
Gene–Nutrient Interactions in Obesity: COBLL1 Genetic Variants Interact with Dietary Fat Intake to Modulate the Incidence of Obesity


Abstract
:1. Introduction
2. Results
2.1. General Characteristics of Participants at Baseline
2.2. Association between Dietary Fat Intake and BMI
2.3. Association between Dietary Fat Intake and Incidence of Obesity
2.4. Association between COBLL1 rs6717858 Genotypes and Incidence of Obesity
2.5. Association between COBLL1 rs6717858 Genotypes, Dietary Fat Intake, and Incidence of Obesity
3. Discussion
4. Materials and Methods
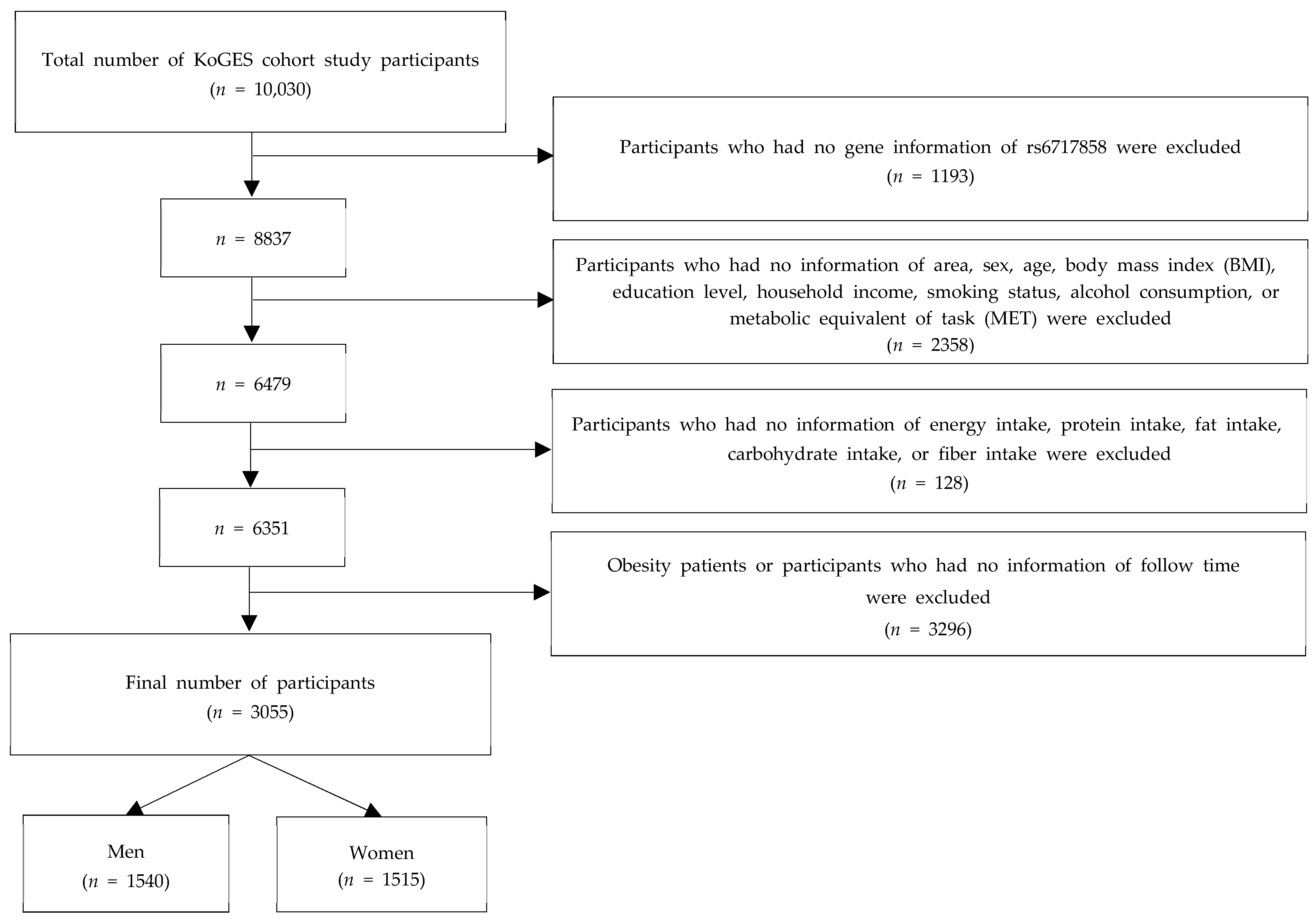
4.1. Data Source and Study Participants
4.2. Dietary Assessment
4.3. Definition of Obesity
4.4. Genotyping and Imputation
4.5. Statistical Analyses
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Blüher, M. Obesity: Global epidemiology and pathogenesis. Nat. Rev. Endocrinol. 2019, 15, 288–298. [Google Scholar] [CrossRef] [PubMed]
- Lee, G.B.; Kim, Y.; Park, S.; Kim, H.C.; Oh, K. Obesity, hypertension, diabetes mellitus, and hypercholesterolemia in Korean adults before and during the COVID-19 pandemic: A special report of the 2020 Korea National Health and Nutrition Examination Survey. Epidemiol. Health 2022, 44, e2022041. [Google Scholar] [CrossRef] [PubMed]
- Magalhaes, C.; Carvalho, M.; Sousa, L.; Caramelli, P.; Gomes, K. Leptin in Alzheimer’s disease. Anal. Chim. Acta 2015, 450, 162–168. [Google Scholar]
- Hoong, C.W.S.; Hussain, I.; Aravamudan, V.M.; Phyu, E.E.; Lin, J.H.X.; Koh, H. Obesity is associated with poor COVID-19 outcomes: A systematic review and meta-analysis. Horm. Metab. Res. 2021, 53, 85–93. [Google Scholar] [CrossRef] [PubMed]
- Pietri, L.; Giorgi, R.; Bégu, A.; Lojou, M.; Koubi, M.; Cauchois, R.; Grangeot, R.; Dubois, N.; Kaplanski, G.; Valéro, R. Excess body weight is an independent risk factor for severe forms of COVID-19. Metabolism 2021, 117, 154703. [Google Scholar] [CrossRef] [PubMed]
- Herbert, A.; Gerry, N.P.; McQueen, M.B.; Heid, I.M.; Pfeufer, A.; Illig, T.; Wichmann, H.E.; Meitinger, T.; Hunter, D.; Hu, F.B. A common genetic variant is associated with adult and childhood obesity. Science 2006, 312, 279–283. [Google Scholar] [CrossRef]
- Farooqi, I.S.; Wangensteen, T.; Collins, S.; Kimber, W.; Matarese, G.; Keogh, J.M.; Lank, E.; Bottomley, B.; Lopez Fernandez, J.; Ferraz Amaro, I. Clinical and molecular genetic spectrum of congenital deficiency of the leptin receptor. N. Engl. J. Med. 2007, 356, 237–247. [Google Scholar] [CrossRef]
- Licinio, J.; Caglayan, S.; Ozata, M.; Yildiz, B.O.; De Miranda, P.B.; O’Kirwan, F.; Whitby, R.; Liang, L.; Cohen, P.; Bhasin, S. Phenotypic effects of leptin replacement on morbid obesity, diabetes mellitus, hypogonadism, and behavior in leptin-deficient adults. Proc. Natl. Acad. Sci. USA 2004, 101, 4531–4536. [Google Scholar] [CrossRef]
- Farooqi, I.S.; Matarese, G.; Lord, G.M.; Keogh, J.M.; Lawrence, E.; Agwu, C.; Sanna, V.; Jebb, S.A.; Perna, F.; Fontana, S. Beneficial effects of leptin on obesity, T cell hyporesponsiveness, and neuroendocrine/metabolic dysfunction of human congenital leptin deficiency. J. Clin. Investig. 2002, 110, 1093–1103. [Google Scholar] [CrossRef]
- Bray, G.A.; Popkin, B.M. Dietary fat intake does affect obesity! Am. J. Clin. Nutr. 1998, 68, 1157–1173. [Google Scholar] [CrossRef]
- Wang, L.; Wang, H.; Zhang, B.; Popkin, B.M.; Du, S. Elevated fat intake increases body weight and the risk of overweight and obesity among Chinese adults: 1991–2015 trends. Nutrients 2020, 12, 3272. [Google Scholar] [CrossRef] [PubMed]
- Hall, K.D.; Bemis, T.; Brychta, R.; Chen, K.Y.; Courville, A.; Crayner, E.J.; Goodwin, S.; Guo, J.; Howard, L.; Knuth, N.D. Calorie for calorie, dietary fat restriction results in more body fat loss than carbohydrate restriction in people with obesity. Cell Metab. 2015, 22, 427–436. [Google Scholar] [CrossRef] [PubMed]
- Cani, P.D.; Bibiloni, R.; Knauf, C.; Waget, A.; Neyrinck, A.M.; Delzenne, N.M.; Burcelin, R. Changes in gut microbiota control metabolic endotoxemia-induced inflammation in high-fat diet–induced obesity and diabetes in mice. Diabetes 2008, 57, 1470–1481. [Google Scholar] [CrossRef] [PubMed]
- Machate, D.J.; Figueiredo, P.S.; Marcelino, G.; Guimarães, R.d.C.A.; Hiane, P.A.; Bogo, D.; Pinheiro, V.A.Z.; Oliveira, L.C.S.d.; Pott, A. Fatty acid diets: Regulation of gut microbiota composition and obesity and its related metabolic dysbiosis. Int. J. Mol. Sci. 2020, 21, 4093. [Google Scholar] [CrossRef]
- Sun, Y.; Ge, X.; Li, X.; He, J.; Wei, X.; Du, J.; Sun, J.; Li, X.; Xun, Z.; Liu, W. High-fat diet promotes renal injury by inducing oxidative stress and mitochondrial dysfunction. Cell Death Dis 2020, 11, 914. [Google Scholar] [CrossRef]
- Maffei, á.; Halaas, J.; Ravussin, E.; Pratley, R.; Lee, G.; Zhang, Y.; Fei, H.; Kim, S.; Lallone, R.; Ranganathan, S. Leptin levels in human and rodent: Measurement of plasma leptin and ob RNA in obese and weight-reduced subjects. Nat. Med. 1995, 1, 1155–1161. [Google Scholar] [CrossRef]
- Hu, S.; Wang, L.; Yang, D.; Li, L.; Togo, J.; Wu, Y.; Liu, Q.; Li, B.; Li, M.; Wang, G. Dietary fat, but not protein or carbohydrate, regulates energy intake and causes adiposity in mice. Cell Metab. 2018, 28, 415–431.e4. [Google Scholar] [CrossRef]
- Wang, H.; Storlien, L.H.; Huang, X.F. Effects of dietary fat types on body fatness, leptin, and ARC leptin receptor, NPY, and AgRP mRNA expression. Am. J. Physiol. Endocrinol. Metab. 2002, 282, E1352–E1359. [Google Scholar] [CrossRef]
- Huang, L.; Li, C. Leptin: A multifunctional hormone. Cell Res. 2000, 10, 81–92. [Google Scholar] [CrossRef]
- Zhang, Y.; Scarpace, P.J. The role of leptin in leptin resistance and obesity. Physiol. Behav. 2006, 88, 249–256. [Google Scholar] [CrossRef]
- Simon, M.S.; Heilbrun, L.K.; Boomer, A.; Kresge, C.; Depper, J.; Kim, P.N.; Valeriote, F.; Martino, S. A randomized trial of a low-fat dietary intervention in women at high risk for breast cancer. Nutr. Cancer. 1997, 27, 136–142. [Google Scholar] [CrossRef]
- Jeffery, R.W.; Hellerstedt, W.L.; French, S.A.; Baxter, J.E. A randomized trial of counseling for fat restriction versus calorie restriction in the treatment of obesity. Int. J. Obes. Relat. Metab. Disord. 1995, 19, 132–137. [Google Scholar]
- Carroll, E.A.; Gerrelli, D.; Gasca, S.; Berg, E.; Beier, D.R.; Copp, A.J.; Klingensmith, J. Cordon-bleu is a conserved gene involved in neural tube formation. Dev. Biol. 2003, 262, 16–31. [Google Scholar] [CrossRef]
- Shungin, D.; Winkler, T.W.; Croteau Chonka, D.C.; Ferreira, T.; Locke, A.E.; Mägi, R.; Strawbridge, R.J.; Pers, T.H.; Fischer, K.; Justice, A.E. New genetic loci link adipose and insulin biology to body fat distribution. Nature 2015, 518, 187–196. [Google Scholar] [CrossRef] [PubMed]
- Scott, R.A.; Lagou, V.; Welch, R.P.; Wheeler, E.; Montasser, M.E.; Luan, J.; Mägi, R.; Strawbridge, R.J.; Rehnberg, E.; Gustafsson, S. Large-scale association analyses identify new loci influencing glycemic traits and provide insight into the underlying biological pathways. Nat. Genet. 2012, 44, 991–1005. [Google Scholar] [CrossRef] [PubMed]
- Morris, A.; Voight, B.; Teslovich, T.; Wellcome Trust Case Control Consortium; Asian Genetic Epidemiology Network–Type 2 Diabetes (AGEN-T2D) Consortium; South Asian Type 2 Diabetes (SAT2D) Consortium; Diabetes Genetics Replication and Meta-analysis (DIAGRAM) Consortium. Large-scale association analysis provides insights into the genetic architecture and pathophysiology of type 2 diabetes. Nat. Genet. 2012, 44, 981–990. [Google Scholar] [PubMed]
- Teslovich, T.M.; Musunuru, K.; Smith, A.V.; Edmondson, A.C.; Stylianou, I.M.; Koseki, M.; Pirruccello, J.P.; Ripatti, S.; Chasman, D.I.; Willer, C.J. Biological, clinical and population relevance of 95 loci for blood lipids. Nature 2010, 466, 707–713. [Google Scholar] [CrossRef]
- Lu, Y.; Day, F.R.; Gustafsson, S.; Buchkovich, M.L.; Na, J.; Bataille, V.; Cousminer, D.L.; Dastani, Z.; Drong, A.W.; Esko, T. New loci for body fat percentage reveal link between adiposity and cardiometabolic disease risk. Nat. Commun. 2016, 7, 10495. [Google Scholar] [CrossRef]
- Wang, Z.; Yan, Z.; Zhang, B.; Rao, Z.; Zhang, Y.; Liu, J.; Yu, L.; Zhao, Y.; Yang, B.; Wu, T.; et al. Identification of a 5-gene signature for clinical and prognostic prediction in gastric cancer patients upon microarray data. Med. Oncol. 2013, 30, 1–11. [Google Scholar] [CrossRef]
- Takayama, K.i.; Suzuki, T.; Fujimura, T.; Takahashi, S.; Inoue, S. COBLL1 modulates cell morphology and facilitates androgen receptor genomic binding in advanced prostate cancer. Proc. Natl. Acad. Sci. USA 2018, 115, 4975–4980. [Google Scholar] [CrossRef]
- Hussein, S.; Abdelazem, A.S.; Abdelmoneem, S.; Abdelnabi, A.S.M.; Khamis, T.; Obaya, A.A.; Ebian, H.F. Evaluation of miRNA 223/125a and COBLL1 Expressions and ROR-1 Levels as Reliable Markers in B-chronic Lymphocytic Leukemia. Asian Pac. J. Cancer Prev. 2022, 23, 2735–2742. [Google Scholar] [CrossRef] [PubMed]
- Han, S.; Kim, S.; Kim, H.; Lee, Y.; Choi, S.; Park, G.; Kim, D.; Lee, A.; Kim, J.; Choi, J. Cobll1 is linked to drug resistance and blastic transformation in chronic myeloid leukemia. Leukemia 2017, 31, 1532–1539. [Google Scholar] [CrossRef] [PubMed]
- Goodarzi, M.O. Genetics of obesity: What genetic association studies have taught us about the biology of obesity and its complications. Lancet Diabetes Endocrinol. 2018, 6, 223–236. [Google Scholar] [CrossRef] [PubMed]
- Gilks, W.P.; Abbott, J.K.; Morrow, E.H. Sex differences in disease genetics: Evidence, evolution, and detection. Trends Genet. 2014, 30, 453–463. [Google Scholar] [CrossRef] [PubMed]
- Heid, I.M.; Jackson, A.U.; Randall, J.C.; Winkler, T.W.; Qi, L.; Steinthorsdottir, V.; Thorleifsson, G.; Zillikens, M.C.; Speliotes, E.K.; Mägi, R. Meta-analysis identifies 13 new loci associated with waist-hip ratio and reveals sexual dimorphism in the genetic basis of fat distribution. Nat. Genet. 2010, 42, 949–960. [Google Scholar] [CrossRef]
- Chen, Z.; Yu, H.; Shi, X.; Warren, C.R.; Lotta, L.A.; Friesen, M.; Meissner, T.B.; Langenberg, C.; Wabitsch, M.; Wareham, N. Functional screening of candidate causal genes for insulin resistance in human preadipocytes and adipocytes. Circ. Res. 2020, 126, 330–346. [Google Scholar] [CrossRef]
- Sun, C.; Förster, F.; Gutsmann, B.; Moulla, Y.; Stroh, C.; Dietrich, A.; Schön, M.R.; Gärtner, D.; Lohmann, T.; Dressler, M. Metabolic Effects of the Waist-To-Hip Ratio Associated Locus GRB14/COBLL1 Are Related to GRB14 Expression in Adipose Tissue. Int. J. Mol. Sci. 2022, 23, 8558. [Google Scholar] [CrossRef]
- Dallner, O.S.; Marinis, J.M.; Lu, Y.H.; Birsoy, K.; Werner, E.; Fayzikhodjaeva, G.; Dill, B.D.; Molina, H.; Moscati, A.; Kutalik, Z. Dysregulation of a long noncoding RNA reduces leptin leading to a leptin-responsive form of obesity. Nat. Med. 2019, 25, 507–516. [Google Scholar] [CrossRef]
- Kilpeläinen, T.O.; Carli, J.F.M.; Skowronski, A.A.; Sun, Q.; Kriebel, J.; Feitosa, M.F.; Hedman, Å.K.; Drong, A.W.; Hayes, J.E.; Zhao, J. Genome-wide meta-analysis uncovers novel loci influencing circulating leptin levels. Nat. Commun. 2016, 7, 10494. [Google Scholar] [CrossRef]
- Randall, J.C.; Winkler, T.W.; Kutalik, Z.; Berndt, S.I.; Jackson, A.U.; Monda, K.L.; Kilpeläinen, T.O.; Esko, T.; Mägi, R.; Li, S. Sex-stratified genome-wide association studies including 270,000 individuals show sexual dimorphism in genetic loci for anthropometric traits. PLoS Genet. 2013, 9, e1003500. [Google Scholar] [CrossRef]
- Knopp, R.H.; Paramsothy, P.; Retzlaff, B.M.; Fish, B.; Walden, C.; Dowdy, A.; Tsunehara, C.; Aikawa, K.; Cheung, M.C. Gender differences in lipoprotein metabolism and dietary response: Basis in hormonal differences and implications for cardiovascular disease. Curr. Atheroscler. Rep. 2005, 7, 472–479. [Google Scholar] [CrossRef] [PubMed]
- Haleem, D.J.; Mahmood, K. Brain serotonin in high-fat diet-induced weight gain, anxiety and spatial memory in rats. Nutr. Neurosci. 2021, 24, 226–235. [Google Scholar] [CrossRef] [PubMed]
- Cavaliere, G.; Viggiano, E.; Trinchese, G.; De Filippo, C.; Messina, A.; Monda, V.; Valenzano, A.; Cincione, R.I.; Zammit, C.; Cimmino, F. Long feeding high-fat diet induces hypothalamic oxidative stress and inflammation, and prolonged hypothalamic AMPK activation in rat animal model. Front. Physiol. 2018, 9, 818. [Google Scholar] [CrossRef]
- McNay, D.E.; Speakman, J.R. High fat diet causes rebound weight gain. Mol. Metab. 2013, 2, 103–108. [Google Scholar] [CrossRef]
- Chakraborty, T.R.; Donthireddy, L.; Adhikary, D.; Chakraborty, S. Long-term high fat diet has a profound effect on body weight, hormone levels, and estrous cycle in mice. Med. Sci. Monit. 2016, 22, 1601. [Google Scholar] [CrossRef]
- Paeratakul, S.; Popkin, B.M.; Keyou, G.; Adair, L.; Stevens, J. Changes in diet and physical activity affect the body mass index of Chinese adults. Int. J. Obes. 1998, 22, 424–431. [Google Scholar] [CrossRef] [PubMed]
- Field, A.E.; Willett, W.C.; Lissner, L.; Colditz, G.A. Dietary fat and weight gain among women in the Nurses’ Health Study. Obesity 2007, 15, 967–976. [Google Scholar] [CrossRef]
- Yang, Y.; Smith Jr, D.L.; Keating, K.D.; Allison, D.B.; Nagy, T.R. Variations in body weight, food intake and body composition after long-term high-fat diet feeding in C57BL/6J mice. Obesity 2014, 22, 2147–2155. [Google Scholar] [CrossRef]
- Ministry of Health and Welfare. The Korean Nutrition Society, Dietary Reference Intakes for Koreans. 2020. Available online: http://www.kns.or.kr/FileRoom/FileRoom_view.asp?idx=108&BoardID=Kdr (accessed on 1 November 2022).
- Pereira, M.A.; Jacobs Jr, D.R.; Van Horn, L.; Slattery, M.L.; Kartashov, A.I.; Ludwig, D.S. Dairy consumption, obesity, and the insulin resistance syndrome in young adults: The CARDIA Study. JAMA 2002, 287, 2081–2089. [Google Scholar] [CrossRef]
- Beulen, Y.; Martínez González, M.A.; Van de Rest, O.; Salas Salvadó, J.; Sorlí, J.V.; Gómez Gracia, E.; Fiol, M.; Estruch, R.; Santos Lozano, J.M.; Schröder, H. Quality of dietary fat intake and body weight and obesity in a mediterranean population: Secondary analyses within the PREDIMED trial. Nutrients 2018, 10, 2011. [Google Scholar] [CrossRef] [PubMed]
- Heshka, J.T.; Jones, P.J. A role for dietary fat in leptin receptor, OB-Rb, function. Life Sci. 2001, 69, 987–1003. [Google Scholar] [CrossRef] [PubMed]
- Dragano, N.R.; Haddad Tovolli, R.; Velloso, L.A. Leptin, neuroinflammation and obesity. Endocr. Immunol. 2017, 48, 84–96. [Google Scholar]
- Myers, M.G., Jr.; Leibel, R.L.; Seeley, R.J.; Schwartz, M.W. Obesity and leptin resistance: Distinguishing cause from effect. Trends Endocrinol. Metab. 2010, 21, 643–651. [Google Scholar] [CrossRef] [PubMed]
- Tchernof, A.; Després, J.P. Sex steroid hormones, sex hormone-binding globulin, and obesity in men and women. Horm. Metab. Res. 2000, 32, 526–536. [Google Scholar] [CrossRef] [PubMed]
- Palmer, B.F.; Clegg, D.J. The sexual dimorphism of obesity. Mol. Cell. Endocrinol. 2015, 402, 113–119. [Google Scholar] [CrossRef]
- Field, A.E.; Colditz, G.A.; Willett, W.C.; Longcope, C.; McKinlay, J.B. The relation of smoking, age, relative weight, and dietary intake to serum adrenal steroids, sex hormones, and sex hormone-binding globulin in middle-aged men. J. Clin. Endocrinol. Metab. 1994, 79, 1310–1316. [Google Scholar]
- Nuttall, F.Q. Body mass index: Obesity, BMI, and health: A critical review. Nutr. Today 2015, 50, 117. [Google Scholar] [CrossRef]
- Hinney, A.; Nguyen, T.T.; Scherag, A.; Friedel, S.; Brönner, G.; Müller, T.D.; Grallert, H.; Illig, T.; Wichmann, H.E.; Rief, W. Genome wide association (GWA) study for early onset extreme obesity supports the role of fat mass and obesity associated gene (FTO) variants. PLoS ONE 2007, 2, e1361. [Google Scholar] [CrossRef]
- Choi, S.; Bae, S.; Park, T. Risk prediction using genome-wide association studies on type 2 diabetes. Genom. Inform. 2016, 14, 138–148. [Google Scholar] [CrossRef]
- Lee, J.Y.; Lee, B.S.; Shin, D.J.; Park, K.W.; Shin, Y.A.; Kim, K.J.; Heo, L.; Lee, J.Y.; Kim, Y.K.; Kim, Y.J. A genome-wide association study of a coronary artery disease risk variant. J. Hum. Genet. 2013, 58, 120–126. [Google Scholar] [CrossRef]
- Shin, D.; Lee, K.W. Dietary carbohydrates interact with AMY1 polymorphisms to influence the incidence of type 2 diabetes in Korean adults. Sci. Rep. 2021, 11, 16788. [Google Scholar] [CrossRef] [PubMed]


{kind=link}
| Variables | Men (n = 1540) | Women (n = 1515) | ||||
|---|---|---|---|---|---|---|
| Non-Obese | Obese | p-Value (1) | Non-Obese | Obese | p-Value (1) | |
| Participants, n | 1228 | 312 | 1200 | 315 | ||
| rs6717858 genotypes | 0.34 | 0.36 | ||||
| TT (n = 2473) | 979 (79.7%) | 241 (77.2%) | 998 (83.2%) | 255 (81.0%) | ||
| CT, CC (n = 582) | 249 (20.3%) | 71 (22.8%) | 202 (16.8%) | 60 (19.1%) | ||
| Age (years) | 51.8 ± 8.9 | 48.9 ± 7.6 | <0.0001 | 50.7 ± 8.9 | 50.9 ± 8.6 | 0.66 |
| Area | 0.002 | 0.57 | ||||
| Ansung | 419 (34.1%) | 72 (23.1%) | 444 (37.0%) | 122 (38.7%) | ||
| Ansan | 809 (65.9%) | 240 (76.9%) | 756 (63.0%) | 193 (61.3%) | ||
| Education level | 0.84 | 0.88 | ||||
| College or lower | 1040 (84.7%) | 260 (83.3%) | 1126 (93.8%) | 295 (93.7%) | ||
| University | 156 (12.7%) | 43 (13.8%) | 69 (5.8%) | 18 (5.7%) | ||
| Graduate school or higher | 32 (2.6%) | 9 (2.9%) | 5 (0.4%) | 2 (0.6%) | ||
| Household income (1 million won/month) | 0.0035 | 0.2 | ||||
| <1 | 336 (27.4%) | 64 (20.5%) | 388 (32.3%) | 117 (37.2%) | ||
| 1–3 | 624 (50.8%) | 155 (49.7%) | 594 (49.5%) | 139 (44.1%) | ||
| >3 | 268 (21.8%) | 93 (29.8%) | 218 (18.2%) | 59 (18.7%) | ||
| BMI (kg/m²) | 22.1 ± 1.8 | 24.0 ± 0.8 | <0.0001 | 22.2 ± 1.7 | 23.9 ± 1.0 | <0.0001 |
| Alcohol consumption | 0.0035 | 0.4 | ||||
| None | 336 (27.4%) | 64 (20.5%) | 850 (70.8%) | 219 (69.5%) | ||
| Past | 624 (50.8%) | 155 (49.7%) | 32 (2.7%) | 13 (4.1%) | ||
| Current | 268 (21.8%) | 93 (29.8%) | 318 (26.5%) | 83 (26.4%) | ||
| Smoking status | 0.16 | 0.96 | ||||
| None | 240 (19.5%) | 61 (19.6%) | 1149 (95.8%) | 301 (95.6%) | ||
| Past | 121 (9.9%) | 20 (6.4%) | 13 (1.1%) | 4 (1.3%) | ||
| Current | 867 (70.6%) | 231 (74.0%) | 38 (3.1%) | 10 (3.1%) | ||
| MET (hours/day) (2) | 170.6 ± 106.6 | 156.0 ± 97.1 | 0.02 | 151.5 ± 92.6 | 157.4 ± 96.2 | 0.32 |
| Biomarkers | ||||||
| Waist circumference (cm) | 78.5 ± 5.8 | 82.6 ± 4.2 | <0.0001 | 74.8 ± 7.1 | 78.8 ± 7.2 | <0.0001 |
| HDL-cholesterol (mg/dL) (3) | 45.7 ± 10.3 | 43.8 ± 10.2 | 0.003 | 47.7 ± 10.3 | 46.8 ± 10.6 | 0.17 |
| Triglyceride (mg/dL) | 155.1 ± 109.8 | 173.3 ± 99.8 | 0.0048 | 129.2 ± 71.73 | 140.1 ± 101.3 | 0.07 |
| Fasting glucose (mg/dL) | 88.9 ± 21.8 | 89.0 ± 29.9 | 0.96 | 82.4 ± 14.7 | 83.5 ± 21.1 | 0.39 |
| Blood pressure | ||||||
| Average systolic blood pressure (mmHg) | 119.4 ± 16.6 | 118.8 ± 16.5 | 0.62 | 115.0 ± 18.4 | 116.2 ± 17.8 | 0.27 |
| Average diastolic blood pressure (mmHg) | 79.7 ± 10.69 | 80.24 ± 10.52 | 0.42 | 75.0 ± 11.3 | 76.4 ± 10.8 | 0.04 |
| Dietary intake | ||||||
| Total energy intake (kcal/day) | 1979 ± 609 | 2070 ± 619 | 0.02 | 1886 ± 696 | 1876 ± 682 | 0.83 |
| Protein intake (% energy) | 13.6 ± 2.3 | 13.9 ± 2.1 | 0.0069 | 13.5 ± 2.4 | 13.5 ± 2.2 | 0.72 |
| Fat intake (% energy) | 15.4 ± 5.1 | 16.4 ± 4.8 | 0.001 | 14.0 ± 5.4 | 14.1 ± 5.1 | 0.78 |
| Carbohydrate intake (% energy) | 69.8 ± 6.6 | 68.5 ± 6.1 | 0.001 | 71.5 ± 7.1 | 71.4 ± 6.6 | 0.86 |
| Fiber intake (g/day) | 6.7 ± 3.0 | 6.9 ± 3.6 | 0.46 | 6.9 ± 3.3 | 7.2 ± 4.3 | 0.23 |
| Variables | Men (n = 1540) | Women (n = 1515) | ||||
|---|---|---|---|---|---|---|
| COBLL1 rs6717858 Genotypes | ||||||
| TT | CT, CC | p-Value (1) | TT | CT, CC | p-Value (1) | |
| Participants, n | 1220 | 320 | 1253 | 262 | ||
| Age (years) | 51.3 ± 8.7 | 50.8 ± 8.7 | 0.33 | 50.4 ± 8.8 | 52.0 ± 9.1 | 0.0082 |
| Area | 0.34 | 0.39 | ||||
| Ansung | 396 (32.5%) | 95 (29.7%) | 462 (36.9%) | 104 (39.7%) | ||
| Ansan | 824 (67.5%) | 225 (70.3%) | 791 (63.1%) | 158 (60.3%) | ||
| Education level | 0.2 | 0.32 | ||||
| College or lower | 1033 (84.7%) | 267 (83.4%) | 1171 (93.5%) | 250 (95.4%) | ||
| University | 151 (12.3%) | 48 (15.0%) | 75 (6.0%) | 12 (4.6%) | ||
| Graduate school or higher | 36 (3.0%) | 5 (1.6%) | 7 (0.5%) | 0 (0%) | ||
| Household income (1 million won/month) | 0.91 | 0.68 | ||||
| <1 | 320 (26.2%) | 80 (25.0%) | 417 (33.3%) | 88 (33.6%) | ||
| 1–3 | 615 (50.4%) | 164 (51.2%) | 602 (48.0%) | 131 (50.0%) | ||
| >3 | 285 (23.4%) | 76 (23.8%) | 234 (18.7%) | 43 (16.4%) | ||
| Body mass index (kg/m²) | 22.5 ± 1.8 | 22.6 ± 1.8 | 0.42 | 22.5 ± 1.7 | 22.4 ± 1.8 | 0.35 |
| Alcohol consumption | 0.4 | 0.16 | ||||
| None | 247 (20.2%) | 54 (16.9%) | 889 (71.0%) | 180 (68.7%) | ||
| Past | 111 (9.1%) | 30 (9.3%) | 41 (3.2%) | 4 (1.5%) | ||
| Current | 862 (70.7%) | 236 (73.8%) | 323 (25.8%) | 78 (29.8%) | ||
| Smoking status | 0.54 | 0.17 | ||||
| None | 242 (19.8%) | 72 (22.5%) | 1201 (95.9%) | 249 (95.0%) | ||
| Past | 370 (30.4%) | 91 (28.4%) | 16 (1.2%) | 1 (0.4%) | ||
| Current | 608 (49.8%) | 157 (49.1%) | 36 (2.9%) | 12 (4.6%) | ||
| MET (hours/day) (2) | 168.2 ± 105.4 | 167.1 ± 102.5 | 0.88 | 151.6 ± 93.6 | 157.9 ± 92.3 | 0.33 |
| Waist circumference (cm) | 79.3 ± 5.7 | 79.2 ± 5.8 | 0.77 | 75.6 ± 7.2 | 75.7 ± 7.6 | 0.78 |
| HDL-cholesterol (mg/dL) (3) | 45.0 ± 10.2 | 46.5 ± 10.6 | 0.03 | 47.6 ± 10.3 | 47.0 ± 10.4 | 0.41 |
| Triglyceride (mg/dL) | 162.0 ± 114.9 | 145.8 ± 76.2 | 0.0027 | 129.7 ± 77.8 | 140.1 ± 83.7 | 0.05 |
| Fasting glucose (mg/dL) | 89.0 ± 21.9 | 89.0 ± 29.5 | 0.96 | 82.7 ± 16.8 | 82.0 ± 13.3 | 0.46 |
| Blood pressure | ||||||
| Average systolic blood pressure (mmHg) | 119.1 ± 16.3 | 119.8 ± 17.8 | 0.56 | 114.7 ± 18.0 | 117.8 ± 19.4 | 0.01 |
| Average diastolic blood pressure (mmHg) | 79.8 ± 10.6 | 79.7 ± 11.0 | 0.83 | 74.9 ± 11.0 | 76.9 ± 12.1 | 0.01 |
| Dietary intake | ||||||
| Total energy intake (kcal/day) | 1998 ± 595 | 1993 ± 676 | 0.89 | 1890 ± 674 | 1854 ± 777 | 0.48 |
| Protein intake (% energy) | 13.5 ± 2.4 | 13.5 ± 2.2 | 0.72 | 13.5 ± 2.3 | 13.4 ± 2.5 | 0.42 |
| Fat intake (% energy) | 15.6 ± 5.1 | 15.7 ± 4.8 | 0.54 | 14.2 ± 5.3 | 13.5 ± 5.5 | 0.04 |
| Carbohydrate intake (% energy) | 69.6 ± 6.6 | 69.3 ± 6.2 | 0.37 | 71.3 ± 6.9 | 72.1 ± 7.2 | 0.08 |
| Fiber intake (g/day) | 6.7 ± 3.0 | 6.9 ± 3.7 | 0.41 | 7.0 ± 3.5 | 7.0 ± 3.9 | 0.98 |
| Men (n = 1540) | Women (n = 1515) | ||||
|---|---|---|---|---|---|
| BMI (kg/m2) | |||||
| Beta ± SE | p-Value | Beta ± SE | p-Value | ||
| Dietary fat (g/1000 kcal) | Model 1 (1) | −0.001 ± 0.009 | 0.89 | −0.004 ± 0.008 | 0.58 |
| Model 2 (2) | −0.015 ± 0.009 | 0.12 | −0.003 ± 0.008 | 0.71 | |
| Dietary Fat (% Energy) | ||||||
|---|---|---|---|---|---|---|
| Men (n = 1540) | Tertile 1 | Tertile 2 | Tertile 3 | |||
| Median (ranges) | 10.7 (2.9–13.2) | 15.4 (13.2–17.5) | 20.3 (17.5–35.1) | |||
| Person-years | 4717.5 | 4653.1 | 4634.0 | |||
| Incident cases (n) | 84/513 | 109/514 | 119/513 | |||
| HR (95% CI) | p-value | HR (95% CI) | p-value | HR (95% CI) | p-value | |
| Model 1 (1) | 1.00 (Ref.) | 1.13 (0.81–1.57) | 0.46 | 1.41 (1.02–1.95) | 0.04 | |
| Model 2 (2) | 1.00 (Ref.) | 1.13 (0.81–1.57) | 0.48 | 1.37 (0.98–1.93) | 0.07 | |
| Women (n = 1515) | Tertile 1 | Tertile 2 | Tertile 3 | |||
| Median (ranges) | 8.9 (1.9–11.5) | 13.7 (11.5–15.9) | 18.9 (15.9–42.0) | |||
| Person-years | 4582.6 | 4622.1 | 4753.4 | |||
| Incident cases (n) | 97/505 | 108/505 | 110/505 | |||
| HR (95% CI) | p-value | HR (95% CI) | p-value | HR (95% CI) | p-value | |
| Model 1 (1) | 1.00 (Ref.) | 0.96 (0.72–1.28) | 0.79 | 1.31 (0.97–1.78) | 0.08 | |
| Model 2 (2) | 1.00 (Ref.) | 0.97 (0.72–1.29) | 0.81 | 1.35 (0.99–1.84) | 0.06 | |
| COBLL1 rs6717858 Genotypes | |||
|---|---|---|---|
| Men (n = 1540) | TT | CT, CC | |
| HR (95% CI) | HR (95% CI) | p-value | |
| Model 1 (1) | 1.00 (Ref.) | 1.08 (0.83–1.41) | 0.58 |
| Model 2 (2) | 1.00 (Ref.) | 1.08 (0.83–1.41) | 0.58 |
| Women (n = 1515) | TT | CT, CC | |
| HR (95% CI) | HR (95% CI) | p-value | |
| Model 1 (1) | 1.00 (Ref.) | 1.29 (0.97–1.71) | 0.09 |
| Model 2 (2) | 1.00 (Ref.) | 1.27 (0.95–1.69) | 0.10 |
| Men (n = 1540) | Women (n = 1515) | ||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Dietary Fat (% energy) | |||||||||||||||
| Tertile 1 | Tertile 2 | Tertile 3 | p-Interaction | Tertile 1 | Tertile 2 | Tertile 3 | p-Interaction | ||||||||
| Median | 10.7 | 15.4 | 20.3 | 8.9 | 13.7 | 18.9 | |||||||||
| HR (95% CI) | p-value | HR (95% CI) | p-value | HR (95% CI) | p-value | HR (95% CI) | p-value | HR (95% CI) | p-value | HR (95% CI) | p-value | ||||
| Model 1 (1) | TT | 1.00 (Ref.) | 1.19 (0.82–1.71) | 0.36 | 1.37 (0.95–1.96) | 0.09 | 0.52 | 1.00 (Ref.) | 1.43 (0.95–2.17) | 0.62 | 1.49 (1.08–2.06) | 0.02 | 0.09 | ||
| CT, CC | 1.08 (0.54–2.15) | 0.82 | 1.03 (0.62–1.70) | 0.91 | 1.66 (1.07–2.58) | 0.03 | 1.50 (0.95–2.37) | 0.09 | 1.68 (1.04–2.72) | 0.03 | 0.90 (0.43–1.88) | 0.78 | |||
| Model 2 (2) | TT | 1.00 (Ref.) | 1.18 (0.82–1.71) | 0.37 | 1.32 (0.91–1.92) | 0.15 | 0.49 | 1.00 (Ref.) | 0.93 (0.67–1.29) | 0.67 | 1.53 (1.10–2.13) | 0.01 | 0.08 | ||
| CT, CC | 1.09 (0.55–2.17) | 0.80 | 1.01 (0.61–1.68) | 0.96 | 1.63 (1.04–2.56) | 0.03 | 1.44 (0.95–2.18) | 0.09 | 1.63 (1.01–2.64) | 0.0458 | 0.92 (0.44–1.94) | 0.84 | |||
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kwak, J.; Shin, D. Gene–Nutrient Interactions in Obesity: COBLL1 Genetic Variants Interact with Dietary Fat Intake to Modulate the Incidence of Obesity. Int. J. Mol. Sci. 2023, 24, 3758. https://doi.org/10.3390/ijms24043758
Kwak J, Shin D. Gene–Nutrient Interactions in Obesity: COBLL1 Genetic Variants Interact with Dietary Fat Intake to Modulate the Incidence of Obesity. International Journal of Molecular Sciences. 2023; 24(4):3758. https://doi.org/10.3390/ijms24043758
Chicago/Turabian StyleKwak, Junkyung, and Dayeon Shin. 2023. "Gene–Nutrient Interactions in Obesity: COBLL1 Genetic Variants Interact with Dietary Fat Intake to Modulate the Incidence of Obesity" International Journal of Molecular Sciences 24, no. 4: 3758. https://doi.org/10.3390/ijms24043758