
Impact of Double-Stranded RNA Internalization on Hematopoietic Progenitors and Krebs-2 Cells and Mechanism
 , ,
, ,
Abstract
:1. Introduction
2. Results
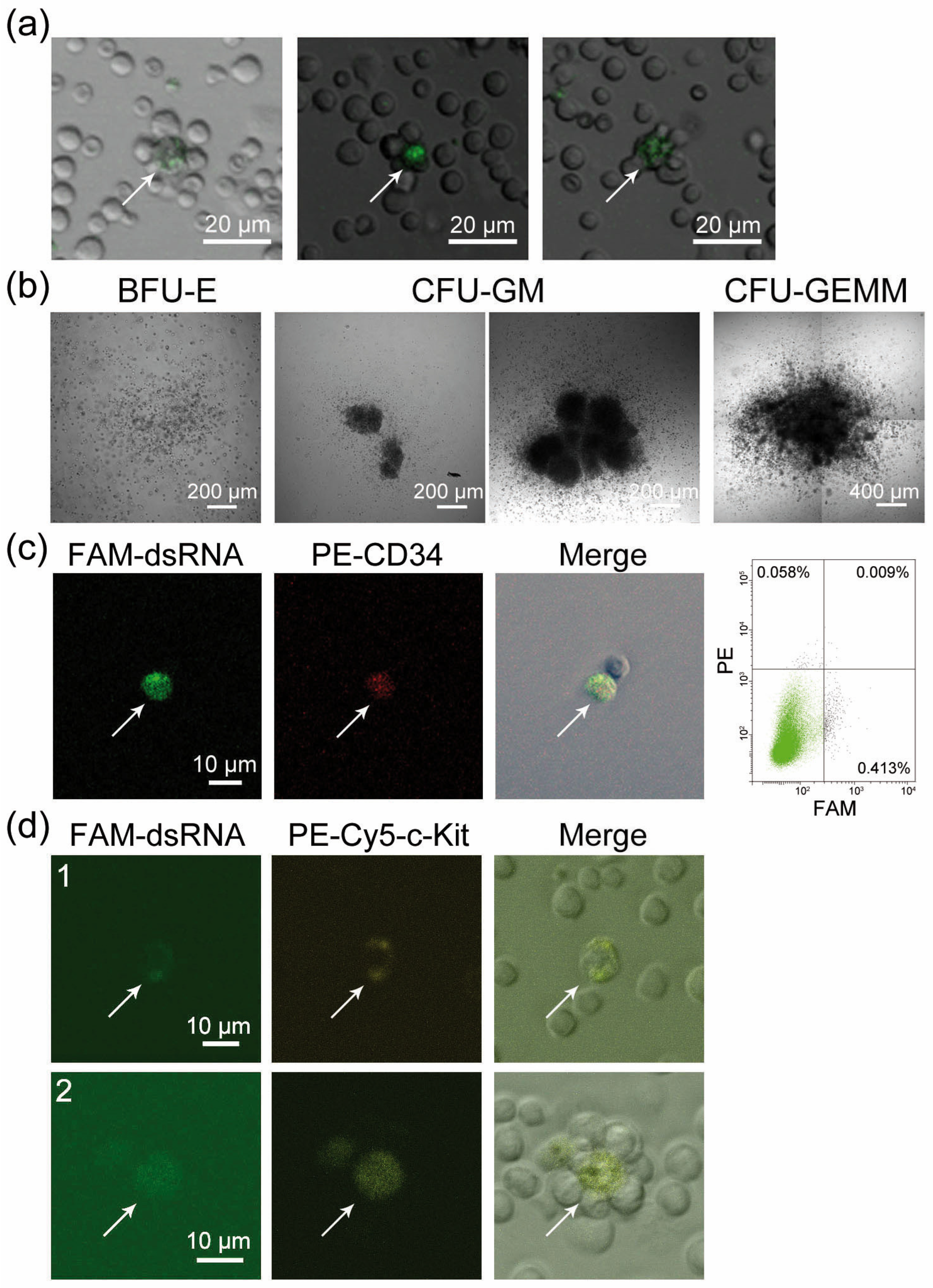
2.1. CD34+ Poorly Differentiated Bone Marrow Cells Are Capable of Internalizing FAM-dsRNA. Treating Bone Marrow Cells with Synthetic dsRNA Stimulates Colony Formation by Hematopoietic Precursors
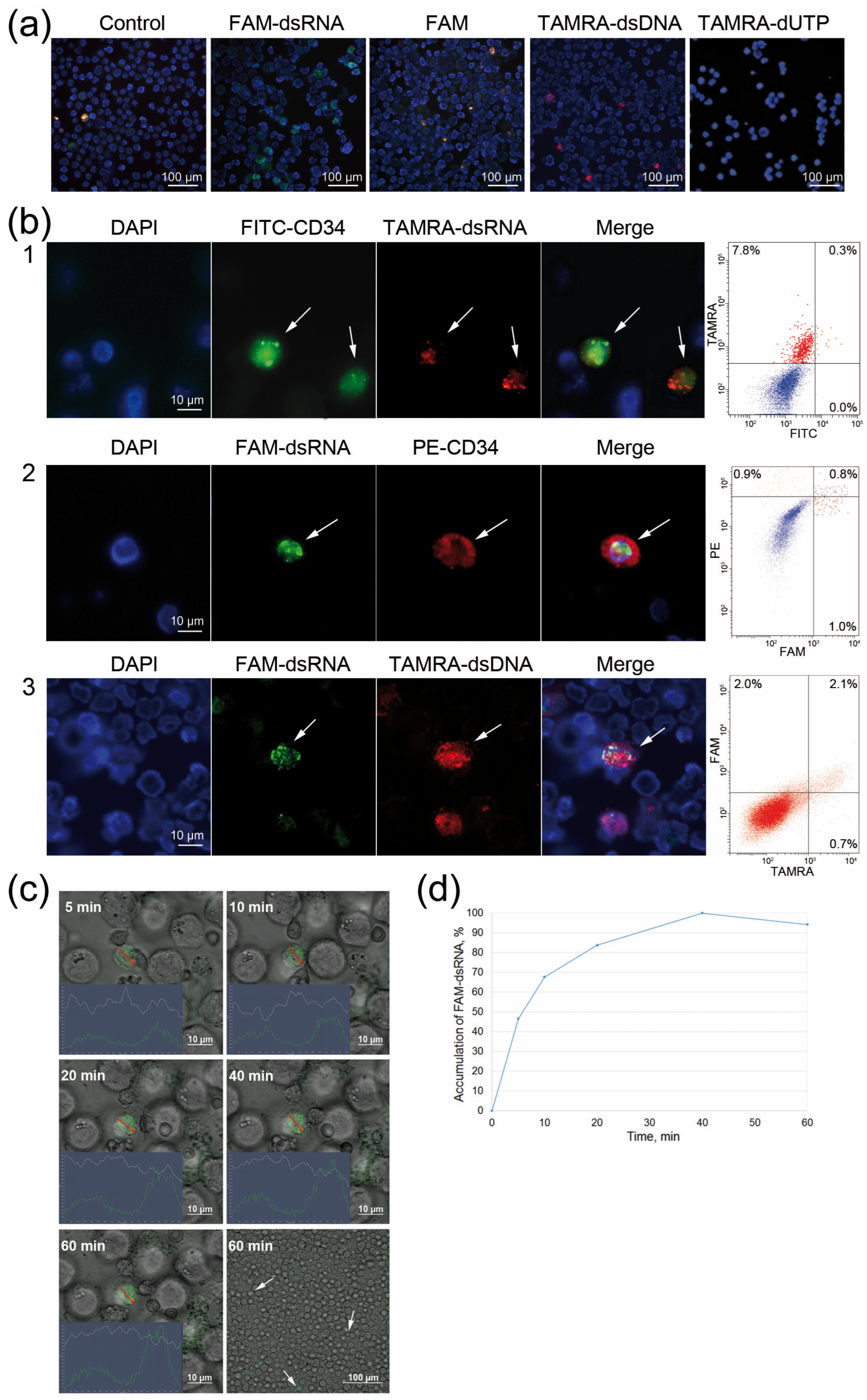
2.2. Comparative Analysis of Internalization of FAM-dsRNA and TAMRA-dsDNA Fragments into CD34+ Krebs-2 Cells
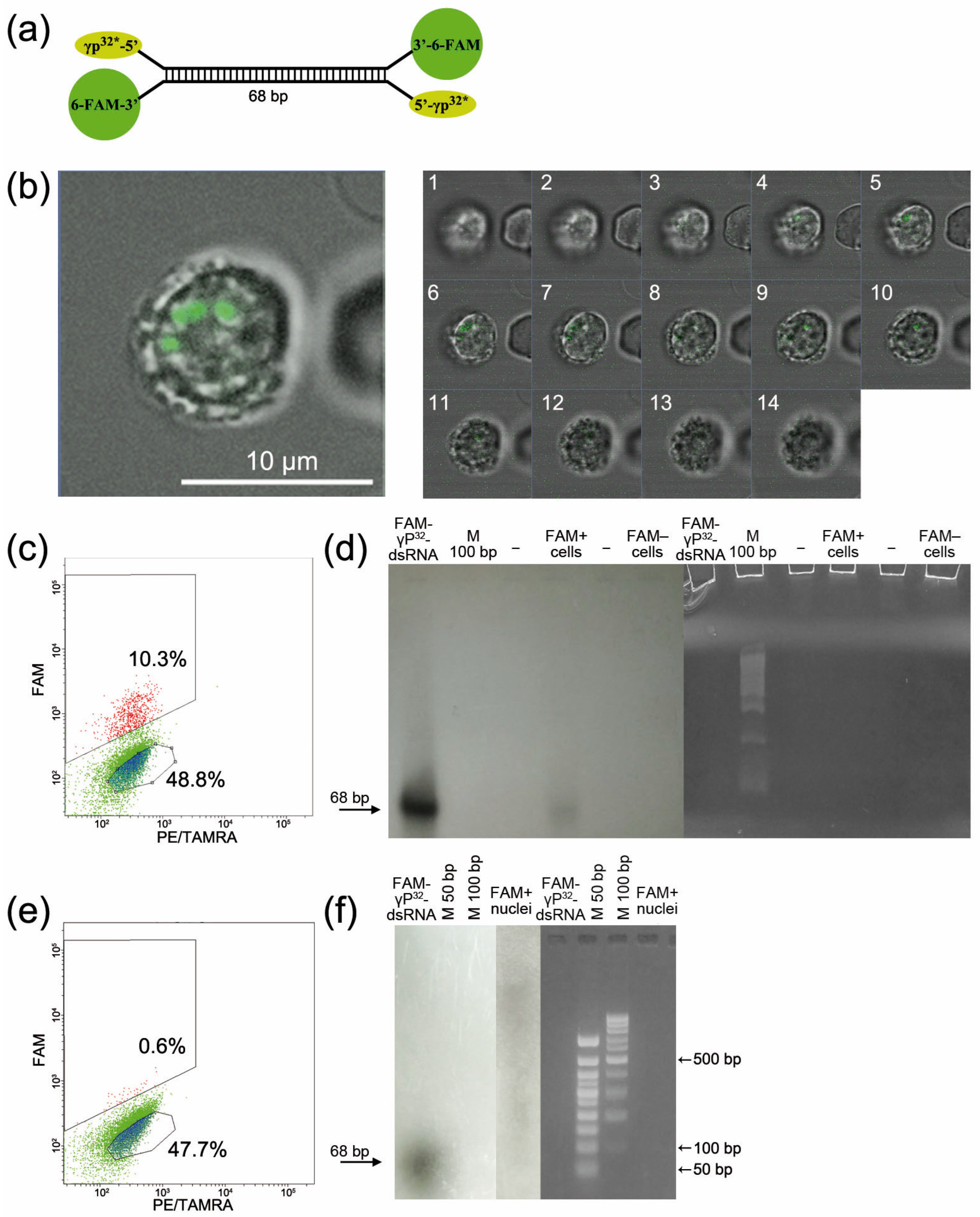
2.3. Direct Evidence for the Internalization of Synthetic dsRNA into a Eukaryotic Cell (Exemplified by Krebs-2 Ascites Cells)
2.4. Direct Evidence for the Internalization of Synthetic dsRNA into a Eukaryotic Cell (Exemplified by Krebs-2 Ascites Cells)
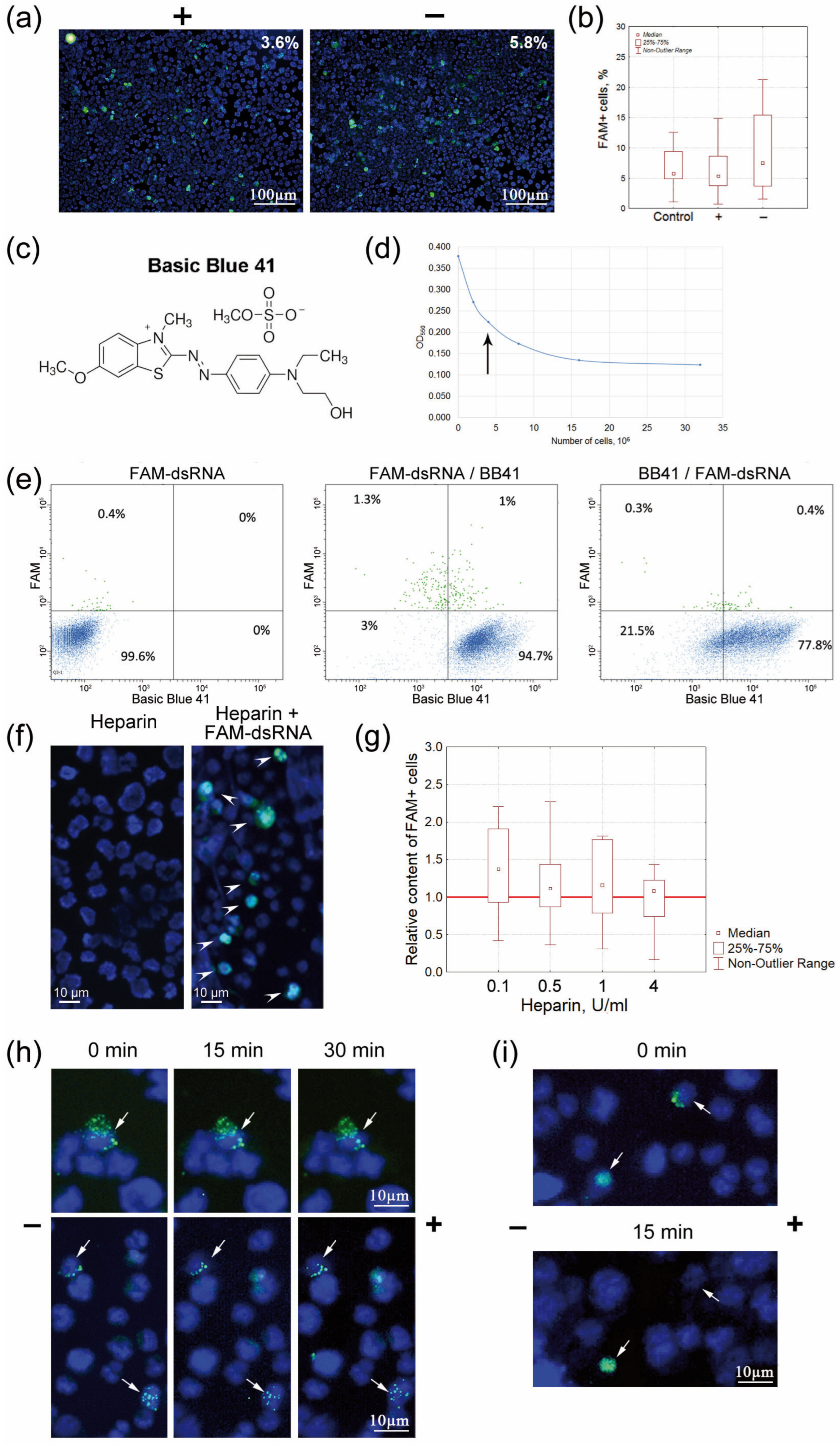
2.4.1. Determining the Modes of Interaction between FAM-dsRNA and Cell Surface Elements
Free-Flow Gel Electrophoresis of Krebs-2 Cells Followed by Quantification of Cells Internalizing FAM-dsRNA at the Opposite Poles of the Electrophoresis Chamber
Assessing the Mode of Internalization of FAM-dsRNA Using the High-Specificity Positively Charged Basic Blue 41 Dye
Assessing the Charge of dsRNA-Internalizing Cells Using Heparin
2.4.2. Characterizing the Changes in the Spatial Position of FAM-dsRNA Associated with the Small Population of Krebs-2 Cells after Microgel Electrophoresis
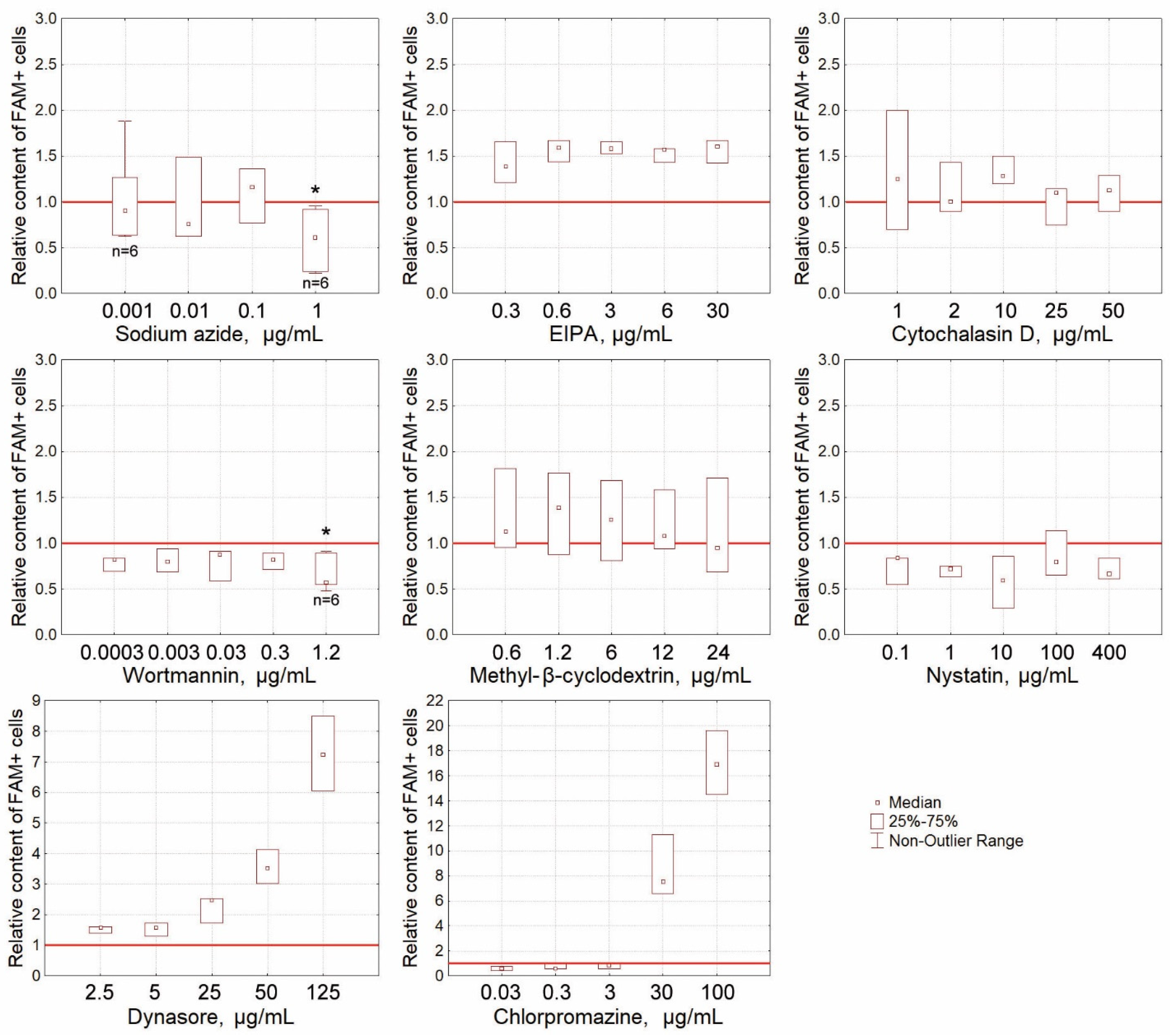
2.4.3. The Effect of Blocking Specific Endocytosis Pathways on the Percentage of FAM+ Krebs-2 Cells
2.4.4. Final note on the internalization section
- (1)
- The localization of FAM labeled material in c-Kit+ (a marker of long-term hematopoietic stem cells) and CD34+ cells (a marker of short-term hematopoietic stem cells and multipotent progenitors);
- (2)
- The induction of the proliferation of primitive precursors treated with a dsRNA preparation;
- (3)
- Radioprotection and the formation of splenic colonies [9];
- (4)
- The rescue of hematopoietic lineage growth [10].
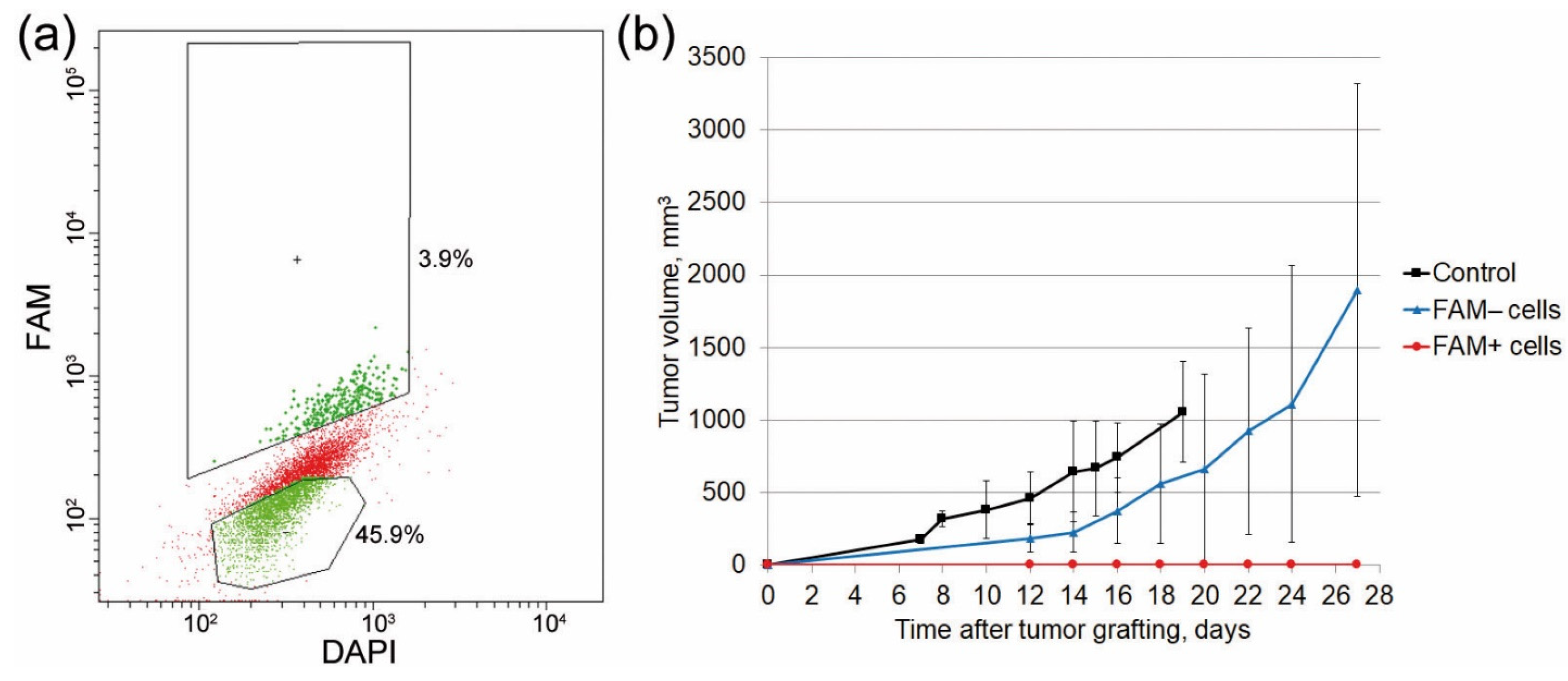
2.5. Analysis of the Tumorigenic Potential of FAM+ Cells
2.6. Identifying the Critical Organ for Homing of FAM+ Hematopoietic Progenitors
3. Discussion
4. Materials and Methods
4.1. Experimental Animals and Cells
4.2. Nucleic Acid Probes
4.3. Internalization of Labeled Nucleic Acids into the Bone Marrow Cells or Krebs-2 Cells
4.4. Stimulation of Bone Marrow Colony Growth by dsRNA
4.5. Autoradiographic Assay
4.6. Microgel Electrophoresis and Membrane Electrophoresis of Cells
4.7. Treating Cells with Endocytosis Inhibitors
4.8. Grafting Cells to Mice
4.9. Analyzing the Stability of γP32-dsRNA in Mouse Bloodstream
4.10. Analysis of the Amount of Radioactive Label in Organs of Mice after Intravenous Injection of fam-γp32-dsrna-Labeled Bone Marrow Cells
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Reisz, J.A.; Bansal, N.; Qian, J.; Zhao, W.; Furdui, C.M. Effects of Ionizing Radiation on Biological Molecules—Mechanisms of Damage and Emerging Methods of Detection. Antioxid. Redox Signal. 2014, 21, 260–292. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bergonié, J.; Tribondeau, L. Interpretation of Some Results from Radiotherapy and an Attempt to Determine a Rational Treatment Technique. 1906. Yale J. Biol. Med. 2003, 76, 181–182. [Google Scholar] [PubMed]
- Vogin, G.; Foray, N. The Law of Bergonié and Tribondeau: A Nice Formula for a First Approximation. Int. J. Radiat. Biol. 2013, 89, 2–8. [Google Scholar] [CrossRef] [PubMed]
- Patt, H.M.; Tyree, E.B.; Straube, R.L.; Smith, D.E. Cysteine Protection against X Irradiation. Science 1949, 110, 213–214. [Google Scholar] [CrossRef] [PubMed]
- Fridovich, I. Superoxide Radical and Superoxide Dismutases. Annu. Rev. Biochem. 1995, 64, 97–112. [Google Scholar] [CrossRef] [PubMed]
- Dent, P.; Yacoub, A.; Contessa, J.; Caron, R.; Amorino, G.; Valerie, K.; Hagan, M.P.; Grant, S.; Schmidt-Ullrich, R. Stress and Radiation-Induced Activation of Multiple Intracellular Signaling Pathways 1. Radiat. Res. 2003, 159, 283–300. [Google Scholar] [CrossRef]
- Wang, Y.; Xu, C.; Du, L.Q.; Cao, J.; Liu, J.X.; Su, X.; Zhao, H.; Fan, F.Y.; Wang, B.; Katsube, T.; et al. Evaluation of the Comet Assay for Assessing the Dose-Response Relationship of DNA Damage Induced by Ionizing Radiation. Int. J. Mol. Sci. 2013, 14, 22449–22461. [Google Scholar] [CrossRef] [Green Version]
- Ward, J.F. DNA Damage Produced by Ionizing Radiation in Mammalian Cells: Identities, Mechanisms of Formation, and Reparability. Prog. Nucleic. Acid. Res. Mol. Biol. 1988, 35, 95–125. [Google Scholar] [CrossRef]
- Ritter, G.; Nikolin, V.; Popova, N.; Proskurina, A.; Kisaretova, P.; Taranov, O.; Dubatolova, T.; Dolgova, E.; Potter, E.; Kirikovich, S.; et al. Characterization of Biological Peculiarities of the Radioprotective Activity of Double-Stranded RNA Isolated from Saccharomyces Cerevisiae. Int. J. Radiat. Biol. 2020, 96, 1173–1191. [Google Scholar] [CrossRef]
- Dubatolova, T.D.; Ritter, G.S.; Proskurina, A.S.; Kisaretova, P.E.; Nikolin, V.P.; Popova, N.A.; Ruzanova, V.S.; Taranov, O.S.; Kolchanov, N.A.; Bogachev, S.S. Changes in the Number and Morphology of Blood Cells in Mice Pretreated with RNA Preparations and Exposed to 8 Gy of Gamma Radiation. Int. J. Radiat. Res. 2022, 20, 361–368. [Google Scholar] [CrossRef]
- Challen, G.A.; Boles, N.; Lin, K.K.Y.; Goodell, M.A. Mouse Hematopoietic Stem Cell Identification and Analysis. Cytom. A 2009, 75, 14–24. [Google Scholar] [CrossRef] [PubMed]
- Dolgova, E.V.; Alyamkina, E.A.; Efremov, Y.R.; Nikolin, V.P.; Popova, N.A.; Tyrinova, T.V.; Kozel, A.V.; Minkevich, A.M.; Andrushkevich, O.M.; Zavyalov, E.L.; et al. Identification of Cancer Stem Cells and a Strategy for Their Elimination. Cancer Biol. Ther. 2014, 15, 1378–1394. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Potter, E.A.; Dolgova, E.V.; Proskurina, A.S.; Minkevich, A.M.; Efremov, Y.R.; Taranov, O.S.; Omigov, V.V.; Nikolin, V.P.; Popova, N.A.; Bayborodin, S.I.; et al. A Strategy to Eradicate Well-Developed Krebs-2 Ascites in Mice. Oncotarget 2016, 7, 11580–11594. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Potter, E.A.; Dolgova, E.V.; Proskurina, A.S.; Zavyalov, E.L.; Taranov, O.S.; Baiborodin, S.I.; Alexander, A. Gene Expression Profiling of Tumor-Initiating Stem Cells from Mouse Krebs-2 Carcinoma Using a Novel Marker of Poorly Differentiated Cells. Oncotarget 2017, 8, 9425–9441. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ritter, G.S.; Dolgova, E.V.; Petrova, D.D.; Efremov, Y.R.; Proskurina, A.S.; Potter, E.A.; Ruzanova, V.S.; Kirikovich, S.S.; Levites, E.V.; Taranov, O.S.; et al. The New General Biological Property of Stem-like Tumor Cells Part I. Peculiarities of the Process of the Double-Stranded DNA Fragments Internalization into Stem-like Tumor Cells. Front. Genet. 2022, 13, 954395. [Google Scholar] [CrossRef] [PubMed]
- Dolgova, E.V.; Potter, E.A.; Proskurina, A.S.; Minkevich, A.M.; Chernych, E.R.; Ostanin, A.A.; Efremov, Y.R.; Bayborodin, S.I.; Nikolin, V.P.; Popova, N.A.; et al. Properties of Internalization Factors Contributing to the Uptake of Extracellular DNA into Tumor-Initiating Stem Cells of Mouse Krebs-2 Cell Line. Stem Cell Res. Ther. 2016, 7, 76. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Höfer, T.; Rodewald, H.R. Differentiation-Based Model of Hematopoietic Stem Cell Functions and Lineage Pathways. Blood 2018, 132, 1106–1113. [Google Scholar] [CrossRef] [Green Version]
- Pinho, S.; Frenette, P.S. Haematopoietic Stem Cell Activity and Interactions with the Niche. Nat. Rev. Mol. Cell Biol. 2019, 20, 303–320. [Google Scholar] [CrossRef]
- Choi, J.S.; Mahadik, B.P.; Harley, B.A.C. Engineering the Hematopoietic Stem Cell Niche: Frontiers in Biomaterial Science. Biotechnol. J. 2015, 10, 1529–1545. [Google Scholar] [CrossRef]
- Boggs, D.R. The Total Marrow Mass of the Mouse: A Simplified Method of Measurement. Am. J. Hematol. 1984, 16, 277–286. [Google Scholar] [CrossRef]
- Likhacheva, A.S.; Nikolin, V.P.; Popova, N.A.; Dubatolova, T.D.; Strunkin, D.N.; Rogachev, V.A.; Sebeleva, T.E.; Erofeev, I.S.; Bogachev, S.S.; Yakubov, L.A.; et al. Integration of Human DNA Fragments into the Cell Genomes of Certain Tissues from Adult Mice Treated with Cytostatic Cyclophosphamide in Combination with Human DNA. Gene Ther. Mol. Biol. 2007, 11, 185–202. [Google Scholar]
- Uehata, T.; Takeuchi, O. RNA Recognition and Immunity-Innate Immune Sensing and Its Posttranscriptional Regulation Mechanisms. Cells 2020, 9, 1701. [Google Scholar] [CrossRef] [PubMed]
- Tatematsu, M.; Funami, K.; Seya, T.; Matsumoto, M. Extracellular RNA Sensing by Pattern Recognition Receptors. J. Innate. Immun. 2018, 10, 398–406. [Google Scholar] [CrossRef] [PubMed]
- Bartok, E.; Hartmann, G. Immune Sensing Mechanisms That Discriminate Self from Altered Self and Foreign Nucleic Acids. Immunity 2020, 53, 54–77. [Google Scholar] [CrossRef] [PubMed]
- Chen, L.L.; Yang, L.; Carmichael, G.G. Molecular Basis for an Attenuated Cytoplasmic DsRNA Response in Human Embryonic Stem Cells. Cell Cycle 2010, 9, 3552–3564. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Garlatti, V.; Martin, L.; Lacroix, M.; Gout, E.; Arlaud, G.J.; Thielens, N.M.; Gaboriaud, C. Structural Insights into the Recognition Properties of Human Ficolins. J. Innate. Immun. 2010, 2, 17–23. [Google Scholar] [CrossRef]
- Gusev, E.Y.; Zotova, N.V.; Zhuravleva, Y.A.; Chereshnev, V.A. Physiological and Pathogenic Role of Scavenger Receptors in Humans. Med. Immunol. 2020, 22, 7–48. [Google Scholar] [CrossRef] [Green Version]
- PrabhuDas, M.R.; Baldwin, C.L.; Bollyky, P.L.; Bowdish, D.M.E.; Drickamer, K.; Febbraio, M.; Herz, J.; Kobzik, L.; Krieger, M.; Loike, J.; et al. A Consensus Definitive Classification of Scavenger Receptors and Their Roles in Health and Disease. J. Immunol. 2017, 198, 3775–3789. [Google Scholar] [CrossRef] [Green Version]
- Alquraini, A.; El Khoury, J. Scavenger Receptors. Curr. Biol. 2020, 30, R790–R795. [Google Scholar] [CrossRef]
- Quesenberry, P.J.; Goldberg, L.; Aliotta, J.; Dooner, M. Marrow Hematopoietic Stem Cells Revisited: They Exist in a Continuum and Are Not Defined by Standard Purification Approaches; Then There Are the Microvesicles. Front. Oncol. 2014, 4, 56. [Google Scholar] [CrossRef] [Green Version]
- Meers, C.; Keskin, H.; Storici, F. DNA Repair by RNA: Templated, or Not Templated, That Is the Question. DNA Repair 2016, 44, 17–21. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Keskin, H.; Shen, Y.; Huang, F.; Patel, M.; Yang, T.; Ashley, K.; Mazin, A.V.; Storici, F. Transcript-RNA-Templated DNA Recombination and Repair. Nature 2014, 515, 436–439. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Emara, M.M.; Ivanov, P.; Hickman, T.; Dawra, N.; Tisdale, S.; Kedersha, N.; Hu, G.F.; Anderson, P. Angiogenin-Induced TRNA-Derived Stress-Induced RNAs Promote Stress-Induced Stress Granule Assembly. J. Biol. Chem. 2010, 285, 10959–10968. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Goncalves, K.A.; Silberstein, L.; Li, S.; Severe, N.; Hu, M.G.; Yang, H.; Scadden, D.T.; Hu, G. fu Angiogenin Promotes Hematopoietic Regeneration by Dichotomously Regulating Quiescence of Stem and Progenitor Cells. Cell 2016, 166, 894–906. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ivanov, P.; Emara, M.M.; Villen, J.; Gygi, S.P.; Anderson, P. Angiogenin-Induced TRNA Fragments Inhibit Translation Initiation. Mol. Cell 2011, 43, 613–623. [Google Scholar] [CrossRef] [Green Version]
- Prehn, J.H.M.; Jirström, E. Angiogenin and TRNA Fragments in Parkinson’s Disease and Neurodegeneration. Acta Pharmacol. Sin. 2020, 41, 442–446. [Google Scholar] [CrossRef]
- Pizzo, E.; Sarcinelli, C.; Sheng, J.; Fusco, S.; Formiggini, F.; Netti, P.; Yu, W.; D’Alessio, G.; Hu, G.F. Ribonuclease/Angiogenin Inhibitor 1 Regulates Stressinduced Subcellular Localization of Angiogenin to Control Growth and Survival. J. Cell Sci. 2013, 126, 4308–4319. [Google Scholar] [CrossRef] [Green Version]
- Peveri, P.; Walz, A.; Dewald, B.; Baggiolini, M. A Novel Neutrophil-Activating Factor Produced by Human Mononuclear Phagocytes. J. Exp. Med. 1988, 167, 1547–1559. [Google Scholar] [CrossRef] [Green Version]
- Strieter, R.M.; Kunkel, S.L.; Showell, H.J.; Marks, R.M. Monokine-Induced Gene Expression of a Human Endothelial Cell-Derived Neutrophil Chemotactic Factor. Biochem. Biophys. Res. Commun. 1988, 156, 1340–1345. [Google Scholar] [CrossRef] [Green Version]
- Kwon, O.J.; Au, B.T.; Collins, P.D.; Adcock, I.M.; Mak, J.C.; Robbins, R.R.; Chung, K.F.; Barnes, P.J. Tumor Necrosis Factor-Induced Interleukin-8 Expression in Cultured Human Airway Epithelial Cells. Am. J. Physiol. 1994, 267, L398–L405. [Google Scholar] [CrossRef] [PubMed]
- Wanninger, J.; Neumeier, M.; Weigert, J.; Bauer, S.; Weiss, T.S.; Schäffler, A.; Krempl, C.; Bleyl, C.; Aslanidis, C.; Schölmerich, J.; et al. Adiponectin-Stimulated CXCL8 Release in Primary Human Hepatocytes Is Regulated by ERK1/ERK2, P38 MAPK, NF-KappaB, and STAT3 Signaling Pathways. Am. J. Physiol. Gastrointest. Liver Physiol. 2009, 297, G611–G618. [Google Scholar] [CrossRef] [PubMed]
- French, B.M.; Sendil, S.; Sepuru, K.M.; Ranek, J.; Burdorf, L.; Harris, D.; Redding, E.; Cheng, X.; Laird, C.T.; Zhao, Y.; et al. Interleukin-8 Mediates Neutrophil-Endothelial Interactions in Pig-to-Human Xenogeneic Models. Xenotransplantation 2018, 25, e12385. [Google Scholar] [CrossRef] [PubMed]
- Kuckleburg, C.J.; Newman, P.J. Neutrophil Proteinase 3 Acts on Protease-Activated Receptor-2 to Enhance Vascular Endothelial Cell Barrier Function. Arterioscler. Thromb. Vasc. Biol. 2013, 33, 275–284. [Google Scholar] [CrossRef] [Green Version]
- Laterveer, L.; Lindley, I.J.; Hamilton, M.S.; Willemze, R.; Fibbe, W.E. Interleukin-8 Induces Rapid Mobilization of Hematopoietic Stem Cells with Radioprotective Capacity and Long-Term Myelolymphoid Repopulating Ability. Blood 1995, 85, 2269–2275. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ha, H.; Debnath, B.; Neamati, N. Role of the CXCL8-CXCR1/2 Axis in Cancer and Inflammatory Diseases. Theranostics 2017, 7, 1543–1588. [Google Scholar] [CrossRef] [PubMed]
- Roberts, D.D.; Kaur, S.; Isenberg, J.S. Regulation of Cellular Redox Signaling by Matricellular Proteins in Vascular Biology, Immunology, and Cancer. Antioxid. Redox Signal. 2017, 27, 874–911. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Maxhimer, J.B.; Soto-Pantoja, D.R.; Ridnour, L.A.; Shih, H.B.; Degraff, W.G.; Tsokos, M.; Wink, D.A.; Isenberg, J.S.; Roberts, D.D. Radioprotection in Normal Tissue and Delayed Tumor Growth by Blockade of CD47 Signaling. Sci. Transl. Med. 2009, 1, 3ra7. [Google Scholar] [CrossRef] [Green Version]
- Wardman, P.; Rothkamm, K.; Folkes, L.K.; Woodcock, M.; Johnston, P.J. Radiosensitization by Nitric Oxide at Low Radiation Doses. Radiat. Res. 2007, 167, 475–484. [Google Scholar] [CrossRef]
- Miller, T.W.; Soto-Pantoja, D.R.; Schwartz, A.L.; Sipes, J.M.; DeGraff, W.G.; Ridnour, L.A.; Wink, D.A.; Roberts, D.D. CD47 Receptor Globally Regulates Metabolic Pathways That Control Resistance to Ionizing Radiation. J. Biol. Chem. 2015, 290, 24858–24874. [Google Scholar] [CrossRef] [Green Version]
- Meijles, D.N.; Sahoo, S.; Al Ghouleh, I.; Amaral, J.H.; Bienes-Martinez, R.; Knupp, H.E.; Attaran, S.; Sembrat, J.C.; Nouraie, S.M.; Rojas, M.M.; et al. The Matricellular Protein TSP1 Promotes Human and Mouse Endothelial Cell Senescence through CD47 and Nox1. Sci. Signal 2017, 10, eaaj1784. [Google Scholar] [CrossRef] [Green Version]
- Dooner, M.; Cerny, J.; Colvin, G.; Demers, D.; Pimentel, J.; Greer, D.; Abedi, M.; McAuliffe, C.; Quesenberry, P. Homing and Conversion of Murine Hematopoietic Stem Cells to Lung. Blood Cells Mol. Dis. 2004, 32, 47–51. [Google Scholar] [CrossRef] [PubMed]
- Eaves, C.J. Hematopoietic Stem Cells: Concepts, Definitions, and the New Reality. Blood 2015, 125, 2605–2613. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dorn, D.C.; Dorn, A. Structural Characterization and Primary in Vitro Cell Culture of Locust Male Germline Stem Cells and Their Niche. Stem. Cell Res. 2011, 6, 112–128. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lévesque, J.P.; Helwani, F.M.; Winkler, I.G. The Endosteal “osteoblastic” Niche and Its Role in Hematopoietic Stem Cell Homing and Mobilization. Leukemia 2010, 24, 1979–1992. [Google Scholar] [CrossRef] [Green Version]
- Lee-Thedieck, C.; Spatz, J.P. Biophysical Regulation of Hematopoietic Stem Cells. Biomater. Sci. 2014, 2, 1548–1561. [Google Scholar] [CrossRef] [PubMed]
- Li, H.; Luo, Q.; Shan, W.; Cai, S.; Tie, R.; Xu, Y.; Lin, Y.; Qian, P.; Huang, H. Biomechanical Cues as Master Regulators of Hematopoietic Stem Cell Fate. Cell. Mol. Life Sci. 2021, 78, 5881–5902. [Google Scholar] [CrossRef]
- Redondo, P.A.; Pavlou, M.; Loizidou, M.; Cheema, U. Elements of the Niche for Adult Stem Cell Expansion. J. Tissue Eng. 2017, 8, 2041731417725464. [Google Scholar] [CrossRef]
- Morita, Y.; Ema, H.; Nakauchi, H. Heterogeneity and Hierarchy within the Most Primitive Hematopoietic Stem Cell Compartment. J. Exp. Med. 2010, 207, 1173–1182. [Google Scholar] [CrossRef] [Green Version]
- Cheng, M.; Qin, G. Progenitor Cell Mobilization and Recruitment: SDF-1, CXCR4, A4-Integrin, and c-Kit. Prog. Mol. Biol. Transl. Sci. 2012, 111, 243–264. [Google Scholar] [CrossRef] [Green Version]
- Wilkinson, A.C.; Igarashi, K.J.; Nakauchi, H. Haematopoietic Stem Cell Self-Renewal in Vivo and Ex Vivo. Nat. Rev. Genet. 2020, 21, 541–554. [Google Scholar] [CrossRef]
- Dorshkind, K.; Höfer, T.; Montecino-Rodriguez, E.; Pioli, P.D.; Rodewald, H.R. Do Haematopoietic Stem Cells Age? Nat. Rev. Immunol. 2020, 20, 196–202. [Google Scholar] [CrossRef] [PubMed]
- Wilson, A.; Trumpp, A. Bone-Marrow Haematopoietic-Stem-Cell Niches. Nat. Rev. Immunol. 2006, 6, 93–106. [Google Scholar] [CrossRef] [PubMed]
- Dolgova, E.V.; Efremov, Y.R.; Orishchenko, K.E.; Andrushkevich, O.M.; Alyamkina, E.A.; Proskurina, A.S.; Bayborodin, S.I.; Nikolin, V.P.; Popova, N.A.; Chernykh, E.R.; et al. Delivery and Processing of Exogenous Double-Stranded DNA in Mouse CD34+ Hematopoietic Progenitor Cells and Their Cell Cycle Changes upon Combined Treatment with Cyclophosphamide and Double-Stranded DNA. Gene 2013, 528, 74–83. [Google Scholar] [CrossRef] [PubMed]
- Dolgova, E.V.; Evdokimov, A.N.; Proskurina, A.S.; Efremov, Y.R.; Bayborodin, S.I.; Potter, E.A.; Popov, A.A.; Petruseva, I.O.; Lavrik, O.I.; Bogachev, S.S. Double-Stranded DNA Fragments Bearing Unrepairable Lesions and Their Internalization into Mouse Krebs-2 Carcinoma Cells. Nucleic. Acid Ther. 2019, 29, 278–290. [Google Scholar] [CrossRef]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Colony Type | Control | dsRNA |
|---|---|---|
| BFU-E | 13.5 ± 0.7 | 9.5 ± 0.7 |
| CFU-GM | 33.5 ± 6.4 | 45.0 ± 0.1 |
| CFU-GEMM | 2.5 ± 0.7 | 2.0 ± 1.4 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ritter, G.S.; Proskurina, A.S.; Meschaninova, M.I.; Potter, E.A.; Petrova, D.D.; Ruzanova, V.S.; Dolgova, E.V.; Kirikovich, S.S.; Levites, E.V.; Efremov, Y.R.; et al. Impact of Double-Stranded RNA Internalization on Hematopoietic Progenitors and Krebs-2 Cells and Mechanism. Int. J. Mol. Sci. 2023, 24, 4858. https://doi.org/10.3390/ijms24054858
Ritter GS, Proskurina AS, Meschaninova MI, Potter EA, Petrova DD, Ruzanova VS, Dolgova EV, Kirikovich SS, Levites EV, Efremov YR, et al. Impact of Double-Stranded RNA Internalization on Hematopoietic Progenitors and Krebs-2 Cells and Mechanism. International Journal of Molecular Sciences. 2023; 24(5):4858. https://doi.org/10.3390/ijms24054858
Chicago/Turabian StyleRitter, Genrikh S., Anastasia S. Proskurina, Maria I. Meschaninova, Ekaterina A. Potter, Daria D. Petrova, Vera S. Ruzanova, Evgeniya V. Dolgova, Svetlana S. Kirikovich, Evgeniy V. Levites, Yaroslav R. Efremov, and et al. 2023. "Impact of Double-Stranded RNA Internalization on Hematopoietic Progenitors and Krebs-2 Cells and Mechanism" International Journal of Molecular Sciences 24, no. 5: 4858. https://doi.org/10.3390/ijms24054858