
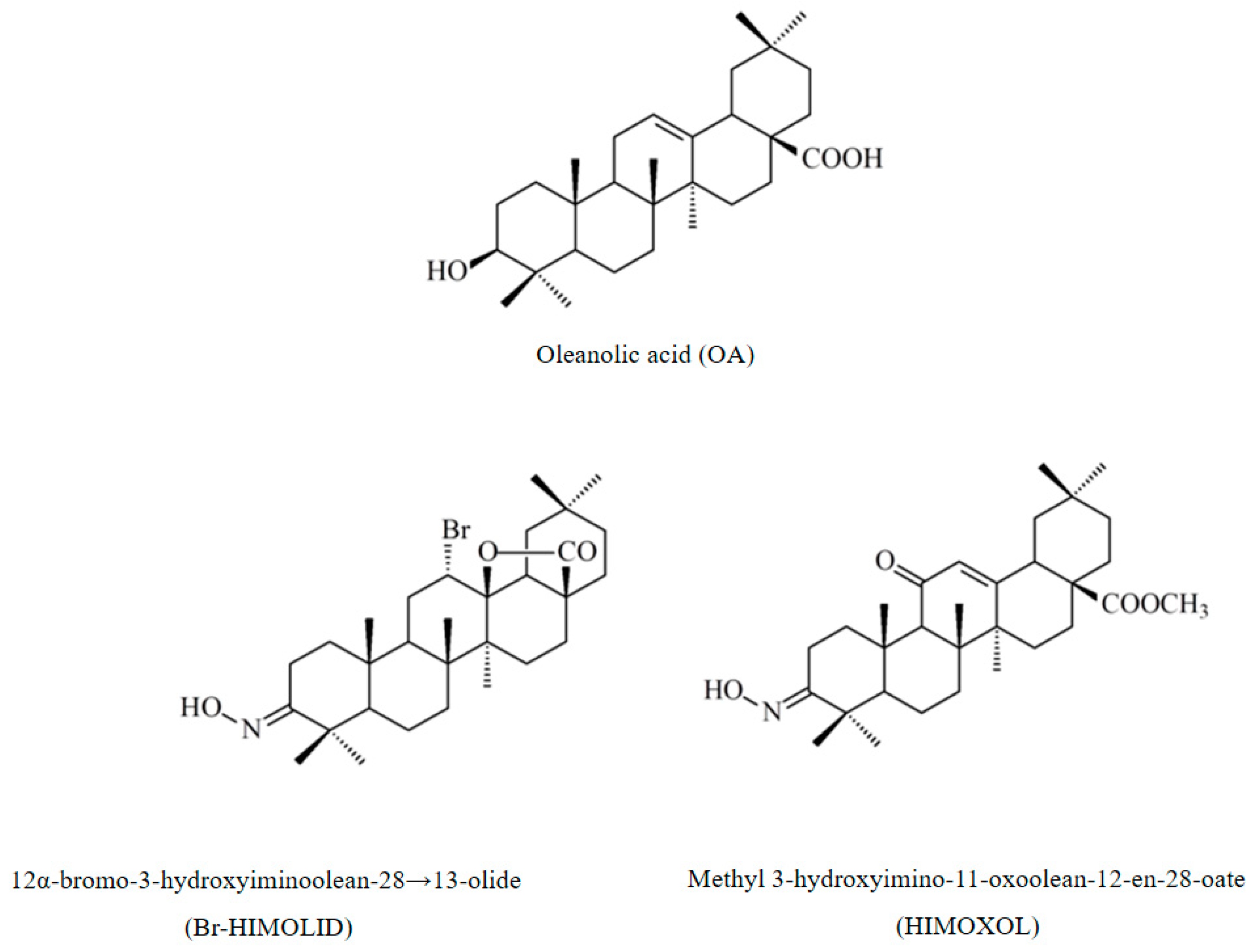
Biological Activity of Oleanolic Acid Derivatives HIMOXOL and Br-HIMOLID in Breast Cancer Cells Is Mediated by ER and EGFR
 ,
,  , and
, and
Abstract
:1. Introduction
2. Results
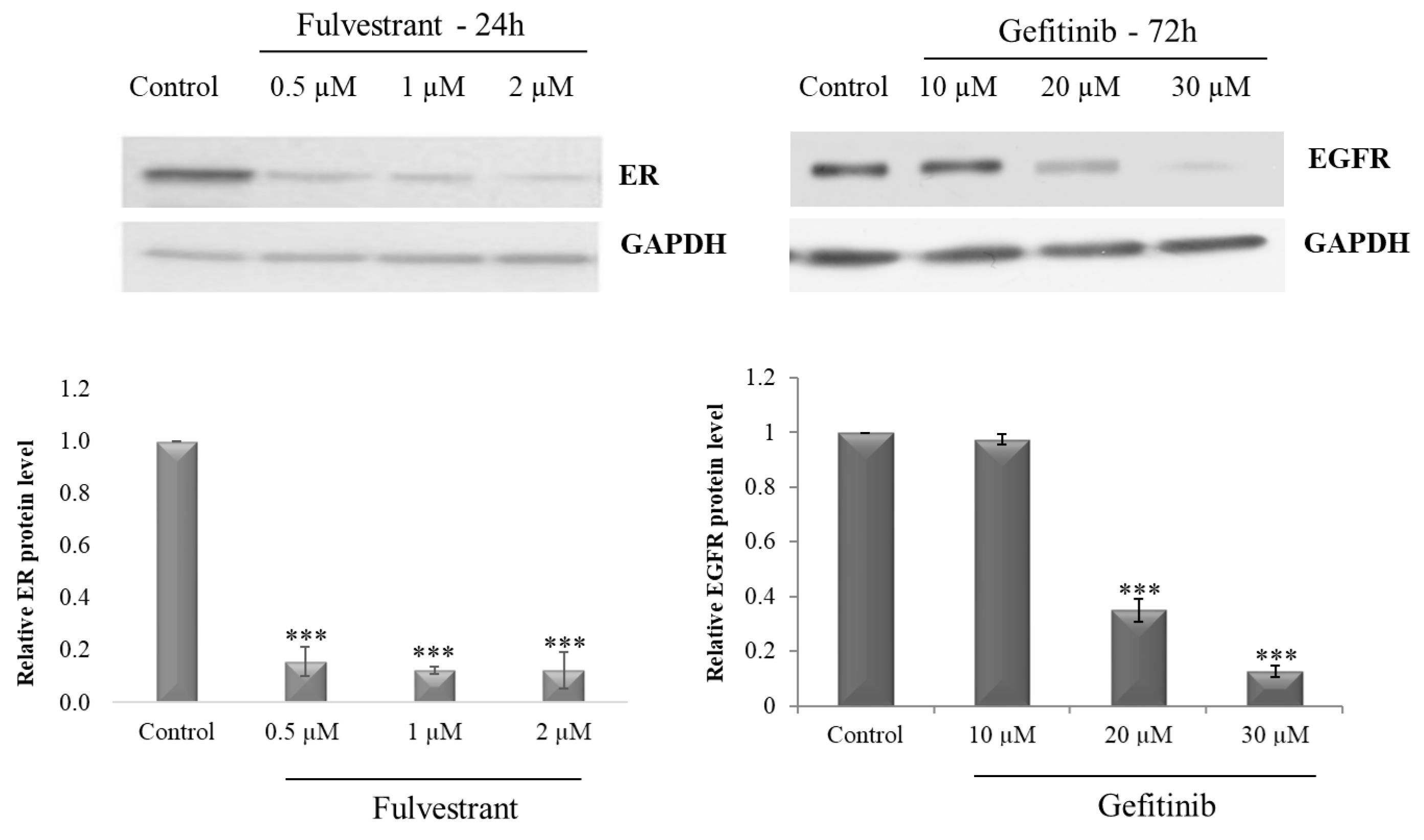
2.1. Optimization of Receptor Blocker Concentration
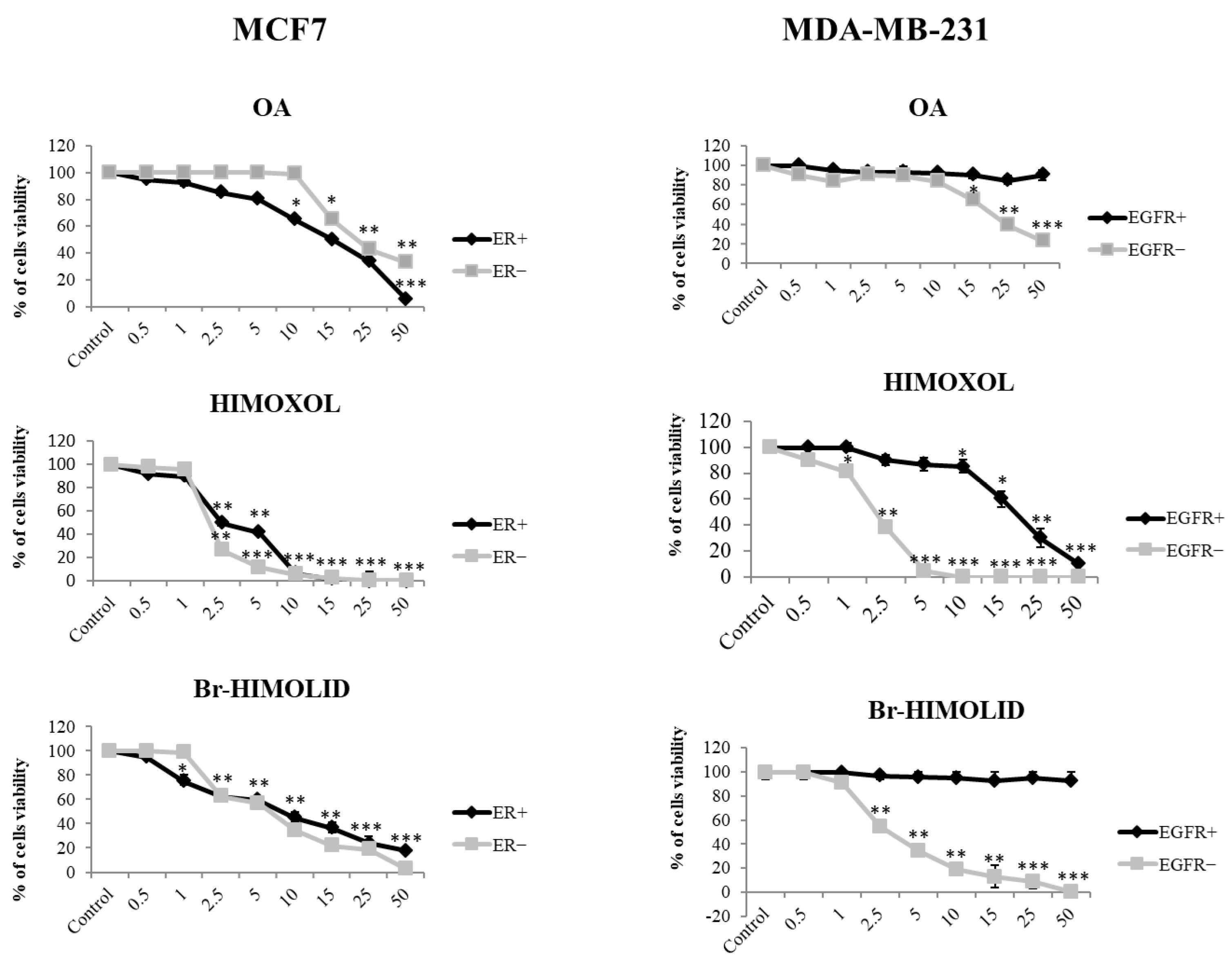
2.2. Contribution of ER and EGFR to the Response of MCF7 and MDA-MB-231 Cells to OA or Its Derivatives
2.3. Contribution of ER and EGFR to the Genotoxic Effect of OA or Its Derivatives in MCF7 and MDA-MB-231 Cells
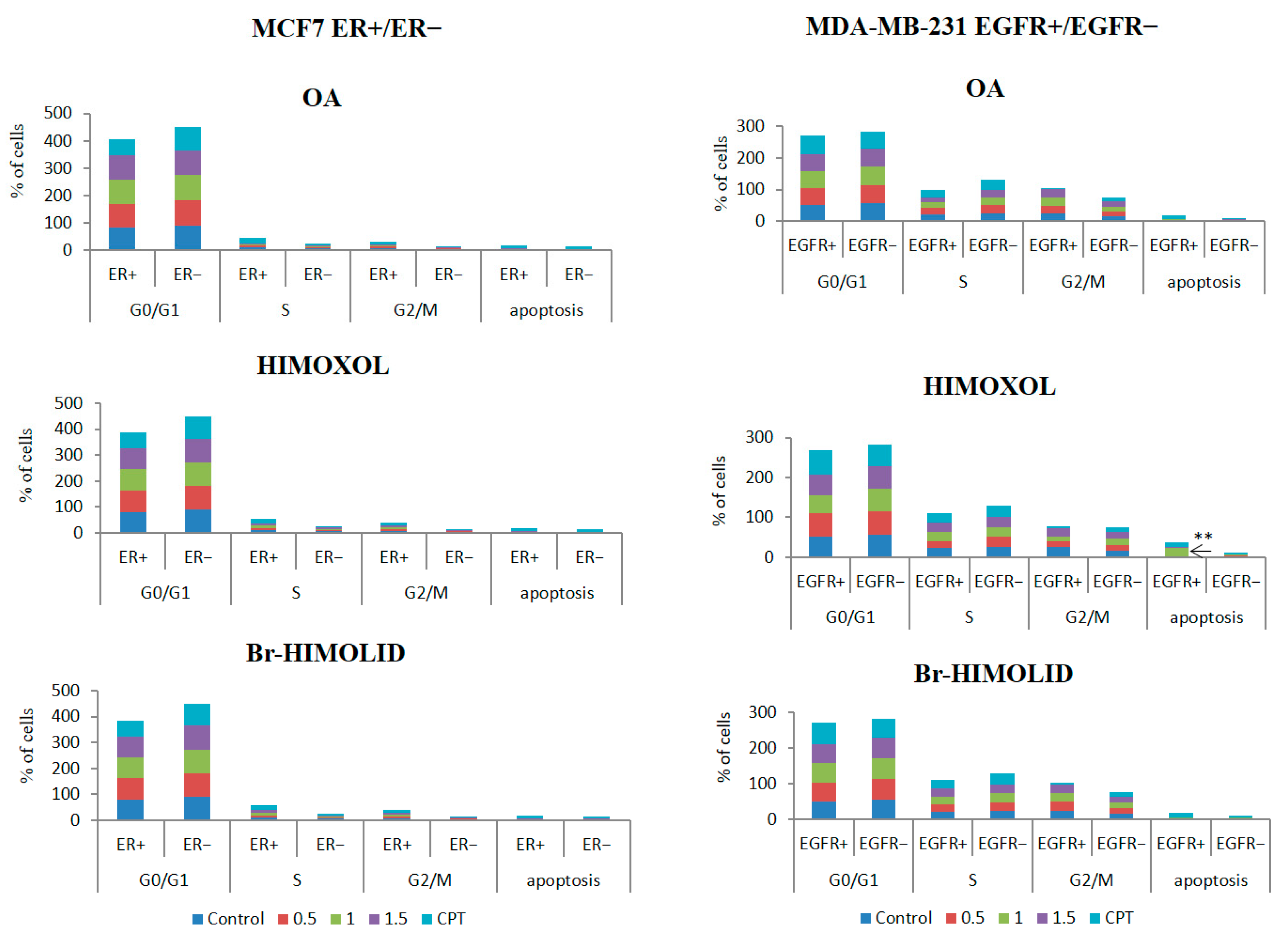
2.4. Contribution of ER and EGFR to the Cell Cycle Distribution in MCF7/ER+/− and MDA-MB-231/EGFR+/− Breast Cancer Cells Exposed to OA or Its Derivatives
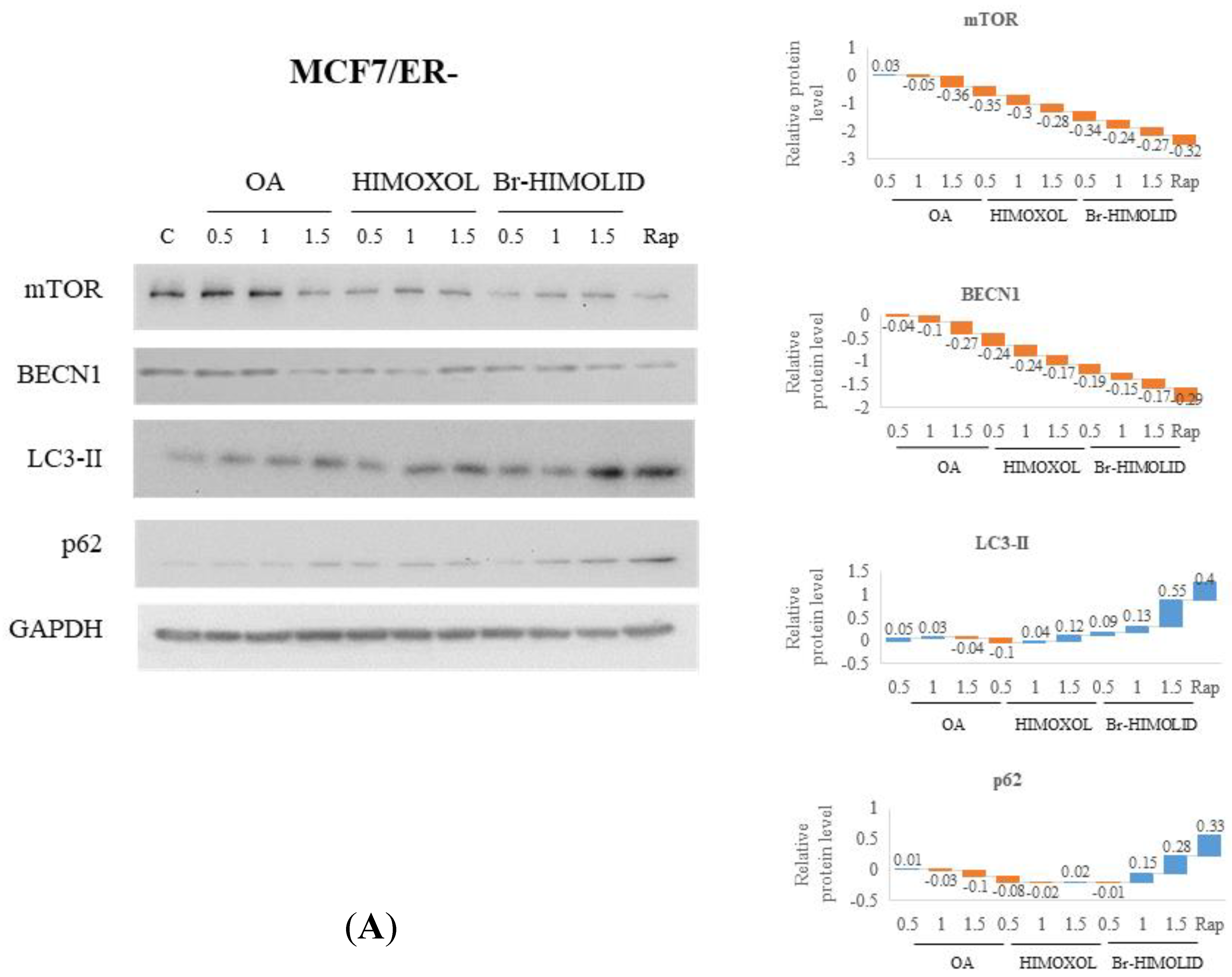
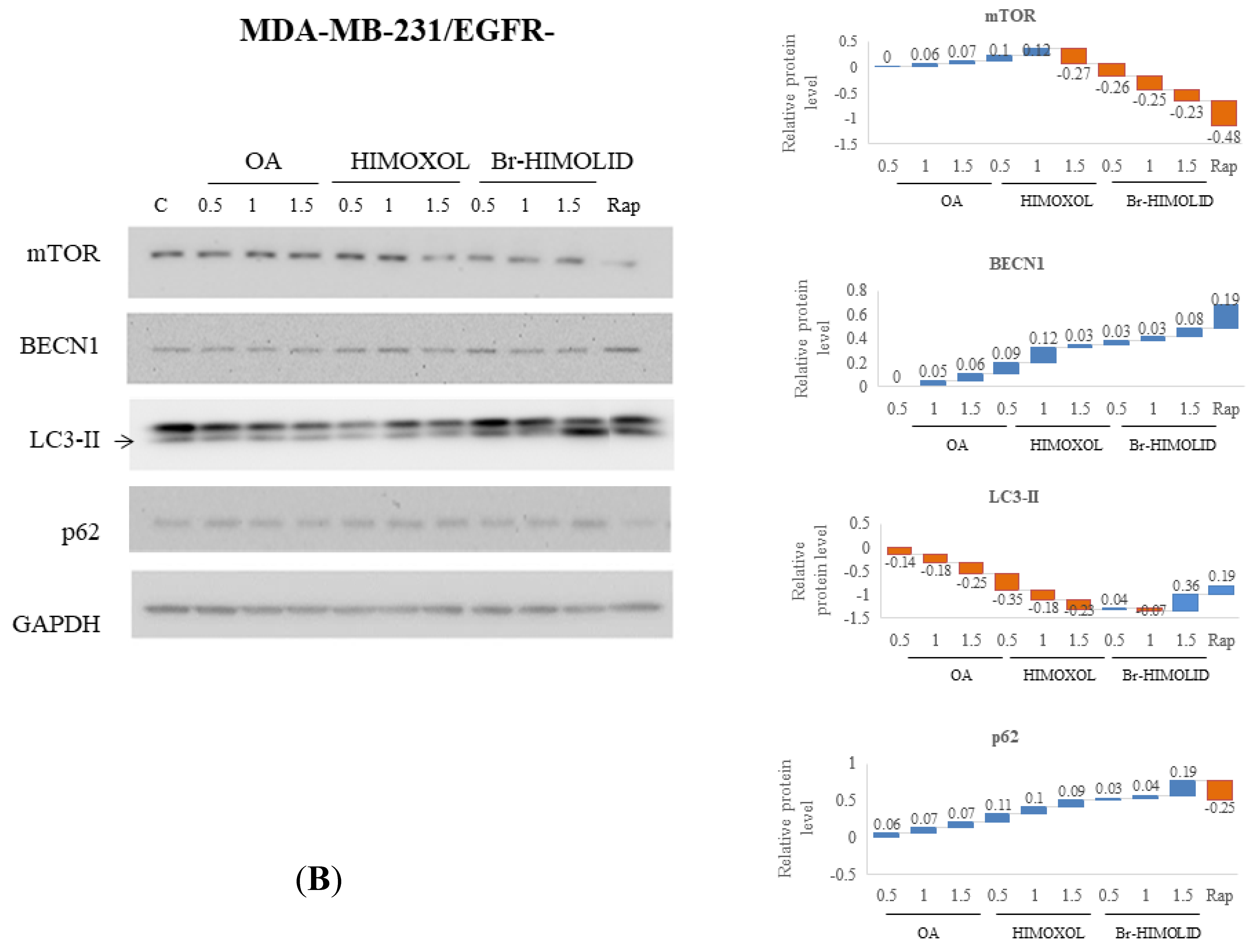
2.5. Evaluation and Verification of Autophagy Pathway
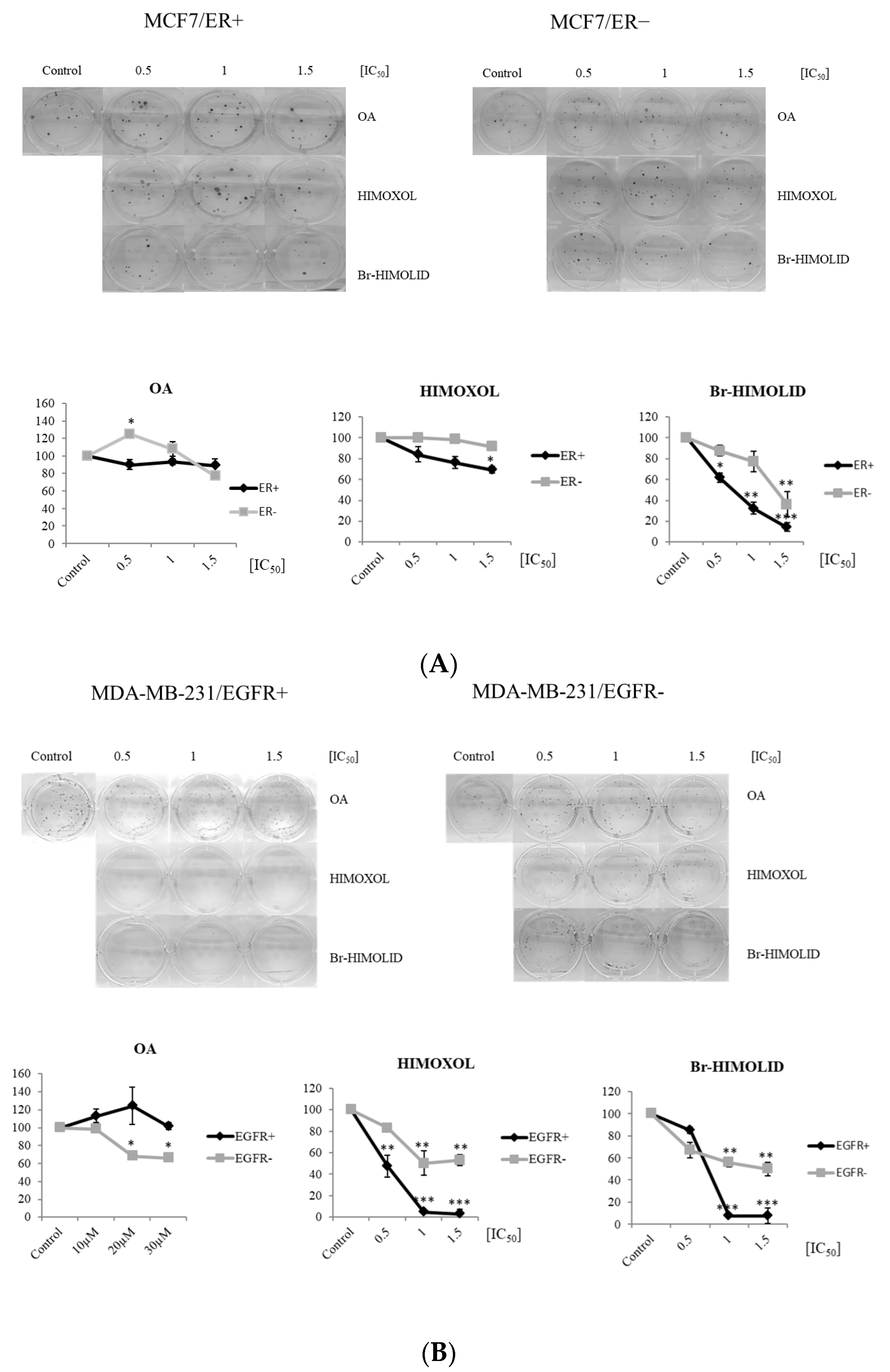
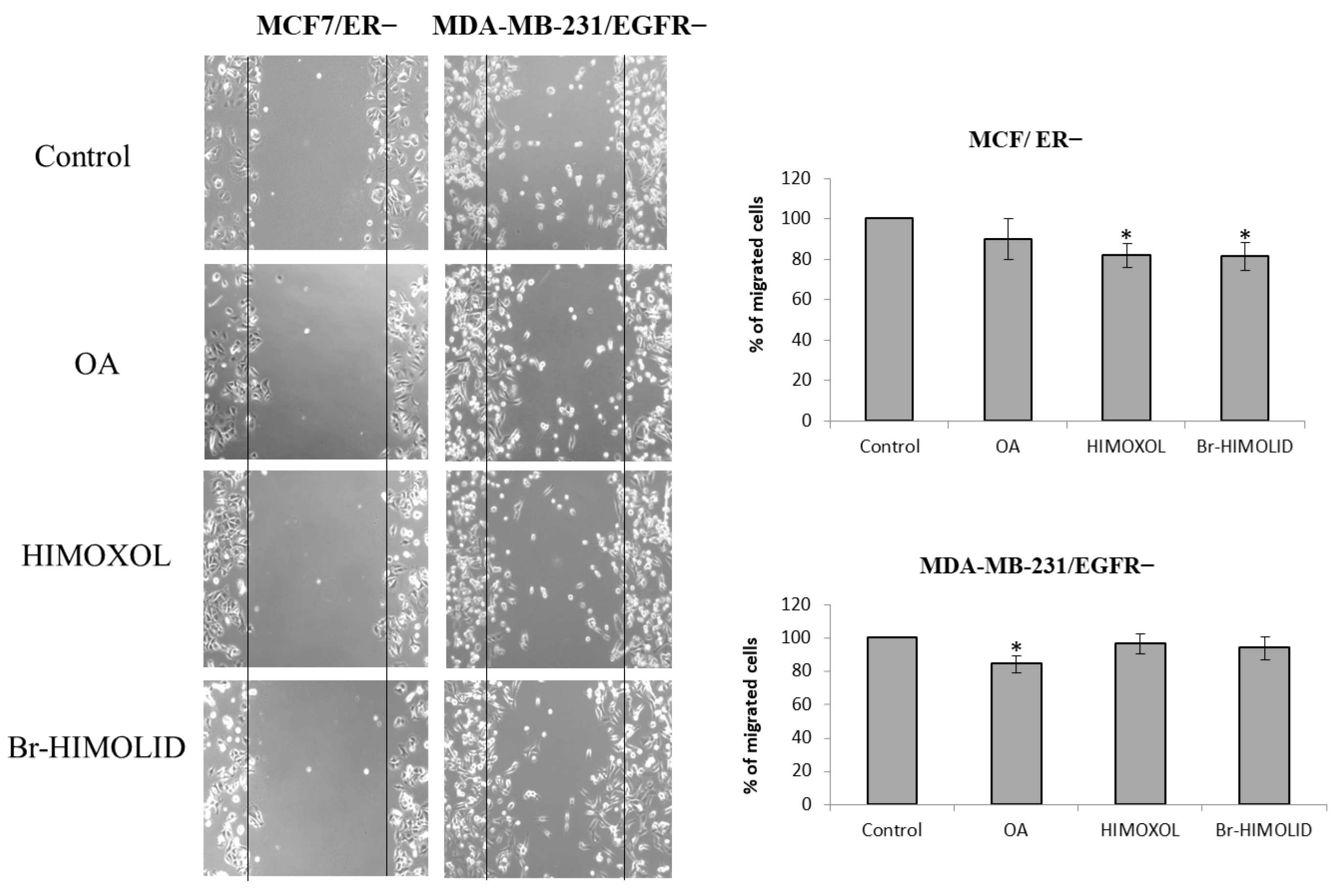
2.6. Contribution of ER and EGFR to Migration Potential of OA and Its Derivatives in Breast Cancer Cells
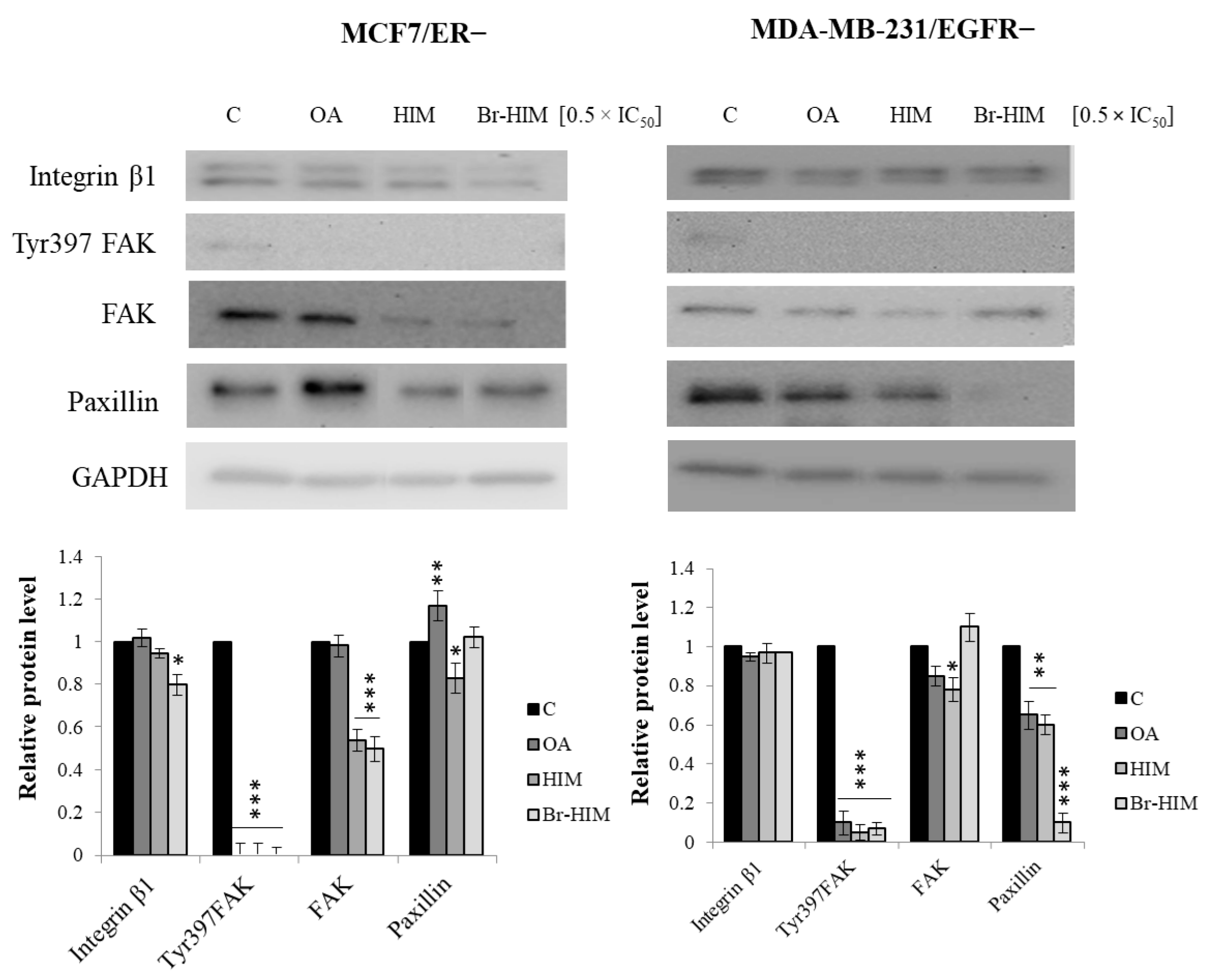
2.7. Contribution of OA and Its Derivatives to the Adhesion and Migration of ER− and EGFR− Deprived Breast Cancer Cells
3. Discussion
3.1. OA Derivatives in Cancer Targeting
3.2. The Mechanistic Aspect of OA and Its Derivatives
4. Materials and Methods
4.1. Compounds and Reagents
4.2. Cell Line and Cell Culture
4.2.1. ER Inhibition with Fulvestrant
4.2.2. EGFR Inhibition with Gefitinib
4.3. Viability Assay
4.4. Colony-Formation Assay
4.5. Cell Cycle Analysis by Flow Cytometry
4.6. Immunodetection
4.7. Wound Healing Assay
4.8. Statistical Analysis
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Available online: https://www.who.int/news-room/fact-sheets/detail/breast-cancer (accessed on 15 January 2023).
- Sun, Y.-S.; Zhao, Z.; Yang, Z.-N.; Xu, F.; Lu, H.-J.; Zhu, Z.-Y.; Shi, W.; Jiang, J.; Yao, P.-P.; Zhu, H.-P. Risk Factors and Preventions of Breast Cancer. Int. J. Biol. Sci. 2017, 13, 1387–1397. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Testa, U.; Castelli, G.; Pelosi, E. Breast Cancer: A Molecularly Heterogenous Disease Needing Subtype-Specific Treatments. Med. Sci. 2020, 8, 18. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Orrantia-Borunda, E.; Anchondo-Nuñez, P.; Acuña-Aguilar, L.E.; Gómez-Valles, F.O.; Ramírez-Valdespino, C.A. Subtypes of Breast Cancer. Exon Publ. 2022, 3, 1–12. [Google Scholar] [CrossRef]
- Cho, N. Molecular subtypes and imaging phenotypes of breast cancer. Ultrasonography 2016, 35, 281–288. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sohail, S.K.; Sarfraz, R.; Imran, M.; Kamran, M.; Qamar, S. Estrogen and Progesterone Receptor Expression in Breast Carcinoma and Its Association With Clinicopathological Variables Among the Pakistani Population. Cureus 2020, 12, e9751. [Google Scholar] [CrossRef]
- Saraiva, D.P.; Cabral, M.G.; Jacinto, A.; Braga, S. How many diseases is triple negative breast cancer: The protagonism of the immune microenvironment. ESMO Open 2017, 2, e000208. [Google Scholar] [CrossRef] [Green Version]
- Carroll, J. EJE PRIZE 2016: Mechanisms of oestrogen receptor (ER) gene regulation in breast cancer. Eur. J. Endocrinol. 2016, 175, R41–R49. [Google Scholar] [CrossRef] [Green Version]
- Yaşar, P.; Ayaz, G.; Muyan, M. Estradiol-Estrogen Receptor α Mediates the Expression of the CXXC5 Gene through the Estrogen Response Element-Dependent Signaling Pathway. Sci. Rep. 2016, 6, 37808. [Google Scholar] [CrossRef] [Green Version]
- Farzad, P. Molecular Pathways of Estrogen Receptor Action. Int. J. Mol. Sci. 2018, 19, 2591. [Google Scholar] [CrossRef] [Green Version]
- Fuentes, N.; Silveyra, P. Estrogen receptor signaling mechanisms. Adv. Protein Chem. Struct. Biol. 2019, 116, 135–170. [Google Scholar] [CrossRef]
- Yaşar, P.; Ayaz, G.; User, S.D.; Güpür, G.; Muyan, M. Molecular mechanism of estrogen-estrogen receptor signaling. Reprod. Med. Biol. 2016, 16, 4–20. [Google Scholar] [CrossRef]
- Assunta, S.; D’Agostino, Y.; Alexandrova, E.; Lamberti, J.; Pecoraro, G.; Memoli, D.; Rocco, D.; Coviello, E.; Giurato, G.; Nassa, G.; et al. Insights into the Role of Estrogen Receptor β in Triple-Negative Breast Cancer. Cancers 2020, 12, 1477. [Google Scholar] [CrossRef]
- Mal, R.; Magner, A.; David, J.; Datta, J.; Vallabhaneni, M.; Kassem, M.; Manouchehri, J.; Willingham, N.; Stover, D.; Vandeusen, J.; et al. Estrogen Receptor Beta (ERβ): A Ligand Activated Tumor Suppressor. Front. Oncol. 2020, 10, 587386. [Google Scholar] [CrossRef]
- Yeh, W.-L.; Shioda, K.; Coser, K.R.; Rivizzigno, D.; McSweeney, K.R.; Shioda, T. Fulvestrant-Induced Cell Death and Proteasomal Degradation of Estrogen Receptor α Protein in MCF-7 Cells Require the CSK c-Src Tyrosine Kinase. PLoS ONE 2013, 8, e60889. [Google Scholar] [CrossRef] [PubMed]
- Abourehab, M.A.; Alqahtani, A.M.; Youssif, B.G.; Gouda, A.M. Globally Approved EGFR Inhibitors: Insights into Their Syntheses, Target Kinases, Biological Activities, Receptor Interactions, and Metabolism. Molecules 2021, 26, 6677. [Google Scholar] [CrossRef] [PubMed]
- Akinnusi, P.A.; Olubode, S.O.; Shodehinde, S.A. Discovery of Promising Inhibitors of Epidermal Growth Factor Receptor (EGFR), Human Epidermal Growth Factor Receptor 2 (HER2), Estrogen Receptor (ER), and Phosphatidylinositol-3-kinase a (PI3Ka) for Personalized Breast Cancer Treatment. Cancer Inform. 2022, 21, 27. [Google Scholar] [CrossRef]
- Kyriakopoulou, K.; Kefali, E.; Piperigkou, Z.; Bassiony, H.; Karamanos, N.K. Advances in targeting epidermal growth factor receptor signaling pathway in mammary cancer. Cell. Signal. 2018, 51, 99–109. [Google Scholar] [CrossRef]
- Guo, P.; Pu, T.; Chen, S.; Qiu, Y.; Zhong, X.; Zheng, H.; Chen, L.; Bu, H.; Ye, F. Breast cancers with EGFR and HER2 co-amplification favor distant metastasis and poor clinical outcome. Oncol. Lett. 2017, 14, 6562–6570. [Google Scholar] [CrossRef] [Green Version]
- Subramaniyan, V.; Fuloria, S.; Gupta, G.; Kumar, D.H.; Sekar, M.; Sathasivam, K.V.; Sudhakar, K.; Alharbi, K.S.; Al-Malki, W.H.; Afzal, O.; et al. A review on epidermal growth factor receptor’s role in breast and non-small cell lung cancer. Chem. Interact. 2021, 351, 109735. [Google Scholar] [CrossRef] [PubMed]
- Du, X.; Yang, B.; An, G.; Assaraf, Y.G.; Cao, X.; Xia, J. Acquired resistance to third-generation EGFR-TKIs and emerging next-generation EGFR inhibitors. Innovations 2021, 2, 100103. [Google Scholar] [CrossRef]
- Cai, W.Q.; Zeng, L.S.; Wang, L.F.; Wang, Y.Y.; Cheng, J.T.; Zhang, Y.; Han, Z.W.; Zhou, Y.; Huang, S.L.; Wang, X.W.; et al. The Latest Battles Between EGFR Monoclonal Antibodies and Resistant Tumor Cells. Front. Oncol. 2020, 10, 1249. [Google Scholar] [CrossRef] [PubMed]
- Iancu, G.; Serban, D.; Badiu, C.D.; Tanasescu, C.; Tudosie, M.S.; Tudor, C.; Costea, D.O.; Zgura, A.; Iancu, R.; Vasile, D.; et al. Tyrosine kinase inhibitors in breast cancer (Review). Exp. Ther. Med. 2022, 23, 114. [Google Scholar] [CrossRef] [PubMed]
- Uribe, M.L.; Marrocco, I.; Yarden, Y. EGFR in Cancer: Signaling Mechanisms, Drugs, and Acquired Resistance. Cancers 2021, 13, 2748. [Google Scholar] [CrossRef] [PubMed]
- Sun, L.-R.; Zhou, W.; Zhang, H.-M.; Guo, Q.-S.; Yang, W.; Li, B.-J.; Sun, Z.-H.; Gao, S.-H.; Cui, R.-J. Modulation of Multiple Signaling Pathways of the Plant-Derived Natural Products in Cancer. Front. Oncol. 2019, 9, 1153. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Castellano, J.M.; Ramos-Romero, S.; Perona, J.S. Oleanolic Acid: Extraction, Characterization and Biological Activity. Nutrients 2022, 14, 623. [Google Scholar] [CrossRef]
- Žiberna, L.; Šamec, D.; Mocan, A.; Nabavi, S.F.; Bishayee, A.; Farooqi, A.A.; Sureda, A.; Nabavi, S.M. Oleanolic Acid Alters Multiple Cell Signaling Pathways: Implication in Cancer Prevention and Therapy. Int. J. Mol. Sci. 2017, 18, 643. [Google Scholar] [CrossRef] [Green Version]
- Tang, Z.Y.; Li, Y.; Tang, Y.T.; Ma, X.D.; Tang, Z.Y. Anticancer activity of oleanolic acid and its derivatives: Recent advances in evidence, target profiling and mechanisms of action. Biomed. Pharm. 2022, 145, 112397. [Google Scholar] [CrossRef] [PubMed]
- Jiang, Q.; Yang, X.; Du, P.; Zhang, H.; Zhang, T. Dual strategies to improve oral bioavailability of oleanolic acid: Enhancing water-solubility, permeability and inhibiting cytochrome P450 isozymes. Eur. J. Pharm. Biopharm. 2016, 99, 65–72. [Google Scholar] [CrossRef]
- Lisiak, N.; Paszel-Jaworska, A.; Bednarczyk-Cwynar, B.; Zaprutko, L.; Kaczmarek, M.; Rybczyńska, M. Methyl 3-hydroxyimino-11-oxoolean-12-en-28-oate (HIMOXOL), a synthetic oleanolic acid derivative, induces both apoptosis and autophagy in MDA-MB-231 breast cancer cells. Chem. Biol. Interact. 2014, 208, 47–57. [Google Scholar] [CrossRef]
- Lisiak, N.; Toton, E.; Rubis, B.; Majer, B.; Rybczynska, M. The Synthetic Oleanane Triterpenoid HIMOXOL Induces Autophagy in Breast Cancer Cells via ERK1/2 MAPK Pathway and Beclin-1 Up-regulation. Anticancer Agents Med. Chem. 2016, 16, 1066–1076. [Google Scholar] [CrossRef] [PubMed]
- Lisiak, N.; Paszel-Jaworska, A.; Totoń, E.; Rubiś, B.; Pakuła, M.; Bednarczyk-Cwynar, B.; Zaprutko, L.; Rybczyńska, M. Semisynthetic oleanane triterpenoids inhibit migration and invasion of human breast cancer cells through downregulated expression of the ITGB1/PTK2/PXN pathway. Chem. Biol. Interact. 2017, 268, 136–147. [Google Scholar] [CrossRef] [PubMed]
- Lisiak, N.M.; Lewicka, I.; Kaczmarek, M.; Kujawski, J.; Bednarczyk-Cwynar, B.; Zaprutko, L.; Rubis, B. Oleanolic Acid’s Semisynthetic Derivatives HIMOXOL and Br-HIMOLID Show Proautophagic Potential and Inhibit Migration of HER2-Positive Breast Cancer Cells In Vitro. Int. J. Mol. Sci. 2021, 22, 11273. [Google Scholar] [CrossRef]
- Mayr, C.; Beyreis, M.; Dobias, H.; Gaisberger, M.; Pichler, M.; Ritter, M.; Jakab, M.; Neureiter, D.; Kiesslich, T. Miniaturization of the Clonogenic Assay Using Confluence Measurement. Int. J. Mol. Sci. 2018, 19, 724. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dong, X.; Li, Y.; Li, W.; Kang, W.; Tang, R.; Wu, W.; Xing, Z.; Zhou, L. The function of Cav-1 in MDA-MB-231 breast cancer cell migration and invasion induced by ectopic ATP5B. Med. Oncol. 2021, 38, 73. [Google Scholar] [CrossRef] [PubMed]
- Nistor, G.; Trandafirescu, C.; Prodea, A.; Milan, A.; Cristea, A.; Ghiulai, R.; Racoviceanu, R.; Mioc, A.; Mioc, M.; Ivan, V.; et al. Semisynthetic Derivatives of Pentacyclic Triterpenes Bearing Heterocyclic Moieties with Therapeutic Potential. Molecules 2022, 27, 6552. [Google Scholar] [CrossRef] [PubMed]
- Ayeleso, T.B.; Given, M.; Mukwevho, E.M. Oleanolic Acid and Its Derivatives: Biological Activities and Therapeutic Potential in Chronic Diseases. Molecules 2017, 22, 1915. [Google Scholar] [CrossRef] [Green Version]
- Shanmugam, M.K.; Dai, X.; Kumar, A.P.; Tan, B.K.H.; Sethi, G.; Bishayee, A. Oleanolic acid and its synthetic derivatives for the prevention and therapy of cancer: Preclinical and clinical evidence. Cancer Lett. 2014, 346, 206–216. [Google Scholar] [CrossRef] [Green Version]
- Yuan, Z.Z.; Suo, Y.R.; Hao, X.Y.; Wang, S.L.; Li, G.; Wang, H.L. Triterpenic Acids from Potentilla parvifolia and Their Protective Effects against Okadaic Acid Induced Neurotoxicity in Differentiated SH-SY5Y Cells. Biol. Pharm. Bull. 2018, 41, 885–890. [Google Scholar] [CrossRef] [Green Version]
- Borella, R.; Forti, L.; Gibellini, L.; Gaetano, M.D.; Biasi, S.D.; Cossarizza, A.; Pinti, M.N. Synthesis and Anticancer Activity of CDDO and CDDO-Me, Two Derivatives of Natural Triterpenoids. Molecules 2019, 24, 4097. [Google Scholar] [CrossRef] [Green Version]
- Ju, W.; Li, N.; Wang, J.; Yu, N.; Lei, Z.; Zhang, L.; Sun, J.; Chen, L. Design and synthesis of novel mitochondria-targeted CDDO derivatives as potential anti-cancer agents. Bioorg. Chem. 2021, 115, 105249. [Google Scholar] [CrossRef]
- Baer-Dubowska, W.; Narożna, M.; Krajka-Kuźniak, V. Anti-Cancer Potential of Synthetic Oleanolic Acid Derivatives and Their Conjugates with NSAIDs. Molecules 2021, 26, 4957. [Google Scholar] [CrossRef] [PubMed]
- Kamran, S.; Sinniah, A.; Abdulghani, M.A.M.; Alshawsh, M.A. Therapeutic Potential of Certain Terpenoids as Anticancer Agents: A Scoping Review. Cancers 2022, 14, 1100. [Google Scholar] [CrossRef] [PubMed]
- Paszel, A.; Rubiś, B.; Bednarczyk-Cwynar, B.; Zaprutko, L.; Kaczmarek, M.; Hofmann, J.; Rybczyńska, M. Oleanolic acid derivative methyl 3,11-dioxoolean-12-en-28-olate targets multidrug resistance related to ABCB1. Pharmacol. Rep. 2011, 63, 1500–1517. [Google Scholar] [CrossRef] [PubMed]
- Şoica, C.; Voicu, M.; Ghiulai, R.; Dehelean, C.; Racoviceanu, R.; Trandafirescu, C.; Roșca, O.J.; Nistor, G.; Mioc, M.; Miocdoi, A. Natural Compounds in Sex Hormone-Dependent Cancers: The Role of Triterpenes as Therapeutic Agents. Front. Endocrinol. 2021, 11, 612396. [Google Scholar] [CrossRef]
- Węsierska-Gądek, J.; Schreiner, T.; Maurer-Granofszky, M.; Waringer, A.; Ranftler, C. Phenol red in the culture medium strongly affects the susceptibility of human MCF-7 cells to roscovitine. Cell. Mol. Biol. Lett. 2007, 12, 280–293. [Google Scholar] [CrossRef]
- Pilgrim, C.R.; McCahill, K.A.; Rops, J.G.; Dufour, J.M.; Russell, K.A.; Koch, T.G. A Review of Fetal Bovine Serum in the Culture of Mesenchymal Stromal Cells and Potential Alternatives for Veterinary Medicine. Front. Vet. Sci. 2022, 9, 557. [Google Scholar] [CrossRef]
- Katzenellenbogen, B.S.; Kendra, K.L.; Norman, M.J.; Berthois, Y. Proliferation, hormonal responsiveness, and estrogen receptor content of MCF-7 human breast cancer cells grown in the short-term and long-term absence of estrogens. Cancer Res. 1987, 47, 4355–4360. [Google Scholar]
- Xie, B.-P.; Shi, L.-Y.; Li, J.-P.; Zeng, Y.; Liu, W.; Tang, S.-Y.; Jia, L.-J.; Zhang, J.; Gan, G.-X. Oleanolic acid inhibits RANKL-induced osteoclastogenesis via ER alpha/miR-503/RANK signaling pathway in RAW264.7 cells. Biomed. Pharmacother. 2019, 117, 109045. [Google Scholar] [CrossRef]
- Zhou, Z.; Dong, Y.; Li, N.; Niu, M.; Wang, S.; Zhou, Y.; Sun, Z.; Chu, P.; Tang, Z. An oleanolic acid derivative, K73-03, inhibits pancreatic cancer cells proliferation in vitro and in vivo via blocking EGFR/Akt pathway. Cell Biol. Int. 2022, 46, 1801–1813. [Google Scholar] [CrossRef]
- Li, Y.; Fan, J.; Ju, D. Neurotoxicity concern about the brain targeting delivery systems. Brain Target. Drug Deliv. Syst. 2018, 15, 377–408. [Google Scholar] [CrossRef]
- Qu, X.; Zou, Z.; Sun, Q.; Luby-Phelps, K.; Cheng, P.; Hogan, R.N.; Gilpin, C.; Levine, B. Autophagy Gene-Dependent Clearance of Apoptotic Cells during Embryonic Development. Cell 2007, 128, 931–946. [Google Scholar] [CrossRef] [Green Version]
- Chen, N.; Karantza-Wadsworth, V. Role and regulation of autophagy in cancer. Biochim. Biophys. Acta 2009, 1793, 1516–1523. [Google Scholar] [CrossRef] [Green Version]
- Hasan, A.; Rizvi, S.F.; Parveen, S.; Pathak, N.; Nazir, A.; Mir, S.S. Crosstalk Between ROS and Autophagy in Tumorigenesis: Understanding the Multifaceted Paradox. Front. Oncol. 2022, 12, 852424. [Google Scholar] [CrossRef] [PubMed]
- Kroemer, G.; Levine, B. Autophagic cell death: The story of a misnomer. Nat. Rev. Mol. Cell Biol. 2008, 9, 1004–1010. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fuhler, G.M.; Tyl, M.R.; Olthof, S.G.; Drayer, A.L.; Blom, N.; Vellenga, E. Distinct roles of the mTOR components Rictor and Raptor in MO7e megakaryocytic cells. Eur. J. Haematol. 2009, 83, 235–245. [Google Scholar] [CrossRef] [PubMed]
- Deleyto-Seldas, N.; Efeyan, A. The mTOR–Autophagy Axis and the Control of Metabolism. Front. Cell Dev. Biol. 2021, 9, 655731. [Google Scholar] [CrossRef] [PubMed]
- Tanida, I.; Ueno, T.; Kominami, E. LC3 and Autophagy. Methods Mol. Biol. 2008, 445, 77–88. [Google Scholar] [CrossRef] [PubMed]
- Seetharaman, S.; Etienne-Manneville, S. Cytoskeletal Crosstalk in Cell Migration. Trends Cell Biol. 2020, 30, 720–735. [Google Scholar] [CrossRef] [PubMed]
- Jiang, W.G.; Sanders, A.J.; Katoh, M.; Ungefroren, H.; Gieseler, F.; Prince, M.; Thompson, S.K.; Zollo, M.; Spano, D.; Dhawan, P.; et al. Tissue invasion and metastasis: Molecular, biological and clinical perspectives. Semin. Cancer Biol. 2015, 35, S244–S275. [Google Scholar] [CrossRef]
- Bauer, M.S.; Baumann, F.; Daday, C.; Redondo, P.; Durner, E.; Jobst, M.A.; Milles, L.F.; Mercadante, D.; Pippig, D.A.; Gaub, H.E.; et al. Structural and mechanistic insights into mechanoactivation of focal adhesion kinase. Proc. Natl. Acad. Sci. USA 2019, 116, 6766–6774. [Google Scholar] [CrossRef] [Green Version]
- Legerstee, K.; Houtsmu, A.B. A Layered View on Focal Adhesions. Biology 2021, 10, 1189. [Google Scholar] [CrossRef] [PubMed]
- Yoon, H.; Dehart, J.P.; Murphy, J.M.; Lim, S.-T.S. Understanding the Roles of FAK in Cancer. J. Histochem. Cytochem. 2014, 63, 114–128. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhou, J.; Yi, Q.; Tang, L. The roles of nuclear focal adhesion kinase (FAK) on Cancer: A focused review. J. Exp. Clin. Cancer Res. 2019, 38, 250. [Google Scholar] [CrossRef] [Green Version]
- Patil, V.M. Structure-activity relationship studies on anti-cancer bioactive natural products. Stud. Nat. Prod. Chem. 2022, 72, 147–199. [Google Scholar] [CrossRef]
- Paul, A.; Hazra, S.; da Silva, M.F.C.G.; Pombeiro, A.J.L. Biological Evaluation of Azo- and Imino-Based Carboxylate Triphenyltin(IV) Compounds. Eur. J. Inorg. Chem. 2020, 2020, 930–941. [Google Scholar] [CrossRef]
- Dostal, J. Essentials of Medical Chemistry and Biochemistry. 2014, pp. 1–211. Available online: https://www.studocu.com/en-gb/document/st-georges-hospital-medical-school/medicine/essentials-of-medical-chemistry-and-biochemistry-textbook/39998012 (accessed on 30 December 2022).
- Taylor, P.G.; Loo, O.A.D.; Bonilla, J.A.; Murillo, R. Anticancer activities of two sesquiterpene lactones, millerenolide and thieleanin isolated from Viguiera sylvatica and Decachaeta thieleana. Fitoterapia 2008, 79, 428–432. [Google Scholar] [CrossRef]
- Tantithanaporn, S.; Wattanapiromsakul, C.; Itharat, A.; Keawpradub, N. Cytotoxic activity of acetogenins and styryl lactones isolated from Goniothalamus undulatus Ridl. root extracts against a lung cancer cell line (COR-L23). Phytomedicine 2011, 18, 486–490. [Google Scholar] [CrossRef]
- de Oca, M.N.M.; Vara, J.; Milla, L.; Rivarola, V.; Ortiz, C.S. Physicochemical Properties and Photodynamic Activity of Novel Derivatives of Triarylmethane and Thiazine. Arch. Pharm. 2013, 346, 255–265. [Google Scholar] [CrossRef]
- Das, A.J.; Das, M.K.; Singh, S.P.; Saikia, P.P.; Singh, N.; Islam, J.; Ansari, A.; Chattopadhyay, P.; Rajamani, P.; Miyaji, T.; et al. Synthesis of salicylic acid phenylethyl ester (SAPE) and its implication in immunomodulatory and anticancer roles. Sci. Rep. 2022, 12, 8735. [Google Scholar] [CrossRef]
- Felzen, V.; Hiebel, C.; Koziollek-Drechsler, I.; Reißig, S.; Wolfrum, U.; Kögel, D.; Brandts, C.; Behl, C.; Morawe, T. Estrogen receptor α regulates non-canonical autophagy that provides stress resistance to neuroblastoma and breast cancer cells and involves BAG3 function. Cell Death Dis. 2015, 6, e1812. [Google Scholar] [CrossRef] [Green Version]
- Tan, X.; Thapa, N.; Sun, Y.; Anderson, R.A. A Kinase-Independent Role for EGF Receptor in Autophagy Initiation. Cell 2015, 160, 145–160. [Google Scholar] [CrossRef] [Green Version]
- Ahmad, V.U.; Atta-ur-Rahman. Pentacyclic Triterpenoids. In Handbook of Natural Products Data; Elsevier Science: Amsterdam, The Netherlands, 1994; Volume 2. [Google Scholar]
- Chen, L.; Zhang, Y.; Kong, X.; Peng, S.; Tian, J. Synthesis and biological evaluation of nitric oxide-releasing derivatives of oleanolic acid as inhibitors of HepG2 cell apoptosis. Bioorg. Med. Chem. Lett. 2011, 17, 2979–2982. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lewis, K.G.; Tucker, D.J. The separation of substituted olean-12-en- 28-oic acid from the corresponding urs-12-en-28-oic acid isomer. Aust. J. Chem. 1983, 36, 2297–2305. [Google Scholar] [CrossRef]
- Zaprutko, L.; Partyka, D.; Bednarczyk-Cwynar, B. Triterpenoids. Part 21: Oleanolic acid azaderivatives as percutaneous transport promoter. Bioorg. Med. Chem. Lett. 2004, 14, 472324. [Google Scholar] [CrossRef] [PubMed]
- Sikora, M.J.; Johnson, M.D.; Lee, A.V.; Oesterreich, S. Endocrine Response Phenotypes Are Altered by Charcoal-Stripped Serum Variability. Endocrinology 2016, 157, 3760–3766. [Google Scholar] [CrossRef] [Green Version]
- Chatuphonprasert, W.; Jarukamjorn, K.; Kuncharoenwirat, N. Effect of phenol red on cell cultures. Isan J. Pharm. Sci. 2021, 17, 13–23. [Google Scholar] [CrossRef]
- Pedersen, M.W.; Pedersen, N.; Ottesen, L.H.; Poulsen, H.S. Differential response to gefitinib of cells expressing normal EGFR and the mutant EGFRvIII. Br. J. Cancer 2005, 93, 915–923. [Google Scholar] [CrossRef]
- Garczarczyk, D.; Toton, E.; Biedermanna, V.; Rosivatza, E.; Rechfelda, F.; Rybczynska, M.; Hofmann, J. Signal transduction of constitutively active protein kinase C epsilon. Cell. Signal. 2009, 21, 745–752. [Google Scholar] [CrossRef] [PubMed]










{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| MCF7 IC50 [µM] | MDA-MB-231 IC50 [µM] | ||||
|---|---|---|---|---|---|
| Receptor Status | ER+ | ER− | EGFR+ | EGFR− | |
| Compound | |||||
| OA | 12.7 ± 0.45 | 31.71 ± 2.98 ** | >50 | 22.47 ± 2.49 ** | |
| HIMOXOL | 3.22 ± 0.42 | 1.63 ± 0.1 ** | 21 | 1.72 ± 0.08 *** | |
| Br-HIMOLID | 6.58 ± 0.74 | 3.53 ± 0.4 ** | >50 | 3.44 ± 0.19 *** | |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Lisiak, N.; Dzikowska, P.; Wisniewska, U.; Kaczmarek, M.; Bednarczyk-Cwynar, B.; Zaprutko, L.; Rubis, B. Biological Activity of Oleanolic Acid Derivatives HIMOXOL and Br-HIMOLID in Breast Cancer Cells Is Mediated by ER and EGFR. Int. J. Mol. Sci. 2023, 24, 5099. https://doi.org/10.3390/ijms24065099
Lisiak N, Dzikowska P, Wisniewska U, Kaczmarek M, Bednarczyk-Cwynar B, Zaprutko L, Rubis B. Biological Activity of Oleanolic Acid Derivatives HIMOXOL and Br-HIMOLID in Breast Cancer Cells Is Mediated by ER and EGFR. International Journal of Molecular Sciences. 2023; 24(6):5099. https://doi.org/10.3390/ijms24065099
Chicago/Turabian StyleLisiak, Natalia, Patrycja Dzikowska, Urszula Wisniewska, Mariusz Kaczmarek, Barbara Bednarczyk-Cwynar, Lucjusz Zaprutko, and Blazej Rubis. 2023. "Biological Activity of Oleanolic Acid Derivatives HIMOXOL and Br-HIMOLID in Breast Cancer Cells Is Mediated by ER and EGFR" International Journal of Molecular Sciences 24, no. 6: 5099. https://doi.org/10.3390/ijms24065099