
Potential of Circulating miRNAs as Molecular Markers in Mood Disorders and Associated Suicidal Behavior


Abstract
:1. Introduction
2. Materials and Methods
3. Overview of miRNA Biogenesis
4. Outlining the Pathogenic Role of miRNA in Neuropsychiatric Brain
5. miRNAs as Clinical Biomarkers in MDD
6. miRNAs as Biomarkers of Treatment Response in MDD Patients
7. Circulating miRNAs in BD
8. Effect of Medications on miRNAs in BD Patients
9. Parallel miRNA Studies Comparing BD and MDD
10. Circulating miRNAs in Suicidality
11. Molecular Profiling of Genes Targeted by miRNAs Derived from MDD and BD Patients
12. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- American Psychiatric Association. DSM 5; American Psychiatric Association: Washington, DC, USA, 2013. [Google Scholar]
- Kalin, N.H. New Insights Into Major Depression and the Treatment of Bipolar Depression. Am. J. Psychiatry 2021, 178, 1071–1074. [Google Scholar] [CrossRef]
- Dome, P.; Rihmer, Z.; Gonda, X. Suicide Risk in Bipolar Disorder: A Brief Review. Medicina 2019, 55, 403. [Google Scholar] [CrossRef] [Green Version]
- Hammen, C.L.; Gotlib, I.H. Handbook of Depression; Guilford Press: New York, NY, USA, 2002. [Google Scholar]
- Miller, J.N.; Black, D.W. Bipolar Disorder and Suicide: A Review. Curr. Psychiatry Rep. 2020, 22, 6. [Google Scholar] [CrossRef] [PubMed]
- Roy, B.; Yoshino, Y.; Allen, L.; Prall, K.; Schell, G.; Dwivedi, Y. Exploiting Circulating MicroRNAs as Biomarkers in Psychiatric Disorders. Mol. Diagn. Ther. 2020, 24, 279–298. [Google Scholar] [CrossRef]
- Roy, B.; Dunbar, M.; Shelton, R.C.; Dwivedi, Y. Identification of MicroRNA-124-3p as a Putative Epigenetic Signature of Major Depressive Disorder. Neuropsychopharmacology 2017, 42, 864–875. [Google Scholar] [CrossRef] [Green Version]
- Lopez, J.P.; Lim, R.; Cruceanu, C.; Crapper, L.; Fasano, C.; Labonte, B.; Turecki, G. miR-1202 is a primate-specific and brain-enriched microRNA involved in major depression and antide-pressant treatment. Nat. Med. 2014, 20, 764–768. [Google Scholar] [CrossRef]
- Issler, O.; Haramati, S.; Paul, E.D.; Maeno, H.; Navon, I.; Zwang, R.; Gil, S.; Mayberg, H.S.; Dunlop, B.W.; Menke, A.; et al. MicroRNA 135 Is Essential for Chronic Stress Resiliency, Antidepressant Efficacy, and Intact Serotonergic Activity. Neuron 2014, 83, 344–360. [Google Scholar] [CrossRef] [Green Version]
- Geaghan, M.; Cairns, M.J. MicroRNA and Posttranscriptional Dysregulation in Psychiatry. Biol. Psychiatry 2015, 78, 231–239. [Google Scholar] [CrossRef] [Green Version]
- Catalanotto, C.; Cogoni, C.; Zardo, G. MicroRNA in Control of Gene Expression: An Overview of Nuclear Functions. Int. J. Mol. Sci. 2016, 17, 1712. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Maffioletti, E.; Tardito, D.; Gennarelli, M.; Chiavetto, L.B. Micro spies from the brain to the periphery: New clues from studies on microRNAs in neuropsychiatric disorders. Front. Cell Neurosci. 2014, 8, 75. [Google Scholar] [CrossRef] [Green Version]
- Ramocki, M.B.; Zoghbi, H.Y. Failure of neuronal homeostasis results in common neuropsychiatric phenotypes. Nature 2008, 455, 912–918. [Google Scholar] [CrossRef] [Green Version]
- Schell, G.; Roy, B.; Prall, K.; Dwivedi, Y. miR-218: A Stress-Responsive Epigenetic Modifier. Non-Coding RNA 2022, 8, 55. [Google Scholar] [CrossRef]
- Gosline, S.J.; Gurtan, A.M.; JnBaptiste, C.K.; Bosson, A.; Milani, P.; Dalin, S.; Matthews, B.J.; Yap, Y.S.; Sharp, P.A.; Fraenkel, E. Elucidating MicroRNA Regulatory Networks Using Transcriptional, Post-transcriptional, and Histone Modification Measurements. Cell Rep. 2015, 14, 310–319. [Google Scholar] [CrossRef] [Green Version]
- Follert, P.; Cremer, H.; Béclin, C. MicroRNAs in brain development and function: A matter of flexibility and stability. Front. Mol. Neurosci. 2014, 7, 5. [Google Scholar] [CrossRef] [Green Version]
- Roy, B.; Wang, Q.; Palkovits, M.; Faludi, G.; Dwivedi, Y. Altered miRNA expression network in locus coeruleus of depressed suicide subjects. Sci. Rep. 2017, 7, 4387. [Google Scholar] [CrossRef] [Green Version]
- Issler, O.; Chen, A. Determining the role of microRNAs in psychiatric disorders. Nat. Rev. Neurosci. 2015, 16, 201–212. [Google Scholar] [CrossRef]
- Yoshino, Y.; Dwivedi, Y. Non-Coding RNAs in Psychiatric Disorders and Suicidal Behavior. Front. Psychiatry 2020, 11, 543893. [Google Scholar] [CrossRef]
- Wiktorowicz, J.E.; Soman, K.V. Discovery of Candidate Biomarkers. Adv. Exp. Med. Biol. 2016, 919, 443–462. [Google Scholar] [CrossRef]
- Peplow, P.V.; Martinez, B. MicroRNAs in blood and cerebrospinal fluid as diagnostic biomarkers of multiple sclerosis and to monitor disease progression. Neural Regen. Res. 2020, 15, 606–619. [Google Scholar] [CrossRef]
- Fries, G.R.; Quevedo, J. Exosomal MicroRNAs as Potential Biomarkers in Neuropsychiatric Disorders. Methods Mol. Biol. 2018, 1733, 79–85. [Google Scholar] [CrossRef] [PubMed]
- Turchinovich, A.; Burwinkel, B. Distinct AGO1 and AGO2 associated miRNA profiles in human cells and blood plasma. RNA Biol. 2012, 9, 1066–1075. [Google Scholar] [CrossRef] [Green Version]
- Rodriguez, A.; Griffiths-Jones, S.; Ashurst, J.L.; Bradley, A. Identification of Mammalian microRNA Host Genes and Transcription Units. Genome Res. 2004, 14, 1902–1910. [Google Scholar] [CrossRef] [Green Version]
- Ha, M.; Kim, V.N. Regulation of microRNA biogenesis. Nat. Rev. Mol. Cell Biol. 2014, 15, 509–524. [Google Scholar] [CrossRef]
- Winter, J.; Jung, S.; Keller, S.; Gregory, R.I.; Diederichs, S. Many roads to maturity: microRNA biogenesis pathways and their regulation. Nat. Cell Biol. 2009, 11, 228–234. [Google Scholar] [CrossRef]
- Daugaard, I.; Hansen, T.B. Biogenesis and Function of Ago-Associated RNAs. Trends Genet. 2017, 33, 208–219. [Google Scholar] [CrossRef]
- Bartel, D.P. MicroRNAs: Target Recognition and Regulatory Functions. Cell 2009, 136, 215–233. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- O’Connor, R.M.; Gururajan, A.; Dinan, T.G.; Kenny, P.J.; Cryan, J.F. All Roads Lead to the miRNome: miRNAs Have a Central Role in the Molecular Pathophysiology of Psychiatric Disorders. Trends Pharmacol. Sci. 2016, 37, 1029–1044. [Google Scholar] [CrossRef] [PubMed]
- Kuehner, J.N.; Bruggeman, E.C.; Wen, Z.; Yao, B. Epigenetic Regulations in Neuropsychiatric Disorders. Front. Genet. 2019, 10, 268. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Deraredj Nadim, W.; Simion, V.; Benedetti, H.; Pichon, C.; Baril, P.; Morisset-Lopez, S. MicroRNAs in Neurocognitive Dysfunctions: New Molecular Targets for Pharmacological Treatments? Curr. Neuropharmacol. 2017, 15, 260–275. [Google Scholar] [CrossRef] [Green Version]
- Forstner, A.J.; Hofmann, A.; Maaser, A.; Sumer, S.; Khudayberdiev, S.; Mühleisen, T.W.; Leber, M.; Schulze, T.G.; Strohmaier, J.; Degenhardt, F.; et al. Genome-wide analysis implicates microRNAs and their target genes in the development of bipolar disorder. Transl. Psychiatry 2015, 5, e678. [Google Scholar] [CrossRef] [Green Version]
- Pittenger, C.; Duman, R.S. Stress, Depression, and Neuroplasticity: A Convergence of Mechanisms. Neuropsychopharmacology 2008, 33, 88–109. [Google Scholar] [CrossRef] [PubMed]
- Dwivedi, Y. Emerging role of microRNAs in major depressive disorder: Diagnosis and therapeutic implications. Dialog-Clin. Neurosci. 2014, 16, 43–61. [Google Scholar] [CrossRef] [PubMed]
- Azevedo, J.A.; Carter, B.S.; Meng, F.; Turner, D.L.; Dai, M.; Schatzberg, A.F.; Barchas, J.D.; Jones, E.G.; Bunney, W.E.; Myers, R.M.; et al. The microRNA network is altered in anterior cingulate cortex of patients with unipolar and bipolar depression. J. Psychiatr. Res. 2016, 82, 58–67. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yoshino, Y.; Roy, B.; Dwivedi, Y. Differential and unique patterns of synaptic miRNA expression in dorsolateral prefrontal cortex of depressed subjects. Neuropsychopharmacology 2021, 46, 900–910. [Google Scholar] [CrossRef]
- Yoshino, Y.; Roy, B.; Dwivedi, Y. Altered miRNA landscape of the anterior cingulate cortex is associated with potential loss of key neuronal functions in depressed brain. Eur. Neuropsychopharmacol. 2020, 40, 70–84. [Google Scholar] [CrossRef] [PubMed]
- Roy, B.; Dunbar, M.; Agrawal, J.; Allen, L.; Dwivedi, Y. Amygdala-Based Altered miRNome and Epigenetic Contribution of miR-128-3p in Conferring Susceptibility to Depression-Like Behavior via Wnt Signaling. Int. J. Neuropsychopharmacol. 2020, 23, 165–177. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Smalheiser, N.R.; Lugli, G.; Rizavi, H.S.; Torvik, V.I.; Turecki, G.; Dwivedi, Y. MicroRNA Expression Is Down-Regulated and Reorganized in Prefrontal Cortex of Depressed Suicide Subjects. PLoS ONE 2012, 7, e33201. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Smalheiser, N.R.; Lugli, G.; Zhang, H.; Rizavi, H.; Cook, E.H.; Dwivedi, Y. Expression of microRNAs and Other Small RNAs in Prefrontal Cortex in Schizophrenia, Bipolar Disorder and Depressed Subjects. PLoS ONE 2014, 9, e86469. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Maffioletti, E.; Cattaneo, A.; Rosso, G.; Maina, G.; Maj, C.; Gennarelli, M.; Tardito, D.; Bocchio-Chiavetto, L. Peripheral whole blood microRNA alterations in major depression and bipolar disorder. J. Affect. Disord. 2016, 200, 250–258. [Google Scholar] [CrossRef] [Green Version]
- Fan, H.-M.; Sun, X.-Y.; Guo, W.; Zhong, A.-F.; Niu, W.; Zhao, L.; Dai, Y.-H.; Guo, Z.-M.; Zhang, L.-Y.; Lu, J. Differential expression of microRNA in peripheral blood mononuclear cells as specific biomarker for major depressive disorder patients. J. Psychiatr. Res. 2014, 59, 45–52. [Google Scholar] [CrossRef] [PubMed]
- Camkurt, M.A.; Coşkun, S.A.; Güneş, M.; Güneş, S.; Yılmaz, M.F.; Görür, A.; Tamer, L. Comparison of plasma MicroRNA levels in drug naive, first episode depressed patients and healthy controls. J. Psychiatr. Res. 2015, 69, 67–71. [Google Scholar] [CrossRef] [PubMed]
- He, S.; Liu, X.; Jiang, K.; Peng, D.; Hong, W.; Fang, Y.; Qian, Y.; Yu, S.; Li, H. Alterations of microRNA-124 expression in peripheral blood mononuclear cells in pre- and post-treatment patients with major depressive disorder. J. Psychiatr. Res. 2016, 78, 65–71. [Google Scholar] [CrossRef] [PubMed]
- Feng, J.; Wang, M.; Li, M.; Yang, J.; Jia, J.; Liu, L.; Zhou, J.; Zhang, C.; Wang, X. Serum miR-221-3p as a new potential biomarker for depressed mood in perioperative patients. Brain Res. 2019, 1720, 146296. [Google Scholar] [CrossRef]
- Wan, Y.; Liu, Y.; Wang, X.; Wu, J.; Liu, K.; Zhou, J.; Liu, L.; Zhang, C. Identification of Differential MicroRNAs in Cerebrospinal Fluid and Serum of Patients with Major Depressive Disorder. PLoS ONE 2015, 10, e0121975. [Google Scholar] [CrossRef]
- Song, M.-F.; Dong, J.-Z.; Wang, Y.-W.; He, J.; Ju, X.; Zhang, L.; Zhang, Y.-H.; Shi, J.-F.; Lv, Y.-Y. CSF miR-16 is decreased in major depression patients and its neutralization in rats induces depression-like behaviors via a serotonin transmitter system. J. Affect. Disord. 2015, 178, 25–31. [Google Scholar] [CrossRef]
- Sun, N.; Lei, L.; Wang, Y.; Yang, C.; Liu, Z.; Li, X.; Zhang, K. Preliminary comparison of plasma notch-associated microRNA-34b and -34c levels in drug naive, first episode depressed patients and healthy controls. J. Affect. Disord. 2016, 194, 109–114. [Google Scholar] [CrossRef]
- He, C.; Bai, Y.; Wang, Z.; Fan, D.; Wang, Q.; Liu, X.; Zhang, H.; Zhang, H.; Zhang, Z.; Yao, H.; et al. Identification of microRNA-9 linking the effects of childhood maltreatment on depression using amygdala connectivity. Neuroimage 2021, 224, 117428. [Google Scholar] [CrossRef]
- Gecys, D.; Dambrauskiene, K.; Simonyte, S.; Patamsyte, V.; Vilkeviciute, A.; Musneckis, A.; Adomaitiene, V. Circulating hsa-let-7e-5p and hsa-miR-125a-5p as Possible Biomarkers in the Diagnosis of Major Depression and Bipolar Disorders. Dis. Markers 2022, 2022, 3004338. [Google Scholar] [CrossRef]
- Hung, Y.-Y.; Wu, M.-K.; Tsai, M.-C.; Huang, Y.-L.; Kang, H.-Y. Aberrant Expression of Intracellular let-7e, miR-146a, and miR-155 Correlates with Severity of Depression in Patients with Major Depressive Disorder and Is Ameliorated after Antidepressant Treatment. Cells 2019, 8, 647. [Google Scholar] [CrossRef] [Green Version]
- Li, Q.S.; Galbraith, D.; Morrison, R.L.; Trivedi, M.H.; Drevets, W.C. Circulating microRNA associated with future relapse status in major depressive disorder. Front. Psychiatry 2022, 13, 937360. [Google Scholar] [CrossRef]
- Roumans, S.; Sundquist, K.; Memon, A.A.; Hedelius, A.; Sundquist, J.; Wang, X. Association of circulating let-7b-5p with major depressive disorder: A nested case-control study. BMC Psychiatry 2021, 21, 616. [Google Scholar] [CrossRef] [PubMed]
- Gururajan, A.; Naughton, M.E.; Scott, K.A.; O’Connor, R.M.; Moloney, G.; Clarke, G.; Dowling, J.; Walsh, A.; Ismail, F.; Shorten, G.; et al. MicroRNAs as biomarkers for major depression: A role for let-7b and let-7c. Transl. Psychiatry 2016, 6, e862. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, H.-P.; Liu, X.-L.; Chen, J.-J.; Cheng, K.; Bai, S.-J.; Zheng, P.; Zhou, C.-J.; Wang, W.; Wang, H.-Y.; Zhong, L.-M.; et al. Circulating microRNA 134 sheds light on the diagnosis of major depressive disorder. Transl. Psychiatry 2020, 10, 95. [Google Scholar] [CrossRef] [Green Version]
- van der Zee, Y.Y.; Eijssen, L.M.; Mews, P.; Ramakrishnan, A.; Alvarez, K.; Lardner, C.K.; Issler, O. Blood miR-144-3p: A novel diagnostic and therapeutic tool for depression. Mol. Psychiatry 2022, 27, 4536–4549. [Google Scholar] [CrossRef] [PubMed]
- Bocchio-Chiavetto, L.; Maffioletti, E.; Bettinsoli, P.; Giovannini, C.; Bignotti, S.; Tardito, D.; Corrada, D.; Milanesi, L.; Gennarelli, M. Blood microRNA changes in depressed patients during antidepressant treatment. Eur. Neuropsychopharmacol. 2013, 23, 602–611. [Google Scholar] [CrossRef]
- Lopez, J.P.; Fiori, L.M.; Cruceanu, C.; Lin, R.; Labonte, B.; Cates, H.M.; Heller, E.A.; Vialou, V.; Ku, S.M.; Gerald, C.; et al. MicroRNAs 146a/b-5 and 425-3p and 24-3p are markers of antidepressant response and regulate MAPK/Wnt-system genes. Nat. Commun. 2017, 8, 15497. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, J.; Meng, H.; Cao, W.; Qiu, T. MiR-335 is involved in major depression disorder and antidepressant treatment through targeting GRM4. Neurosci. Lett. 2015, 606, 167–172. [Google Scholar] [CrossRef]
- Enatescu, V.R.; Papava, I.; Enatescu, I.; Antonescu, M.; Anghel, A.; Seclaman, E.; Marian, C. Circulating Plasma Micro RNAs in Patients with Major Depressive Disorder Treated with Antidepres-sants: A Pilot Study. Psychiatry Investig. 2016, 13, 549–557. [Google Scholar] [CrossRef] [Green Version]
- Fang, Y.; Qiu, Q.; Zhang, S.; Sun, L.; Li, G.; Xiao, S.; Li, X. Changes in miRNA-132 and miR-124 levels in non-treated and citalopram-treated patients with depression. J. Affect. Disord. 2018, 227, 745–751. [Google Scholar] [CrossRef]
- Lin, C.-C.; Tsai, M.-C.; Lee, C.-T.; Sun, M.-H.; Huang, T.-L. Antidepressant treatment increased serum miR-183 and miR-212 levels in patients with major depressive disorder. Psychiatry Res. 2018, 270, 232–237. [Google Scholar] [CrossRef]
- Wei, Z.-X.; Xie, G.-J.; Mao, X.; Zou, X.-P.; Liao, Y.-J.; Liu, Q.-S.; Wang, H.; Cheng, Y. Exosomes from patients with major depression cause depressive-like behaviors in mice with involvement of miR-139-5p-regulated neurogenesis. Neuropsychopharmacology 2020, 45, 1050–1058. [Google Scholar] [CrossRef] [PubMed]
- Liang, J.-Q.; Liao, H.-R.; Xu, C.-X.; Li, X.-L.; Wei, Z.-X.; Xie, G.-J.; Cheng, Y. Serum Exosome-Derived miR-139-5p as a Potential Biomarker for Major Depressive Disorder. Neuropsychiatr. Dis. Treat. 2020, 16, 2689–2693. [Google Scholar] [CrossRef] [PubMed]
- Hung, Y.-Y.; Chou, C.-K.; Yang, Y.-C.; Fu, H.-C.; Loh, E.-W.; Kang, H.-Y. Exosomal let-7e, miR-21-5p, miR-145, miR-146a and miR-155 in Predicting Antidepressants Response in Patients with Major Depressive Disorder. Biomedicines 2021, 9, 1428. [Google Scholar] [CrossRef] [PubMed]
- Saeedi, S.; Nagy, C.; Ibrahim, P.; Théroux, J.-F.; Wakid, M.; Fiori, L.M.; Yang, J.; Rotzinger, S.; Foster, J.A.; Mechawar, N.; et al. Neuron-derived extracellular vesicles enriched from plasma show altered size and miRNA cargo as a function of antidepressant drug response. Mol. Psychiatry 2021, 26, 7417–7424. [Google Scholar] [CrossRef]
- Ceylan, D.; Tufekci, K.U.; Keskinoglu, P.; Genc, S.; Özerdem, A. Circulating exosomal microRNAs in bipolar disorder. J. Affect. Disord. 2019, 262, 99–107. [Google Scholar] [CrossRef]
- Fries, G.R.; Lima, C.N.; Valvassori, S.S.; Zunta-Soares, G.; Soares, J.C.; Quevedo, J. Preliminary investigation of peripheral extracellular vesicles’ microRNAs in bipolar disorder. J. Affect. Disord. 2019, 255, 10–14. [Google Scholar] [CrossRef]
- Tekdemir, R.; Selvi, Y.; Altınbaş, K.; Koçak, N. Decreased miR-15b-5p/miR-155-5p levels and increased miR-134-5p/miR-652-3p levels among BD patients under lithium treatment. J. Affect. Disord. 2022, 317, 6–14. [Google Scholar] [CrossRef]
- Tabano, S.; Caldiroli, A.; Terrasi, A.; Colapietro, P.; Grassi, S.; Carnevali, G.S.; Fontana, L.; Serati, M.; Vaira, V.; Altamura, A.C.; et al. A miRNome analysis of drug-free manic psychotic bipolar patients versus healthy controls. Eur. Arch. Psychiatry Clin. Neurosci. 2020, 270, 893–900. [Google Scholar] [CrossRef]
- Lee, S.-Y.; Lu, R.-B.; Wang, L.-J.; Chang, C.-H.; Lu, T.; Wang, T.-Y.; Tsai, K.-W. Serum miRNA as a possible biomarker in the diagnosis of bipolar II disorder. Sci. Rep. 2020, 10, 1131. [Google Scholar] [CrossRef] [Green Version]
- Tekin, S.S.; Erdal, M.E.; Asoğlu, M.; Ay, Ö.İ.; Ay, M.E.; Yılmaz, Ş.G. Biomarker potential of hsa-miR-145-5p in peripheral whole blood of manic bipolar I patients. Braz. J. Psychiatry 2022, 40, 378–387. [Google Scholar] [CrossRef]
- Martins, H.C.; Gilardi, C.; Sungur A, Ö.; Winterer, J.; Pelzl, M.A.; Bicker, S.; Schratt, G. Bipolar-associated miR-499-5p controls neuroplasticity by downregulating the Cav1.2 subunit CACNB2. EMBO Rep. 2022, 23, e54420. [Google Scholar] [CrossRef] [PubMed]
- Camkurt, M.A.; Karababa, I.F.; Erdal, M.E.; Kandemir, S.B.; Fries, G.R.; Bayazıt, H.; Ay, M.E.; Kandemir, H.; Ay, I.; Coşkun, S.; et al. MicroRNA dysregulation in manic and euthymic patients with bipolar disorder. J. Affect. Disord. 2020, 261, 84–90. [Google Scholar] [CrossRef] [PubMed]
- Lim, C.H.; Zainal, N.Z.; Kanagasundram, S.; Zain, S.M.; Mohamed, Z. Preliminary examination of microRNA expression profiling in bipolar disorder I patients during antipsy-chotic treatment. Am. J. Med. Genet. B Neuropsychiatr. Genet. 2016, 171, 867–874. [Google Scholar] [CrossRef]
- Chen, Y.; Shi, J.; Liu, H.; Wang, Q.; Chen, X.; Tang, H.; Yan, R.; Yao, Z.; Lu, Q. Plasma microRNA Array Analysis Identifies Overexpressed miR-19b-3p as a Biomarker of Bipolar Depression Distinguishing From Unipolar Depression. Front. Psychiatry 2020, 11, 757. [Google Scholar] [CrossRef] [PubMed]
- Lee, S.-Y.; Wang, T.-Y.; Lu, R.-B.; Wang, L.-J.; Chang, C.-H.; Chiang, Y.-C.; Tsai, K.-W. Peripheral BDNF correlated with miRNA in BD-II patients. J. Psychiatr. Res. 2020, 136, 184–189. [Google Scholar] [CrossRef] [PubMed]
- Walker, R.M.; Rybka, J.; Anderson, S.M.; Torrance, H.S.; Boxall, R.; Sussmann, J.E.; Porteous, D.J.; McIntosh, A.M.; Evans, K.L. Preliminary investigation of miRNA expression in individuals at high familial risk of bipolar disorder. J. Psychiatr. Res. 2015, 62, 48–55. [Google Scholar] [CrossRef] [Green Version]
- Kosten, T.; Ye, A.; Nielsen, D.; Patriquin, M.; Palat, A.; Rezaei, H.; Salas, R.; Harding, M.; Haile, C.; Gunaratne, P.; et al. Specific miRNAs associated with treatment response during hospitalization for suicidal ideation. Res. Sq. 2022. [Google Scholar] [CrossRef]
- Paska, A.V.; Alič, U.; Zupanc, T.; Kouter, K. Suicide and Changes in Expression of Neuronal miRNA Predicted by an Algorithm Search through miRNA Databases. Genes 2022, 13, 562. [Google Scholar] [CrossRef]
- Wang, Q.; Roy, B.; Turecki, G.; Shelton, R.C.; Dwivedi, Y. Role of Complex Epigenetic Switching in Tumor Necrosis Factor-α Upregulation in the Prefrontal Cortex of Suicide Subjects. Am. J. Psychiatry 2018, 175, 262–274. [Google Scholar] [CrossRef] [Green Version]
- Belzeaux, R.; Fiori, L.M.; Lopez, J.P.; Boucekine, M.; Boyer, L.; Blier, P.; Farzan, F.; Frey, B.N.; Giacobbe, P.; Lam, R.W.; et al. Predicting Worsening Suicidal Ideation With Clinical Features and Peripheral Expression of Messenger RNA and MicroRNA During Antidepressant Treatment. J. Clin. Psychiatry 2019, 80, 22559. [Google Scholar] [CrossRef]
- Zhao, L.; Yang, X.; Cui, L.; Wei, J.; Ni, P.; Li, M.; Wang, Y.; He, Y.; Li, X.; Liang, S.; et al. Increased expression of a novel miRNA in peripheral blood is negatively correlated with hippocampal volume in patients with major depressive disorder. J. Affect. Disord. 2019, 245, 205–212. [Google Scholar] [CrossRef] [PubMed]
- Clausen, A.R.; Durand, S.; Petersen, R.L.; Staunstrup, N.H.; Qvist, P. Circulating miRNAs as Potential Biomarkers for Patient Stratification in Bipolar Disorder: A Combined Review and Data Mining Approach. Genes 2022, 13, 1038. [Google Scholar] [CrossRef] [PubMed]
- Rong, H.; Liu, T.B.; Yang, K.J.; Yang, H.C.; Wu, D.H.; Liao, C.P.; Hong, F.; Yang, H.Z.; Wan, F.; Ye, X.Y.; et al. MicroRNA-134 plasma levels before and after treatment for bipolar mania. J. Psychiatr. Res. 2011, 45, 92–95. [Google Scholar] [CrossRef]
- Kouter, K.; Paska, A.V. Biomarkers for suicidal behavior: miRNAs and their potential for diagnostics through liquid biopsy—A systematic review. Epigenomics 2020, 12, 2219–2235. [Google Scholar] [CrossRef]
- Baldessarini, R.; Tondo, L. Suicidal Risks in 12 DSM-5 Psychiatric Disorders. J. Affect. Disord. 2020, 271, 66–73. [Google Scholar] [CrossRef]
- Serra, G.; De Crescenzo, F.; Maisto, F.; Galante, J.R.; Iannoni, M.E.; Trasolini, M.; Maglio, G.; Tondo, L.; Baldessarini, R.J.; Vicari, S. Suicidal behavior in juvenile bipolar disorder and major depressive disorder patients: Systematic review and meta-analysis. J. Affect. Disord. 2022, 311, 572–581. [Google Scholar] [CrossRef]
- Bosch, M.; Meyer-Lindenberg, A. Environmental Exposures and Depression: Biological Mechanisms and Epidemiological Evi-dence. Annu. Rev. Public Health 2019, 40, 239–259. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Matsuda, S.; Ikeda, Y.; Murakami, M.; Nakagawa, Y.; Tsuji, A.; Kitagishi, Y. Roles of PI3K/AKT/GSK3 Pathway Involved in Psychiatric Illnesses. Diseases 2019, 7, 22. [Google Scholar] [CrossRef] [Green Version]
- Duman, R.S.; Voleti, B. Signaling pathways underlying the pathophysiology and treatment of depression: Novel mecha-nisms for rapid-acting agents. Trends Neurosci. 2012, 35, 47–56. [Google Scholar] [CrossRef] [Green Version]
- Ludtmann, M.H.; Boeckeler, K.; Williams, R.S. Molecular pharmacology in a simple model system: Implicating MAP kinase and phosphoinositide signalling in bipolar disorder. Semin. Cell Dev. Biol. 2011, 22, 105–113. [Google Scholar] [CrossRef] [Green Version]
- Van den Berg, M.M.J.; Krauskopf, J.; Ramaekers, J.G.; Kleinjans, J.C.S.; Prickaerts, J.; Briedé, J.J. Circulating microRNAs as potential biomarkers for psychiatric and neurodegenerative disorders. Prog. Neurobiol. 2019, 185, 101732. [Google Scholar] [CrossRef] [PubMed]
- Witwer, K.W. Circulating MicroRNA Biomarker Studies: Pitfalls and Potential Solutions. Clin. Chem. 2015, 61, 56–63. [Google Scholar] [CrossRef] [PubMed]
- Xia, X.; Wang, Y.; Huang, Y.; Zhang, H.; Lu, H.; Zheng, J.C. Exosomal miRNAs in central nervous system diseases: Biomarkers, pathological mediators, protective factors and therapeutic agents. Prog. Neurobiol. 2019, 183, 101694. [Google Scholar] [CrossRef] [PubMed]
- Mayr, M.; Zampetaki, A. Analytical challenges and technical limitations in assessing circulating MiRNAs. Thromb. Haemost. 2012, 108, 592–598. [Google Scholar] [CrossRef] [Green Version]
- Kok, M.; de Ronde, M.; Moerland, P.; Ruijter, J.; Creemers, E.; Pinto-Sietsma, S. Small sample sizes in high-throughput miRNA screens: A common pitfall for the identification of miRNA biomarkers. Biomol. Detect. Quantif. 2018, 15, 1–5. [Google Scholar] [CrossRef]
- Saeedi, S.; Israel, S.; Nagy, C.; Turecki, G. The emerging role of exosomes in mental disorders. Transl. Psychiatry 2019, 9, 122. [Google Scholar] [CrossRef] [Green Version]
- Agliardi, C.; Clerici, M. Blood extracellular vesicles (EVs) of central nervous system origin: A window into the brain. Neural Regen. Res. 2020, 15, 55–56. [Google Scholar] [CrossRef]
- Torres-Berrío, A.; Nouel, D.; Cuesta, S.; Parise, E.M.; Restrepo-Lozano, J.M.; Larochelle, P.; Nestler, E.J.; Flores, C. MiR-218: A molecular switch and potential biomarker of susceptibility to stress. Mol. Psychiatry 2020, 25, 951–964. [Google Scholar] [CrossRef]
- Torres-Berrío, A.; Morgunova, A.; Giroux, M.; Cuesta, S.; Nestler, E.J.; Flores, C. miR-218 in Adolescence Predicts and Mediates Vulnerability to Stress. Biol. Psychiatry 2021, 89, 911–919. [Google Scholar] [CrossRef]
- Ochi, S.; Dwivedi, Y. Dissecting early life stress-induced adolescent depression through epigenomic approach. Mol. Psychiatry 2023, 28, 141–153. [Google Scholar] [CrossRef]
- Arnow, B.A.; Blasey, C.; Williams, L.M.; Palmer, D.M.; Rekshan, W.; Schatzberg, A.F.; Rush, A. J Depression Subtypes in Predicting Antidepressant Response: A Report From the iSPOT-D Trial. Am. J. Psychiatry 2015, 172, 743–750. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- McIntyre, R.S.; Alda, M.; Baldessarini, R.J.; Bauer, M.; Berk, M.; Correll, C.U.; Fagiolini, A.; Fountoulakis, K.; Frye, M.A.; Grunze, H.; et al. The clinical characterization of the adult patient with bipolar disorder aimed at personalization of management. World Psychiatry 2022, 21, 364–387. [Google Scholar] [CrossRef] [PubMed]
- Jimenez-Treviño, L.; Blasco-Fontecilla, H.; Braquehais, M.D.; Ceverino-Dominguez, A.; Baca-Garcia, E. Endophenotypes and suicide behaviour. Actas Espanolas Psiquiatr. 2011, 39, 61–69. [Google Scholar]
- Bi, B.; Liu, W.; Zhou, D.; Fu, X.; Qin, X.; Wu, J. Personality traits and suicide attempts with and without psychiatric disorders: Analysis of impulsivity and neuroticism. BMC Psychiatry 2017, 17, 294. [Google Scholar] [CrossRef] [Green Version]
- Pompili, M.; Serafini, G.; Innamorati, M.; Dominici, G.; Ferracuti, S.; Kotzalidis, G.D.; Serra, G.; Girardi, P.; Janiri, L.; Tatarelli, R.; et al. Suicidal Behavior and Alcohol Abuse. Int. J. Environ. Res. Public Health 2010, 7, 1392–1431. [Google Scholar] [CrossRef] [Green Version]
- Stapel, B.; Xiao, K.; Gorinski, N.; Schmidt, K.; Pfanne, A.; Fiedler, J.; Richter, I.; Vollbrecht, A.-L.; Thum, T.; Kahl, K.G.; et al. MicroRNAs as novel peripheral markers for suicidality in patients with major depressive disorder. Front. Psychiatry 2022, 13, 1020530. [Google Scholar] [CrossRef]




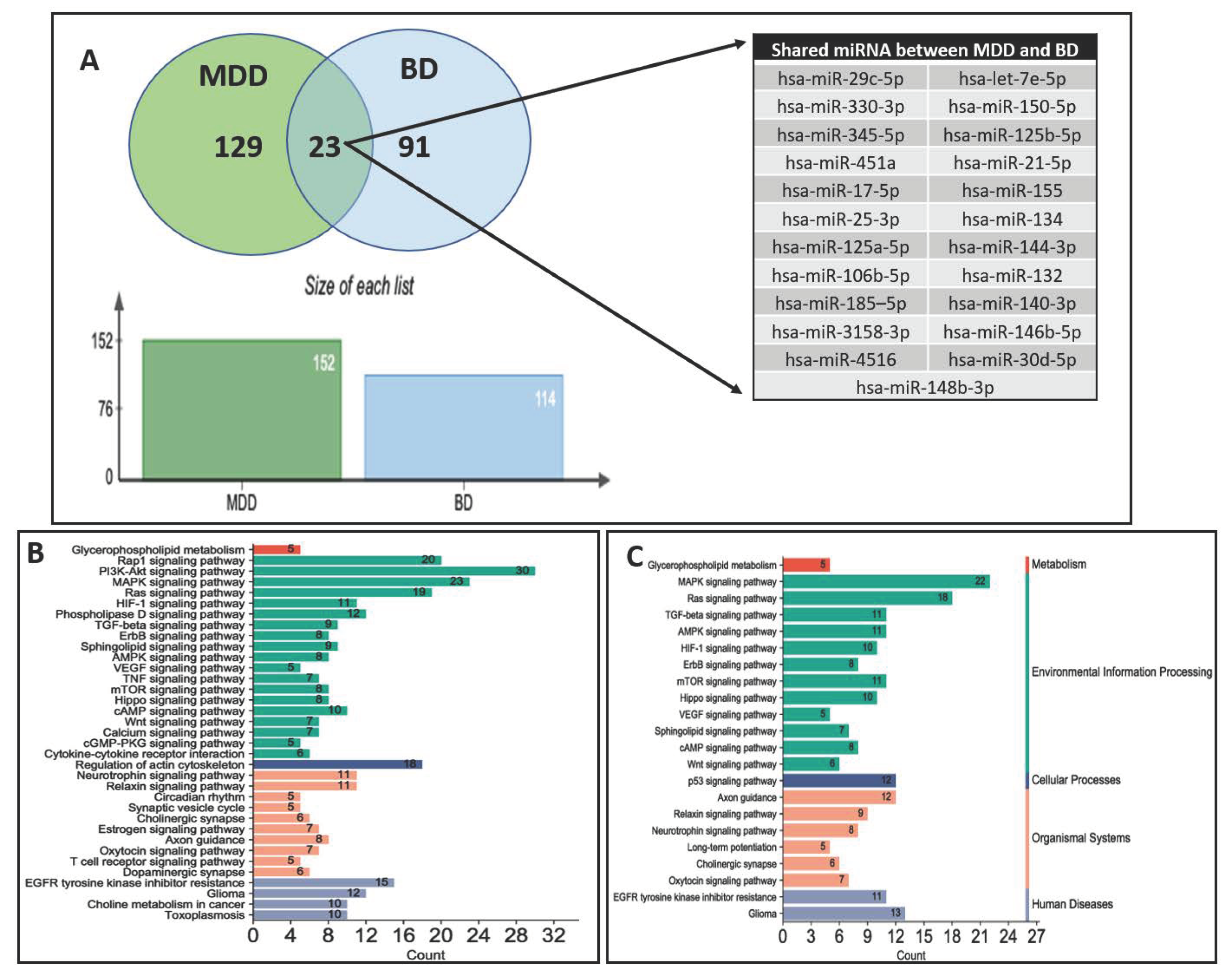{kind=link}
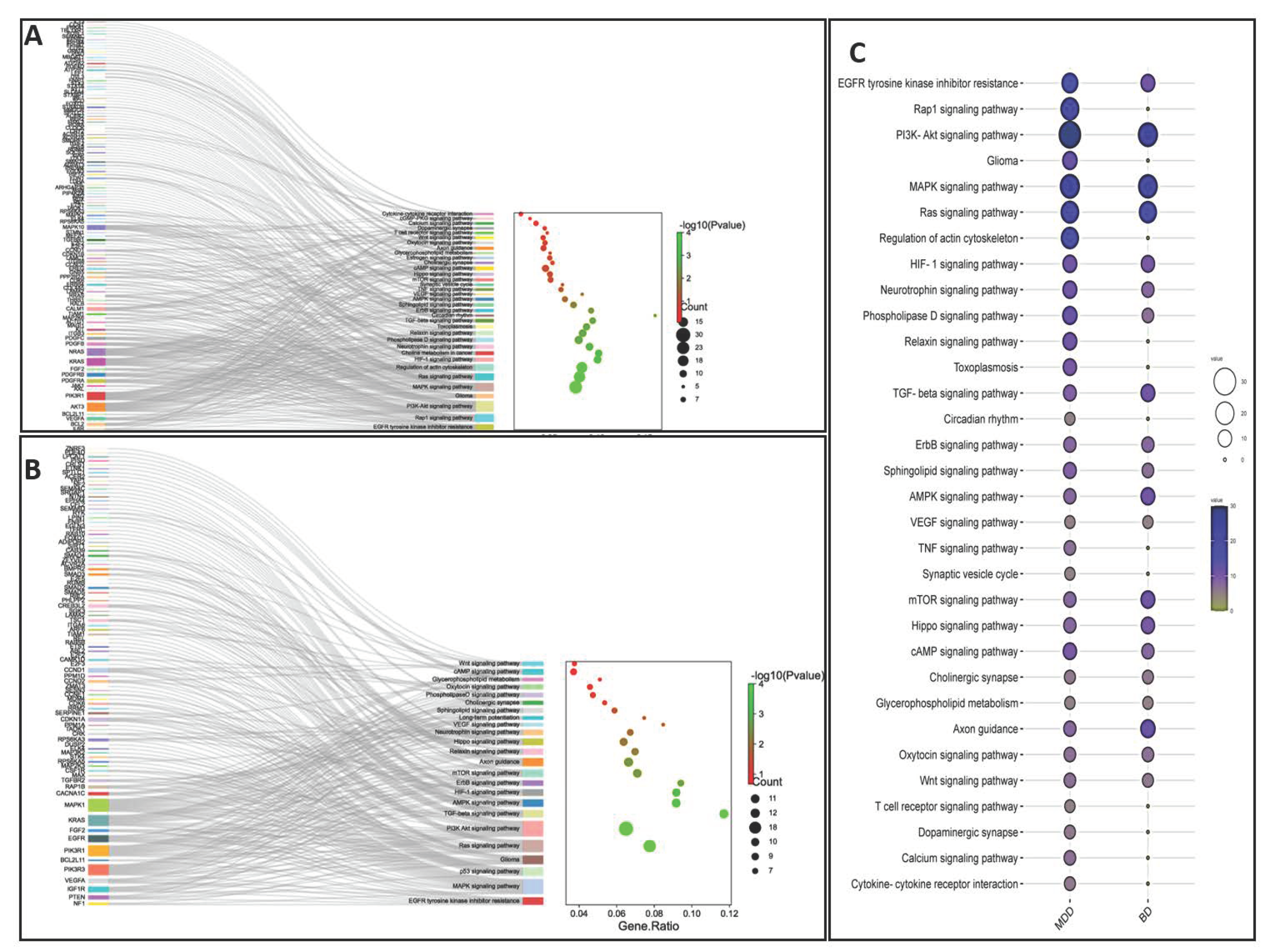{kind=link}
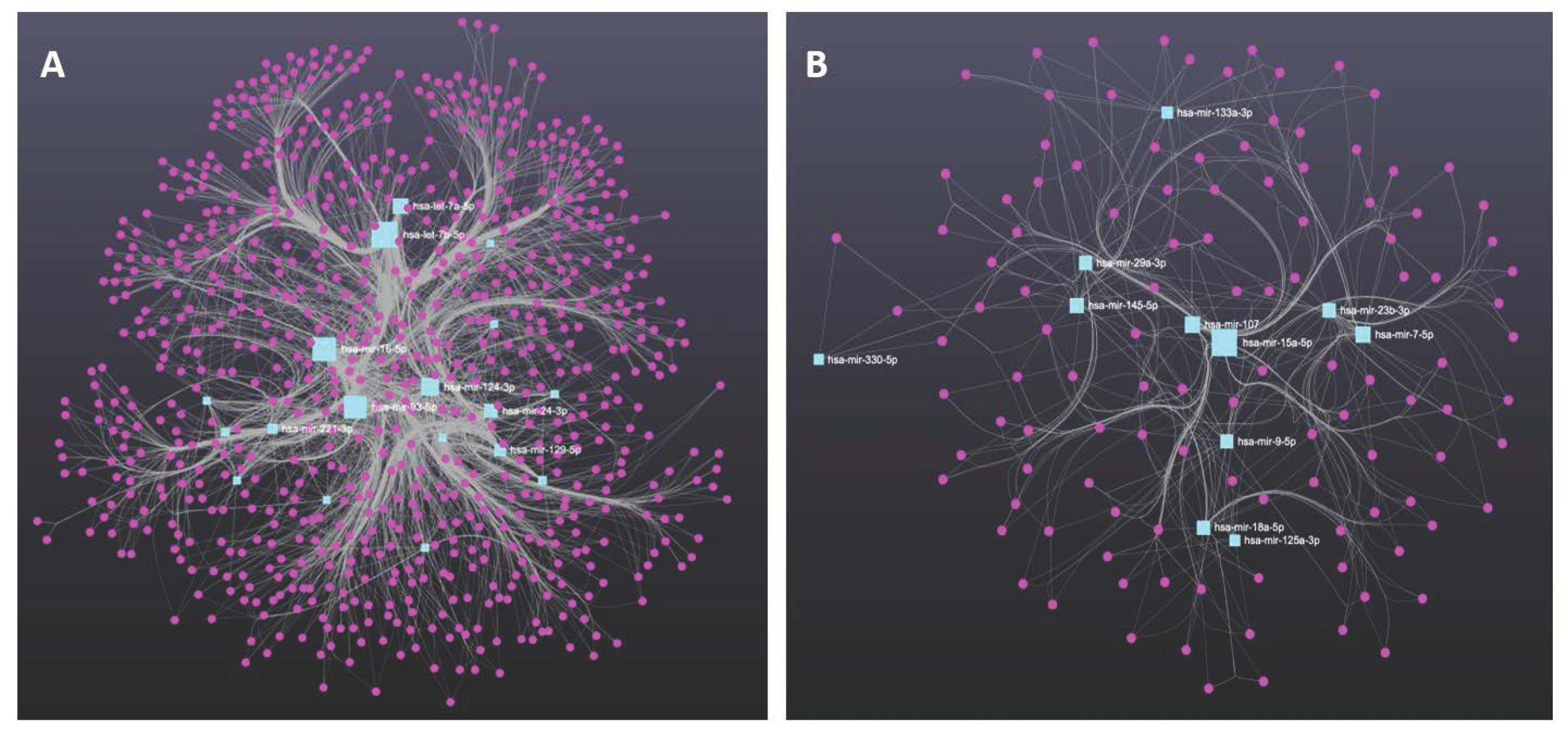{kind=link}
| Samples | MiRNA Expression Changes | Reference |
|---|---|---|
| Major Depressive Disorder | ||
| Peripheral blood | let-7d-5p, miR-1915-3p, miR-29c-5p, let-7f-5p, miR-330-3p, miR-425-3p, miR-24-3p, let-7a-5p, miR-199a-5p, miR-345-5p | [41] |
| Blood mononuclear cells | miR-26b, miR-29b, miR-146b, miR-1244, miR- 4485, miR-1972, miR-4498, miR-4743, miR-874, miR-338 | [42] |
| Plasma | miR-320a, miR-451a, miR-17-5p, miR-223-3p, miR-25-3p, miR-126-3p, miR-16-5p, miR-93-5p | [43] |
| Serum | miR-124-3p | [7] |
| Blood mononuclear cells | miR-124 | [44] |
| Serum | miR-221-3p | [45] |
| Cerebrospinal fluid and serum | miR-125a-5p, let-7d-3p, miR-30a-5p, miR-34a-5p, miR-221-3p, miR-29b-3p, miR-10a-5p, miR-375, miR-155–5p, miR-33a-5p, miR-139–5p, miR-451a, miR-15b-5p miR-106b-5p, miR-590–5p, miR-185–5p | [46] |
| Cerebrospinal fluid and blood | miR-16 | [47] |
| Peripheral blood leukocyte | miR-34b-5p and miR-34c-5p | [48] |
| Serum | miR-9 | [49] |
| Plasma | miR-483-5p, let-7f-5p, let-7e-5p, miR-122-5p, miR-125a-5p, miR-150-5p, miR-139-3p, miR-193a-5p, miR-125b-5p, miR-197-3p, miR-483-3p | [50] |
| Blood mononuclear cells | let-7e, miR-21-5p miR-145, miR-223, miR-146a, and miR-155 | [51] |
| Serum | miR-199b-5p, miR-215-5p, miR-200a-3p, miR-143-3p | [52] |
| Plasma | let-7b-5p | [53] |
| Peripheral blood | let-7b, let-7c | [54] |
| Plasma | miR-134 | [55] |
| Peripheral blood | miR-144-3p | [56] |
| Peripheral blood | miR-130, miR-505, miR-29b-2, miR-26b, miR-22, miR-26a, miR-664, miR-494, let-7d, let-7g, let-7e, miR-34c-5p, let-7f, miR-629, miR-106, miR-103, miR-191, miR-128, miR-502-3p, miR-374b, miR-132, miR-30d, miR-500, miR-770-5p, miR-589, miR-183, miR-574-3p, miR-140-3p, miR-335, miR-361-5p | [57] |
| Plasma | miR-425-3p, miR-24-3p, miR-503-5p, miR-146a-5p, miR-215-5p, miR-3074-5p, miR-1180-3p, miR-425-5p, miR-324-5p, miR-146b-5p, miR-6750-3p, miR-6511a-3p, miR-361-5p, miR-3173-5p, miR-2110, miR-3605-3p, miR-6881-3p, miR-30e-5p, miR-423-3p, miR-361-3p, miR-3184-5p, miR-636 | [58] |
| Peripheral blood | miR-335 | [59] |
| Plasma | miR-1193, miR-3173-3p, miR-3154, miR-129-5p, miR-3661, miR-1287, miR-532-3p, miR-2278, miR-3150a-3p, miR-3909, miR-151-5p, miR-99b, miR-937, miR-676, miR-223, miR-181b, miR-489, miR-637, miR-608, miR-26a | [60] |
| Plasma | miR-132, miR-124 | [61] |
| Whole blood | miR-183, miR-212 | [62] |
| Blood exosome | miR-139-5p | [63] |
| Serum exosome | miR-139-5p | [64] |
| Serum exosome | let-7e, miR-21-5p, miR-145, miR-146a, miR-155 | [65] |
| Plasma exosome, neuron-derived plasma exosomes | miR-423-3p, miR-191-5p, miR-486-5p, miR-30d-5p, miR-425-5p, miR-25-3p, miR-21-5p, miR-335-5p, miR-126-5p | [66] |
| Bipolar Disorder | ||
| Exosomes from plasma | miR-185-5p, miR-25-3p, miR-92a-3p, miR-376b-3p, let-7i-5p, miR-484, miR-652–3p, miR-142-3p, miR-30b-5p, miR-126–3p, miR-15a-5p, miR-126-5p, miR-301a-3p | [67] |
| Plasma | miR-21-5p, miR-22-3p, miR-29-c3p, miR-92a-3p, miR-142-3p, miR-1185-2-3p, miR-3135b, miR-3194-5p, miR-4516, miR-6090, miR-6791-5p, miR-6808-5p, miR-7975, miR-7977, miR-133a-3p, miR-188-5p, miR-451a, miR-671-5p, miR-1227-5p, miR-1238-3p, miR-1268b, miR-1281, miR-3620-5p, miR-4433a-5p, miR-5739, miR-6068, miR-6125, miR-6727-5p, miR-6775-5p, miR-6800-3p, miR-6821-5p, miR-7108-5p, miR-8060 | [68] |
| Plasma | miR-132, miR-134, miR-152, miR-607, miR-633, miR-652, miR-15b, miR-155 | [69] |
| Plasma | miR-150-5p, miR-25-3p, miR-451a, miR-144-3p, miR-363-3p, miR-4454+miR-7975, miR-873-3p, miR-548al, miR-598-3p, miR-4443, miR-551a, miR-6721-5p | [70] |
| Serum | miR-7-5p, miR-23b-3p, miR-142-3p, miR-221-5p, miR-370-3p | [71] |
| Peripheral whole blood | miR-376a-3p, miR-3680-5p, miR-4253-5p, miR-4482-3p, miR-145-5p | [72] |
| Blood mononuclear cells | miR-499-5p | [73] |
| Whole blood | Manic episode: miR-9-5p, miR-29a-3p, miR-106a-5p, miR-106b-5p, miR-107, miR-125a-3p, miR-125b-5p (compared to controls), miR-106a-5p, miR-107 (compared to euthymic episode). euthymic episode: miR-29a-3p, miR-106b-5p, miR-107, miR-125a-3p (compared to controls) | [74] |
| Blood | miR-15a-5p, miR-17-3p, miR-17-5p, miR-18a-5p, miR-19b-3p, miR-20a-5p, miR-27a-3p, miR-30b-5p, miR-106a-5p, miR-106b-5p, miR-145-5p, miR-148b-3p, miR-210-3p, miR-339-5p, Asenapine: miR-92b-5p, miR-1343-5p. Risperidone: miR-146b-5p, miR-664b-5p, miR-6778-5p | [75] |
| Plasma | let-7e-5p, miR-125a-5p (compared to controls) | [50] |
| Plasma | miR-134 (compared to MDD), miR-134 (compared to controls) | [55] |
| Plasma | miR19b-3p | [76] |
| Blood mononuclear cells | miR-21-3p, miR-29c-5p, miR-30d-5p, miR-140-3p, miR-330-3p, miR-330-5p, miR-345-5p, miR-378a-5p, miR-720-5p, miR-1973-5p, miR-3158-3p, miR-4521-5p, miR-1915-5p, miR-1972-5p, miR-4440-5p, miR-4793-3p | [41] |
| Serum | (Correlation with BDNF: miR7-5p, miR221-5p, miR370-3p) | [77] |
| Whole blood | miR-15b, miR-132, miR-652 | [78] |
| Suicidal Behavior | ||
| Plasma | miR-424-5p, miR-378i, miR-6724-5p, and miR-10b-5p | [79] |
| Biocomputation analysis | miR-27b-3p, miR-124-3p, miR-129-5p, miR-381-3p, miR-3135b, miR-4516 and miR-4286 | [80] |
| Blood mononuclear cells | miR-19a-3p | [81] |
| Whole blood | miR-3688 and miR-5695 | [82] |
| Name | Accession | Location | Sequence |
|---|---|---|---|
| Major Depressive Disorder and Bipolar Disorder Enriched miRNAs | |||
| hsa-let-7a-5p | MIMAT0000062 | chr22:46,112,752-46,112,773 (+); chr11:122,146,568-122,146,589 (−); chr9:94,175,962-94,175,983 (+) | UGAGGUAGUAGGUUGUAUAGUU |
| hsa-let-7b-5p | MIMAT0000063 | chr22:46,113,691-46,113,712 (+) | UGAGGUAGUAGGUUGUGUGGUU |
| hsa-let-7d-3p | MIMAT0004484 | chr9:94,178,895-94,178,916 (+) | CUAUACGACCUGCUGCCUUUCU |
| hsa-let-7d-5p | MIMAT0000065 | chr9:94,178,841-94,178,862 (+) | AGAGGUAGUAGGUUGCAUAGUU |
| hsa-let-7f-5p | MIMAT0000067 | chr9:94,176,353-94,176,374 (+); chrX:53,557,246-53,557,267 (−) | UGAGGUAGUAGAUUGUAUAGUU |
| hsa-miR-10a-5p | MIMAT0000253 | chr17:48,579,904-48,579,926 (−) | UACCCUGUAGAUCCGAAUUUGUG |
| hsa-miR-1180-3p | MIMAT0005825 | chr17:19,344,513-19,344,534 (−) | UUUCCGGCUCGCGUGGGUGUGU |
| hsa-miR-1193 | MIMAT0015049 | chr14:101,030,061-101,030,083 (+) | GGGAUGGUAGACCGGUGACGUGC |
| hsa-miR-122-5p | MIMAT0000421 | chr18:58,451,088-58,451,109 (+) | UGGAGUGUGACAAUGGUGUUUG |
| hsa-miR-124-3p | MIMAT0000422 | chr20:63,178,552-63,178,571 (+); chr8:9,903,401-9,903,420 (−); chr8:64,379,210-64,379,229 (+) | UAAGGCACGCGGUGAAUGCC |
| hsa-miR-1244 | MIMAT0005896 | chr12:9,239,472-9,239,497 (−); chr12:12,112,006-12,112,031 (+); chr2:231,713,368-231,713,393 (+); chr5:118,974,640-118,974,665 (+) | AAGUAGUUGGUUUGUAUGAGAUGGUU |
| hsa-miR-126-3p | MIMAT0000445 | chr9:136,670,653-136,670,674 (+) | UCGUACCGUGAGUAAUAAUGCG |
| hsa-miR-126-5p | MIMAT0000444 | chr9:136,670,616-136,670,636 (+) | CAUUAUUACUUUUGGUACGCG |
| hsa-miR-1275 | MIMAT0005929 | chr6:34,000,018-34,000,034 (−) | GUGGGGGAGAGGCUGUC |
| hsa-miR-129-5p | MIMAT0000242 | chr7:128,207,876-128,207,896 (+); chr11:43,581,408-43,581,428 (+) | CUUUUUGCGGUCUGGGCUUGC |
| hsa-miR-139-3p | MIMAT0004552 | chr11:72,615,066-72,615,088 (−) | UGGAGACGCGGCCCUGUUGGAGU |
| hsa-miR-139-5p | MIMAT0000250 | chr11:72,615,102-72,615,124 (−) | UCUACAGUGCACGUGUCUCCAGU |
| hsa-miR-143-3p | MIMAT0000435 | chr5:149,428,978-149,428,998 (+) | UGAGAUGAAGCACUGUAGCUC |
| hsa-miR-146a-5p | MIMAT0000449 | chr5:160,485,372-160,485,393 (+) | UGAGAACUGAAUUCCAUGGGUU |
| hsa-miR-15b-5p | MIMAT0000417 | chr3:160,404,607-160,404,628 (+) | UAGCAGCACAUCAUGGUUUACA |
| hsa-miR-16-5p | MIMAT0000069 | chr3:160,404,754-160,404,775 (+); chr13:50,049,027-50,049,048 (−) | UAGCAGCACGUAAAUAUUGGCG |
| hsa-miR-191-5p | MIMAT0000440 | chr3:49,020,672-49,020,694 (−) | CAACGGAAUCCCAAAAGCAGCUG |
| hsa-miR-1915-3p | MIMAT0007892 | chr10:21,496,576-21,496,595 (−) | CCCCAGGGCGACGCGGCGGG |
| hsa-miR-193a-5p | MIMAT0004614 | chr17:31,560,016-31,560,037 (+) | UGGGUCUUUGCGGGCGAGAUGA |
| hsa-miR-197-3p | MIMAT0000227 | chr1:109,598,940-109,598,961 (+) | UUCACCACCUUCUCCACCCAGC |
| hsa-miR-1972 | MIMAT0009447 | chr16:15,010,329-15,010,350 (−); chr16:70,030,393-70,030,414 (+) | UCAGGCCAGGCACAGUGGCUCA |
| hsa-miR-199a-5p | MIMAT0000231 | chr19:10,817,469-10,817,491 (−); chr1:172,144,592-172,144,614 (−) | CCCAGUGUUCAGACUACCUGUUC |
| hsa-miR-199b-5p | MIMAT0000263 | chr9:128,244,783-128,244,805 (−) | CCCAGUGUUUAGACUAUCUGUUC |
| hsa-miR-200a-3p | MIMAT0000682 | chr1:1,167,916-1,167,937 (+) | UAACACUGUCUGGUAACGAUGU |
| hsa-miR-2110 | MIMAT0010133 | chr10:114,174,151-114,174,172 (−) | UUGGGGAAACGGCCGCUGAGUG |
| hsa-miR-215-5p | MIMAT0000272 | chr1:220,117,916-220,117,936 (−) | AUGACCUAUGAAUUGACAGAC |
| hsa-miR-221-3p | MIMAT0000278 | chrX:45,746,180-45,746,202 (−) | AGCUACAUUGUCUGCUGGGUUUC |
| hsa-miR-223-3p | MIMAT0000280 | chrX:66,018,937-66,018,958 (+) | UGUCAGUUUGUCAAAUACCCCA |
| hsa-miR-2278 | MIMAT0011778 | chr9:94,809,977-94,809,998 (+) | GAGAGCAGUGUGUGUUGCCUGG |
| hsa-miR-24-3p | MIMAT0000080 | chr9:95,086,064-95,086,085 (+); chr19:13,836,289-13,836,310 (−) | UGGCUCAGUUCAGCAGGAACAG |
| hsa-miR-29b-3p | MIMAT0000100 | chr7:130,877,467-130,877,489 (−); chr1:207,802,450-207,802,472 (−) | UAGCACCAUUUGAAAUCAGUGUU |
| hsa-miR-3074-5p | MIMAT0019208 | chr9:95,086,063-95,086,083 (−) | GUUCCUGCUGAACUGAGCCAG |
| hsa-miR-30a-5p | MIMAT0000087 | chr6:71,403,595-71,403,616 (−) | UGUAAACAUCCUCGACUGGAAG |
| hsa-miR-30e-3p | MIMAT0000693 | chr1:40,754,413-40,754,434 (+) | CUUUCAGUCGGAUGUUUACAGC |
| hsa-miR-30e-5p | MIMAT0000692 | chr1:40,754,371-40,754,392 (+) | UGUAAACAUCCUUGACUGGAAG |
| hsa-miR-3150a-3p | MIMAT0015023 | chr8:95,072,962-95,072,983 (+) | CUGGGGAGAUCCUCGAGGUUGG |
| hsa-miR-3154 | MIMAT0015028 | chr9:128,244,956-128,244,977 (−) | CAGAAGGGGAGUUGGGAGCAGA |
| hsa-miR-3173-3p | MIMAT0015048 | chr14:95,137,923-95,137,944 (−) | AAAGGAGGAAAUAGGCAGGCCA |
| hsa-miR-3173-5p | MIMAT0019214 | chr14:95,137,961-95,137,982 (−) | UGCCCUGCCUGUUUUCUCCUUU |
| hsa-miR-3184-5p | MIMAT0015064 | chr17:30,117,128-30,117,151 (−) | UGAGGGGCCUCAGACCGAGCUUUU |
| hsa-miR-320a | MIMAT0000510 | chr8:22,244,975-22,244,996 (−) | AAAAGCUGGGUUGAGAGGGCGA |
| hsa-miR-324-5p | MIMAT0000761 | chr17:7,223,342-7,223,364 (−) | CGCAUCCCCUAGGGCAUUGGUGU |
| hsa-miR-335-5p | MIMAT0000765 | chr7:130,496,126-130,496,148 (+) | UCAAGAGCAAUAACGAAAAAUGU |
| hsa-miR-33a-5p | MIMAT0000091 | chr22:41,900,949-41,900,969 (+) | GUGCAUUGUAGUUGCAUUGCA |
| hsa-miR-34a-5p | MIMAT0000255 | chr1:9,151,735-9,151,756 (−) | UGGCAGUGUCUUAGCUGGUUGU |
| hsa-miR-34b-5p | MIMAT0000685 | chr11:111,512,950-111,512,972 (+) | UAGGCAGUGUCAUUAGCUGAUUG |
| hsa-miR-34c-5p | MIMAT0000686 | chr11:111,513,451-111,513,473 (+) | AGGCAGUGUAGUUAGCUGAUUGC |
| hsa-miR-3605-3p | MIMAT0017982 | chr1:33,332,405-33,332,427 (−) | CCUCCGUGUUACCUGUCCUCUAG |
| hsa-miR-361-3p | MIMAT0004682 | chrX:85,903,641-85,903,663 (−) | UCCCCCAGGUGUGAUUCUGAUUU |
| hsa-miR-361-5p | MIMAT0000703 | chrX:85,903,681-85,903,702 (−) | UUAUCAGAAUCUCCAGGGGUAC |
| hsa-miR-3611 | MIMAT0017988 | chr10:35,079,609-35,079,629 (−) | UUGUGAAGAAAGAAAUUCUUA |
| hsa-miR-3661 | MIMAT0018082 | chr5:134,225,777-134,225,798 (+) | UGACCUGGGACUCGGACAGCUG |
| hsa-miR-375 | MIMAT0000728 | chr2:219,001,648-219,001,669 (−) | UUUGUUCGUUCGGCUCGCGUGA |
| hsa-miR-3909 | MIMAT0018183 | chr22:35,335,710-35,335,731 (+) | UGUCCUCUAGGGCCUGCAGUCU |
| hsa-miR-3944-5p | MIMAT0019231 | chr10:133,371,621-133,371,641 (−) | UGUGCAGCAGGCCAACCGAGA |
| hsa-miR-423-3p | MIMAT0001340 | chr17:30,117,131-30,117,153 (+) | AGCUCGGUCUGAGGCCCCUCAGU |
| hsa-miR-425-3p | MIMAT0001343 | chr3:49,020,159-49,020,180 (−) | AUCGGGAAUGUCGUGUCCGCCC |
| hsa-miR-425-5p | MIMAT0003393 | chr3:49,020,199-49,020,221 (−) | AAUGACACGAUCACUCCCGUUGA |
| hsa-miR-4498 | MIMAT0019033 | chr12:120,155,473-120,155,494 (−) | UGGGCUGGCAGGGCAAGUGCUG |
| hsa-miR-4532 | MIMAT0019071 | chr20:57,895,399-57,895,415 (+) | CCCCGGGGAGCCCGGCG |
| hsa-miR-483-3p | MIMAT0002173 | chr11:2,134,142-2,134,162 (−) | UCACUCCUCUCCUCCCGUCUU |
| hsa-miR-483-5p | MIMAT0004761 | chr11:2,134,181-2,134,202 (−) | AAGACGGGAGGAAAGAAGGGAG |
| hsa-miR-486-5p | MIMAT0002177 | chr8:41,660,444-41,660,465 (+); chr8:41,660,484-41,660,505 (−) | UCCUGUACUGAGCUGCCCCGAG |
| hsa-miR-502-3p | MIMAT0004775 | chrX:50,014,649-50,014,670 (+) | AAUGCACCUGGGCAAGGAUUCA |
| hsa-miR-503-5p | MIMAT0002874 | chrX:134,546,371-134,546,393 (−) | UAGCAGCGGGAACAGUUCUGCAG |
| hsa-miR-532-3p | MIMAT0004780 | chrX:50,003,204-50,003,225 (+) | CCUCCCACACCCAAGGCUUGCA |
| hsa-miR-574-3p | MIMAT0003239 | chr4:38,868,092-38,868,113 (+) | CACGCUCAUGCACACACCCACA |
| hsa-miR-608 | MIMAT0003276 | chr10:100,975,000-100,975,024 (+) | AGGGGUGGUGUUGGGACAGCUCCGU |
| hsa-miR-636 | MIMAT0003306 | chr17:76,736,466-76,736,488 (−) | UGUGCUUGCUCGUCCCGCCCGCA |
| hsa-miR-637 | MIMAT0003307 | chr19:3,961,429-3,961,452 (−) | ACUGGGGGCUUUCGGGCUCUGCGU |
| hsa-miR-6511a-3p | MIMAT0025479 | chr16:14,925,980-14,926,001 (+); chr16:16,324,631-16,324,652 (+); chr16:16,368,919-16,368,940 (+); chr16:18,344,015-18,344,036 (−) | CCUCACCAUCCCUUCUGCCUGC |
| hsa-miR-6750-3p | MIMAT0027401 | chr11:64,898,363-64,898,383 (−) | GAACUCACCCUCUGCUCCCAG |
| hsa-miR-6881-3p | MIMAT0027663 | chr15:74,411,361-74,411,382 (−) | AUCCUCUUUCGUCCUUCCCACU |
| hsa-miR-770-5p | MIMAT0003948 | chr14:100,852,409-100,852,431 (+) | UCCAGUACCACGUGUCAGGGCCA |
| hsa-miR-93-5p | MIMAT0000093 | chr7:100,093,815-100,093,837 (−) | CAAAGUGCUGUUCGUGCAGGUAG |
| Bipolar Disorder Enriched miRNAs | |||
| hsa-let-7i-5p | MIMAT0000415 | chr12:62,603,691-62,603,712 (+) | UGAGGUAGUAGUUUGUGCUGUU |
| hsa-miR-106a-5p | MIMAT0000103 | chrX:134,170,244-134,170,266 (−) | AAAAGUGCUUACAGUGCAGGUAG |
| hsa-miR-107 | MIMAT0000104 | chr10:89,592,756-89,592,778 (−) | AGCAGCAUUGUACAGGGCUAUCA |
| hsa-miR-1185-2-3p | MIMAT0022713 | chr14:101,044,250-101,044,271 (+) | AUAUACAGGGGGAGACUCUCAU |
| hsa-miR-1227-5p | MIMAT0022941 | chr19:2,234,133-2,234,149 (−) | GUGGGGCCAGGCGGUGG |
| hsa-miR-1238-3p | MIMAT0005593 | chr19:10,552,183-10,552,202 (+) | CUUCCUCGUCUGUCUGCCCC |
| hsa-miR-125a-3p | MIMAT0004602 | chr19:51,693,306-51,693,327 (+) | ACAGGUGAGGUUCUUGGGAGCC |
| hsa-miR-1268b | MIMAT0018925 | chr17:80,098,831-80,098,850 (+) | CGGGCGUGGUGGUGGGGGUG |
| hsa-miR-1281 | MIMAT0005939 | chr22:41,092,545-41,092,561 (+) | UCGCCUCCUCCUCUCCC |
| hsa-miR-133a-3p | MIMAT0000427 | chr18:21,825,712-21,825,733 (−); chr20:62,564,970-62,564,991 (+) | UUUGGUCCCCUUCAACCAGCUG |
| hsa-miR-1343-5p | MIMAT0027038 | chr11:34,941,851-34,941,872 (+) | UGGGGAGCGGCCCCCGGGUGGG |
| hsa-miR-142-3p | MIMAT0000434 | chr17:58,331,245-58,331,267 (−) | UGUAGUGUUUCCUACUUUAUGGA |
| hsa-miR-145-5p | MIMAT0000437 | chr5:149,430,661-149,430,683 (+) | GUCCAGUUUUCCCAGGAAUCCCU |
| hsa-miR-15a-5p | MIMAT0000068 | chr13:50,049,167-50,049,188 (−) | UAGCAGCACAUAAUGGUUUGUG |
| hsa-miR-17-3p | MIMAT0000071 | chr13:91,350,655-91,350,676 (+) | ACUGCAGUGAAGGCACUUGUAG |
| hsa-miR-188-5p | MIMAT0000457 | chrX:50,003,517-50,003,537 (+) | CAUCCCUUGCAUGGUGGAGGG |
| hsa-miR-18a-5p | MIMAT0000072 | chr13:91,350,756-91,350,778 (+) | UAAGGUGCAUCUAGUGCAGAUAG |
| hsa-miR-1915-5p | MIMAT0007891 | chr10:21,496,609-21,496,630 (−) | ACCUUGCCUUGCUGCCCGGGCC |
| hsa-miR-19b-3p | MIMAT0000074 | chr13:91,351,245-91,351,267 (+); chrX:134,169,683-134,169,705 (−) | UGUGCAAAUCCAUGCAAAACUGA |
| hsa-miR-20a-5p | MIMAT0000075 | chr13:91,351,072-91,351,094 (+) | UAAAGUGCUUAUAGUGCAGGUAG |
| hsa-miR-21-3p | MIMAT0004494 | chr17:59,841,311-59,841,331 (+) | CAACACCAGUCGAUGGGCUGU |
| hsa-miR-210-3p | MIMAT0000267 | chr11:568,112-568,133 (−) | CUGUGCGUGUGACAGCGGCUGA |
| hsa-miR-22-3p | MIMAT0000077 | chr17:1,713,914-1,713,935 (−) | AAGCUGCCAGUUGAAGAACUGU |
| hsa-miR-221-5p | MIMAT0004568 | chrX:45,746,221-45,746,242 (−) | ACCUGGCAUACAAUGUAGAUUU |
| hsa-miR-23b-3p | MIMAT0000418 | chr9:95,085,265-95,085,285 (+) | AUCACAUUGCCAGGGAUUACC |
| hsa-miR-27a-3p | MIMAT0000084 | chr19:13,836,447-13,836,467 (−) | UUCACAGUGGCUAAGUUCCGC |
| hsa-miR-29a-3p | MIMAT0000086 | chr7:130,876,748-130,876,769 (−) | UAGCACCAUCUGAAAUCGGUUA |
| hsa-miR-301a-3p | MIMAT0000688 | chr17:59,151,149-59,151,171 (−) | CAGUGCAAUAGUAUUGUCAAAGC |
| hsa-miR-30b-5p | MIMAT0000420 | chr8:134,800,570-134,800,591 (−) | UGUAAACAUCCUACACUCAGCU |
| hsa-miR-3135b | MIMAT0018985 | chr6:32,749,952-32,749,973 (−) | GGCUGGAGCGAGUGCAGUGGUG |
| hsa-miR-3194-5p | MIMAT0015078 | chr20:51,452,948-51,452,968 (−) | GGCCAGCCACCAGGAGGGCUG |
| hsa-miR-330-5p | MIMAT0004693 | chr19:45,639,049-45,639,070 (−) | UCUCUGGGCCUGUGUCUUAGGC |
| hsa-miR-339-5p | MIMAT0000764 | chr7:1,022,990-1,023,012 (−) | UCCCUGUCCUCCAGGAGCUCACG |
| hsa-miR-3620-5p | MIMAT0022967 | chr1:228,097,285-228,097,306 (+) | GUGGGCUGGGCUGGGCUGGGCC |
| hsa-miR-363-3p | MIMAT0000707 | chrX:134,169,382-134,169,403 (−) | AAUUGCACGGUAUCCAUCUGUA |
| hsa-miR-3680-5p | MIMAT0018106 | chr16:29,599,231-29,599,252 (−); chr16:21,506,101-21,506,122 (−) | GACUCACUCACAGGAUUGUGCA |
| hsa-miR-370-3p | MIMAT0000722 | chr14:100,911,186-100,911,207 (+) | GCCUGCUGGGGUGGAACCUGGU |
| hsa-miR-376a-3p | MIMAT0000729 | chr14:101,040,825-101,040,845 (+); chr14:101,040,118-101,040,138 (+) | AUCAUAGAGGAAAAUCCACGU |
| hsa-miR-376b-3p | MIMAT0002172 | chr14:101,040,497-101,040,518 (+) | AUCAUAGAGGAAAAUCCAUGUU |
| hsa-miR-378a-5p | MIMAT0000731 | chr5:149,732,829-149,732,850 (+) | CUCCUGACUCCAGGUCCUGUGU |
| hsa-miR-4433a-5p | MIMAT0020956 | chr2:64,340,770-64,340,790 (+) | CGUCCCACCCCCCACUCCUGU |
| hsa-miR-4443 | MIMAT0018961 | chr3:48,196,572-48,196,588 (+) | UUGGAGGCGUGGGUUUU |
| hsa-miR-4454 | MIMAT0018976 | chr4:163,093,607-163,093,626 (−) | GGAUCCGAGUCACGGCACCA |
| hsa-miR-4482-3p | MIMAT0020958 | chr10:104,268,341-104,268,362 (−) | UUUCUAUUUCUCAGUGGGGCUC |
| hsa-miR-4793-3p | MIMAT0019966 | chr3:48,644,201-48,644,223 (−) | UCUGCACUGUGAGUUGGCUGGCU |
| hsa-miR-484 | MIMAT0002174 | chr16:15,643,301-15,643,322 (+) | UCAGGCUCAGUCCCCUCCCGAU |
| hsa-miR-548al | MIMAT0019024 | chr11:74,399,301-74,399,322 (+) | AACGGCAAUGACUUUUGUACCA |
| hsa-miR-551a | MIMAT0003214 | chr1:3,560,710-3,560,730 (−) | GCGACCCACUCUUGGUUUCCA |
| hsa-miR-5739 | MIMAT0023116 | chr22:28,459,925-28,459,944 (+) | GCGGAGAGAGAAUGGGGAGC |
| hsa-miR-598-3p | MIMAT0003266 | chr8:11,035,221-11,035,242 (−) | UACGUCAUCGUUGUCAUCGUCA |
| hsa-miR-6068 | MIMAT0023693 | chr1:63,326,964-63,326,984 (−) | CCUGCGAGUCUCCGGCGGUGG |
| hsa-miR-607 | MIMAT0003275 | chr10:96,828,684-96,828,704 (−) | GUUCAAAUCCAGAUCUAUAAC |
| hsa-miR-6090 | MIMAT0023715 | chr11:128,522,430-128,522,448 (+) | GGGGAGCGAGGGGCGGGGC |
| hsa-miR-6125 | MIMAT0024598 | chr12:62,260,418-62,260,437 (+) | GCGGAAGGCGGAGCGGCGGA |
| hsa-miR-633 | MIMAT0003303 | chr17:62,944,275-62,944,297 (+) | CUAAUAGUAUCUACCACAAUAAA |
| hsa-miR-664b-5p | MIMAT0022271 | chrX:154,768,596-154,768,619 (+) | UGGGCUAAGGGAGAUGAUUGGGUA |
| hsa-miR-671-5p | MIMAT0003880 | chr7:151,238,449-151,238,471 (+) | AGGAAGCCCUGGAGGGGCUGGAG |
| hsa-miR-6721-5p | MIMAT0025852 | chr6:32,170,084-32,170,106 (−) | UGGGCAGGGGCUUAUUGUAGGAG |
| hsa-miR-6727-5p | MIMAT0027355 | chr1:1,312,539-1,312,561 (−) | CUCGGGGCAGGCGGCUGGGAGCG |
| hsa-miR-6775-5p | MIMAT0027450 | chr16:87,834,631-87,834,655 (−) | UCGGGGCAUGGGGGAGGGAGGCUGG |
| hsa-miR-6778-5p | MIMAT0027456 | chr17:18,340,860-18,340,881 (−) | AGUGGGAGGACAGGAGGCAGGU |
| hsa-miR-6791-5p | MIMAT0027482 | chr19:6,736,752-6,736,773 (−) | CCCCUGGGGCUGGGCAGGCGGA |
| hsa-miR-6800-3p | MIMAT0027501 | chr19:49,832,076-49,832,096 (+) | CACCUCUCCUGGCAUCGCCCC |
| hsa-miR-6808-5p | MIMAT0027516 | chr1:1,339,682-1,339,703 (−) | CAGGCAGGGAGGUGGGACCAUG |
| hsa-miR-6821-5p | MIMAT0027542 | chr22:49,962,866-49,962,888 (+) | GUGCGUGGUGGCUCGAGGCGGGG |
| hsa-miR-7-5p | MIMAT0000252 | chr9:83,969,812-83,969,834 (−); chr19:4,770,700-4,770,722 (+); chr15:88,611,856-88,611,878 (+) | UGGAAGACUAGUGAUUUUGUUGU |
| hsa-miR-7108-5p | MIMAT0028113 | chr19:2,434,980-2,435,000 (−) | GUGUGGCCGGCAGGCGGGUGG |
| hsa-miR-7975 | MIMAT0031178 | chr19:55,123,225-55,123,242 (−) | AUCCUAGUCACGGCACCA |
| hsa-miR-7977 | MIMAT0031180 | chr3:176,515,103-176,515,120 (+) | UUCCCAGCCAACGCACCA |
| hsa-miR-8060 | MIMAT0030987 | chr3:96,360,006-96,360,029 (+) | CCAUGAAGCAGUGGGUAGGAGGAC |
| hsa-miR-873-3p | MIMAT0022717 | chr9:28,888,889-28,888,910 (−) | GGAGACUGAUGAGUUCCCGGGA |
| hsa-miR-9-5p | MIMAT0000441 | chr5:88,666,902-88,666,924 (−); chr1:156,420,392-156,420,414 (−); chr15:89,368,032-89,368,054 (+) | UCUUUGGUUAUCUAGCUGUAUGA |
| hsa-miR-92a-3p | MIMAT0000092 | chr13:91,351,361-91,351,382 (+); chrX:134,169,544-134,169,565 (−) | UAUUGCACUUGUCCCGGCCUGU |
| hsa-miR-92b-5p | MIMAT0004792 | chr1:155,195,196-155,195,217 (+) | AGGGACGGGACGCGGUGCAGUG |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Roy, B.; Ochi, S.; Dwivedi, Y. Potential of Circulating miRNAs as Molecular Markers in Mood Disorders and Associated Suicidal Behavior. Int. J. Mol. Sci. 2023, 24, 4664. https://doi.org/10.3390/ijms24054664
Roy B, Ochi S, Dwivedi Y. Potential of Circulating miRNAs as Molecular Markers in Mood Disorders and Associated Suicidal Behavior. International Journal of Molecular Sciences. 2023; 24(5):4664. https://doi.org/10.3390/ijms24054664
Chicago/Turabian StyleRoy, Bhaskar, Shinichiro Ochi, and Yogesh Dwivedi. 2023. "Potential of Circulating miRNAs as Molecular Markers in Mood Disorders and Associated Suicidal Behavior" International Journal of Molecular Sciences 24, no. 5: 4664. https://doi.org/10.3390/ijms24054664