 [70] | ● | ● | ● | ● | ● | ● | ● | ● | ● | ● | ● | ● | ● | 18 | Moderate |
| Johnson et al., 2017 [71] | ● | ● | ● | ● | ● | ● | ● | ● | ● | ● | ● | ● | ● | 21 | High |
| Latif et al., 2017 [43] | ● | ● | ● | ● | ● | ● | ● | ● | ● | ● | ● | ● | ● | 23 | High |
| Mikkilineni et al., 2017 [53] | ● | ● | ● | ● | ● | ● | ● | ● | ● | ● | ● | ● | ● | 15 | Moderate |
| Sáenz-de-Santa-María et al., 2017 [54] | ● | ● | ● | ● | ● | ● | ● | ● | ● | ● | ● | ● | ● | 18 | Moderate |
| Wang et al., 2017 [67] | ● | ● | ● | ● | ● | ● | ● | ● | ● | ● | ● | ● | ● | 14 | Moderate |
| Cao et al., 2018 [61] | ● | ● | ● | ● | ● | ● | ● | ● | ● | ● | ● | ● | ● | 20 | Moderate |
| Li et al., 2018 [72] | ● | ● | ● | ● | ● | ● | ● | ● | ● | ● | ● | ● | ● | 21 | High |
| Roseweir et al., 2018 [58] | ● | ● | ● | ● | ● | ● | ● | ● | ● | ● | ● | ● | ● | 21 | High |
| Silva et al., 2018 [44] | ● | ● | ● | ● | ● | ● | ● | ● | ● | ● | ● | ● | ● | 21 | High |
| Zhang et al., 2018 [75] | ● | ● | ● | ● | ● | ● | ● | ● | ● | ● | ● | ● | ● | 22 | High |
| Afonso et al., 2019 [65] | ● | ● | ● | ● | ● | ● | ● | ● | ● | ● | ● | ● | ● | 21 | High |
| Chen et al., 2019 [62] | ● | ● | ● | ● | ● | ● | ● | ● | ● | ● | ● | ● | ● | 22 | High |
| Sukeda et al., 2019 [47] | ● | ● | ● | ● | ● | ● | ● | ● | ● | ● | ● | ● | ● | 18 | Moderate |
| Zheng et al., 2019 [63] | ● | ● | ● | ● | ● | ● | ● | ● | ● | ● | ● | ● | ● | 20 | Moderate |
| Dell’Anno et al., 2020 [52] | ● | ● | ● | ● | ● | ● | ● | ● | ● | ● | ● | ● | ● | 12 | Low |
| Bonatelli et al., 2021 [55] | ● | ● | ● | ● | ● | ● | ● | ● | ● | ● | ● | ● | ● | 22 | High |
| Eskuri et al., 2021 [66] | ● | ● | ● | ● | ● | ● | ● | ● | ● | ● | ● | ● | ● | 22 | High |
| Leu et al., 2021 [46] | ● | ● | ● | ● | ● | ● | ● | ● | ● | ● | ● | ● | ● | 21 | High |
| Tong et al., 2021 [68] | ● | ● | ● | ● | ● | ● | ● | ● | ● | ● | ● | ● | ● | 21 | High |
| Yokoo et al., 2021 [49] | ● | ● | ● | ● | ● | ● | ● | ● | ● | ● | ● | ● | ● | 19 | Moderate |
| Hiltunen et al., 2022 [48] | ● | ● | ● | ● | ● | ● | ● | ● | ● | ● | ● | ● | ● | 18 | Moderate |
| Sun et al., 2022 [73] | ● | ● | ● | ● | ● | ● | ● | ● | ● | ● | ● | ● | ● | 24 | High |
| Zhao et al., 2022 [64] | ● | ● | ● | ● | ● | ● | ● | ● | ● | ● | ● | ● | ● | 20 | Moderate |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Silva, A.; Cerqueira, M.C.; Rosa, B.; Sobral, C.; Pinto-Ribeiro, F.; Costa, M.F.; Baltazar, F.; Afonso, J. Prognostic Value of Monocarboxylate Transporter 1 Overexpression in Cancer: A Systematic Review. Int. J. Mol. Sci. 2023, 24, 5141. https://doi.org/10.3390/ijms24065141
Silva A, Cerqueira MC, Rosa B, Sobral C, Pinto-Ribeiro F, Costa MF, Baltazar F, Afonso J. Prognostic Value of Monocarboxylate Transporter 1 Overexpression in Cancer: A Systematic Review. International Journal of Molecular Sciences. 2023; 24(6):5141. https://doi.org/10.3390/ijms24065141
Chicago/Turabian StyleSilva, Ana, Mónica Costa Cerqueira, Beatriz Rosa, Catarina Sobral, Filipa Pinto-Ribeiro, Marta Freitas Costa, Fátima Baltazar, and Julieta Afonso. 2023. "Prognostic Value of Monocarboxylate Transporter 1 Overexpression in Cancer: A Systematic Review" International Journal of Molecular Sciences 24, no. 6: 5141. https://doi.org/10.3390/ijms24065141