
Heat-Killed Enterococcus faecalis Inhibit FL83B Hepatic Lipid Accumulation and High Fat Diet-Induced Fatty Liver Damage in Rats by Activating Lipolysis through the Regulation the AMPK Signaling Pathway

 ,
, {kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results
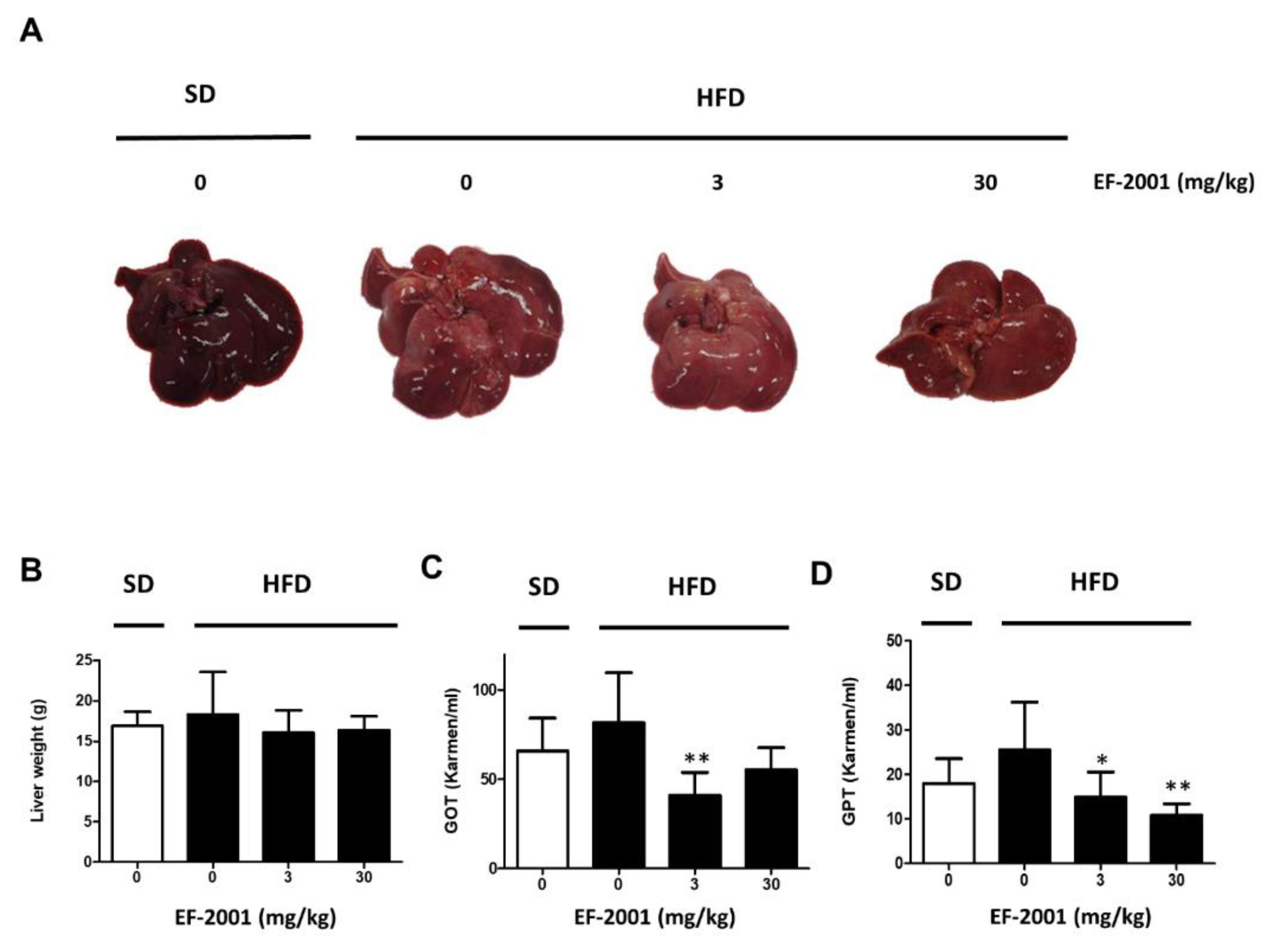
2.1. EF-2001 Intake Effectively Prevents Fatty Liver Tissue and Liver Damage in HFD-Induced Rats
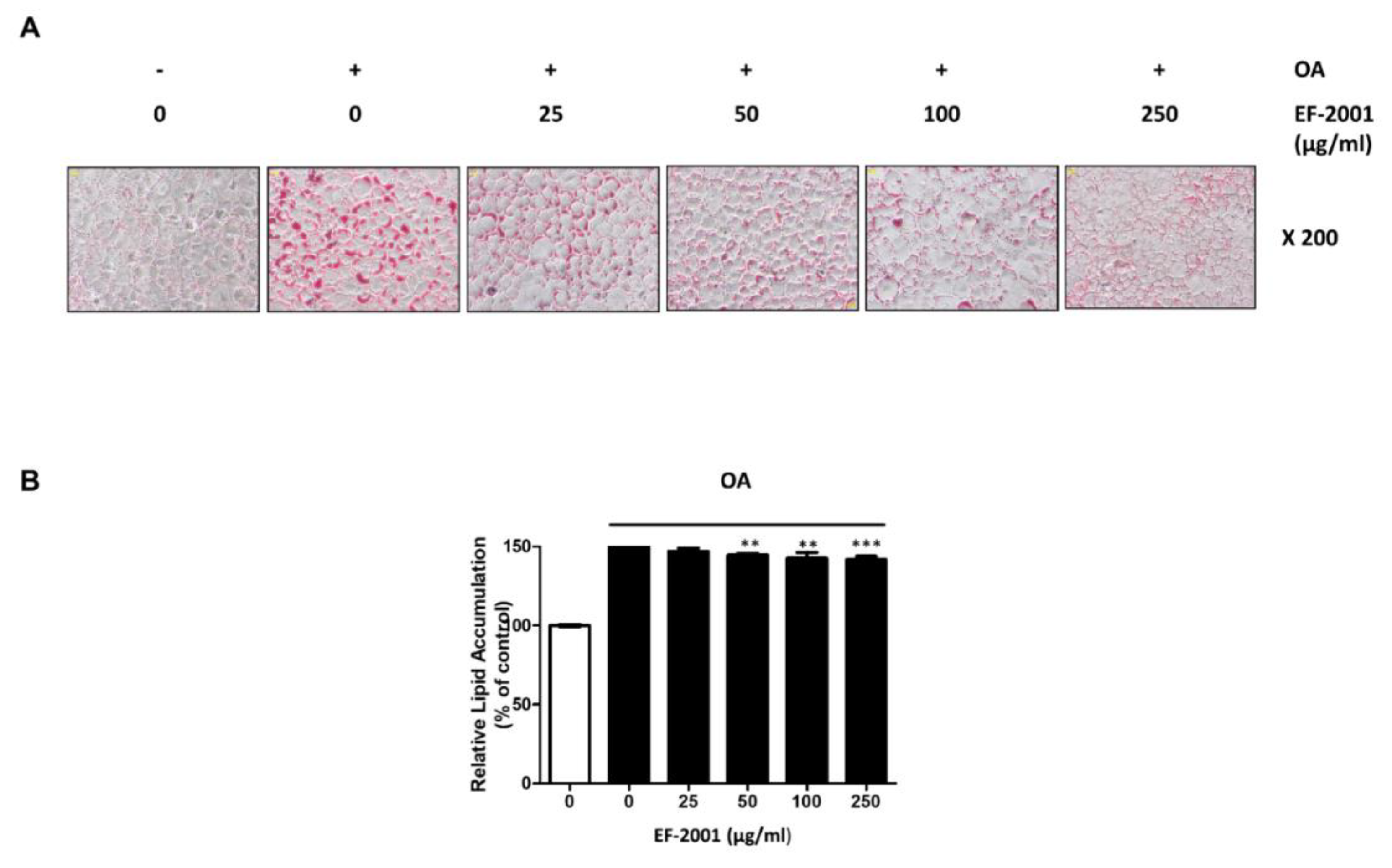
2.2. Effect of EF-2001 on Oleic Acid-Induced Hepatic Lipid Accumulation in FL83Bs
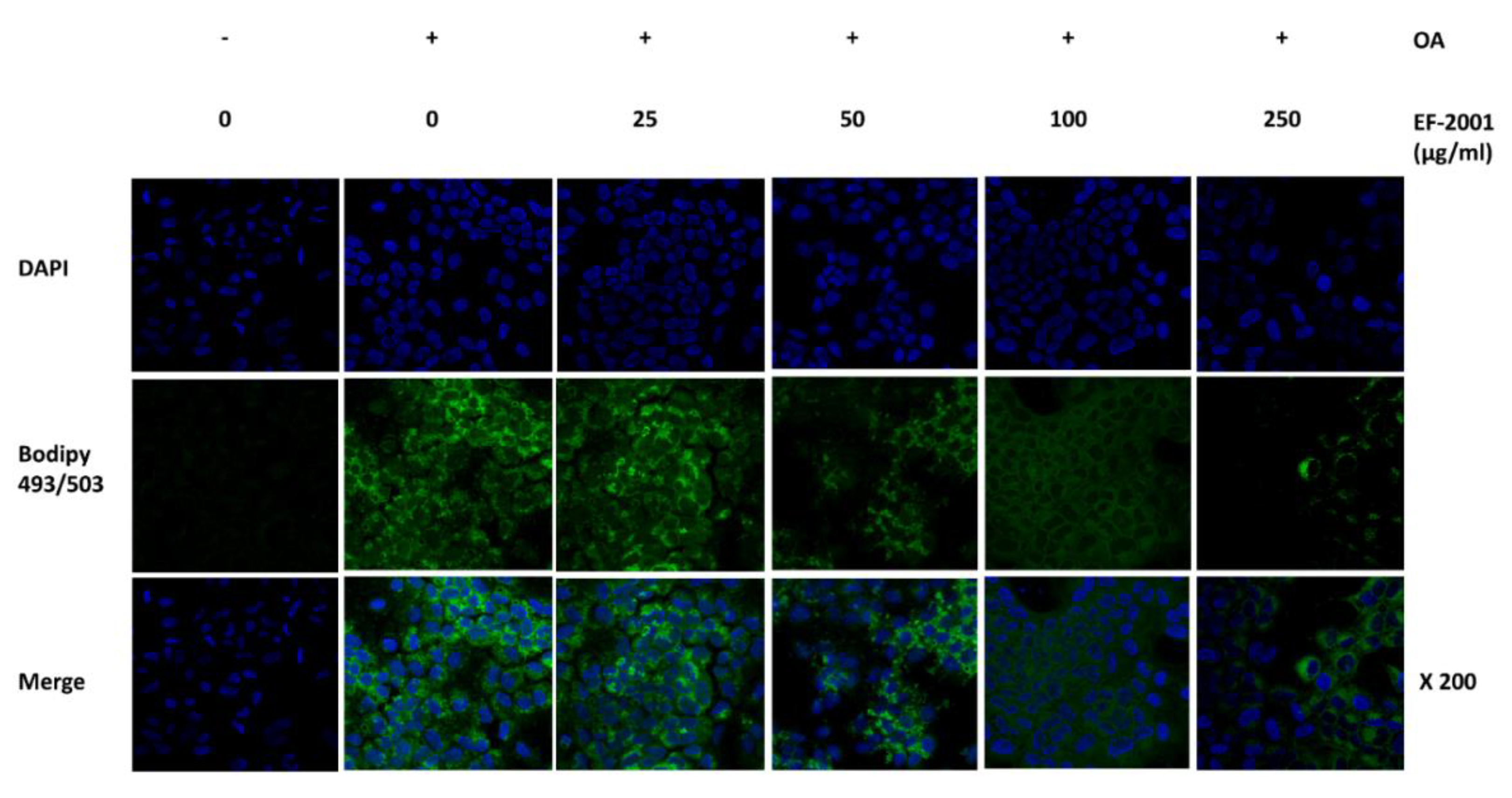
2.3. Effect of EF-2001 on Neutral Lipid Droplet of Oleic Acid-Induced FL83Bs Hepatic Lipid Accumulation
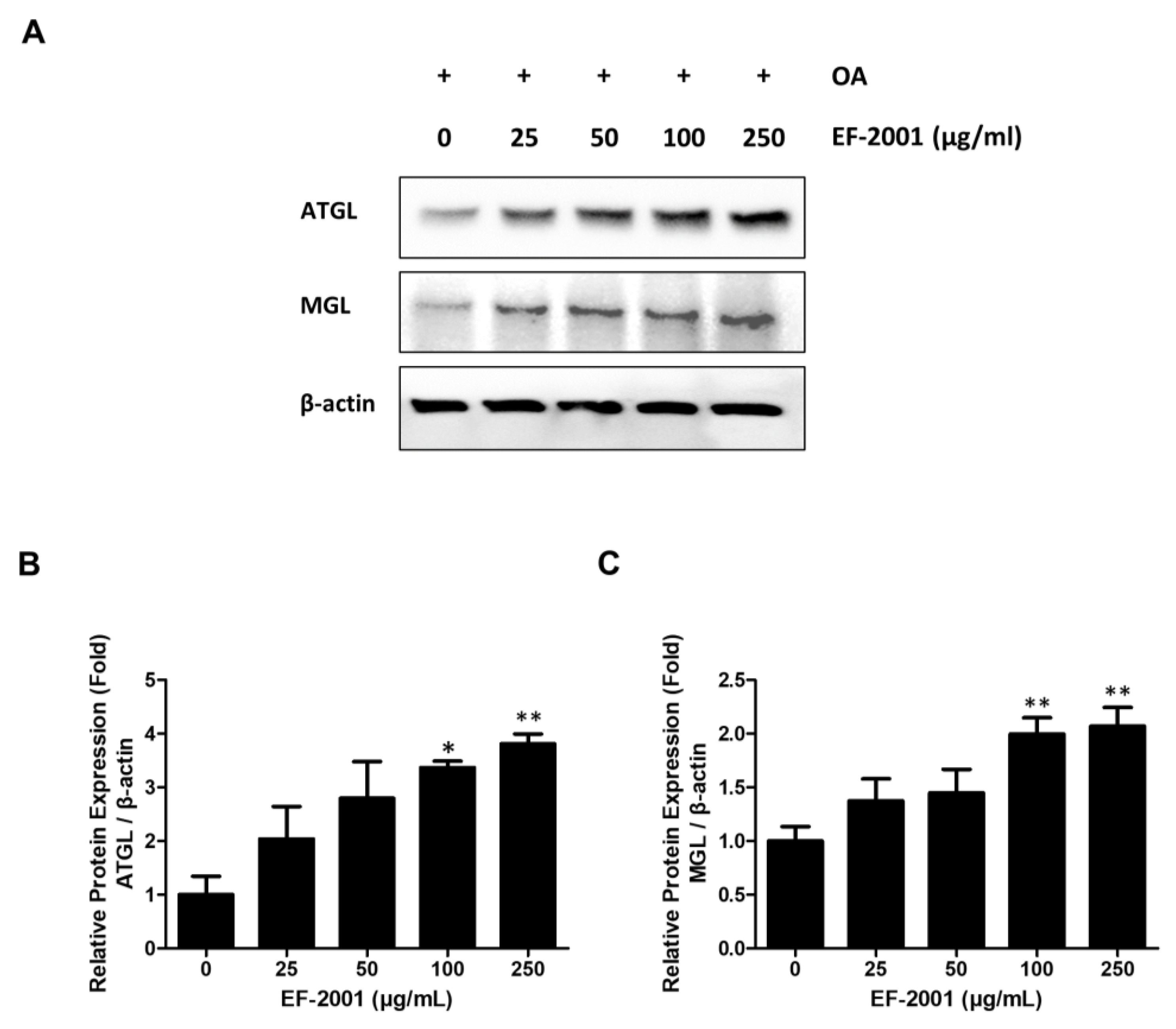
2.4. Effects of EF-2001 on the Expression of Lipase Enzyme Protein in FL83Bs
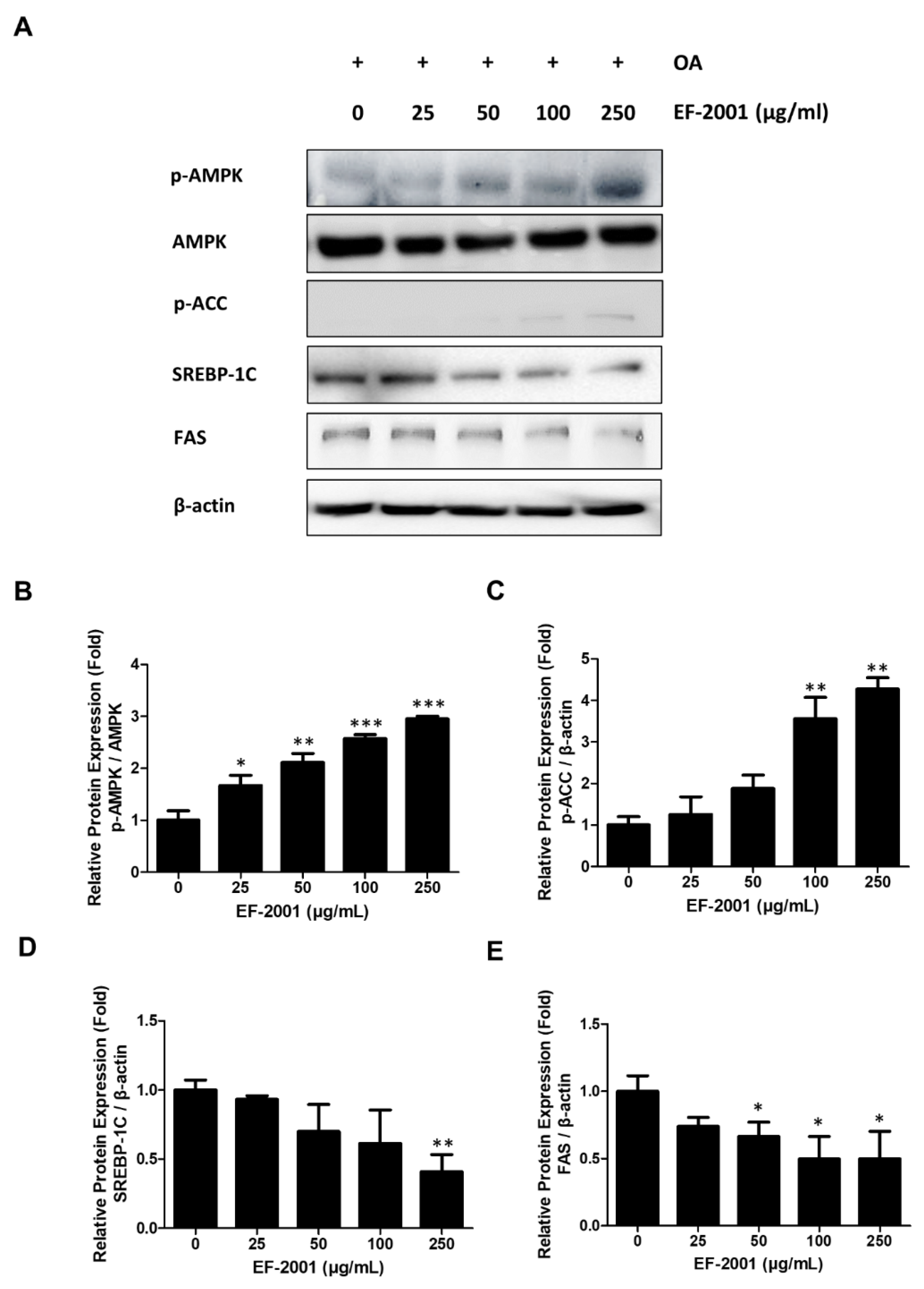
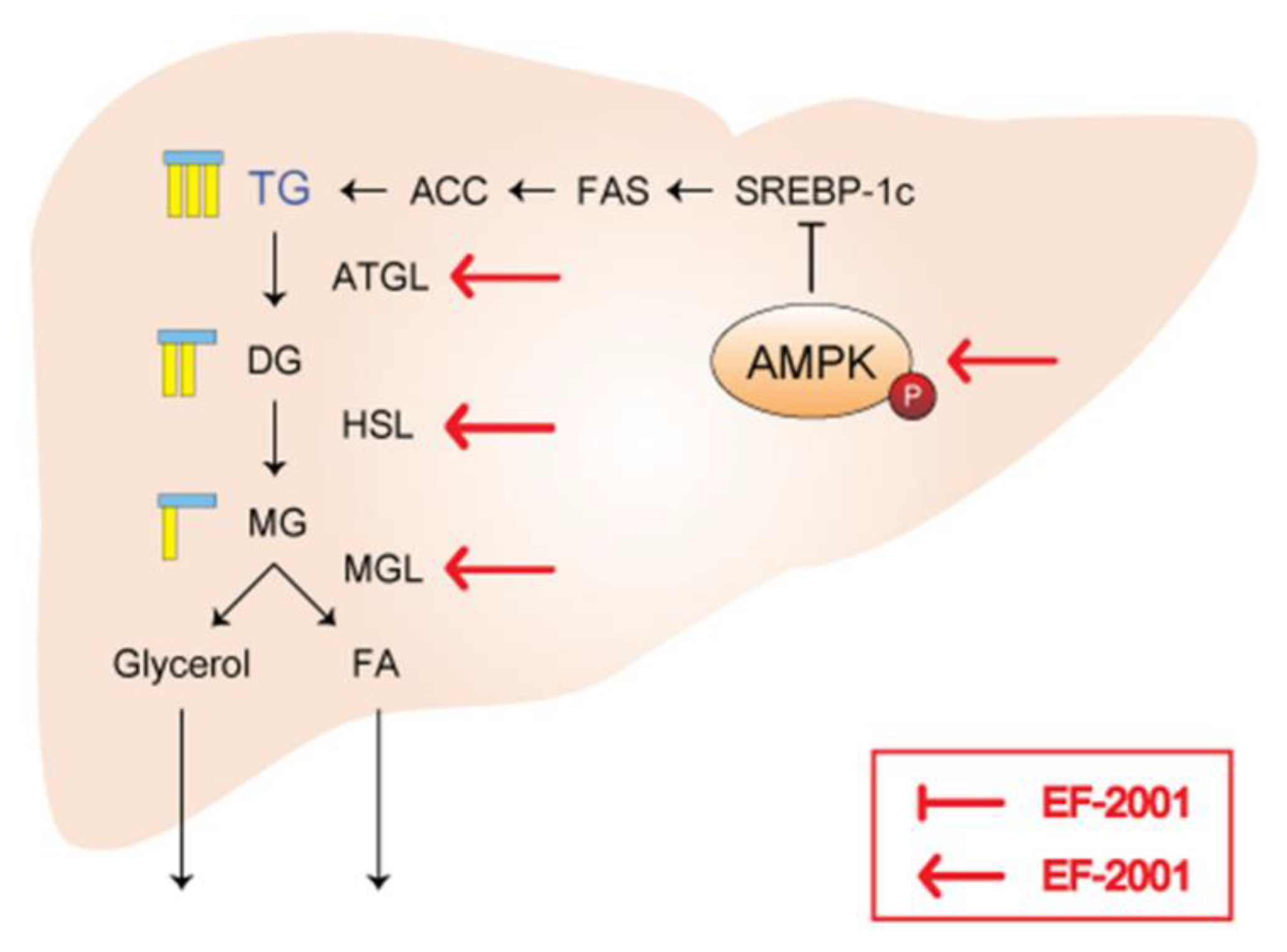
2.5. Effects of EF-2001 on the Expression of AMPK and SREBP Signaling Pathway
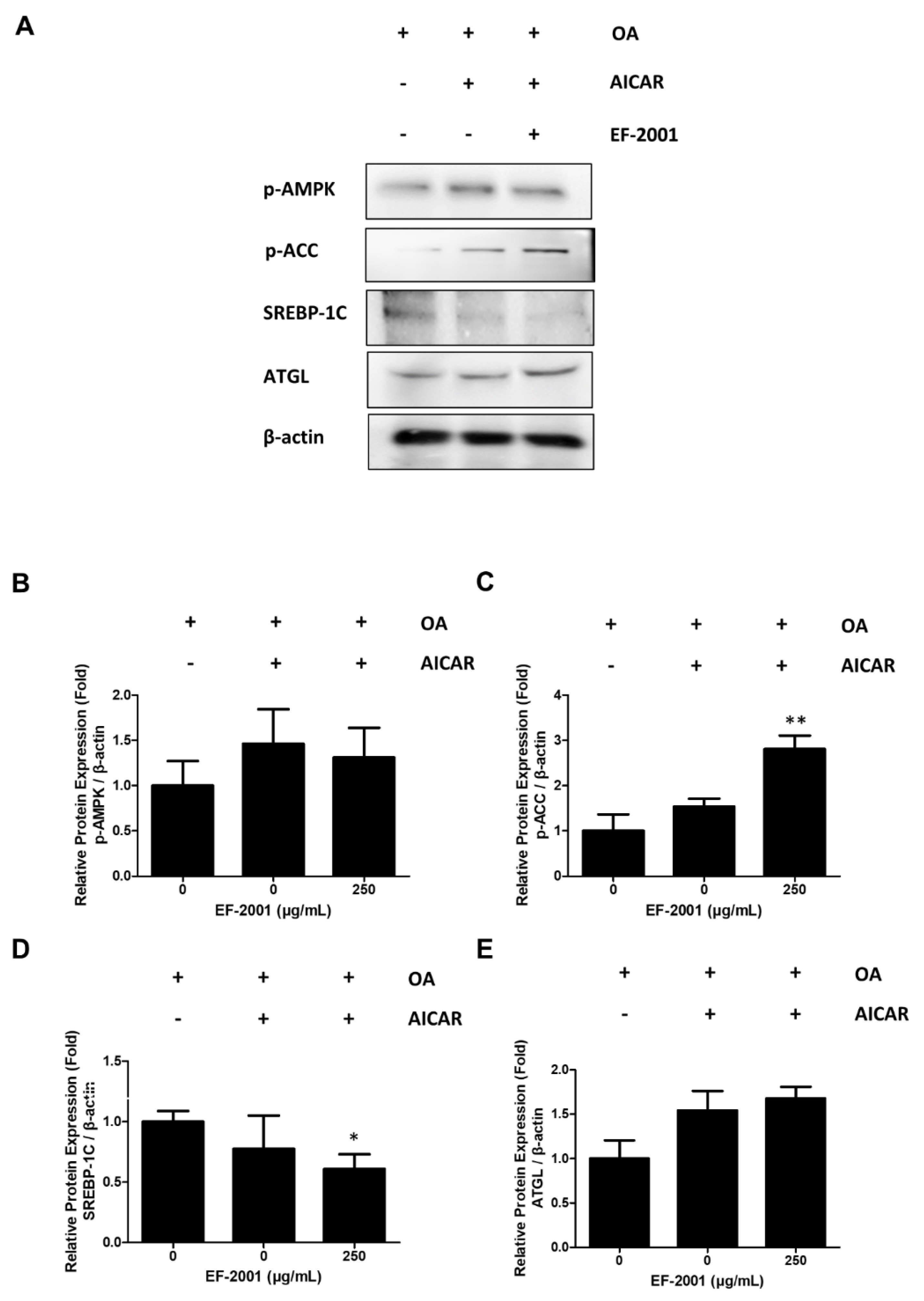
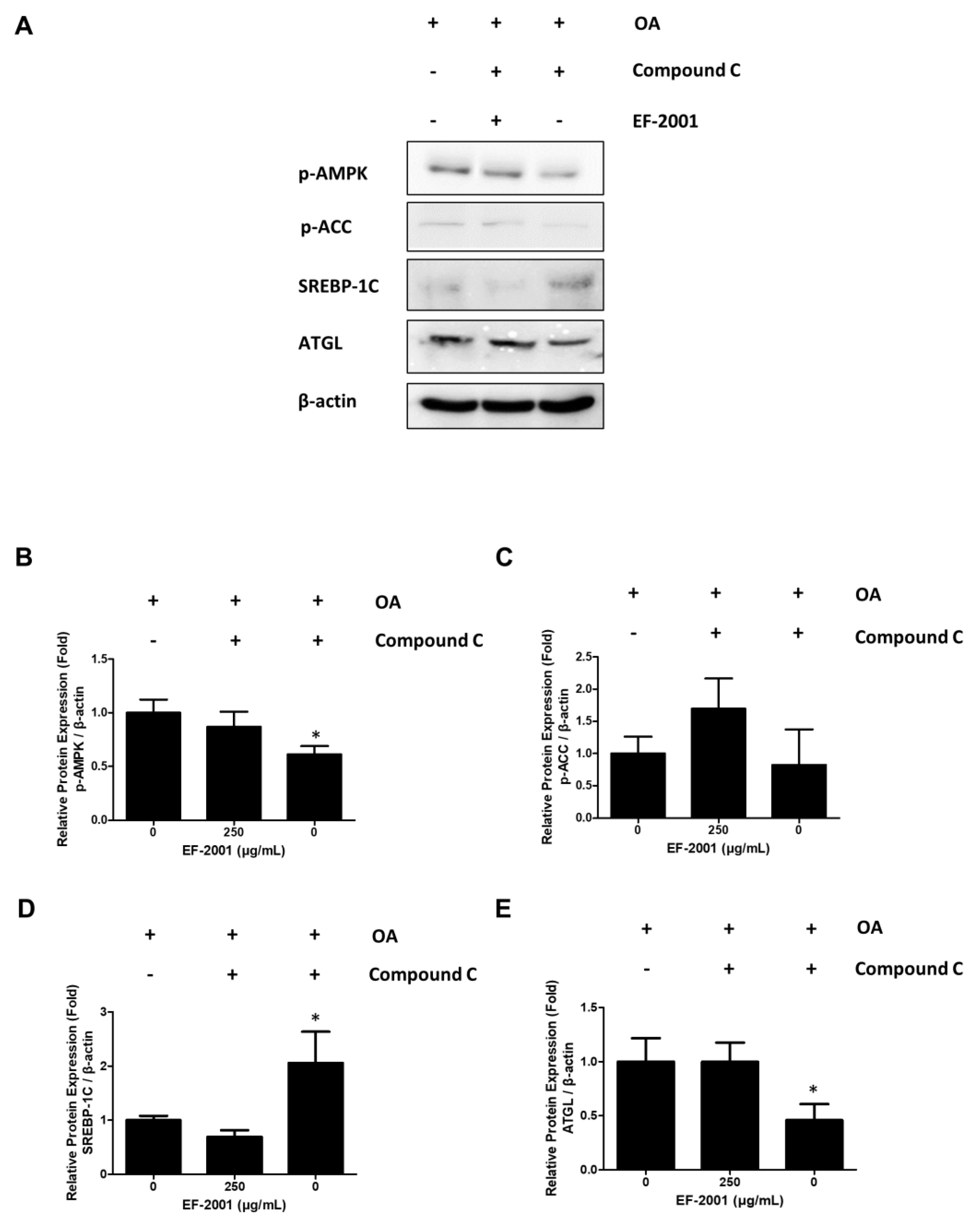
2.6. Effects of EF-2001 on AMPK Targeted Signaling Pathway
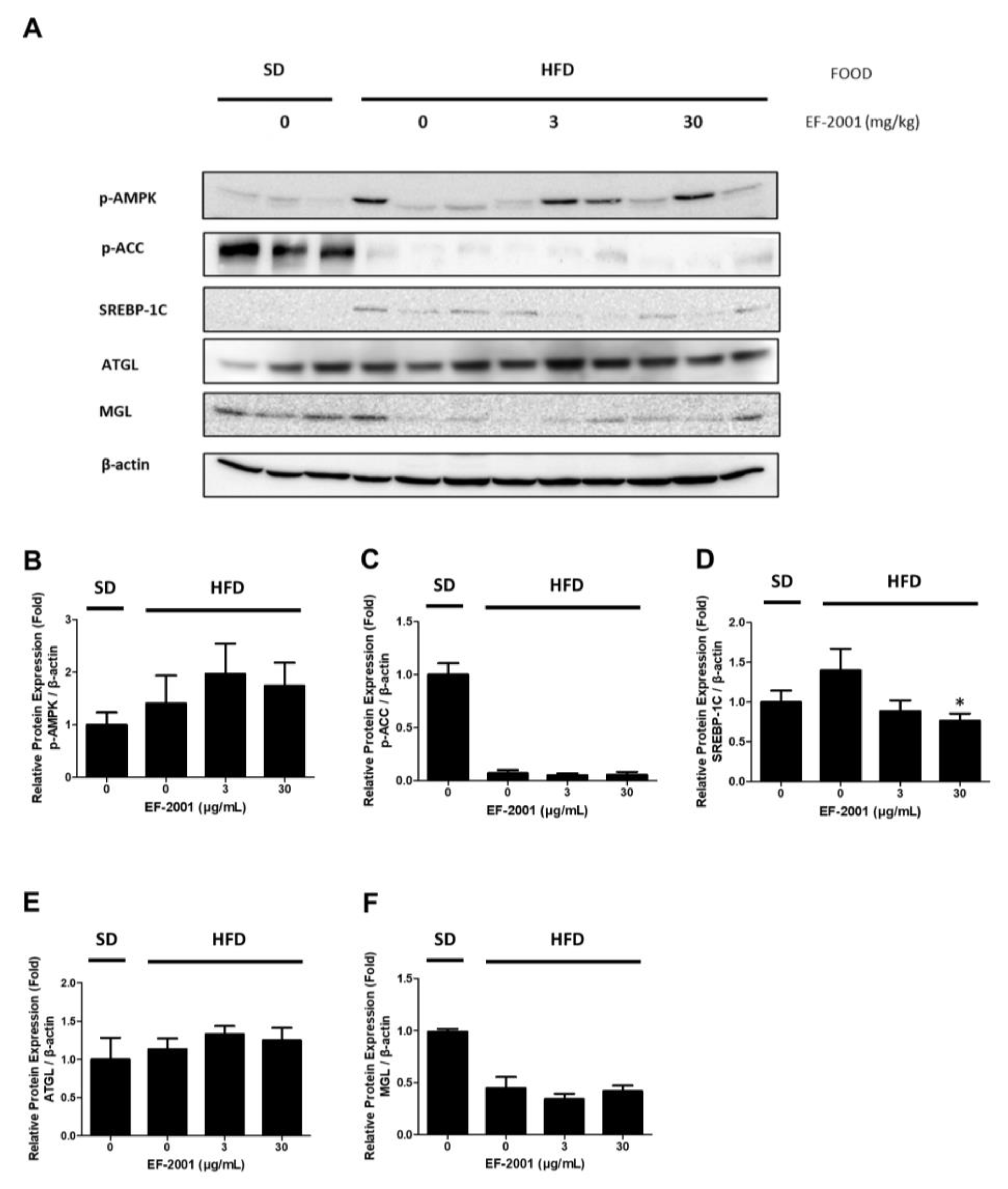
2.7. Effects of EF-2001 AMPK Signaling Pathway on HFD Induced Fatty Liver
3. Discussion
4. Materials and Methods
4.1. Preparation of Heat-Killed Enterococcus faecalis (EF-2001)
4.2. Chemical Reagent
4.3. Animal Experiments
4.4. Serological Analysis
4.5. Cell Culture and Induced Fatty Liver Cells
4.6. Oil Red O Staining of FL83B Hepatocyte
4.7. Western Blot Analysis
4.8. Confocal Microscopy
4.9. Statistical Analysis
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Chang, H.-H.; Moro, A.; Takakura, K.; Su, H.-Y.; Mo, A.; Nakanishi, M.; Waldron, R.T.; French, S.W.; Dawson, D.W.; Hines, O.J. Incidence of pancreatic cancer is dramatically increased by a high fat, high calorie diet in KrasG12D mice. PLoS ONE 2017, 12, e0184455. [Google Scholar] [CrossRef] [PubMed]
- Kim, J.H.; Moon, J.S. Secular trends in pediatric overweight and obesity in Korea. J. Obes. Metab. Syndr. 2020, 29, 12. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yang, H.K.; Han, K.; Cho, J.-H.; Yoon, K.-H.; Cha, B.-Y.; Lee, S.-H. Ambient temperature and prevalence of obesity: A nationwide population-based study in Korea. PLoS ONE 2015, 10, e0141724. [Google Scholar] [CrossRef] [PubMed]
- Qiao, Y.; Sun, J.; Xia, S.; Li, L.; Li, Y.; Wang, P.; Shi, Y.; Le, G. Effects of different Lactobacillus reuteri on inflammatory and fat storage in high-fat diet-induced obesity mice model. J. Funct. Foods 2015, 14, 424–434. [Google Scholar] [CrossRef]
- Ducharme, N.A.; Bickel, P.E. Minireview: Lipid droplets in lipogenesis and lipolysis. Endocrinology 2008, 149, 942–949. [Google Scholar] [CrossRef] [Green Version]
- Lehrskov, L.L.; Christensen, R.H. The role of interleukin-6 in glucose homeostasis and lipid metabolism. In Seminars in Immunopathology; Springer: Berlin/Heidelberg, Germany, 2019; pp. 491–499. [Google Scholar]
- Bardini, G.; Rotella, C.M.; Giannini, S. Dyslipidemia and diabetes: Reciprocal impact of impaired lipid metabolism and Beta-cell dysfunction on micro-and macrovascular complications. Rev. Diabet. Stud. RDS 2012, 9, 82. [Google Scholar] [CrossRef]
- Shin, M.-R.; Shin, S.H.; Roh, S.-S. Diospyros kaki and Citrus unshiu mixture improves disorders of lipid metabolism in nonalcoholic fatty liver disease. Can. J. Gastroenterol. Hepatol. 2020, 2020, 8812634. [Google Scholar] [CrossRef]
- Qiu, X.; Ye, Q.; Sun, M.; Wang, L.; Tan, Y.; Wu, G. Saturated hydrogen improves lipid metabolism disorders and dysbacteriosis induced by a high-fat diet. Exp. Biol. Med. 2020, 245, 512–521. [Google Scholar] [CrossRef]
- Targher, G.; Corey, K.E.; Byrne, C.D.; Roden, M. The complex link between NAFLD and type 2 diabetes mellitus—Mechanisms and treatments. Nat. Rev. Gastroenterol. Hepatol. 2021, 18, 599–612. [Google Scholar] [CrossRef]
- Rodriguez, L.A.; Kanaya, A.M.; Shiboski, S.C.; Fernandez, A.; Herrington, D.; Ding, J.; Bradshaw, P.T. Does NAFLD mediate the relationship between obesity and type 2 diabetes risk? evidence from the multi-ethnic study of atherosclerosis (MESA). Ann. Epidemiol. 2021, 63, 15–21. [Google Scholar] [CrossRef]
- Huang, W.-C.; Liao, P.-C.; Huang, C.-H.; Hu, S.; Huang, S.-C.; Wu, S.-J. Osthole attenuates lipid accumulation, regulates the expression of inflammatory mediators, and increases antioxidants in FL83B cells. Biomed. Pharmacother. 2017, 91, 78–87. [Google Scholar] [CrossRef]
- Lee, H.; Lee, M.; Kim, H.S. Effects of Micro-current stimulation on lipid metabolism in oleic acid-induced non-alcoholic fatty liver disease in FL83B cells. J. Biomed. Eng. Res. 2022, 43, 1–10. [Google Scholar]
- Huang, W.-C.; Chen, Y.-L.; Liu, H.-C.; Wu, S.-J.; Liou, C.-J. Ginkgolide C reduced oleic acid-induced lipid accumulation in HepG2 cells. Saudi Pharm. J. 2018, 26, 1178–1184. [Google Scholar] [CrossRef] [PubMed]
- Ricchi, M.; Odoardi, M.R.; Carulli, L.; Anzivino, C.; Ballestri, S.; Pinetti, A.; Fantoni, L.I.; Marra, F.; Bertolotti, M.; Banni, S. Differential effect of oleic and palmitic acid on lipid accumulation and apoptosis in cultured hepatocytes. J. Gastroenterol. Hepatol. 2009, 24, 830–840. [Google Scholar] [CrossRef]
- Yeh, Y.-T.; Cho, Y.-Y.; Hsieh, S.-C.; Chiang, A.-N. Chinese olive extract ameliorates hepatic lipid accumulation in vitro and in vivo by regulating lipid metabolism. Sci. Rep. 2018, 8, 1057. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liou, C.-J.; Wei, C.-H.; Chen, Y.-L.; Cheng, C.-Y.; Wang, C.-L.; Huang, W.-C. Fisetin protects against hepatic steatosis through regulation of the Sirt1/AMPK and fatty acid β-oxidation signaling pathway in high-fat diet-induced obese mice. Cell. Physiol. Biochem. 2018, 49, 1870–1884. [Google Scholar] [CrossRef]
- Uen, W.C.; Shi, Y.C.; Choong, C.Y.; Tai, C.J. Cordycepin suppressed lipid accumulation via regulating AMPK activity and mitochondrial fusion in hepatocytes. J. Food Biochem. 2018, 42, e12569. [Google Scholar] [CrossRef]
- Liou, C.-J.; Wu, S.-J.; Shen, S.-C.; Chen, L.-C.; Chen, Y.-L.; Huang, W.-C. Acacetin Protects against Non-Alcoholic Fatty Liver Disease by Regulating Lipid Accumulation and Inflammation in Mice. Int. J. Mol. Sci. 2022, 23, 4687. [Google Scholar] [CrossRef]
- Kang, J.I.; Hong, J.-Y.; Lee, H.-J.; Bae, S.Y.; Jung, C.; Park, H.J.; Lee, S.K. Anti-tumor activity of yuanhuacine by regulating AMPK/mTOR signaling pathway and actin cytoskeleton organization in non-small cell lung cancer cells. PLoS ONE 2015, 10, e0144368. [Google Scholar] [CrossRef] [Green Version]
- Chen, L.; Wang, J.; You, Q.; He, S.; Meng, Q.; Gao, J.; Wu, X.; Shen, Y.; Sun, Y.; Wu, X. Activating AMPK to restore tight junction assembly in intestinal epithelium and to attenuate experimental colitis by metformin. Front. Pharmacol. 2018, 9, 761. [Google Scholar] [CrossRef] [Green Version]
- Yao, H.; Tao, X.; Xu, L.; Qi, Y.; Yin, L.; Han, X.; Xu, Y.; Zheng, L.; Peng, J. Dioscin alleviates non-alcoholic fatty liver disease through adjusting lipid metabolism via SIRT1/AMPK signaling pathway. Pharmacol. Res. 2018, 131, 51–60. [Google Scholar] [CrossRef] [PubMed]
- Wang, L.; Chen, L.; Tan, Y.; Wei, J.; Chang, Y.; Jin, T.; Zhu, H. Betaine supplement alleviates hepatic triglyceride accumulation of apolipoprotein E deficient mice via reducing methylation of peroxisomal proliferator-activated receptor alpha promoter. Lipids Health Dis. 2013, 12, 34. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Xi, Y.; Wu, M.; Li, H.; Dong, S.; Luo, E.; Gu, M.; Shen, X.; Jiang, Y.; Liu, Y.; Liu, H. Baicalin attenuates high fat diet-induced obesity and liver dysfunction: Dose-response and potential role of CaMKKβ/AMPK/ACC pathway. Cell. Physiol. Biochem. 2015, 35, 2349–2359. [Google Scholar] [CrossRef] [PubMed]
- Ding, R.-B.; Bao, J.; Deng, C.-X. Emerging roles of SIRT1 in fatty liver diseases. Int. J. Biol. Sci. 2017, 13, 852. [Google Scholar] [CrossRef]
- Quan, H.Y.; Kim, S.J.; Jo, H.K.; Kim, G.W.; Chung, S.H. Betulinic acid alleviates non-alcoholic fatty liver by inhibiting SREBP1 activity via the AMPK–mTOR–SREBP signaling pathway. Biochem. Pharmacol. 2013, 85, 1330–1340. [Google Scholar] [CrossRef]
- Zhang, H.; Cook, J.; Nickel, J.; Yu, R.; Stecker, K.; Myers, K.; Dean, N.M. Reduction of liver Fas expression by an antisense oligonucleotide protects mice from fulminant hepatitis. Nat. Biotechnol. 2000, 18, 862–867. [Google Scholar] [CrossRef]
- Tardelli, M.; Bruschi, F.V.; Trauner, M. The role of metabolic lipases in the pathogenesis and management of liver disease. Hepatology 2020, 72, 1117–1126. [Google Scholar] [CrossRef] [Green Version]
- Han, C.; Wen, X.; Zheng, Q.; Li, H. Effect of starvation on activities and mRNA expression of lipoprotein lipase and hormone-sensitive lipase in tilapia (Oreochromis Niloticus× O. Areus). Fish Physiol. Biochem. 2011, 37, 113–122. [Google Scholar] [CrossRef]
- Koyama, Y.; Brenner, D.A. Liver inflammation and fibrosis. J. Clin. Investig. 2017, 127, 55–64. [Google Scholar] [CrossRef]
- Kraegen, E.W.; Clark, P.W.; Jenkins, A.B.; Daley, E.A.; Chisholm, D.J.; Storlien, L.H. Development of muscle insulin resistance after liver insulin resistance in high-fat–fed rats. Diabetes 1991, 40, 1397–1403. [Google Scholar] [CrossRef] [PubMed]
- Wang, K.; Cao, G.; Zhang, H.; Li, Q.; Yang, C. Effects of Clostridium butyricum and Enterococcus faecalis on growth performance, immune function, intestinal morphology, volatile fatty acids, and intestinal flora in a piglet model. Food Funct. 2019, 10, 7844–7854. [Google Scholar] [CrossRef] [PubMed]
- Mishra, A.K.; Ghosh, A.R. Probiotic Enterococcus faecalis AG5 mitigated high fat diet induced obesity and produced propionic acid stimulated apoptosis in 3T3-L1 pre-adipocyte. Life Sci. 2020, 261, 118292. [Google Scholar] [CrossRef]
- Takahashi, K.; Nakagawasai, O.; Nemoto, W.; Odaira, T.; Sakuma, W.; Onogi, H.; Nishijima, H.; Furihata, R.; Nemoto, Y.; Iwasa, H. Effect of Enterococcus faecalis 2001 on colitis and depressive-like behavior in dextran sulfate sodium-treated mice: Involvement of the brain-gut axis. J. Neuroinflammation 2019, 16, 201. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Adams, C.A. The probiotic paradox: Live and dead cells are biological response modifiers. Nutr. Res. Rev. 2010, 23, 37–46. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Takahashi, K.; Kurokawa, K.; Hong, L.; Miyagawa, K.; Mochida-Saito, A.; Iwasa, M.; Iwasa, H.; Nakagawasai, O.; Tadano, T.; Takeda, H. Antidepressant effects of Enterococcus faecalis 2001 through the regulation of prefrontal cortical myelination via the enhancement of CREB/BDNF and NF-κB p65/LIF/STAT3 pathways in olfactory bulbectomized mice. J. Psychiatr. Res. 2022, 148, 137–148. [Google Scholar] [CrossRef]
- Choi, E.J.; Iwasa, M.; Han, K.I.; Kim, W.J.; Tang, Y.; Hwang, Y.J.; Kim, E.K. Heat-killed Enterococcus faecalis EF-2001 ameliorates atopic dermatitis in a murine model. Nutrients 2016, 8, 146. [Google Scholar] [CrossRef] [Green Version]
- Jeon, D.B.; Shin, H.G.; Lee, B.W.; Jeong, S.H.; Kim, J.H.; Ha, J.H.; Lee, I.C. Effect of heat-killed Enterococcus faecalis EF-2001 on ethanol-induced acute gastric injury in mice: Protective effect of EF-2001 on acute gastric ulcer. Hum. Exp. Toxicol. 2020, 39, 721–733. [Google Scholar] [CrossRef]
- Choi, M.-S.; Chang, S.-J.; Chae, Y.; Lee, M.-H.; Kim, W.-J.; Iwasa, M.; Han, K.-I.; Kim, W.-J.; Kim, T.-J. Anti-inflammatory effect of Enterococcus faecalis, EF-2001. J. Life Sci. 2018, 28, 1361–1368. [Google Scholar]
- Chang, S.-J.; Lee, M.-H.; Kim, W.-J.; Chae, Y.; Iwasa, M.; Han, K.-I.; Kim, W.-J.; Kim, T.-J. Effect of heat-killed Enterococcus faecalis, EF-2001 on C2C12 myoblast damage induced by oxidative stress and muscle volume decreased by sciatic denervation in C57BL/6 mice. J. Life Sci. 2019, 29, 215–222. [Google Scholar]
- Choi, Y.J.; Fan, M.; Tang, Y.; Iwasa, M.; Han, K.I.; Lee, H.; Hwang, J.Y.; Lee, B.; Kim, E.K. Heat-Killed and Live Enterococcus faecalis Attenuates Enlarged Prostate in an Animal Model of Benign Prostatic Hyperplasia. J. Microbiol. Biotechnol. 2021, 31, 1134–1143. [Google Scholar] [CrossRef]
- Huang, W.; Wang, G.; Xia, Y.; Xiong, Z.; Ai, L. Bile salt hydrolase-overexpressing Lactobacillus strains can improve hepatic lipid accumulation in vitro in an NAFLD cell model. Food Nutr. Res. 2020, 64. [Google Scholar] [CrossRef]
- Kitadokoro, K.; Tanaka, M.; Hikima, T.; Okuno, Y.; Yamamoto, M.; Kamitani, S. Crystal structure of pathogenic Staphylococcus aureus lipase complex with the anti-obesity drug orlistat. Sci. Rep. 2020, 10, 5469. [Google Scholar] [CrossRef] [Green Version]
- Choi, W.J.; Dong, H.J.; Jeong, H.U.; Ryu, D.W.; Song, S.M.; Kim, Y.R.; Jung, H.H.; Kim, T.H.; Kim, Y.H. Lactobacillus plantarum LMT1-48 exerts anti-obesity effect in high-fat diet-induced obese mice by regulating expression of lipogenic genes. Sci. Rep. 2020, 10, 869. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Won, S.M.; Chen, S.; Park, K.W.; Yoon, J.H. Isolation of lactic acid bacteria from kimchi and screening of Lactobacillus sakei ADM14 with anti-adipogenic effect and potential probiotic properties. LWT 2020, 126, 109296. [Google Scholar] [CrossRef]
- Park, J.E.; Oh, S.H.; Cha, Y.S. Lactobacillus Brevis OPK-3 from kimchi prevents obesity and modulates the expression of adipogenic and pro-inflammatory genes in adipose tissue of diet-Induced obese mice. Nutrients 2020, 12, 604. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lee, J.-H.; Woo, K.-J.; Kim, M.-A.; Hong, J.; Kim, J.; Kim, S.-H.; Han, K.-I.; Iwasa, M.; Kim, T.-J. Heat-killed Enterococcus faecalis prevents adipogenesis and high fat diet-induced obesity by inhibition of lipid accumulation through inhibiting C/EBP-α and PPAR-γ in the insulin signaling pathway. Nutrients 2022, 14, 1308. [Google Scholar] [CrossRef]
- Kondoh, M.; Shimada, T.; Fukada, K.; Morita, M.; Katada, K.; Higashimura, Y.; Yoshikawa, T. Beneficial effects of heat-treated Enterococcus faecalis FK-23 on high-fat diet-induced hepatic steatosis in mice. Br. J. Nutr. 2014, 112, 868–875. [Google Scholar] [CrossRef] [Green Version]
- Fan, M.; Choi, Y.J.; Wedamulla, N.E.; Tang, Y.; Han, K.I.; Hwang, J.Y.; Kim, E.K. Heat-Killed Enterococcus faecalis EF-2001 attenuate lipid accumulation in diet-induced obese (DIO) mice by activating AMPK signaling in liver. Foods 2022, 11, 575. [Google Scholar] [CrossRef]
- Makadia, S.S.; Blaha, M.; Keenan, T.; Ndumele, C.; Jones, S.; DeFilippis, A.; Martin, S.; Kohli, P.; Conceicao, R.; Carvalho, J.; et al. Relation of hepatic steatosis to atherogenic dyslipidemia. Am. J. Cardiol. 2013, 112, 1599–1604. [Google Scholar] [CrossRef]
- Wu, S.-J.; Huang, W.-C.; Yu, M.-C.; Chen, Y.-L.; Shen, S.-C.; Yeh, K.-W.; Liou, C.-J. Tomatidine ameliorates obesity-induced nonalcoholic fatty liver disease in mice. J. Nutr. Biochem. 2021, 91, 108602. [Google Scholar] [CrossRef]
- Sun, J.; Ji, H.; Li, X.-X.; Shi, X.-C.; Du, Z.-Y.; Chen, L.-Q. Lipolytic enzymes involving lipolysis in Teleost: Synteny, structure, tissue distribution, and expression in grass carp (Ctenopharyngodon idella). Comp. Biochem. Physiol. Part B Biochem. Mol. Biol. 2016, 198, 110–118. [Google Scholar] [CrossRef]
- Nam, Y.K.; Park, S.J.; Kim, M.H.; Yang, W.M. Pharmacopuncture of Taraxacum platycarpum extract reduces localized fat by regulating the lipolytic pathway. Biomed. Pharmacother. 2021, 141, 111905. [Google Scholar] [CrossRef] [PubMed]
- Hardie, D.G.; Ashford, M.L. AMPK: Regulating energy balance at the cellular and whole body levels. Physiology 2014, 29, 99–107. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, Y.; Xu, S.; Mihaylova, M.M.; Zheng, B.; Hou, X.; Jiang, B.; Park, O.; Luo, Z.; Lefai, E.; Shyy, J.Y.-J. AMPK phosphorylates and inhibits SREBP activity to attenuate hepatic steatosis and atherosclerosis in diet-induced insulin-resistant mice. Cell Metab. 2011, 13, 376–388. [Google Scholar] [CrossRef] [Green Version]
- Seo, M.S.; Kim, J.H.; Kim, H.J.; Chang, K.C.; Park, S.W. Honokiol activates the LKB1–AMPK signaling pathway and attenuates the lipid accumulation in hepatocytes. Toxicol. Appl. Pharmacol. 2015, 284, 113–124. [Google Scholar] [CrossRef] [PubMed]
- Kuo, Y.-T.; Lin, T.-H.; Chen, W.-L.; Lee, H.-M. Alpha-lipoic acid induces adipose triglyceride lipase expression and decreases intracellular lipid accumulation in HepG2 cells. Eur. J. Pharmacol. 2012, 692, 10–18. [Google Scholar] [CrossRef]
- Zordoky, B.N.; Nagendran, J.; Pulinilkunnil, T.; Kienesberger, P.C.; Masson, G.; Waller, T.J.; Kemp, B.E.; Steinberg, G.R.; Dyck, J.R. AMPK-dependent inhibitory phosphorylation of ACC is not essential for maintaining myocardial fatty acid oxidation. Circ. Res. 2014, 115, 518–524. [Google Scholar] [CrossRef] [Green Version]
- Louis, P.; Scott, K.P.; Duncan, S.H.; Flint, H.J. Understanding the effects of diet on bacterial metabolism in the large intestine. J. Appl. Microbiol. 2007, 102, 1197–1208. [Google Scholar] [CrossRef]
- Aron-Wisnewsky, J.; Clement, K. The gut microbiome, diet, and links to cardiometabolic and chronic disorders. Nat. Rev. Nephrol. 2016, 12, 169–181. [Google Scholar] [CrossRef]
- Tang, W.H.W.; Wang, Z.; Levison, B.S.; Koeth, R.A.; Britt, E.B.; Fu, X.; Wu, Y.; Hazen, S.L. Intestinal microbial metabolism of phosphatidylcholine and cardiovascular risk. N. Engl. J. Med. 2013, 368, 1575–1584. [Google Scholar] [CrossRef] [Green Version]
- Kitai, T.; Tang, W.H.W. Gut microbiota in cardiovascular disease and heart failure. Clin. Sci. 2018, 132, 85–91. [Google Scholar] [CrossRef]
- Peng, J.; Xiao, X.; Zhang, X. Interaction between gut microbiome and cardiovascular disease. Life Sci. 2018, 214, 153–157. [Google Scholar] [CrossRef]
- Tang, W.H.W.; Hazen, S.L. The Gut microbiome and its role in cardiovascular diseases. Circulation 2017, 135, 1008–1010. [Google Scholar] [CrossRef]
- Ray, K. Manipulating the gut microbiota to combat alcoholic hepatitis. Nat. Rev. Gastroenterol. Hepatol. 2020, 17, 3. [Google Scholar] [CrossRef]
- Lang, S.; Demir, M.; Duan, Y.; Martin, A.; Schnabl, B. Cytolysin-positive Enterococcus faecalis is not increased in patients with non-alcoholic steatohepatitis. Liver Int. 2020, 40, 860–865. [Google Scholar] [CrossRef]
- Tan, J.; Hu, R.; Li, B.; Li, Y.; Yang, X.; He, Z.; Wu, S. Protocatechuic acid as a phenolic intermediate to ameliorate non-alcoholic fatty liver disease by inhibiting Enterococcus faecalis in mice. Res. Sq. 2020. [Google Scholar] [CrossRef]
- Kim, J.M. Roles of Enteric microbial composition and metabolism in health and diseases. Korean J. Gastroenterol. 2013, 62, 191–205. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Henao-Mejia, J.; Elinav, E.; Jin, C.; Hao, L.; Mehal, W.Z.; Strowig, T.; Thaiss, C.A.; Kau, A.L.; Eisenbarth, S.C.; Jurczak, M.J.; et al. Inflammasome-mediated dysbiosis regulates progression of NAFLD and obesity. Nature 2012, 482, 179–185. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Furusawa, Y.; Obata, Y.; Fukuda, S.; Endo, T.A.; Nakato, G.; Takahashi, D.; Nakanishi, Y.; Uetake, C.; Kato, K.; Kato, T.; et al. Commensal microbe-derived butyrate induces the differentiation of colonic regulatory T cells. Nature 2013, 504, 446–450. [Google Scholar] [CrossRef]
- Frost, G.; Sleeth, M.L.; Sahuri-Arisoylu, M.; Lizarbe, B.; Cerdan, S.; Brody, L.; Anastasovska, J.; Ghourab, S.; Hankir, M.; Zhang, S.; et al. The short-chain fatty acid acetate reduces appetite via a central homeostatic mechanism. Nat. Commun. 2014, 5, 3611. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Den Besten, G.; Bleeker, A.; Gerding, A.; van Eunen, K.; Havinga, R.; van Dijk, T.H.; Oosterveer, M.H.; Jonker, J.W.; Groen, A.K.; Reijngoud, D.J.; et al. Short-chain fatty acids protect against high-fat diet-induced obesity via a PPARgamma-dependent switch from lipogenesis to fat oxidation. Diabetes 2015, 64, 2398. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Osadchiy, V.; Martin, C.R.; Mayer, E.A. The gut brain axis and the microbiome: Mechanisms and clinical implications. Clin. Gastroenterol. Hepatol. 2019, 17, 322–332. [Google Scholar] [CrossRef] [PubMed]









Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Lee, J.-H.; Woo, K.-J.; Hong, J.; Han, K.-I.; Kim, H.S.; Kim, T.-J. Heat-Killed Enterococcus faecalis Inhibit FL83B Hepatic Lipid Accumulation and High Fat Diet-Induced Fatty Liver Damage in Rats by Activating Lipolysis through the Regulation the AMPK Signaling Pathway. Int. J. Mol. Sci. 2023, 24, 4486. https://doi.org/10.3390/ijms24054486
Lee J-H, Woo K-J, Hong J, Han K-I, Kim HS, Kim T-J. Heat-Killed Enterococcus faecalis Inhibit FL83B Hepatic Lipid Accumulation and High Fat Diet-Induced Fatty Liver Damage in Rats by Activating Lipolysis through the Regulation the AMPK Signaling Pathway. International Journal of Molecular Sciences. 2023; 24(5):4486. https://doi.org/10.3390/ijms24054486
Chicago/Turabian StyleLee, Jin-Ho, Keun-Jung Woo, Joonpyo Hong, Kwon-Il Han, Han Sung Kim, and Tack-Joong Kim. 2023. "Heat-Killed Enterococcus faecalis Inhibit FL83B Hepatic Lipid Accumulation and High Fat Diet-Induced Fatty Liver Damage in Rats by Activating Lipolysis through the Regulation the AMPK Signaling Pathway" International Journal of Molecular Sciences 24, no. 5: 4486. https://doi.org/10.3390/ijms24054486