
HSPA5 Promotes the Proliferation, Metastasis and Regulates Ferroptosis of Bladder Cancer

Abstract
:1. Introduction
2. Results
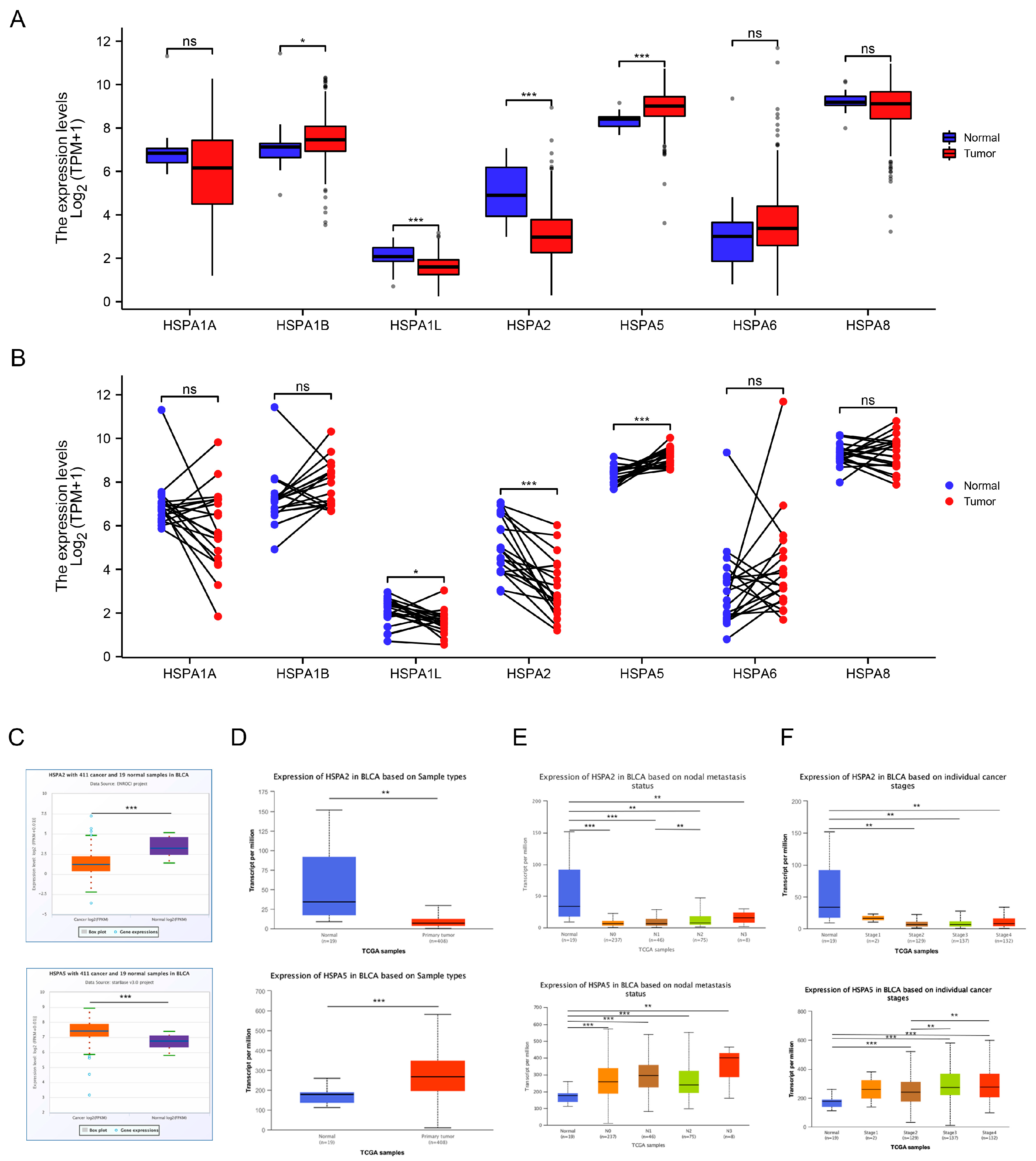
2.1. The Expression Levels of HSP70s in Bladder Cancer
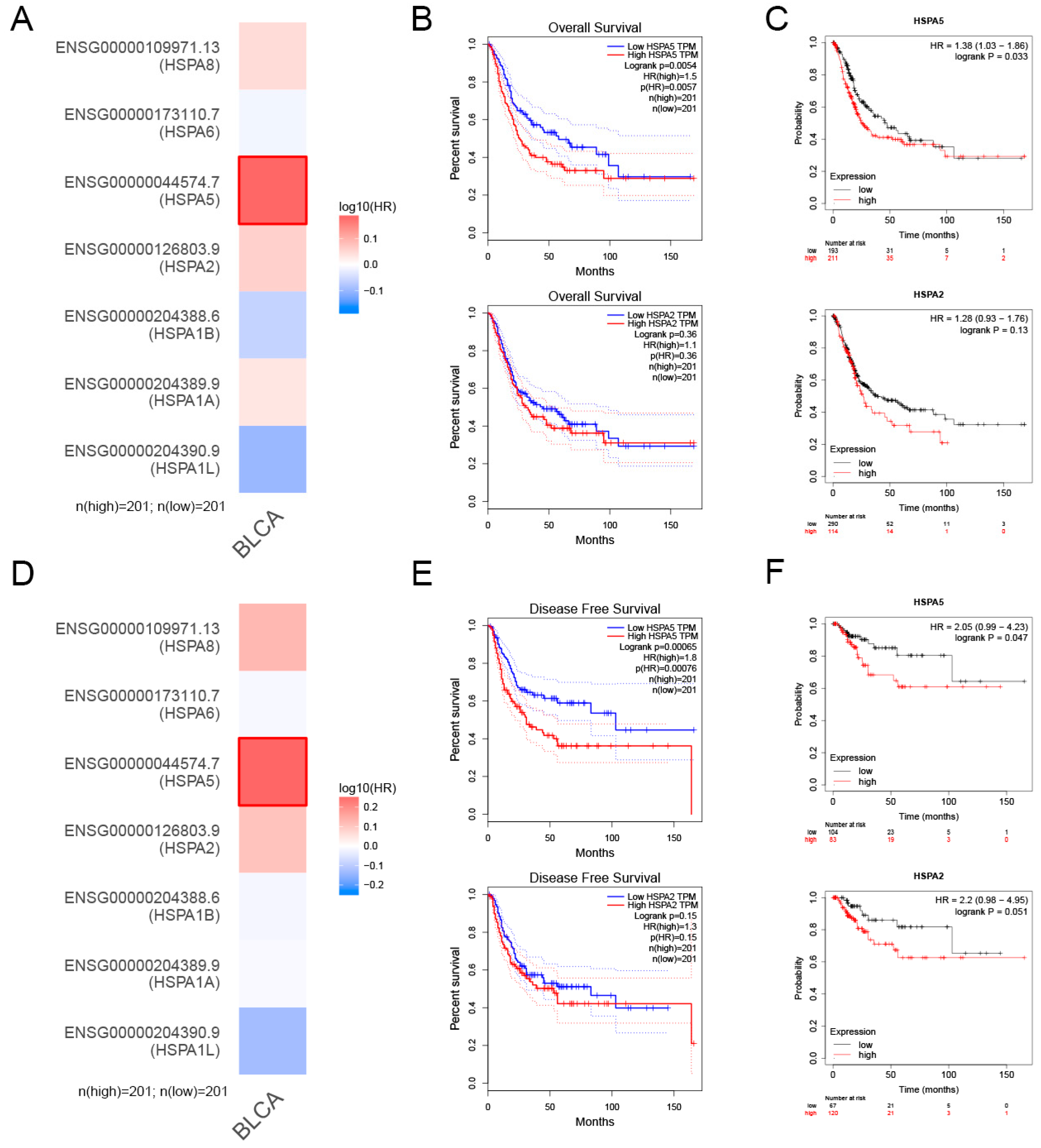
2.2. The Relationship between HSPA2 and HSPA5 Expression and Patient Prognosis
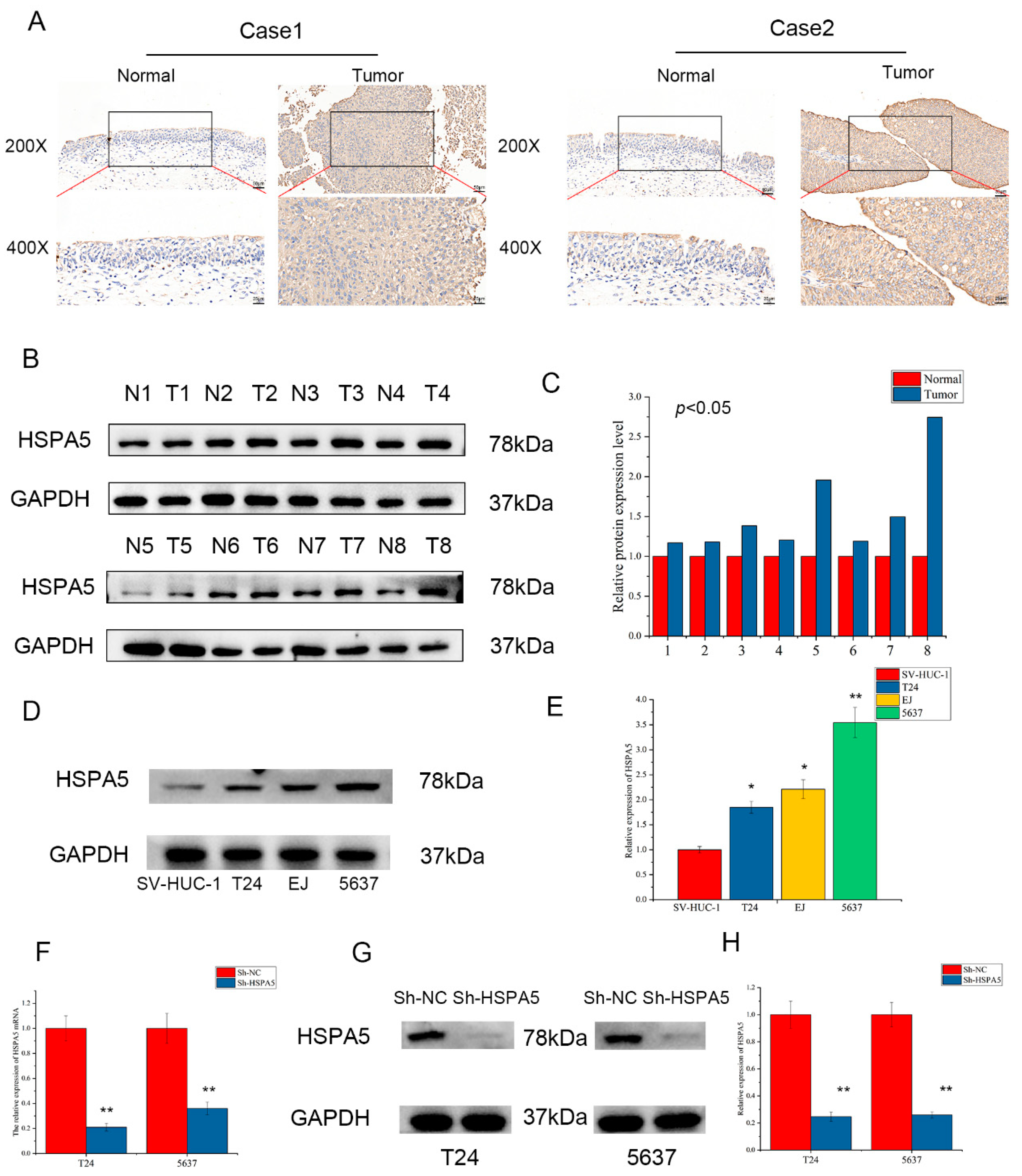
2.3. The Expression of HSPA5 in Clinical Specimens and Cell Lines
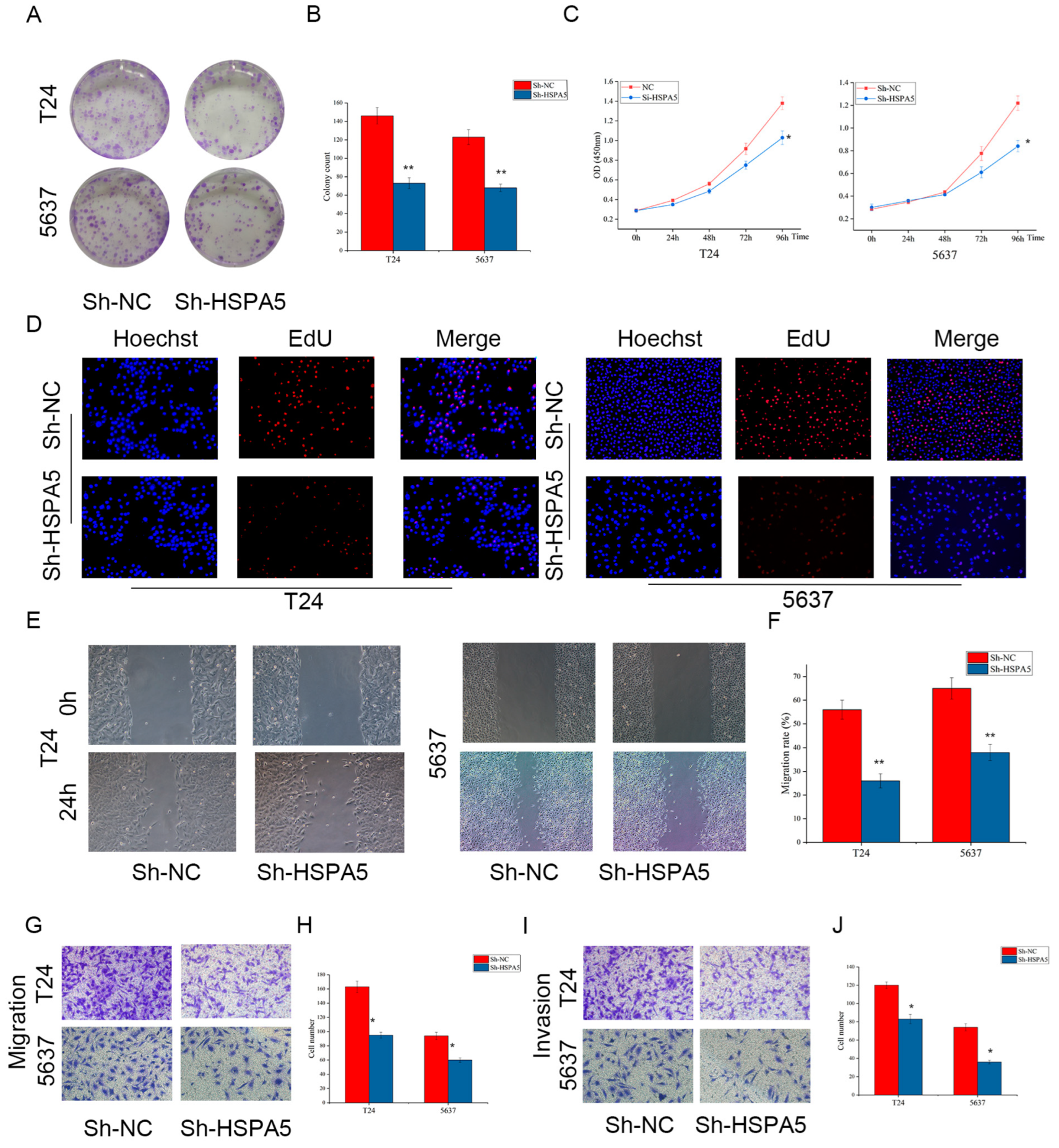
2.4. Downregulating HSPA5 Inhibits BCa Cell Growth and Metastasis In Vitro
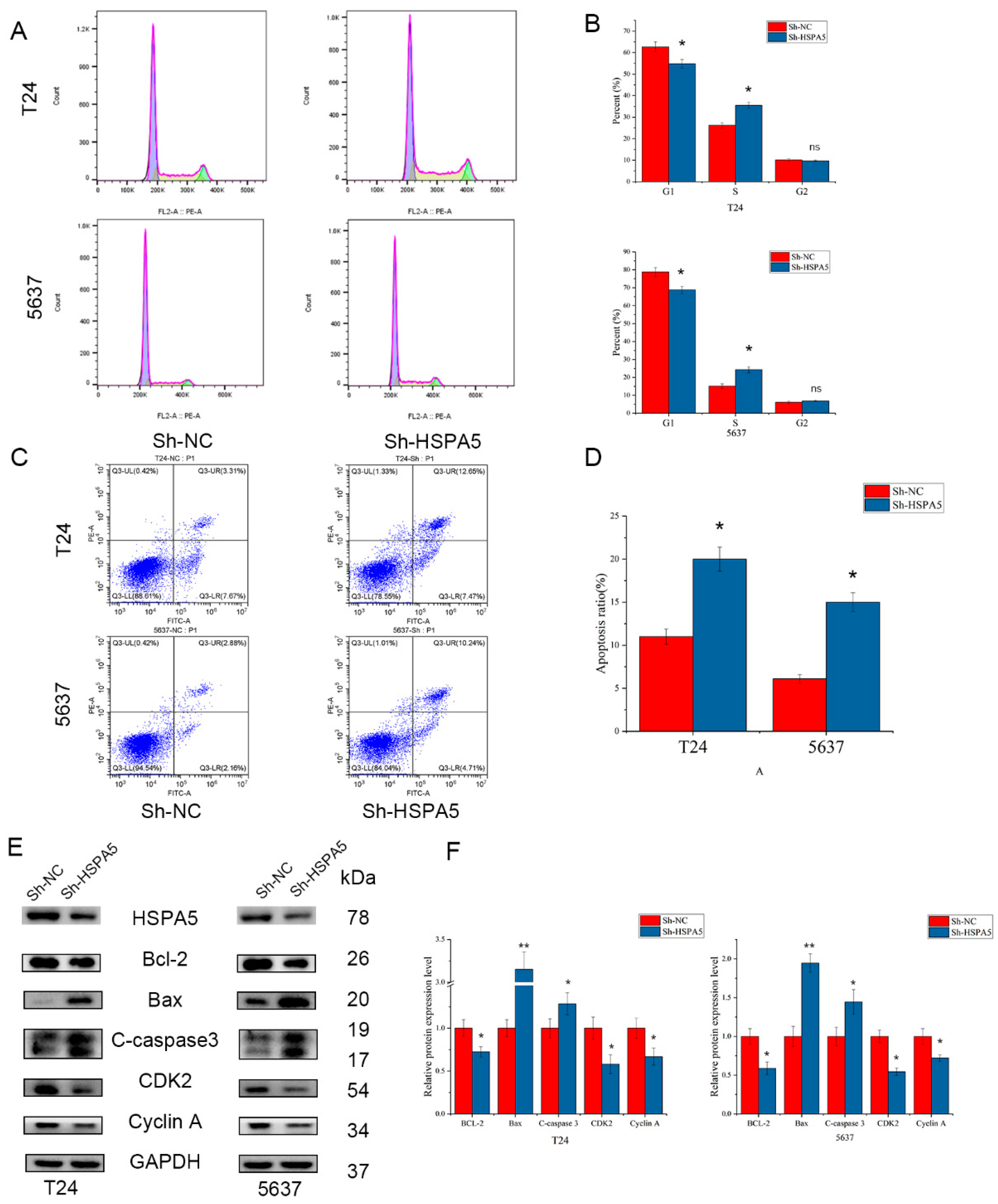
2.5. Effects of HSPA5 on the Cell Cycle and Apoptosis in BCa Cells
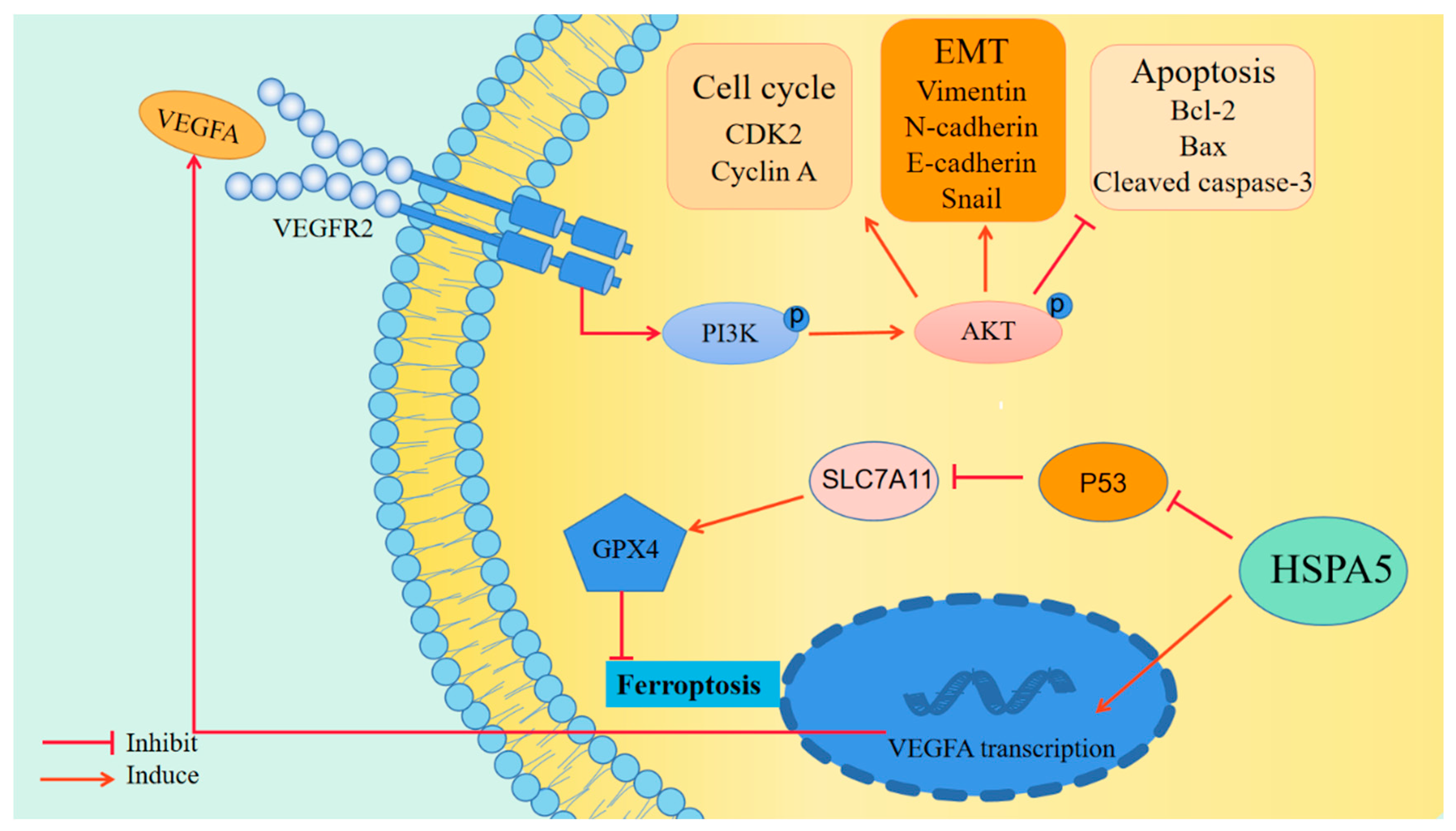
2.6. Inhibiting HSPA5 Suppresses BCa Progression by Downregulating the VEGFA/VEGFR2 Pathway
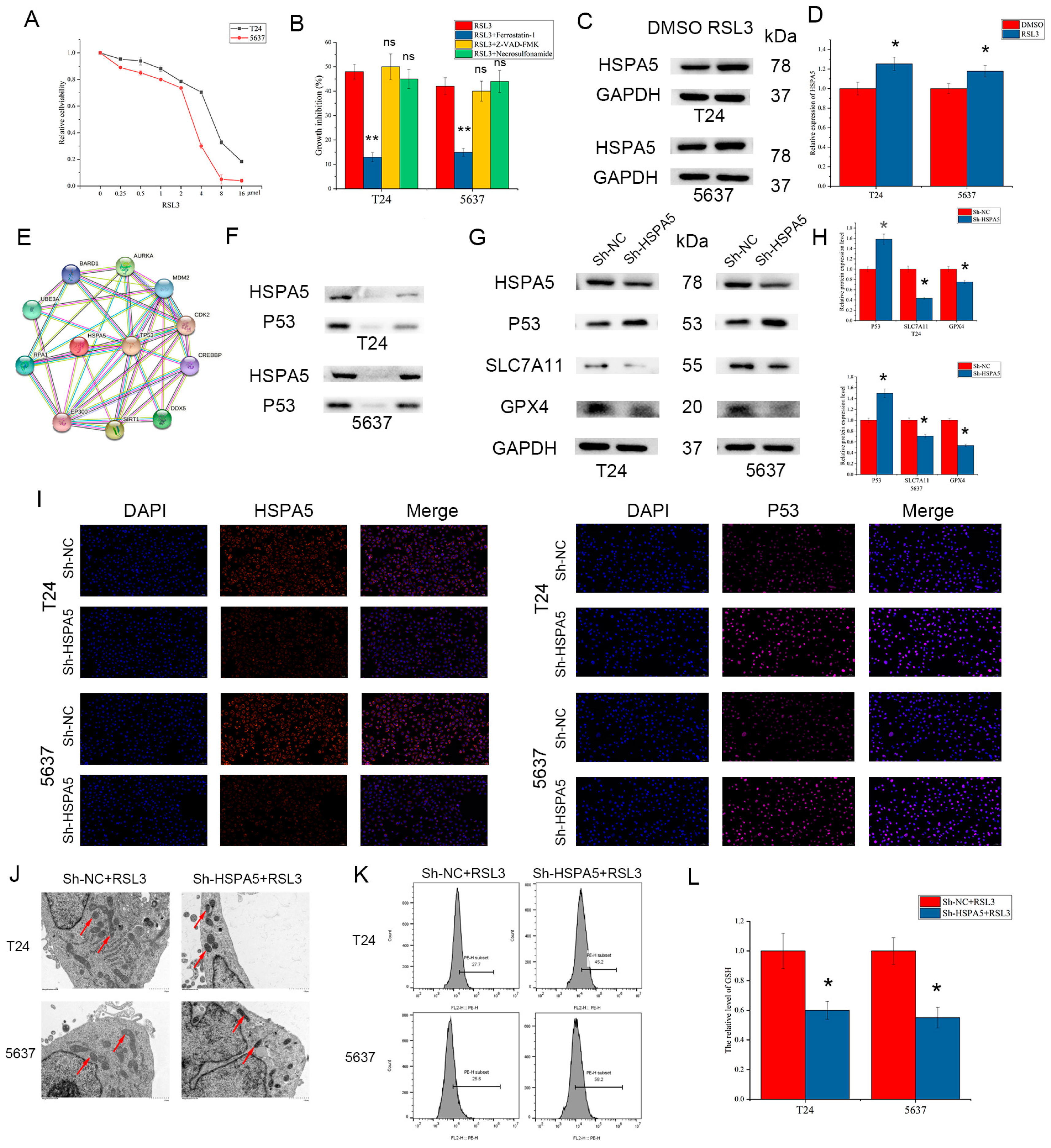
2.7. HSPA5 Inhibition Enhances BCa Cell Ferroptosis
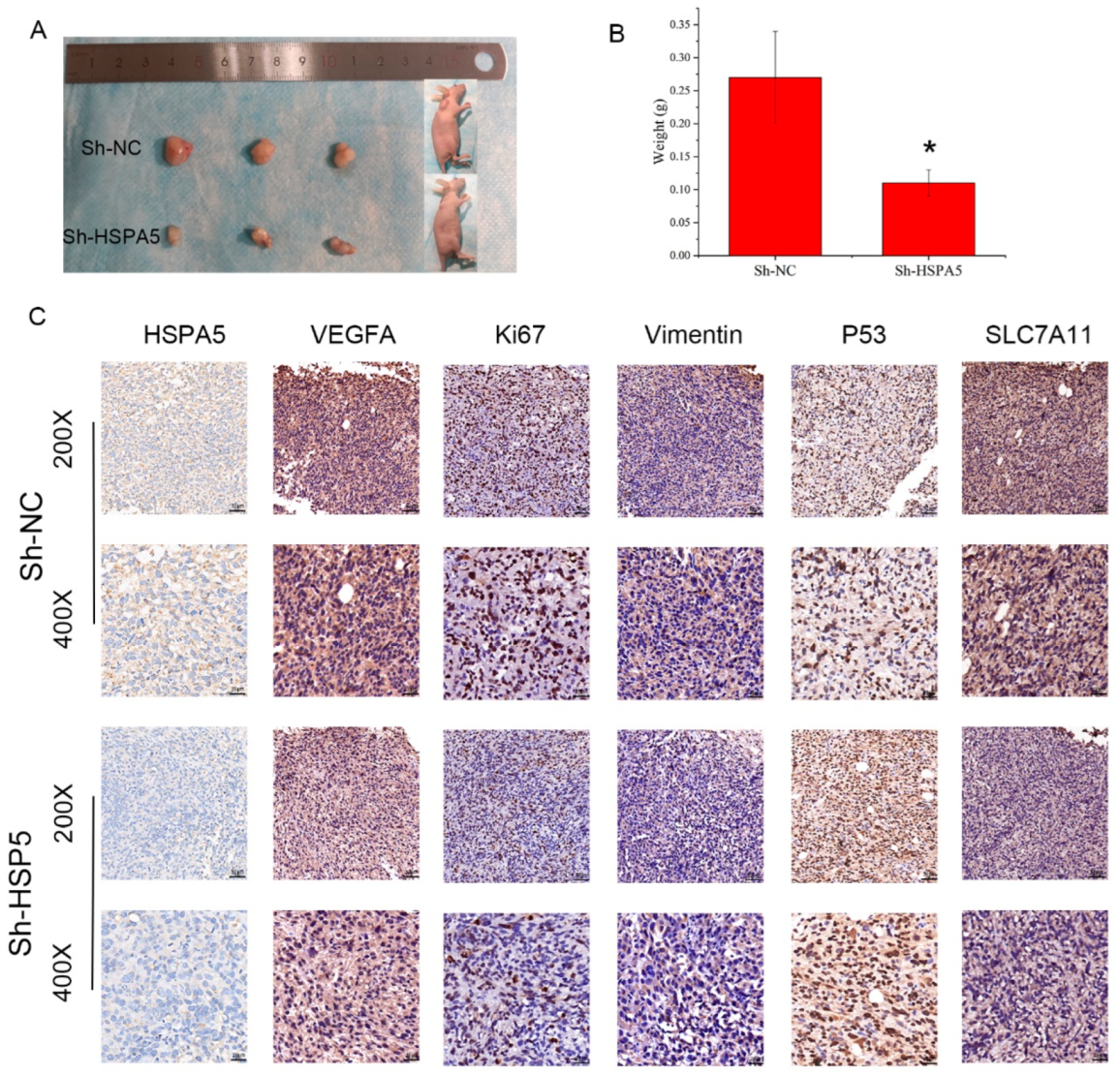
2.8. HSPA5 Knockdown Inhibits Xenograft Growth In Vivo
3. Discussion
4. Materials and Methods
4.1. Patient Samples
4.2. Cell Culture and Treatment
4.3. Cell Transfection Assay
4.4. Quantitative Real-Time PCR
4.5. Western Blotting
4.6. Immunoprecipitation (IP)
4.7. CCK-8 Assay
4.8. Colony Formation Assay
4.9. EdU Assay
4.10. Wound Healing Assay
4.11. Transwell Assay
4.12. Cell Cycle Assay
4.13. Cell Apoptosis Assay
4.14. Reactive Oxygen Species (ROS) Analysis
4.15. Glutathione (GSH) Assay
4.16. Immunofluorescence (IF) and Immunohistochemistry (IHC)
4.17. Xenograft Assay
4.18. Statistical Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Siegel, R.L.; Miller, K.D.; Fuchs, H.E. Cancer statistics, 2022. CA A Cancer J. Clin. 2022, 72, 7–33. [Google Scholar] [CrossRef]
- Lenis; Andrew, T.; Patrick, M.L.; Chamie, K. Bladder cancer: A review. JAMA 2020, 324, 1980–1991. [Google Scholar] [CrossRef]
- Tran, L.; Xiao, J.F.; Agarwal, N.; Duex, J.E. Advances in bladder cancer biology and therapy. Nat. Rev. Cancer 2020, 21, 104–121. [Google Scholar] [CrossRef]
- Minoli, M.; Kiener, M.; Thalmann, G.N.; Julio, M.K.; Seiler, R. Evolution of urothelial bladder cancer in the context of molecular classifications. Int. J. Mol. Sci. 2020, 21, 5670. [Google Scholar] [CrossRef]
- Soheila, B.; Reza, B.; Andre, K.B. Artificial intelligence: A promising frontier in bladder cancer diagnosis and outcome prediction. Crit. Rev. Oncol./Hematol. 2022, 171, 103601. [Google Scholar]
- Patel; Vaibhav, G.; William, K.O.; Matthew, D. Treatment of muscle-invasive and advanced bladder cancer in 2020. CA A Cancer J. Clin. 2020, 70, 404–423. [Google Scholar] [CrossRef]
- Erica, C.O.T.; Benjamin, P.; Jeremy, T.Y.C. The use of artificial intelligence for the diagnosis of bladder cancer: A review and perspectives. Curr. Opin. Urol. 2021, 31, 397–403. [Google Scholar]
- Ibrahim, I.M.; Abdelmalek, D.H.; Elfiky, A.A. GRP78: A cell’s response to stress. Life Sci. 2019, 226, 156–163. [Google Scholar] [CrossRef]
- Jiewen, F.; Chunli, W.; Jiayue, H.; Lianmei, Z.; Ju, Z.; Srinivasa, B.K.; Shiyi, S.; Jiangzhou, P.; Amrish, S.; Junjiang, F. Evaluation and characterization of HSPA5 (GRP78) expression profiles in normal individuals and cancer patients with COVID-19. Int. J. Biol. Sci. 2021, 17, 897–910. [Google Scholar]
- Xiaoli, Y.; Yi, T.; Yulin, X.; Yueyue, G.; Feng, Y.; Xinghua, Z. Endoplasmic reticulum stress-induced exosomal miR-27a-3p promotes immune escape in breast cancer via regulating PD-L1 expression in macrophages. J. Cell Mol. Med. 2020, 24, 9560–9573. [Google Scholar]
- Lager, T.W.; Conner, C.; Keating, C.R.; Warshaw, J.N.; Panopoulos, A.D. Cell surface GRP78 and Dermcidin cooperate to regulate breast cancer cell migration through Wnt signaling. Oncogene 2021, 40, 4050–4059. [Google Scholar] [CrossRef]
- Samanta, S.; Yang, S.; Debnath, B.; Xue, D.; Kuang, Y.; Ramkumar, K.; Lee, A.S.; Ljungman, M.; Neamati, N. The hydroxyquinoline analogue YUM70 inhibits GRP78 to induce ER Stress-Mediated apoptosis in pancreatic cancer. Cancer Res. 2021, 81, 1883–1895. [Google Scholar] [CrossRef]
- Dauer, P.; Sharma, N.S.; Gupta, V.K.; Nomura, A.; Dudeja, V.; Saluja, A.; Banerjee, S. GRP78-mediated antioxidant response and ABC transporter activity confers chemoresistance to pancreatic cancer cells. Mol. Oncol. 2018, 12, 1498–1512. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sun-Yong, K.; Jeong, K.H.; Haeng-Jun, K.; Ho, K.D.; Ho, H.J.; Kwon, B.H.; Keunho, L.; Chul-Ho, K. HSPA5 negatively regulates lysosomal activity through ubiquitination of MUL1 in head and neck cancer. Autophagy 2018, 14, 385–403. [Google Scholar]
- Du, B.; Dai, X.M.; Li, S.; Qi, G.L.; Cao, G.X.; Zhong, Y.; Yin, P.D.; Yang, X.S. MiR-30c regulates cisplatin-induced apoptosis of renal tubular epithelial cells by targeting Bnip3L and Hspa5. Cell Death Dis. 2017, 8, e2987. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gao, S.; Cheng, Q.C.; Hu, Y.G.; Tan, Z.Z.; Chen, L.; Liu, S.W.; Kang, Q.Y.; Wei, T. LncRNA AK148321 alleviates neuroinflammation in LPS-stimulated BV2 microglial cell through regulating microRNA-1199-5p/HSPA5 axis. Life Sci. 2021, 266, 118863. [Google Scholar] [CrossRef]
- Yibing, C.; Yanjun, M.; Xiaofei, Z.; Qian, M.; Yucen, S.; Liwei, Z.; Dandan, W.; Jinliang, X.; Benxin, H.; Haolong, L.; et al. Dihydroartemisinin-induced unfolded protein response feedback attenuates ferroptosis via PERK/ATF4/HSPA5 pathway in glioma cells. J. Exp. Clin. Cancer Res. 2019, 38, 402. [Google Scholar]
- Apte, R.S.; Chen, D.S.; Ferrara, N. VEGF in signaling and disease: Beyond discovery and development. Cell 2019, 176, 1248–1264. [Google Scholar] [CrossRef] [Green Version]
- Ferrara, N.; Adamis, A.P. Ten years of anti-vascular endothelial growth factor therapy. Nat. Rev. Drug Discov. 2016, 15, 385–403. [Google Scholar] [CrossRef] [Green Version]
- Chen, C.; Zhiguo, H.; Xiaoye, M.; Yanmin, S.; Xiangmin, L.; Xiaogang, L.; Mu, Z. The circular RNA 001971/miR-29c-3p axis modulates colorectal cancer growth, metastasis, and angiogenesis through VEGFA. J. Exp. Clin. Cancer Res. 2020, 39, 91. [Google Scholar] [CrossRef]
- Sinem, K.; Veli-Matti, L.; Kari, A. Vascular endothelial growth factor signaling in development and disease. Development 2018, 145, dev151019. [Google Scholar]
- Lijun, L.; Kaiyuan, H.; Chenxi, H.; Yixuan, W.; Shikun, Y.; Panrong, Z.; Lei, W.; Youyou, X.; Yun, Q.; Wen, S.; et al. Autophagy inhibition potentiates the anti-angiogenic property of multikinase inhibitor anlotinib through JAK2/STAT3/VEGFA signaling in non-small cell lung cancer cells. J. Exp. Clin. Cancer Res. 2019, 38, 71. [Google Scholar]
- Dixon, S.J.; Lemberg, K.M.; Lamprecht, M.R.; Skouta, R.; Zaitsev, E.M.; Gleason, C.E.; Patel, D.N.; Bauer, A.J.; Cantley, A.M.; Yang, W.S.; et al. Ferroptosis: An Iron-Dependent form of nonapoptotic cell death. Cell 2012, 149, 1060–1072. [Google Scholar] [CrossRef] [Green Version]
- Xin, C.; Rui, K.; Guido, K.; Daolin, T. Broadening horizons: The role of ferroptosis in cancer. Nat. Rev. Clin. Oncol. 2021, 18, 280–296. [Google Scholar]
- Jiang, X.; Stockwell, B.R.; Conrad, M. Ferroptosis: Mechanisms, biology and role in disease. Nat. Rev. Mol. Cell Bio. 2021, 22, 266–282. [Google Scholar]
- Hirschhorn, T.; Stockwell, B.R. The development of the concept of ferroptosis. Free Radic. Biol. Med. 2018, 133, 130–143. [Google Scholar]
- Jie, L.; Feng, C.; He-Liang, Y.; Zi-Jian, H.; Zhi-Tao, L.; Ning, M.; Bei, S.; Gang, W. Ferroptosis: Past, present and future. Cell Death Dis. 2020, 11, 88. [Google Scholar]
- Hassannia, B.; Vandenabeele, P.; Berghe, T.V. Targeting ferroptosis to iron out cancer. Cancer Cell 2019, 35, 830–849. [Google Scholar] [CrossRef]
- Dingshan, L.; Yongsheng, L. The interaction between ferroptosis and lipid metabolism in cancer. Signal Transduct. Target. Ther. 2020, 5, 108. [Google Scholar]
- Facchini, G.; Cavaliere, C.; Romis, L.; Mordente, S.; Facchini, S.; Iovane, G.; Capasso, M.; D’Errico, D.; Liguori, C.; Formato, R.; et al. Advanced/metastatic bladder cancer: Current status and future directions. Eur. Rev. Med. Pharmacol. 2020, 24, 11536–11552. [Google Scholar]
- Laukhtina, E.; Hassler, M.R.; Pradere, B.; Yanagisawa, T.; Quhal, F.; Rajwa, P.; Sari, M.R.; Konig, F.; Pallauf, M.; Kawada, T.; et al. Circulating tumour DNA is a strong predictor of outcomes in patients treated with systemic therapy for urothelial carcinoma. Eur. Urol. Focus 2022, 8, 1683–1686. [Google Scholar] [CrossRef]
- de Kruijff, I.E.; Beije, N.; Martens, J.; de Wit, R.; Boormans, J.L.; Sleijfer, S. Liquid biopsies to select patients for perioperative chemotherapy in muscle-invasive bladder cancer: A systematic review. Eur. Urol. Oncol. 2021, 4, 204–214. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jin, R.; Zhao, A.; Han, S.; Zhang, D.; Sun, H.; Li, M.; Su, D.; Liang, X. The interaction of S100A16 and GRP78 actives endoplasmic reticulum stress-mediated through the IRE1alpha/XBP1 pathway in renal tubulointerstitial fibrosis. Cell Death Dis. 2021, 12, 942. [Google Scholar] [CrossRef]
- Aran, G.; Sanjurjo, L.; Barcena, C.; Simon-Coma, M.; Téllez, É.; Vázquez-Vitali, M.; Garrido, M.; Guerra, L.; Díaz, E.; Ojanguren, I.; et al. CD5L is upregulated in hepatocellular carcinoma and promotes liver cancer cell proliferation and antiapoptotic responses by binding to HSPA5 (GRP78). FASEB J. Off. Publ. Fed. Am. Soc. Exp. Biol. 2018, 32, 3878–3891. [Google Scholar] [CrossRef] [Green Version]
- Ferrara, N.; Mass, R.D.; Campa, C.; Kim, R. Targeting VEGF-A to treat cancer and age-related macular degeneration. Annu. Rev. Med. 2007, 58, 491–504. [Google Scholar] [CrossRef]
- Lu, Y.; Qin, T.; Li, J.; Wang, L.; Zhang, Q.; Jiang, Z.; Mao, J. MicroRNA-140-5p inhibits invasion and angiogenesis through targeting VEGF-A in breast cancer. Cancer Gene Ther. 2017, 24, 386–392. [Google Scholar] [CrossRef] [Green Version]
- Feng, H.; Jin, Z.; Liang, J.; Zhao, Q.; Zhan, L.; Yang, Z.; Yan, J.; Kuang, J.; Cheng, X.; Qiu, W. FOXK2 transcriptionally activating VEGFA induces apatinib resistance in anaplastic thyroid cancer through VEGFA/VEGFR1 pathway. Oncogene 2021, 40, 6115–6129. [Google Scholar] [CrossRef]
- Keklikoglou, I.; Kadioglu, E.; Bissinger, S.; Langlois, B.; Bellotti, A.; Orend, G.; Ries, C.H.; De Palma, M. Periostin limits tumor response to VEGFA inhibition. Cell Rep. 2018, 22, 2530–2540. [Google Scholar] [CrossRef] [Green Version]
- Zhu, S.; Zhang, Q.; Sun, X.; Zeh, H.R.; Lotze, M.T.; Kang, R.; Tang, D. HSPA5 regulates ferroptotic cell death in cancer cells. Cancer Res. 2017, 77, 2064–2077. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jiang, L.; Kon, N.; Li, T.; Wang, S.J.; Su, T.; Hibshoosh, H.; Baer, R.; Gu, W. Ferroptosis as a p53-mediated activity during tumour suppression. Nature 2015, 520, 57–62. [Google Scholar] [CrossRef] [Green Version]
- Wang, Q.; Liu, Z.; Zhai, G.; Yu, X.; Ke, S.; Shao, H.; Guo, J. Overexpression of GATA5 inhibits prostate cancer progression by regulating PLAGL2 via the FAK/PI3K/AKT pathway. Cancers 2022, 14, 2074. [Google Scholar] [CrossRef] [PubMed]











{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Forward Primer 5′–3′ | Reverse Primer 5′–3′ | |
|---|---|---|
| GAPDH | TGACTTCAACAGCGACACCCA | CACCCTGTTGCTGTAGCCAAA |
| HSPA5 | GGGGTGAGGGGAGGGAGTATTTG | GCTGGGAGACTGAGGTGGAAGG |
| VEGFA | CTTCGCTTACTCTCACCTGCTTCTG | GCTGTCATGGGCTGCTTCTTCC |
| Dilution | s | ||||
|---|---|---|---|---|---|
| Antibody | Specificity | WB | IHC | IF | Supplier |
| HSPA5 | Rabbit | 1:2000 | 1:100 | 1:100 | Proteintech |
| VEGFA | Rabbit | 1:500 | 1:100 | 1:100 | Wanlebio |
| VEGFR2 | Rabbit | 1:500 | - | - | ABclonal |
| p-VEGFR2 | Rabbit | 1:500 | - | - | ABclonal |
| PI3K | Mouse | 1:1000 | - | - | Proteintech |
| p-PI3K | Rabbit | 1:1000 | - | - | ABclonal |
| AKT | Rabbit | 1:1000 | - | - | ABclonal |
| p-AKT | Rabbit | 1:1000 | - | - | ABclonal |
| E-Cadherin | Rabbit | 1:1000 | - | - | Proteintech |
| N-Cadherin | Rabbit | 1:1000 | - | - | Proteintech |
| Vimentin | Rabbit | 1:2000 | 1:100 | 1:100 | Proteintech |
| CDK2 | Rabbit | 1:1000 | 1:100 | - | Wanlebio |
| Cyclin A | Rabbit | 1:500 | - | - | Wanlebio |
| P53 | Rabbit | 1:1000 | 1:100 | 1:100 | Proteintech |
| SLC7A11 | Rabbit | 1:1000 | 1:100 | - | Proteintech |
| GPX4 | Mouse | 1:1000 | - | - | Proteintech |
| Ki-67 | Mouse | - | 1:100 | - | Proteintech |
| GAPDH | Rabbit | 1:1000 | - | - | Servicebio |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Wang, Q.; Ke, S.; Liu, Z.; Shao, H.; He, M.; Guo, J. HSPA5 Promotes the Proliferation, Metastasis and Regulates Ferroptosis of Bladder Cancer. Int. J. Mol. Sci. 2023, 24, 5144. https://doi.org/10.3390/ijms24065144
Wang Q, Ke S, Liu Z, Shao H, He M, Guo J. HSPA5 Promotes the Proliferation, Metastasis and Regulates Ferroptosis of Bladder Cancer. International Journal of Molecular Sciences. 2023; 24(6):5144. https://doi.org/10.3390/ijms24065144
Chicago/Turabian StyleWang, Qinghua, Shuai Ke, Zelin Liu, Haoren Shao, Mu He, and Jia Guo. 2023. "HSPA5 Promotes the Proliferation, Metastasis and Regulates Ferroptosis of Bladder Cancer" International Journal of Molecular Sciences 24, no. 6: 5144. https://doi.org/10.3390/ijms24065144