
Evaluation of L-Alanine Metabolism in Bacteria and Whole-Body Distribution with Bacterial Infection Model Mice

 , , , and
, , , and
Abstract
:1. Introduction
2. Results
2.1. Accumulation of 3H-L-Ala, 3H-L-Met, 3H-L-Glu, and 3H-L-His in E. coli K-12
2.2. Accumulation of 3H-L-Ala, 3H-L-Met, 3H-L-Glu, and 3H-L-His in E. coli EC-14
2.3. Comparing the Accumulation of 3H-L-Ala in E. coli K-12 and EC-14
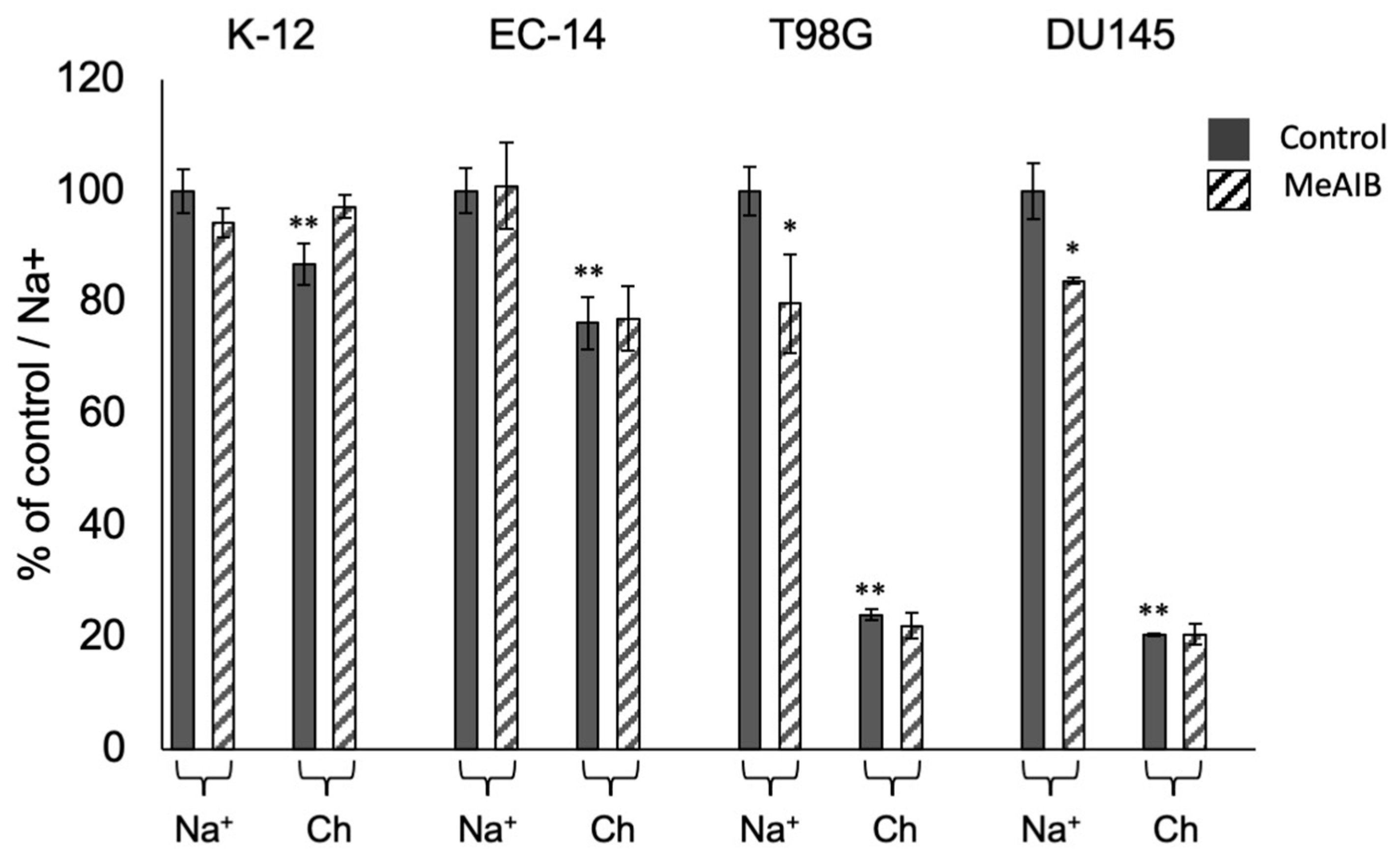
2.4. Inhibition of Amino Acid Accumulation in Bacteria and Cancer Cells
2.5. Biological Distribution of E. coli EC-14 Infection Model Mice Using 3H-L-Ala
3. Discussion
4. Materials and Methods
4.1. Bacterial Strain and Culture Conditions
4.2. Human Tumor Cell Line and Culture Conditions
4.3. Accumulation of 3H-L-Ala, 3H-L-Met, 3H-L-Glu, and 3H-L-His in E. coli K-12 and EC-14
4.4. Inhibition of Amino Acid Accumulation in Bacteria
4.5. Inhibition of Amino Acid Accumulation in Cancer Cells
4.6. Biological Distribution of E. coli EC-14 Infection Model Mice Using 3H-L-Ala
4.7. Statistical Analysis
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Cassini, A.; Högberg, L.D.; Plachouras, D.; Quattrocchi, A.; Hoxha, A.; Struelens, M.J.; Suetens, C.C.; Monnet, D.L. Attributable deaths and disability-adjusted life-years caused by infections with antibiotic-resistant bacteria in the EU and the European Economic Area in 2015: A population-level modelling analysis. Lancet Infect. Dis. 2018, 19, 56–66. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kraker, M.E.A.; Stewardson, A.J.; Harbarth, S. Will 10 Million People Die a Year due to Antimicrobial Resistance by 2050? PLoS Med. 2016, 13, e1002184. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- O’Neill, J. The Review on Antimicrobial Resistance. In Tackling Drug Resistant Infections Globally: Final Report and Recommendations; Wellcome Trust and the Department of Health of United Kingdom Government: London, UK, 2016. [Google Scholar]
- Bunschoten, A.; Welling, M.M.; Termaat, M.F.; Sathekge, M.; van Leeuwen, F.W.B. Development and Prospects of Dedicated Tracers for the Molecular Imaging of Bacterial Infections. Bioconjug. Chem. 2013, 24, 1971–1989. [Google Scholar] [CrossRef]
- Vogel, S.J.; Tank, M.; Goodyear, N. Variation in detection limits between bacterial growth phases and precision of an ATP bioluminescence system. Lett. Appl. Microbiol. 2014, 58, 370–375. [Google Scholar] [CrossRef] [PubMed]
- Schöder, H.; Larson, S.M. Positron emission tomography for prostate, bladder, and renal cancer. Semin. Nucl. Med. 2004, 34, 274–292. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hess, S.; Blomberg, B.A. The pivotal role of FDG-PET/CT in modern medicine. Acad. Radiol. 2014, 21, 232–249. [Google Scholar] [CrossRef]
- Duclos, V.; Iep, A.; Gomez, L.; Goldfarb, L.; Besson, F.L. PET Molecular Imaging: A Holistic Review of Current Practice and Emerging Perspectives for Diagnosis, Therapeutic Evaluation and Prognosis in Clinical Oncology. Int. J. Mol. Sci. 2021, 16, 4159. [Google Scholar] [CrossRef]
- Kumar, R.; Basu, S.; Torigian, D.; Anand, V.; Zhuang, H.; Alavi, A. Role of modern imaging techniques for diagnosis of infection in the era of 18F-fluorodeoxyglucose positron emission tomography. Clin. Microbiol. Rev. 2008, 21, 209–224. [Google Scholar] [CrossRef] [Green Version]
- Love, C.; Tomas, M.B.; Tronco, G.G.; Palestro, C.J. FDG PET of infection and inflammation. Radiographics 2005, 25, 1368–1375. [Google Scholar] [CrossRef] [Green Version]
- Heuker, M.; Sijbesma, J.W.A.; Aguilar, S.R.; de Jong, J.R.; Boersma, H.H.; Luurtsema, G.; Elsinga, P.H.; Glaudemans, A.W.J.M.; Van Dam, G.M.; Van Dijl, J.M.; et al. In vitro imaging of bacteria using 18F-fluorodeoxyglucose micro positron emission tomography. Sci. Rep. 2017, 7, 4973. [Google Scholar] [CrossRef]
- Britton, K.E.; Wareham, D.W.; Das, S.S.; Solanki, K.K.; Amaral, H.; Bhatnagar, A.; Katamihardja, A.H.; Malamitsi, J.; Moustafa, H.M.; Soroa, V.E.; et al. Imaging bacterial infection with 99mTc-ciprofloxacin (Infecton). J. Clin. Pathol. 2002, 55, 817–823. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bonnet, M.; Lagier, J.C.; Raoult, D.; Khelaifia, S. Bacterial culture through selective and non-selective conditions: The evolution of culture media in clinical microbiology. New Microbes New Infect. 2020, 30, 100622. [Google Scholar] [CrossRef] [PubMed]
- Okudaira, H.; Shikano, N.; Nishii, R.; Miyagi, T.; Yoshimoto, M.; Kobayashi, M.; Ohe, K.; Nakanishi, T.; Tamai, I.; Namiki, M.; et al. Putative transport mechanism and intracellular fate of trans-1-amino-3-18F-fluorocyclobutanecarboxylic acid in human prostate cancer. J. Nucl. Med. 2011, 52, 822–829. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kobayashi, M.; Mizutani, A.; Nishi, K.; Nakajima, S.; Shikano, N.; Nishii, R.; Fukuchi, K.; Kawai, K. Differences in accumulation and the transport mechanism of L- and D-methionine in high- and low-grade human glioma cells. Nucl. Med. Biol. 2017, 44, 78–82. [Google Scholar] [CrossRef] [PubMed]
- Kobayashi, M.; Hashimoto, F.; Ohe, K.; Nadamura, T.; Nishi, K.; Shikano, N.; Nishii, R.; Higashi, T.; Okazawa, H.; Kawai, K. Transport mechanism of 11C-labeled L-and D-methionine in human-derived tumor cells. Nucl. Med. Biol. 2012, 39, 1213–1218. [Google Scholar] [CrossRef]
- Nishii, R.; Higashi, T.; Kagawa, S.; Arimoto, M.; Kishibe, Y.; Takahashi, M.; Yamada, S.; Saiki, M.; Arakawa, Y.; Yamauchi, H.; et al. Differential Diagnosis between Low-Grade and High-Grade Astrocytoma Using System A Amino Acid Transport PET Imaging with C-11-MeAIB: A Comparison Study with C-11-Methionine PET Imaging. Contrast Media Mol. Imaging 2018, 2018, 1292746. [Google Scholar] [CrossRef] [Green Version]
- Zhou, Y.; Imlay, J.A. Escherichia coli K-12 Lacks a High-Affinity Assimilatory Cysteine Importer. mBio 2020, 11, e01073-20. [Google Scholar] [CrossRef]
- Auer, G.K.; Weibel, D.B. Bacterial Cell Mechanics. Biochemistry 2017, 56, 3710–3724. [Google Scholar] [CrossRef]
- Vollmer, W.; Blanot, D.; de Pedro, M.A. Peptidoglycan structure and architecture. FEMS Microbiol. Rev. 2008, 32, 149–167. [Google Scholar] [CrossRef] [Green Version]
- Muranaka, Y.; Mizutani, A.; Kobayashi, M.; Nakamoto, K.; Matsue, M.; Takagi, F.; Okazaki, K.; Nishi, K.; Yamazaki, K.; Nishii, R.; et al. 123I-BMIPP, a Radiopharmaceutical for Myocardial Fatty Acid Metabolism Scintigraphy, Could Be Utilized in Bacterial Infection Imaging. Pharmaceutics 2022, 14, 1008. [Google Scholar] [CrossRef]
- Schneider, D.W.; Heitner, T.; Alicke, B.; Light, D.R.; McLean, K.; Satozawa, N.; Parry, G.; Yoo, J.; Lewis, J.S.; Parry, R. In vivo biodistribution, PET imaging, and tumor accumulation of 86Y- and 111In-antimindin/RG-1, engineered antibody fragments in LNCaP tumor-bearing nude mice. J. Nucl. Med. 2009, 50, 435–443. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wick, W.E.; Preston, D.A.; White, W.A.; Gordee, R.S. Compound 64716, a new synthetic antibacterial agent. Antimicrob. Agents Chemother. 1973, 4, 415–420. [Google Scholar] [CrossRef] [PubMed] [Green Version]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Time after 3H-L-Ala Injection | 15 min | 60 min | 15 min | 60 min |
|---|---|---|---|---|
| Organ (%ID/g) | 2 h after Infection | 8 h after Infection | ||
| Blood | 6.54 ± 0.57 | 5.64 ± 0.35 | 5.10 ± 0.55 | 3.05 ± 0.14 |
| Heart | 4.33 ± 0.44 | 3.87 ± 0.37 | 3.90 ± 0.24 | 3.65 ± 0.26 |
| Lung | 7.66 ± 1.57 | 5.60 ± 1.31 | 6.17 ± 0.67 | 5.22 ± 0.30 |
| Liver | 4.26 ± 0.70 | 4.88 ± 1.03 | 3.96 ± 0.65 | 4.23 ± 0.54 |
| Spleen | 17.11 ± 2.39 | 15.64 ± 4.53 | 9.94 ± 0.73 | 7.60 ± 1.22 |
| Kidney | 8.90 ± 0.67 | 7.78 ± 0.54 | 7.06 ± 1.10 | 6.76 ± 0.74 |
| Infected muscle | 5.14 ± 0.57 | 4.25 ± 1.13 | 4.52 ± 1.13 | 3.99 ± 0.69 |
| Control muscle | 4.89 ± 0.81 | 3.55 ± 0.63 | 3.99 ± 0.89 | 3.92 ± 0.36 |
| Infected muscle /blood | 0.79 | 0.75 | 0.89 | 1.31 |
| Infected muscle /control muscle | 1.05 | 1.20 | 1.13 | 1.02 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Muranaka, Y.; Matsue, M.; Mizutani, A.; Kobayashi, M.; Sato, K.; Kondo, A.; Nishiyama, Y.; Ohata, S.; Nishi, K.; Yamazaki, K.; et al. Evaluation of L-Alanine Metabolism in Bacteria and Whole-Body Distribution with Bacterial Infection Model Mice. Int. J. Mol. Sci. 2023, 24, 4775. https://doi.org/10.3390/ijms24054775
Muranaka Y, Matsue M, Mizutani A, Kobayashi M, Sato K, Kondo A, Nishiyama Y, Ohata S, Nishi K, Yamazaki K, et al. Evaluation of L-Alanine Metabolism in Bacteria and Whole-Body Distribution with Bacterial Infection Model Mice. International Journal of Molecular Sciences. 2023; 24(5):4775. https://doi.org/10.3390/ijms24054775
Chicago/Turabian StyleMuranaka, Yuka, Miki Matsue, Asuka Mizutani, Masato Kobayashi, Kakeru Sato, Ami Kondo, Yuri Nishiyama, Shusei Ohata, Kodai Nishi, Kana Yamazaki, and et al. 2023. "Evaluation of L-Alanine Metabolism in Bacteria and Whole-Body Distribution with Bacterial Infection Model Mice" International Journal of Molecular Sciences 24, no. 5: 4775. https://doi.org/10.3390/ijms24054775