

Overexpression of a Novel Noxo1 Mutant Increases Ros Production and Noxo1 Relocalisation

 ,
, 
Abstract
:1. Introduction
2. Results
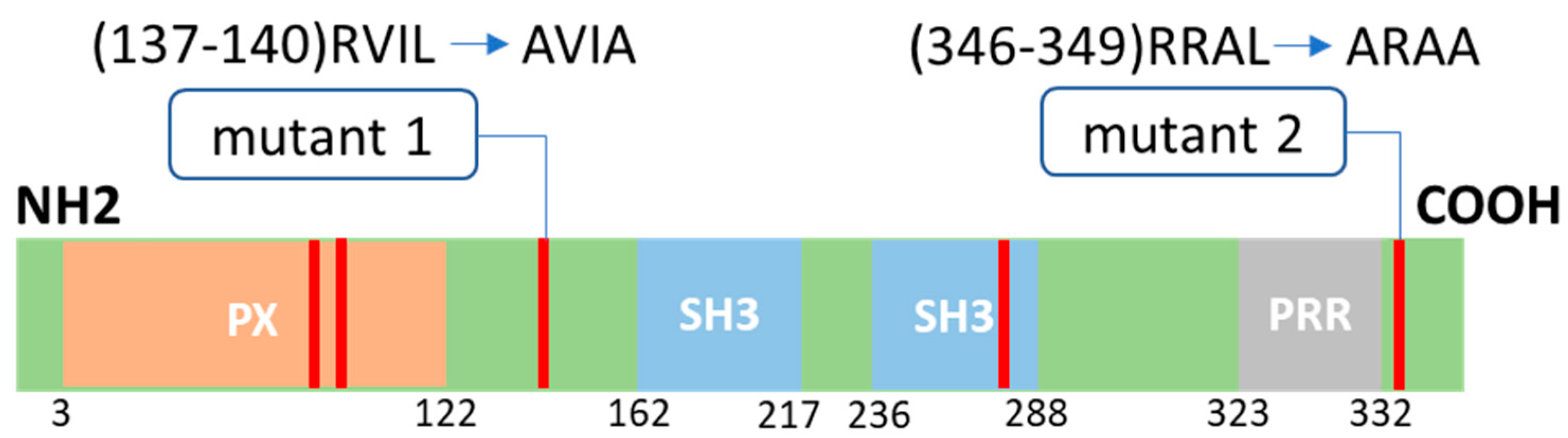
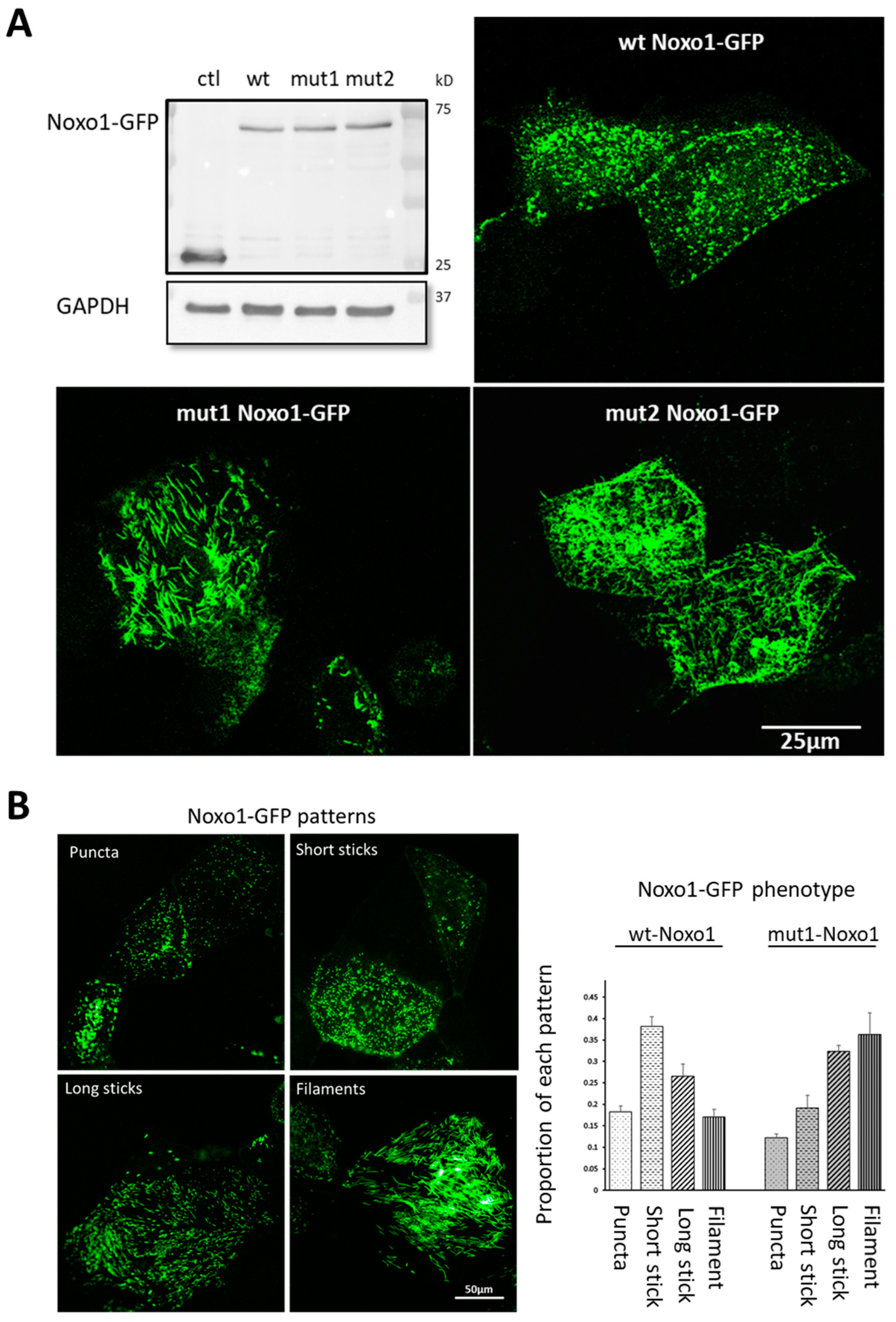
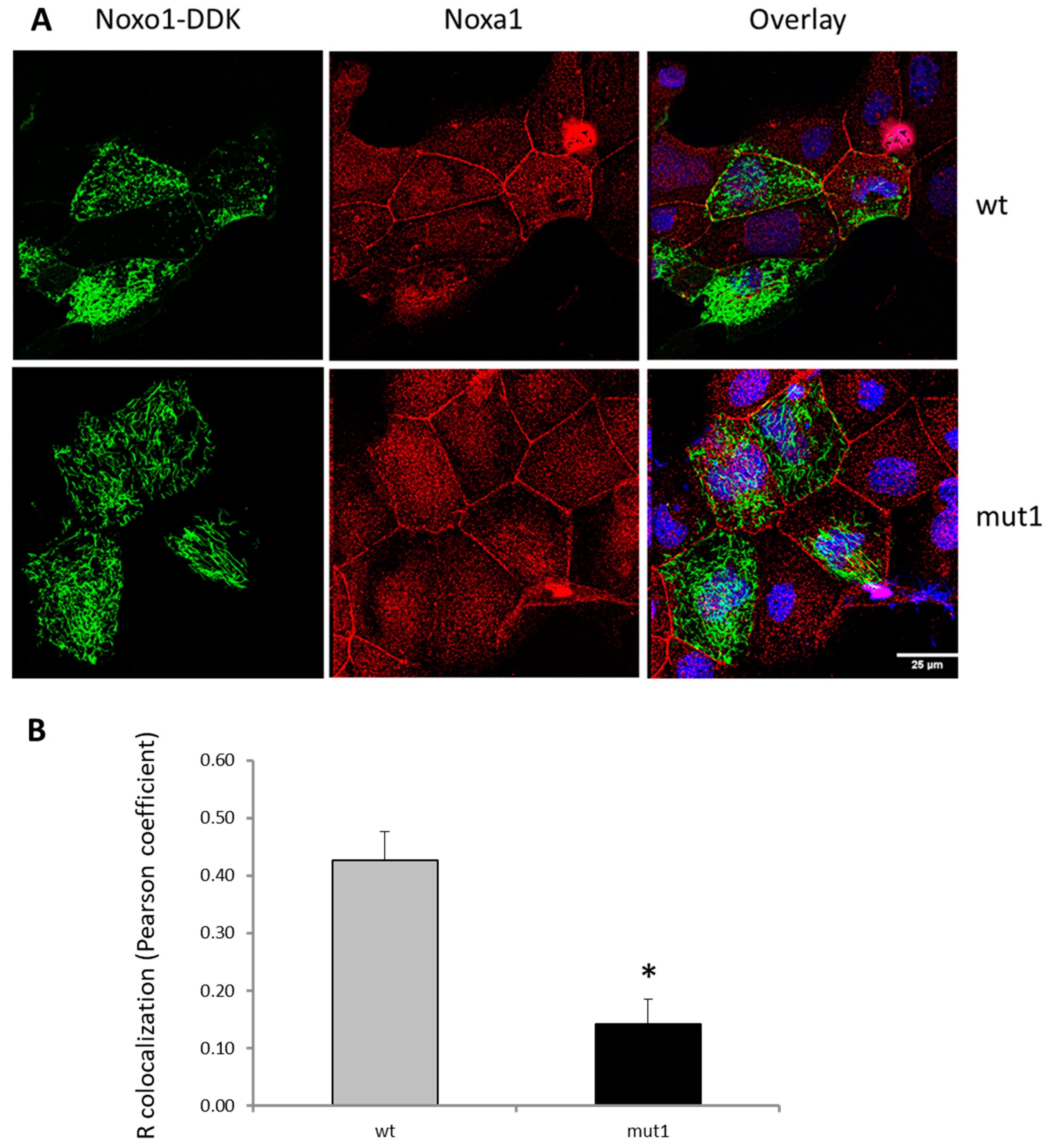
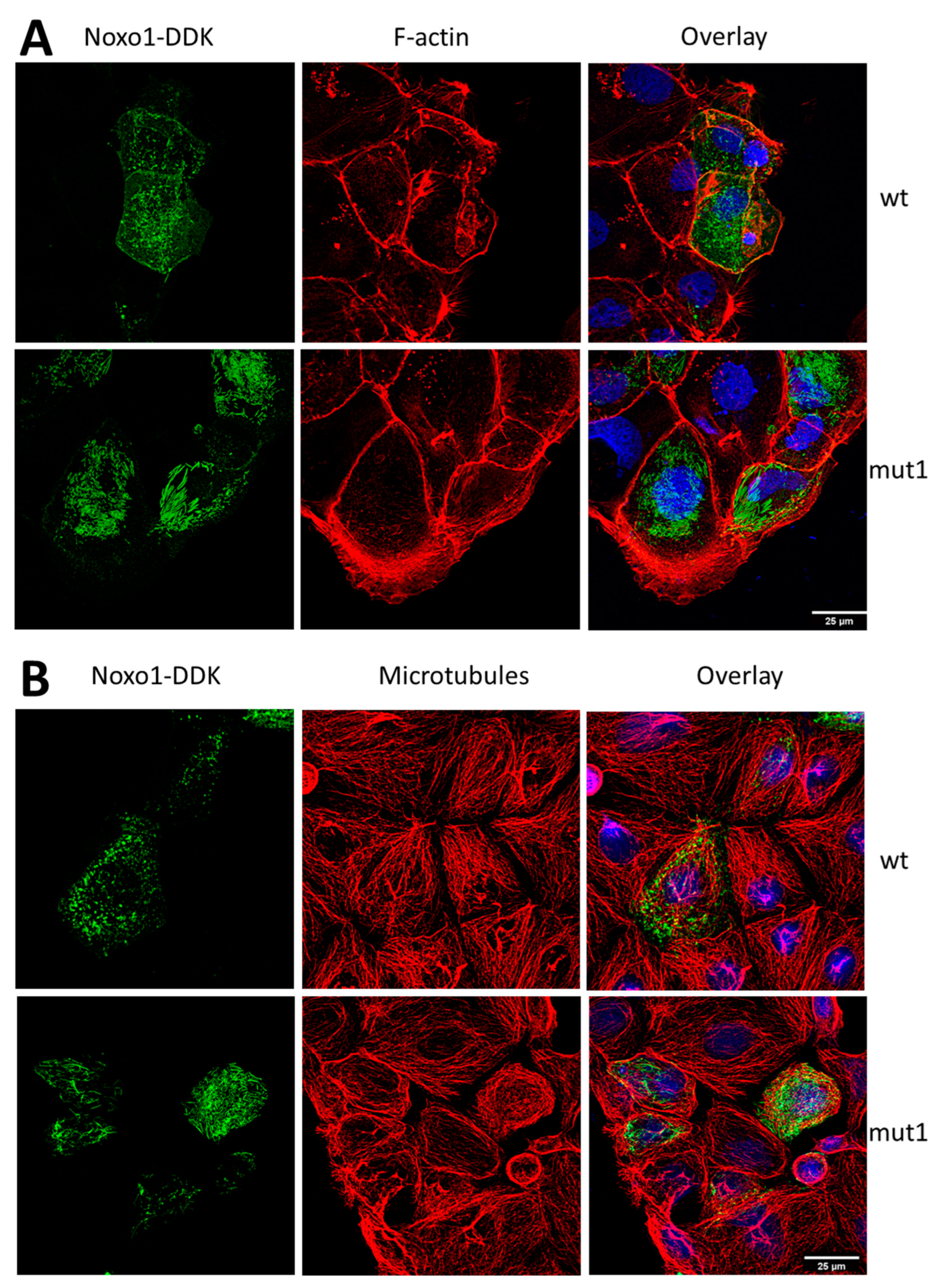
2.1. D-Box Mutations Mut1 of Noxo1 Induces a More Filamentous Pattern in Caco2 Cells
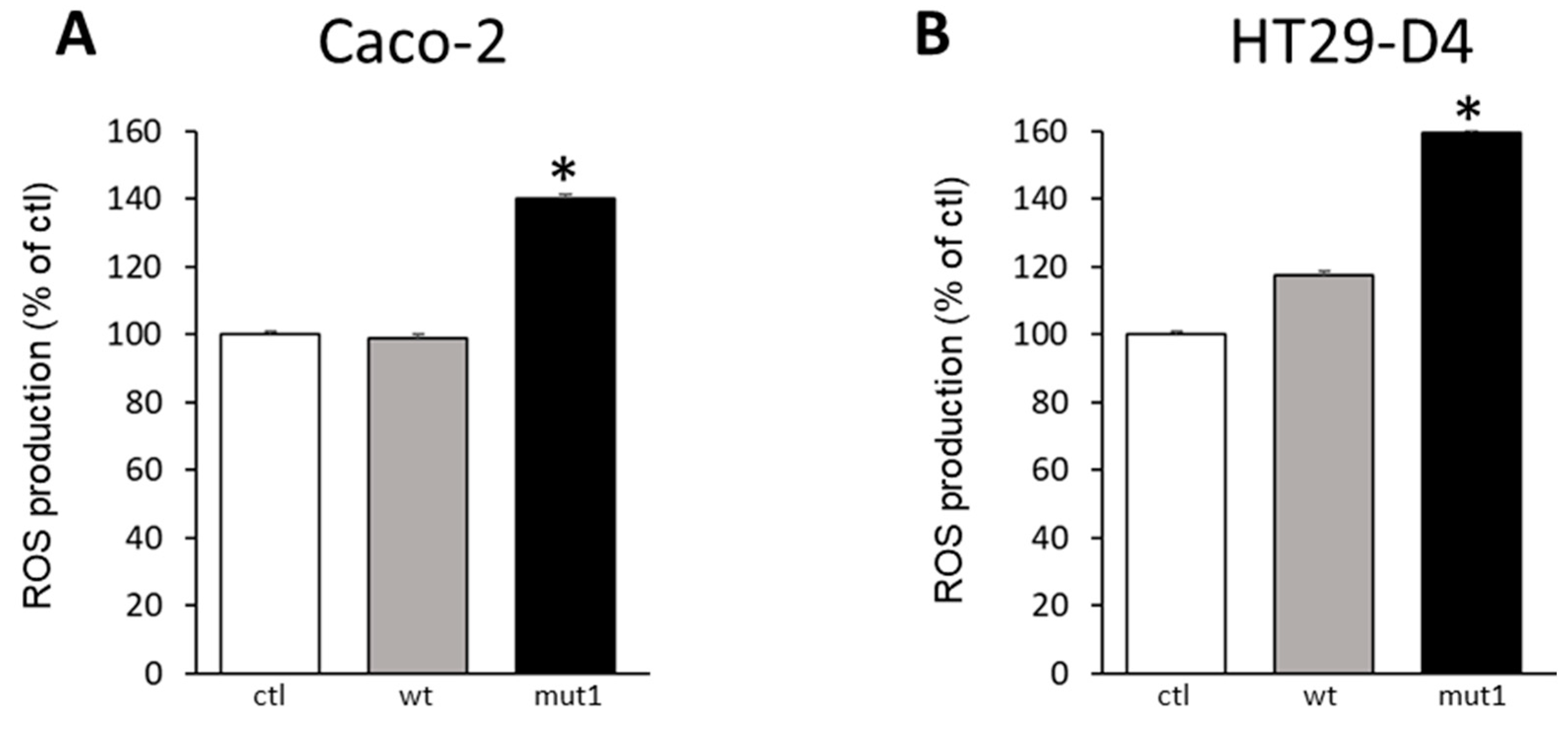
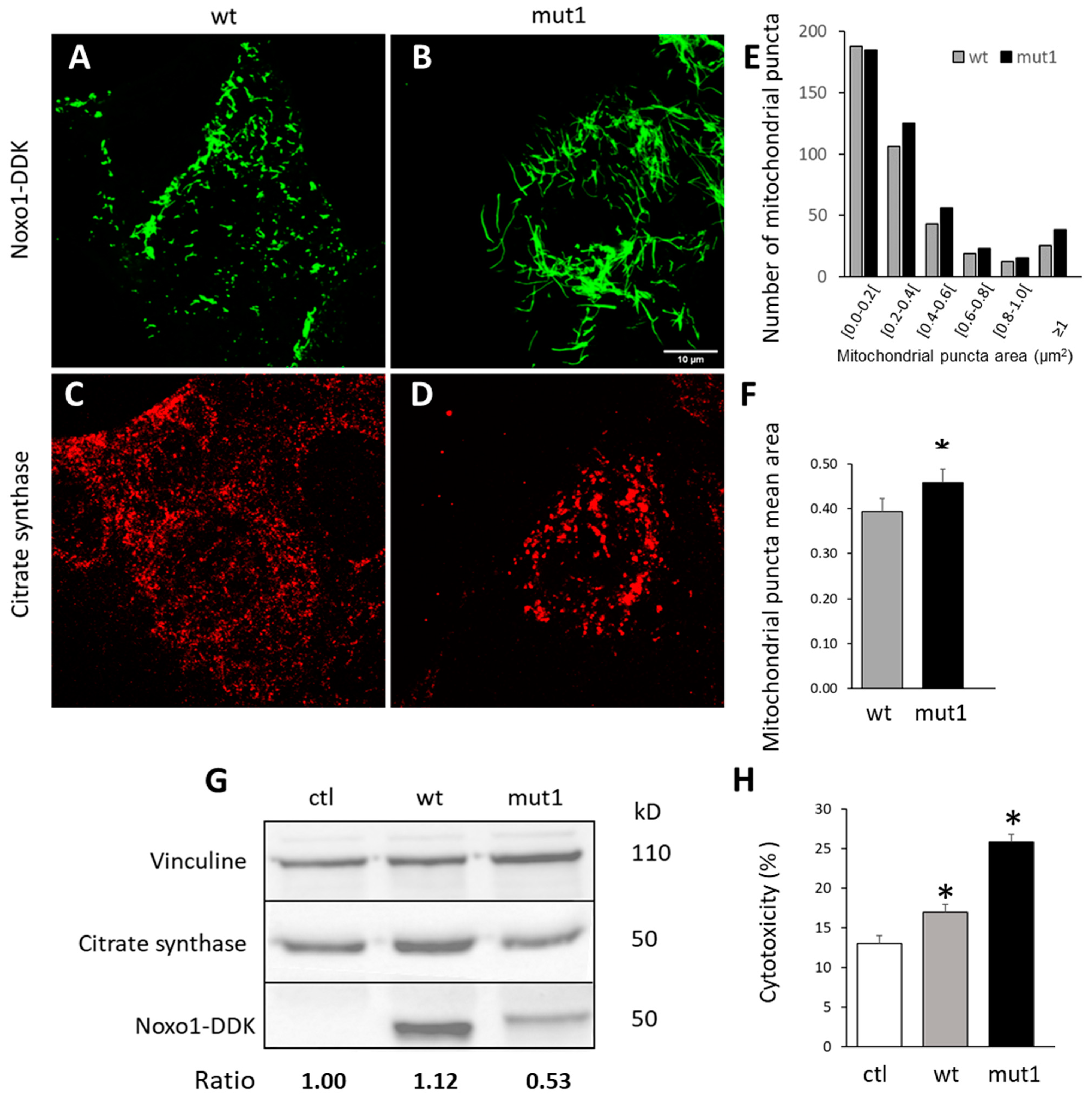
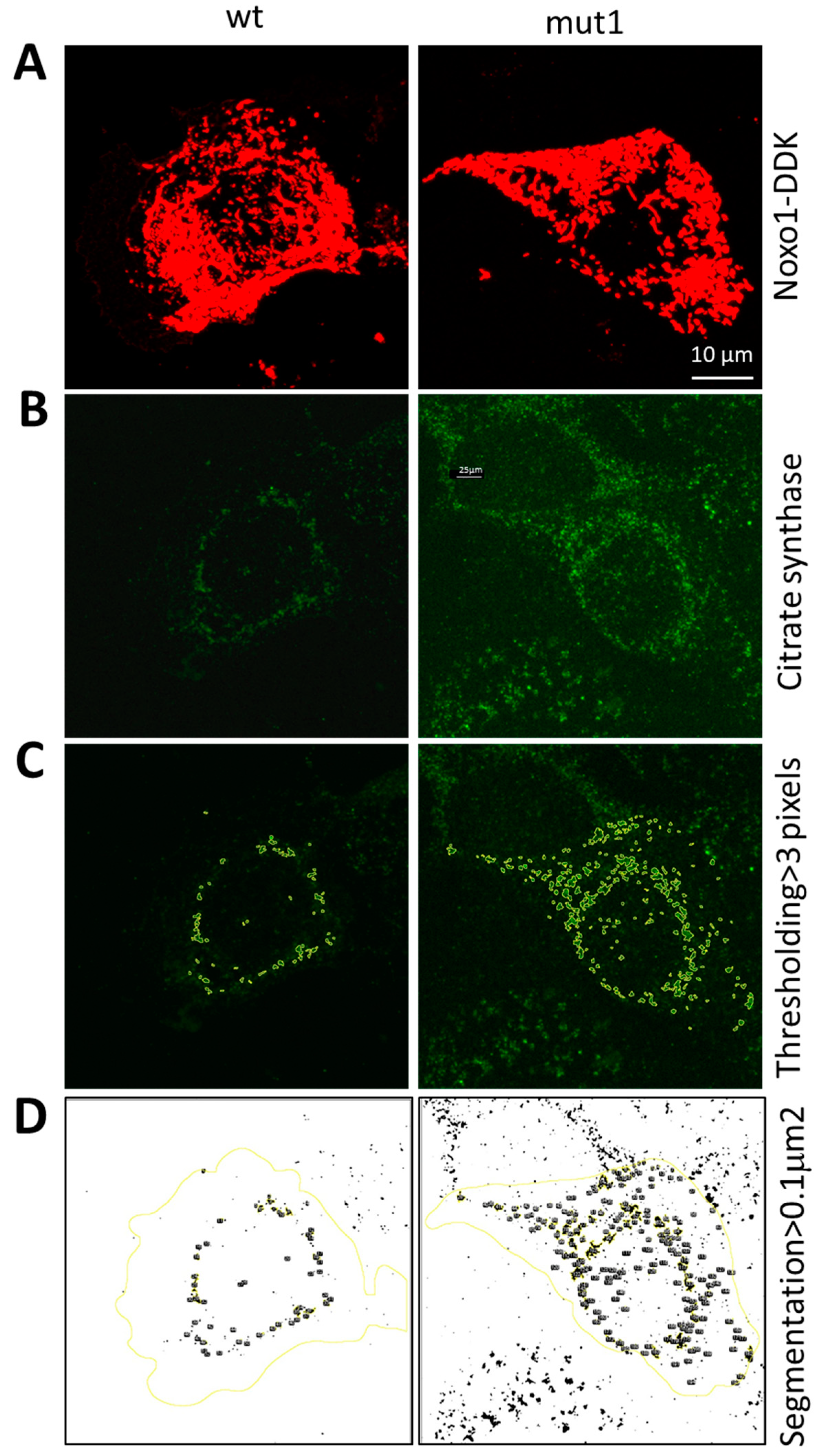
2.2. Noxo1 D-Box Mutation Increases ROS Production, Affects Mitochondrial Organization, and Increases Cytotoxicity in Colorectal Cancer Cell Lines
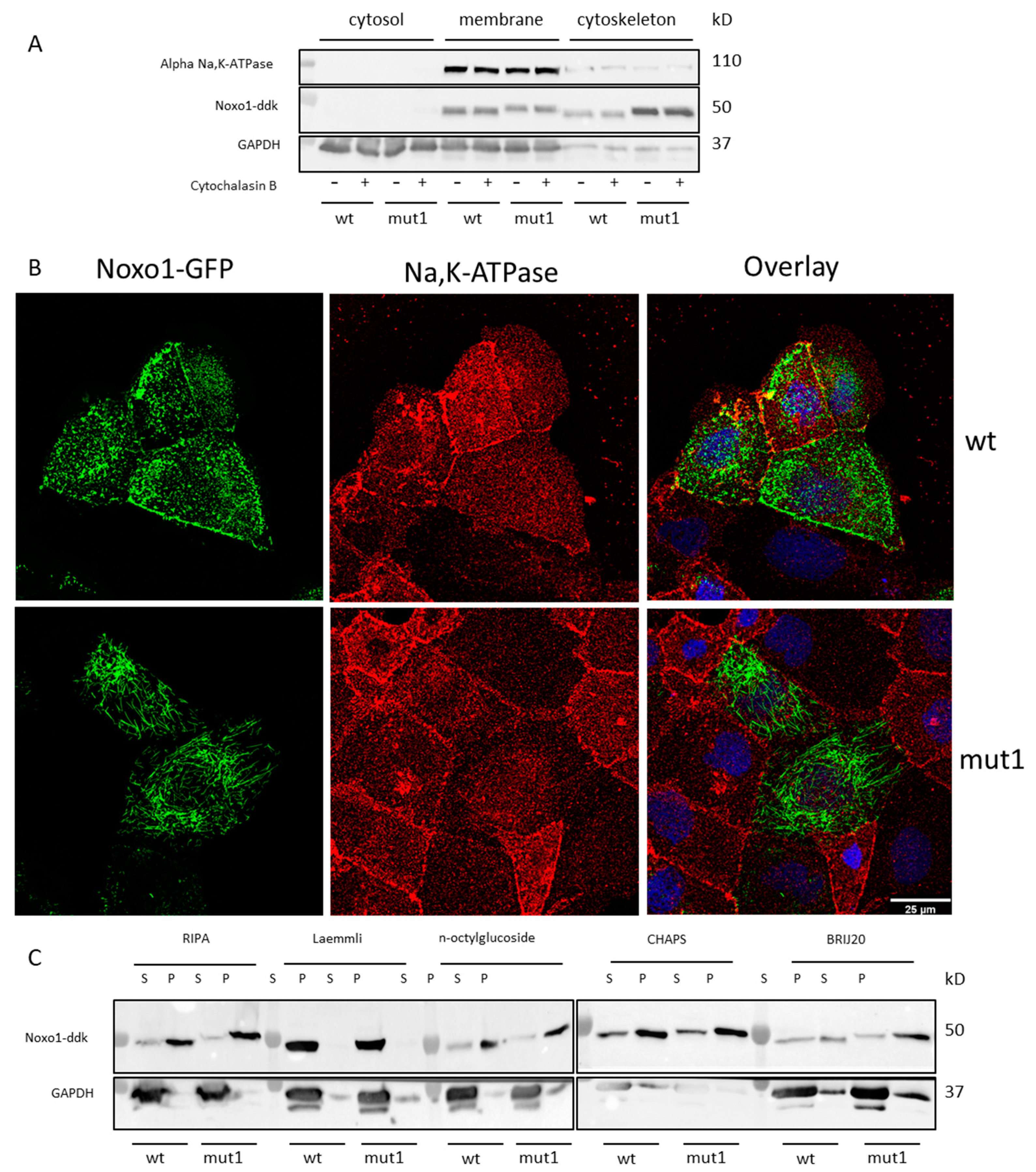
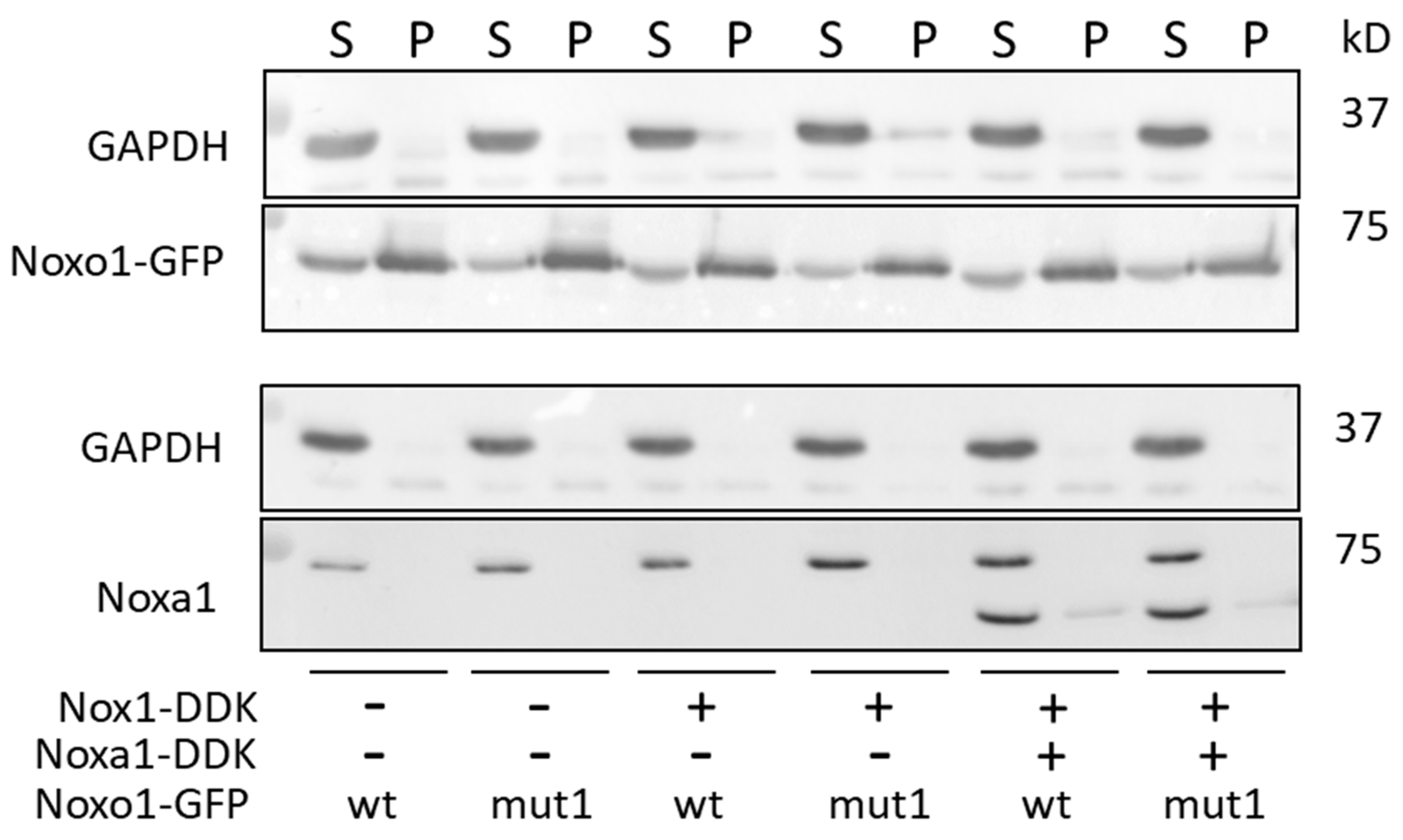
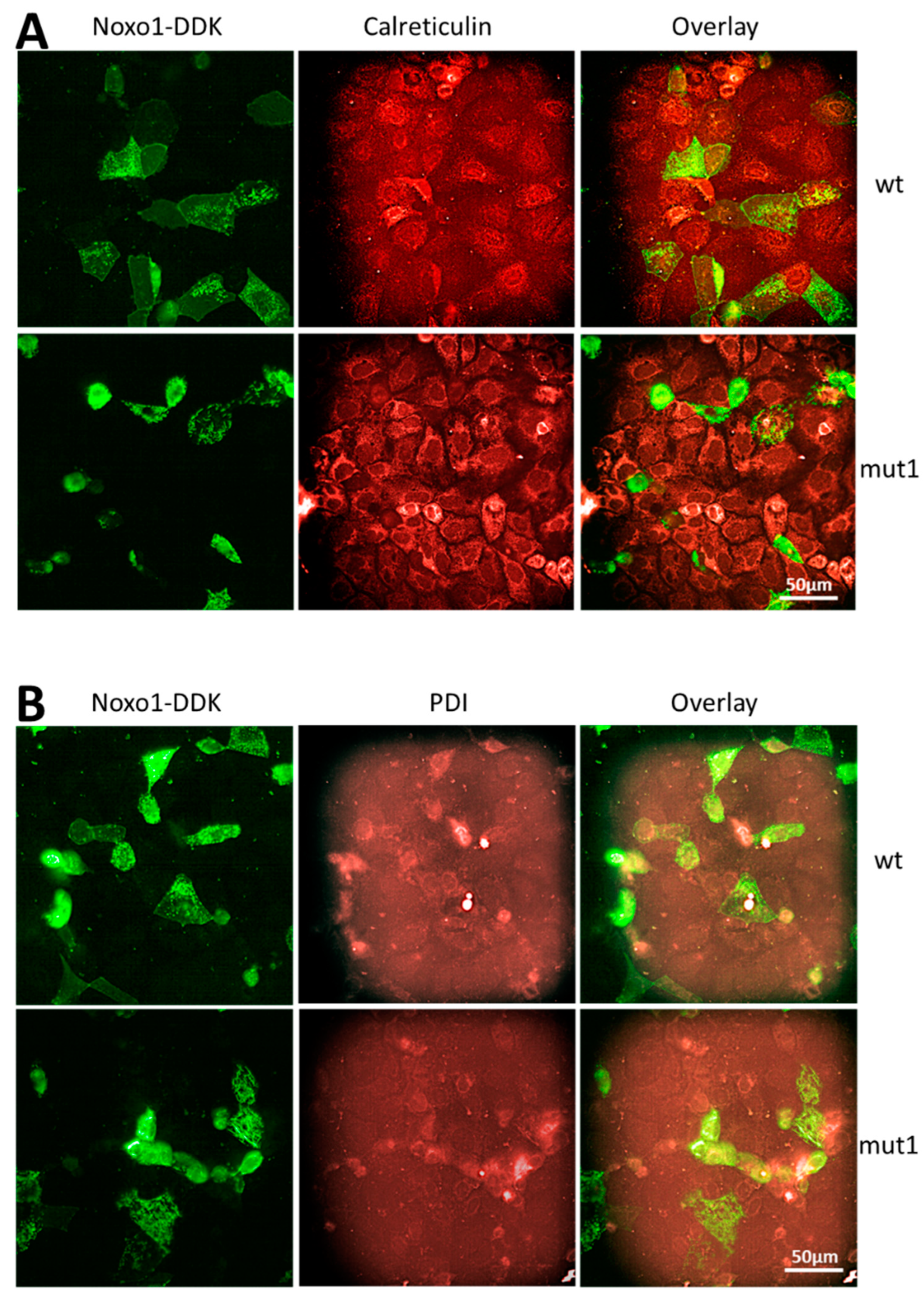
2.3. Mut1 D-Box Mutation Affects Noxa1/Noxo1 Interaction and Leads to a Translocation from the Membrane Compartment to a Cytoskeletal Insoluble Fraction Compared to wt Noxo1 in Caco-2 Cell Line
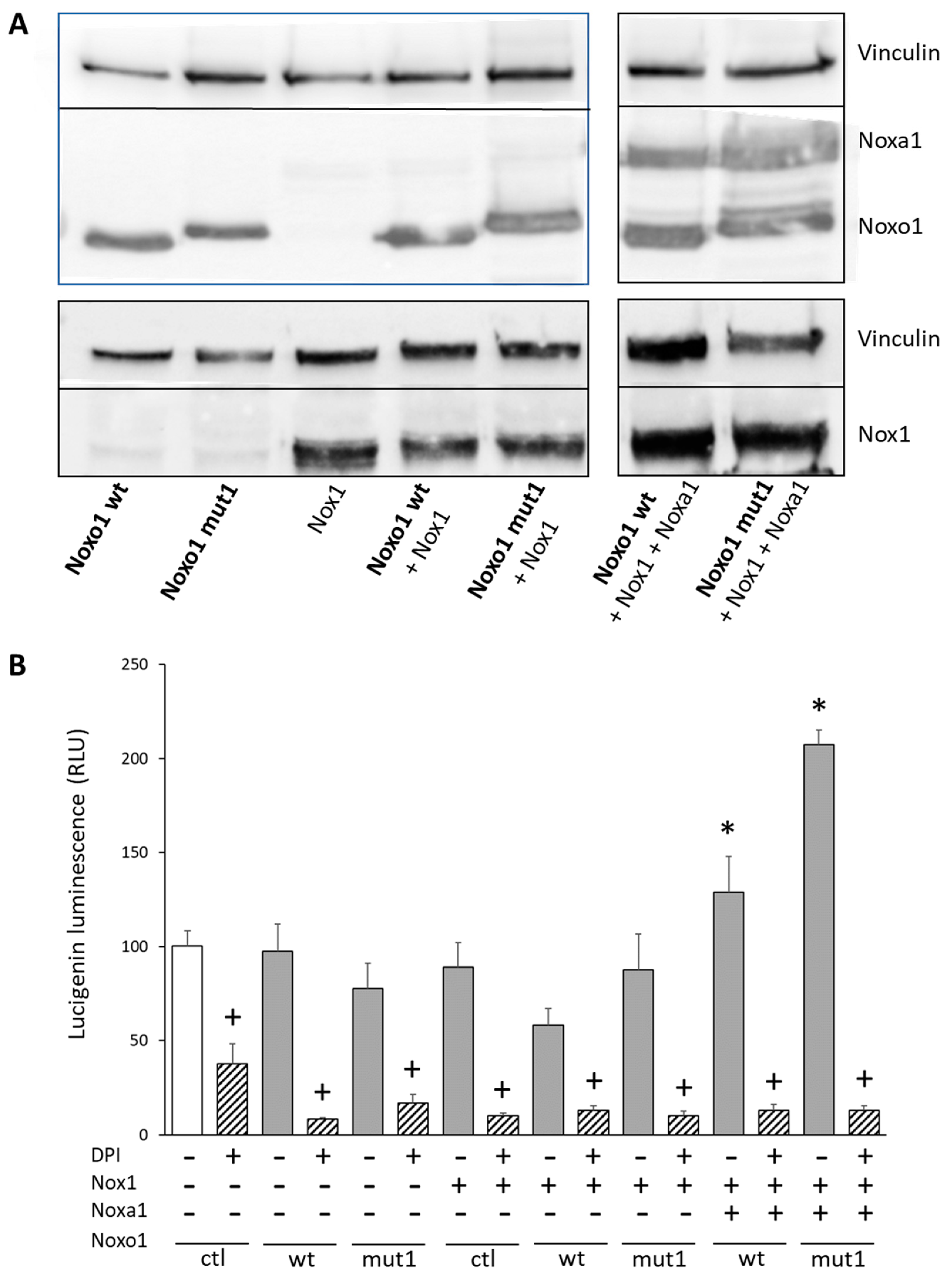
2.4. D-Box Mutation Mut1 in Noxo1 Increases ROS Production through Nox1-Dependant NADPH Oxidase
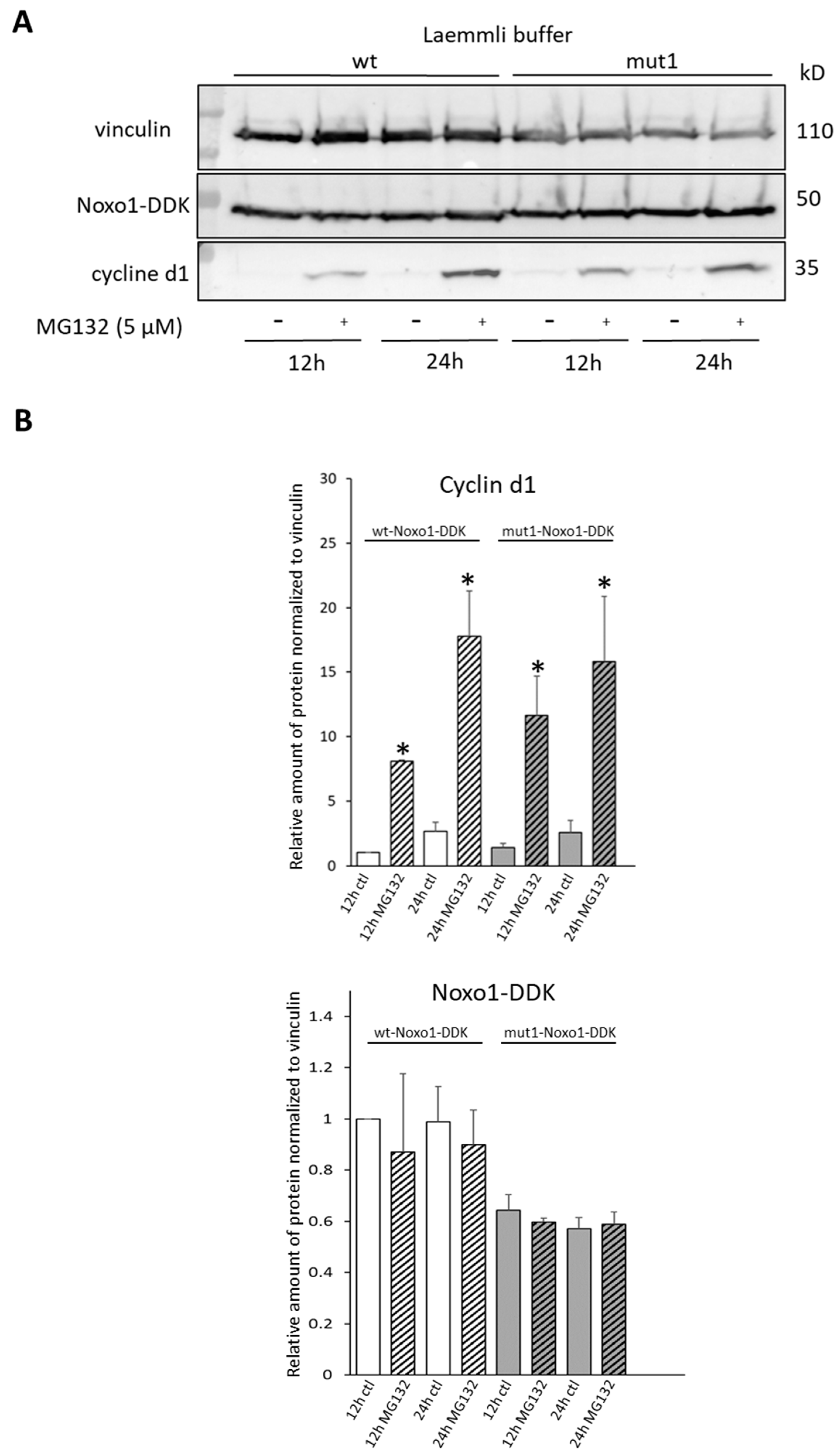
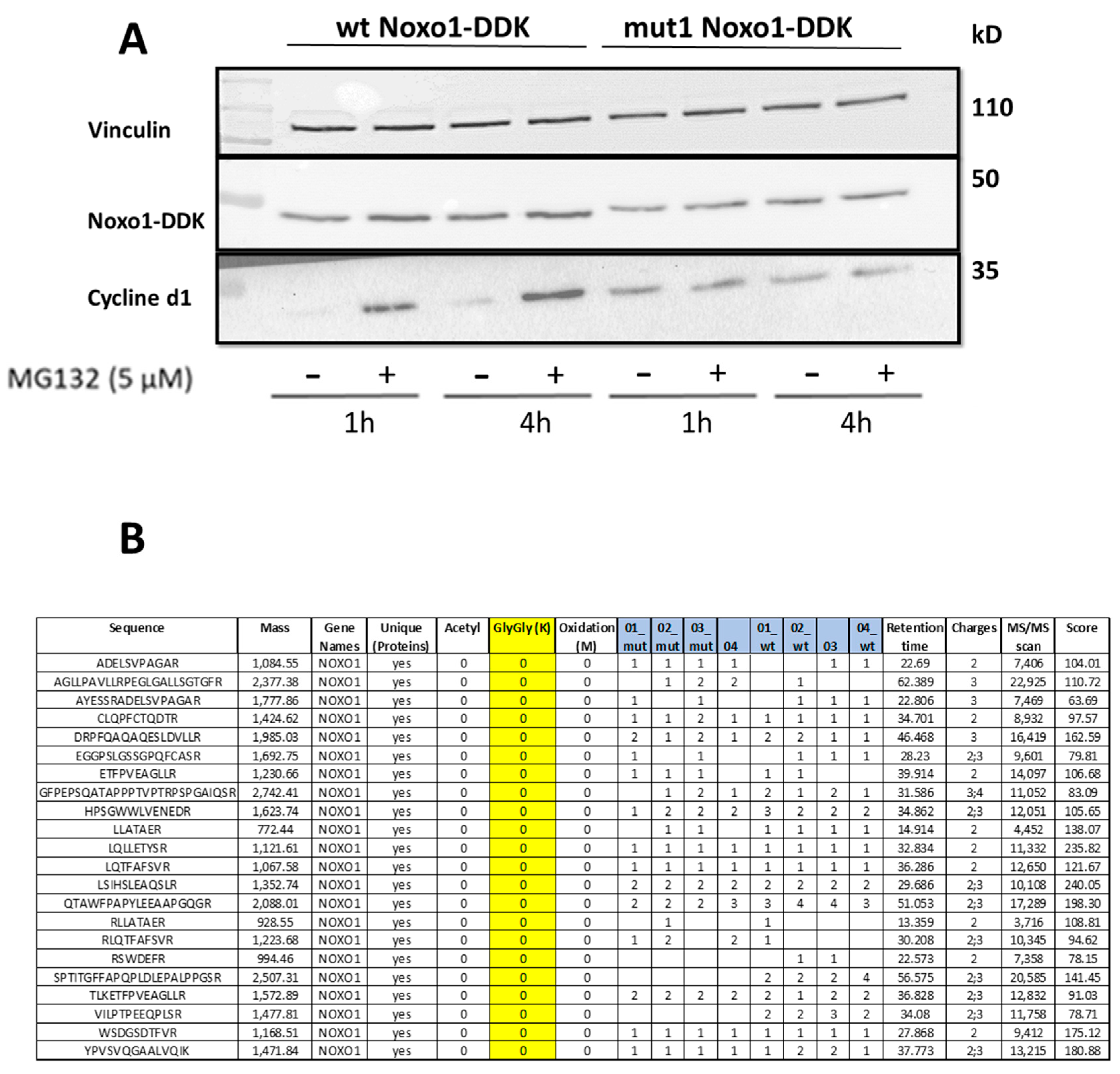
2.5. D-Box Mutation Mut1 in Noxo1 Does Not Stabilize Protein Turnover
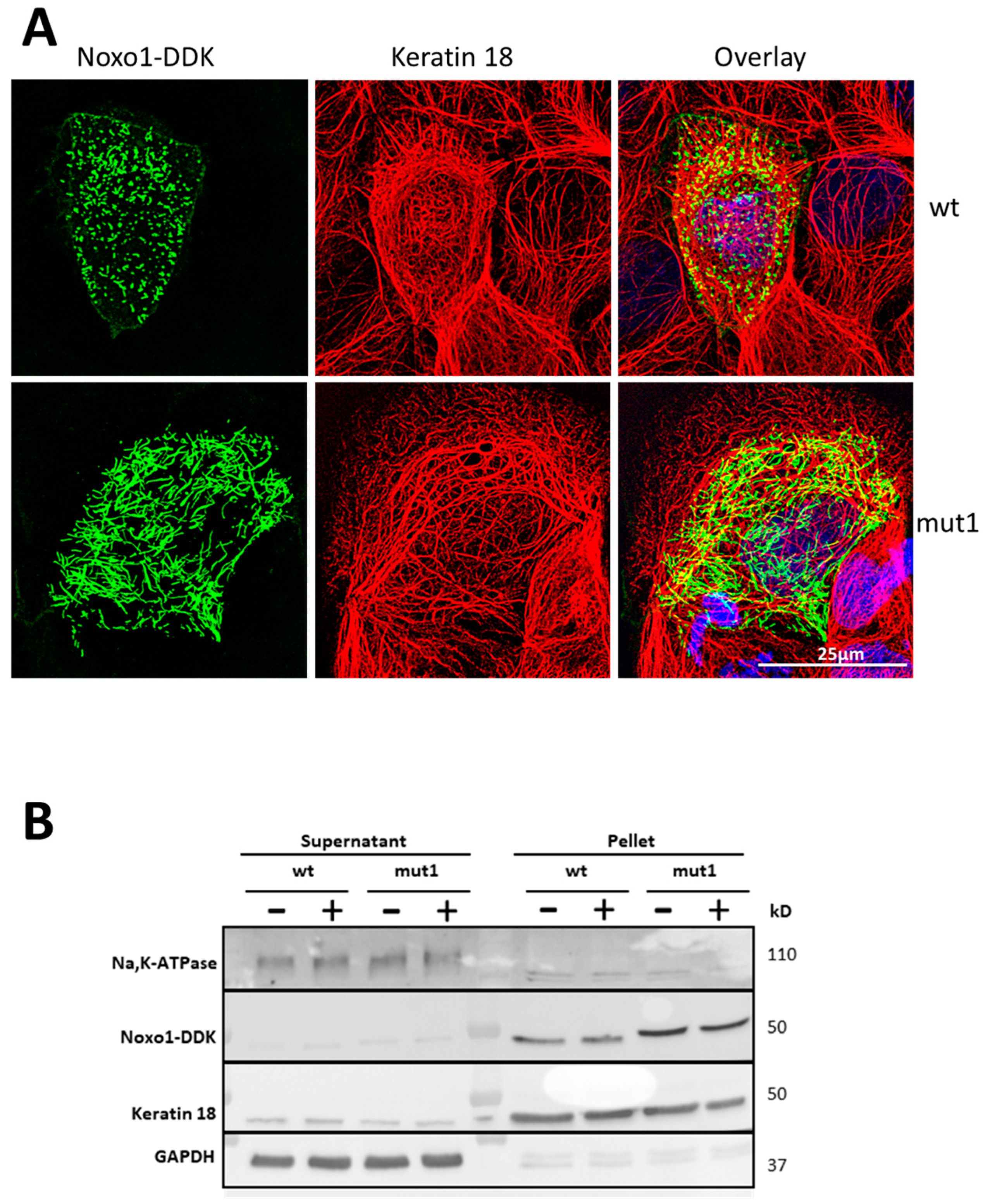
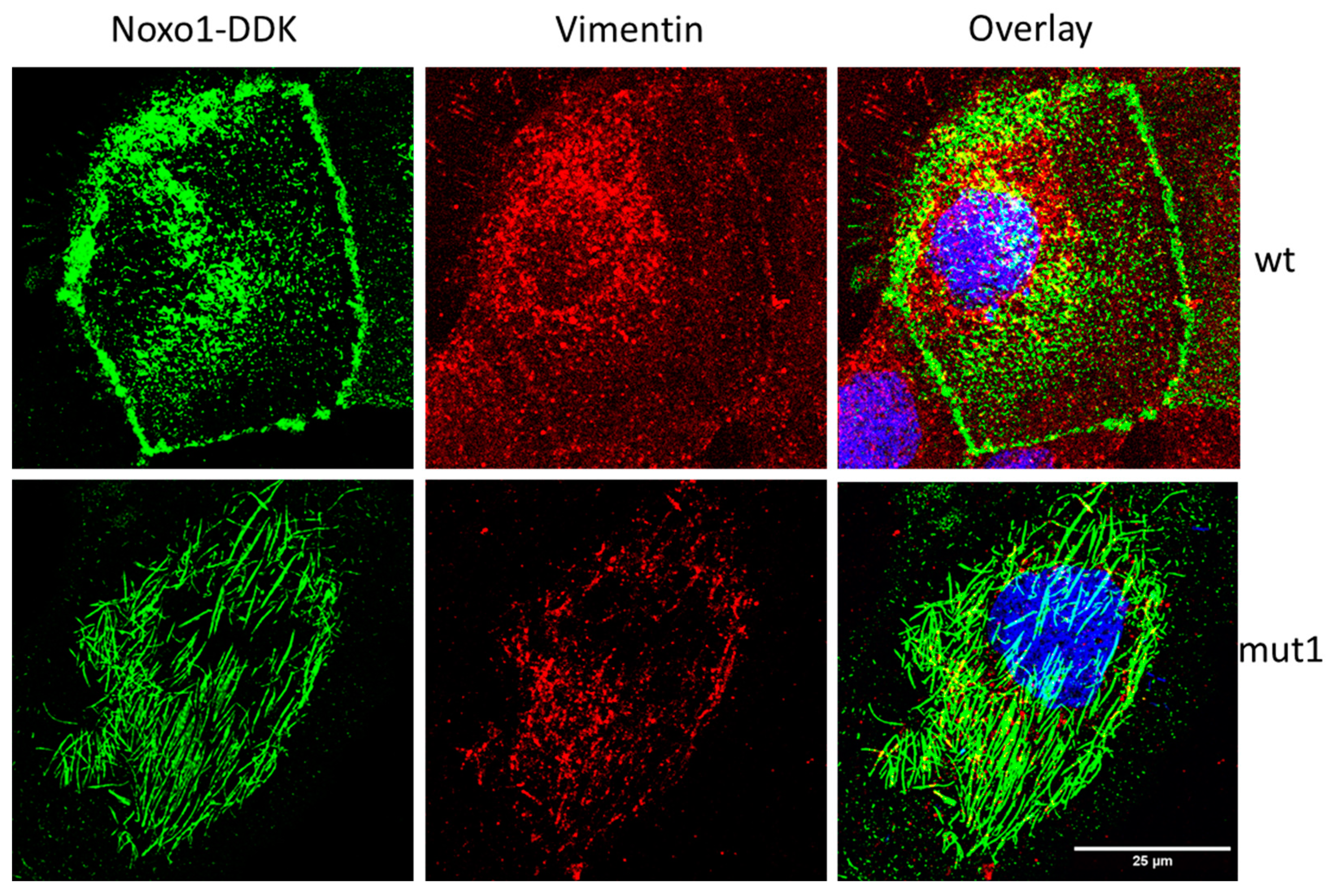
2.6. D-Box Mutation Mut1 in Noxo1 Leads to Association of Noxo1 with Intermediate Filaments
3. Discussion
4. Materials and Methods
4.1. Materials and Reagents
4.2. Cell Culture and Transfections
4.3. Subcellular Fractionation, Cell Extracts, and Immunoblot Analysis
4.4. Measurement of Superoxide Production
4.5. Immunofluorescence
4.6. Cytotoxicity Test and Blue Trypan Exclusion Test
4.7. Data and Statistical Analysis
4.8. High Content Screening (HCS) Data Analysis
4.9. Statistical Analysis
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
Appendix A




References
- Liu, X.; Hussain, R.; Mehmood, K.; Tang, Z.; Zhang, H.; Li, Y. Mitochondrial-Endoplasmic Reticulum Communication-Mediated Oxidative Stress and Autophagy. BioMed Res. Int. 2022, 2022, 6459585. [Google Scholar] [CrossRef]
- Chocry, M.; Leloup, L. The NADPH Oxidase Family and Its Inhibitors. Antioxid. Redox Signal. 2020, 33, 332–353. [Google Scholar] [CrossRef] [PubMed]
- Makhezer, N.; Ben Khemis, M.; Liu, D.; Khichane, Y.; Marzaioli, V.; Tlili, A.; Mojallali, M.; Pintard, C.; Letteron, P.; Hurtado-Nedelec, M.; et al. NOX1-Derived ROS Drive the Expression of Lipocalin-2 in Colonic Epithelial Cells in Inflammatory Conditions. Mucosal Immunol. 2019, 12, 117–131. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Begum, R.; Thota, S.; Abdulkadir, A.; Kaur, G.; Bagam, P.; Batra, S. NADPH Oxidase Family Proteins: Signaling Dynamics to Disease Management. Cell. Mol. Immunol. 2022, 19, 660–686. [Google Scholar] [CrossRef]
- Bedard, K.; Krause, K.-H. The NOX Family of ROS-Generating NADPH Oxidases: Physiology and Pathophysiology. Physiol. Rev. 2007, 87, 245–313. [Google Scholar] [CrossRef]
- Sylvester, A.L.; Zhang, D.X.; Ran, S.; Zinkevich, N.S. Inhibiting NADPH Oxidases to Target Vascular and Other Pathologies: An Update on Recent Experimental and Clinical Studies. Biomolecules 2022, 12, 823. [Google Scholar] [CrossRef]
- Liu, D.; Marie, J.-C.; Pelletier, A.-L.; Song, Z.; Ben-Khemis, M.; Boudiaf, K.; Pintard, C.; Leger, T.; Terrier, S.; Chevreux, G.; et al. Protein Kinase CK2 Acts as a Molecular Brake to Control NADPH Oxidase 1 Activation and Colon Inflammation. Cell. Mol. Gastroenterol. Hepatol. 2022, 13, 1073–1093. [Google Scholar] [CrossRef] [PubMed]
- Kim, J.-S.; Diebold, B.A.; Babior, B.M.; Knaus, U.G.; Bokoch, G.M. Regulation of Nox1 Activity via Protein Kinase A-Mediated Phosphorylation of NoxA1 and 14-3-3 Binding. J. Biol. Chem. 2007, 282, 34787–34800. [Google Scholar] [CrossRef] [Green Version]
- Kroviarski, Y.; Debbabi, M.; Bachoual, R.; Périanin, A.; Gougerot-Pocidalo, M.-A.; El-Benna, J.; Dang, P.M.-C. Phosphorylation of NADPH Oxidase Activator 1 (NOXA1) on Serine 282 by MAP Kinases and on Serine 172 by Protein Kinase C and Protein Kinase A Prevents NOX1 Hyperactivation. FASEB J. Off. Publ. Fed. Am. Soc. Exp. Biol. 2010, 24, 2077–2092. [Google Scholar] [CrossRef]
- Gianni, D.; Taulet, N.; DerMardirossian, C.; Bokoch, G.M. C-Src-Mediated Phosphorylation of NoxA1 and Tks4 Induces the Reactive Oxygen Species (ROS)-Dependent Formation of Functional Invadopodia in Human Colon Cancer Cells. Mol. Biol. Cell 2010, 21, 4287–4298. [Google Scholar] [CrossRef] [Green Version]
- Debbabi, M.; Kroviarski, Y.; Bournier, O.; Gougerot-Pocidalo, M.-A.; El-Benna, J.; Dang, P.M.-C. NOXO1 Phosphorylation on Serine 154 Is Critical for Optimal NADPH Oxidase 1 Assembly and Activation. FASEB J. Off. Publ. Fed. Am. Soc. Exp. Biol. 2013, 27, 1733–1748. [Google Scholar] [CrossRef] [PubMed]
- Yamamoto, A.; Takeya, R.; Matsumoto, M.; Nakayama, K.I.; Sumimoto, H. Phosphorylation of Noxo1 at Threonine 341 Regulates Its Interaction with Noxa1 and the Superoxide-Producing Activity of Nox1. FEBS J. 2013, 280, 5145–5159. [Google Scholar] [CrossRef] [PubMed]
- 1Jayaprakash, S.; Hegde, M.; BharathwajChetty, B.; Girisa, S.; Alqahtani, M.S.; Abbas, M.; Sethi, G.; Kunnumakkara, A.B. Unraveling the Potential Role of NEDD4-like E3 Ligases in Cancer. Int. J. Mol. Sci. 2022, 23, 12380. [Google Scholar] [CrossRef] [PubMed]
- Lee, J.; Lee, Y.; Jung, Y.M.; Park, J.H.; Yoo, H.S.; Park, J. Discovery of E3 Ligase Ligands for Target Protein Degradation. Molecules 2022, 27, 6515. [Google Scholar] [CrossRef] [PubMed]
- Okoye, C.N.; Rowling, P.J.E.; Itzhaki, L.S.; Lindon, C. Counting Degrons: Lessons From Multivalent Substrates for Targeted Protein Degradation. Front. Physiol. 2022, 13, 913063. [Google Scholar] [CrossRef] [PubMed]
- Kovacic, H.N.; Irani, K.; Goldschmidt-Clermont, P.J. Redox Regulation of Human Rac1 Stability by the Proteasome in Human Aortic Endothelial Cells. J. Biol. Chem. 2001, 276, 45856–45861. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Torrino, S.; Visvikis, O.; Doye, A.; Boyer, L.; Stefani, C.; Munro, P.; Bertoglio, J.; Gacon, G.; Mettouchi, A.; Lemichez, E. The E3 Ubiquitin-Ligase HACE1 Catalyzes the Ubiquitylation of Active Rac1. Dev. Cell 2011, 21, 959–965. [Google Scholar] [CrossRef] [Green Version]
- Haq, S.; Sarodaya, N.; Karapurkar, J.K.; Suresh, B.; Jo, J.K.; Singh, V.; Bae, Y.S.; Kim, K.-S.; Ramakrishna, S. CYLD Destabilizes NoxO1 Protein by Promoting Ubiquitination and Regulates Prostate Cancer Progression. Cancer Lett. 2022, 525, 146–157. [Google Scholar] [CrossRef]
- Joo, J.H.; Oh, H.; Kim, M.; An, E.J.; Kim, R.-K.; Lee, S.-Y.; Kang, D.H.; Kang, S.W.; Keun Park, C.; Kim, H.; et al. NADPH Oxidase 1 Activity and ROS Generation Are Regulated by Grb2/Cbl-Mediated Proteasomal Degradation of NoxO1 in Colon Cancer Cells. Cancer Res. 2016, 76, 855–865. [Google Scholar] [CrossRef] [Green Version]
- Yanagida, M.; Yamashita, Y.M.; Tatebe, H.; Ishii, K.; Kumada, K.; Nakaseko, Y. Control of Metaphase-Anaphase Progression by Proteolysis: Cyclosome Function Regulated by the Protein Kinase A Pathway, Ubiquitination and Localization. Philos. Trans. R. Soc. Lond. B Biol. Sci. 1999, 354, 1559–1569; discussion 1569–1570. [Google Scholar] [CrossRef] [Green Version]
- Barford, D. Structure, Function and Mechanism of the Anaphase Promoting Complex (APC/C). Q. Rev. Biophys. 2011, 44, 153–190. [Google Scholar] [CrossRef] [PubMed]
- Juhasz, A.; Ge, Y.; Markel, S.; Chiu, A.; Matsumoto, L.; van Balgooy, J.; Roy, K.; Doroshow, J.H. Expression of NADPH Oxidase Homologues and Accessory Genes in Human Cancer Cell Lines, Tumours and Adjacent Normal Tissues. Free Radic. Res. 2009, 43, 523–532. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ježek, P.; Jabůrek, M.; Porter, R.K. Uncoupling Mechanism and Redox Regulation of Mitochondrial Uncoupling Protein 1 (UCP1). Biochim. Biophys. Acta Bioenerg. 2019, 1860, 259–269. [Google Scholar] [CrossRef] [PubMed]
- Youle, R.J.; van der Bliek, A.M. Mitochondrial Fission, Fusion, and Stress. Science 2012, 337, 1062–1065. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Xu, X.; Liu, Y.; Luan, J.; Liu, R.; Wang, Y.; Liu, Y.; Xu, A.; Zhou, B.; Han, F.; Shang, W. Effect of Downregulated Citrate Synthase on Oxidative Phosphorylation Signaling Pathway in HEI-OC1 Cells. Proteome Sci. 2022, 20, 14. [Google Scholar] [CrossRef] [PubMed]
- Baghirova, S.; Hughes, B.G.; Hendzel, M.J.; Schulz, R. Sequential Fractionation and Isolation of Subcellular Proteins from Tissue or Cultured Cells. MethodsX 2015, 2, 440–445. [Google Scholar] [CrossRef]
- Kyriatzis, G.; Bernard, A.; Bôle, A.; Pflieger, G.; Chalas, P.; Masse, M.; Lécorché, P.; Jacquot, G.; Ferhat, L.; Khrestchatisky, M. Neurotensin Receptor 2 Is Induced in Astrocytes and Brain Endothelial Cells in Relation to Neuroinflammation Following Pilocarpine-Induced Seizures in Rats. Glia 2021, 69, 2618–2643. [Google Scholar] [CrossRef]
- Sbai, O.; Soussi, R.; Bole, A.; Khrestchatisky, M.; Esclapez, M.; Ferhat, L. The Actin Binding Protein α-Actinin-2 Expression Is Associated with Dendritic Spine Plasticity and Migrating Granule Cells in the Rat Dentate Gyrus Following Pilocarpine-Induced Seizures. Exp. Neurol. 2021, 335, 113512. [Google Scholar] [CrossRef]
- Van Dyke, R.W. Heterotrimeric G Protein Subunits Are Located on Rat Liver Endosomes. BMC Physiol. 2004, 4, 1. [Google Scholar] [CrossRef] [Green Version]
- Ushio-Fukai, M. Localizing NADPH Oxidase-Derived ROS. Sci. STKE Signal Transduct. Knowl. Environ. 2006, 2006, re8. [Google Scholar] [CrossRef]
- Sattayakhom, A.; Ittiwat, W.; Stremmel, W.; Chamulitrat, W. Redox Regulation of Cytokeratin 18 Protein by NADPH Oxidase 1 in Preneoplastic Human Epithelial Cells. J. Cancer Res. Clin. Oncol. 2011, 137, 1669–1678. [Google Scholar] [CrossRef] [PubMed]
- Altenhöfer, S.; Kleikers, P.W.M.; Radermacher, K.A.; Scheurer, P.; Rob Hermans, J.J.; Schiffers, P.; Ho, H.; Wingler, K.; Schmidt, H.H.H.W. The NOX Toolbox: Validating the Role of NADPH Oxidases in Physiology and Disease. Cell. Mol. Life Sci. 2012, 69, 2327–2343. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yamano, H.; Tsurumi, C.; Gannon, J.; Hunt, T. The Role of the Destruction Box and Its Neighbouring Lysine Residues in Cyclin B for Anaphase Ubiquitin-Dependent Proteolysis in Fission Yeast: Defining the D-Box Receptor. EMBO J. 1998, 17, 5670–5678. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cheng, G.; Lambeth, J.D. Alternative MRNA Splice Forms of NOXO1: Differential Tissue Expression and Regulation of Nox1 and Nox3. Gene 2005, 356, 118–126. [Google Scholar] [CrossRef] [PubMed]
- Ueyama, T.; Lekstrom, K.; Tsujibe, S.; Saito, N.; Leto, T.L. Subcellular Localization and Function of Alternatively Spliced Noxo1 Isoforms. Free Radic. Biol. Med. 2007, 42, 180–190. [Google Scholar] [CrossRef] [Green Version]
- Zhang, J.; Hu, S.; Li, Y. KRT18 Is Correlated with the Malignant Status and Acts as an Oncogene in Colorectal Cancer. Biosci. Rep. 2019, 39, BSR20190884. [Google Scholar] [CrossRef] [Green Version]
- Hilenski, L.L.; Clempus, R.E.; Quinn, M.T.; Lambeth, J.D.; Griendling, K.K. Distinct Subcellular Localizations of Nox1 and Nox4 in Vascular Smooth Muscle Cells. Arterioscler. Thromb. Vasc. Biol. 2004, 24, 677–683. [Google Scholar] [CrossRef]
- Schindelin, J.; Arganda-Carreras, I.; Frise, E.; Kaynig, V.; Longair, M.; Pietzsch, T.; Preibisch, S.; Rueden, C.; Saalfeld, S.; Schmid, B.; et al. Fiji: An open-source platform for biological-image analysis. Nat. Methods 2012, 9, 676–682. [Google Scholar] [CrossRef] [Green Version]












{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Antibodies | Species | Reference; Supplier | Dilution for | |
|---|---|---|---|---|
| IF | WB | |||
| Anti-DDK | Mouse | TA50011-100; Origene Technologies | 1/400 | 1/1000 |
| Anti-DDK | Rabbit | D6w5b mab; Cell signaling | 1/400 | 1/1000 |
| Anti-vinculin | Mouse | V9264; Sigma Aldrich | 1/200 | 1/5000 |
| Anti-Noxa1 | Mouse | SC398873; SantaCruz Biotechnologies | 1/50 | 1/500 |
| Anti-Nox1 | Goat | Ab121009; Abcam | 1/5000 | |
| Anti-cyclin d1 | Rabbit | 2978; Cell signaling | 1/500 | |
| Phalloidin | P1951; Sigma Aldrich | 1/2000 | ||
| Anti-calreticuline | Mouse | ADISPA601; Enzo Life Sciences | 1/200 | |
| Anti-PDI | Mouse | ADISPA891; Enzo Life Sciences | 1/400 | |
| Anti-Vimentin | Rabbit | R28 3932; Cell Signaling | 2/200 | |
| Anti Alpha-tubulin | Rabbit | Ab18251; Abcam | 1/800 | |
| Anti-Keratin kit | Rabbit and Mouse | 9384; Cell signaling | ||
| Anti Citrate synthase | Mouse | SC390693; SantaCruz Biotechnologies | 1/400 | |
| Anti-GAD 67 | Mouse | MAB5406; Sigma Aldrich | 1/500 | |
| Anti-Synaptophysin | Mouse | 01011; Synaptic Systems | 1/500 | |
| Anti-GFP | Rabbit | SC9996; SantaCruz Biotechnologies | 1/500 | |
| Anti-GAPDH | Mouse | G8795; Sigma Aldrich | 1/5000 | |
| Anti-Keratin 18 | Mouse | Keratin 18 (DC10), Cell Signaling | 1/800 | 1/2000 |
| Anti-Na,K-ATPase | Mouse | Ab7671; Abcam | 1/100 | 1/2000 |
| Alexa Fluor 488 anti-mouse | Goat | A-11029; Invitrogen | 1/800 | |
| Alexa Fluor 546 anti-rabbit | Goat | A-11010; Invitrogen | 1/800 | |
| Alexa Fluor 594 anti-rabbit | Goat | Ab15008; Abcam, | 1/800 | |
| Alexa Fluor 647 anti-mouse | Goat | Ab150115; Abcam | 1/800 | |
| HRP anti-mouse | Horse | 7076S; Cell signaling | 1/5000 | |
| HRP anti-rabbit | Goat | 7074S; Cell signaling | 1/2500 | |
| HRP anti-goat | Donkey | IR 705-035-003; Jackson Immunoresearch | 1/2500 | |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Benssouina, F.-Z.; Parat, F.; Villard, C.; Leloup, L.; Garrouste, F.; Sabatier, J.-m.; Ferhat, L.; Kovacic, H. Overexpression of a Novel Noxo1 Mutant Increases Ros Production and Noxo1 Relocalisation. Int. J. Mol. Sci. 2023, 24, 4663. https://doi.org/10.3390/ijms24054663
Benssouina F-Z, Parat F, Villard C, Leloup L, Garrouste F, Sabatier J-m, Ferhat L, Kovacic H. Overexpression of a Novel Noxo1 Mutant Increases Ros Production and Noxo1 Relocalisation. International Journal of Molecular Sciences. 2023; 24(5):4663. https://doi.org/10.3390/ijms24054663
Chicago/Turabian StyleBenssouina, Fatima-Zahra, Fabrice Parat, Claude Villard, Ludovic Leloup, Françoise Garrouste, Jean-marc Sabatier, Lotfi Ferhat, and Hervé Kovacic. 2023. "Overexpression of a Novel Noxo1 Mutant Increases Ros Production and Noxo1 Relocalisation" International Journal of Molecular Sciences 24, no. 5: 4663. https://doi.org/10.3390/ijms24054663