

Stable Expression of dmiR-283 in the Brain Promises Positive Effects in Endurance Exercise on Sleep–Wake Behavior in Aging Drosophila


Abstract
:1. Introduction
2. Results
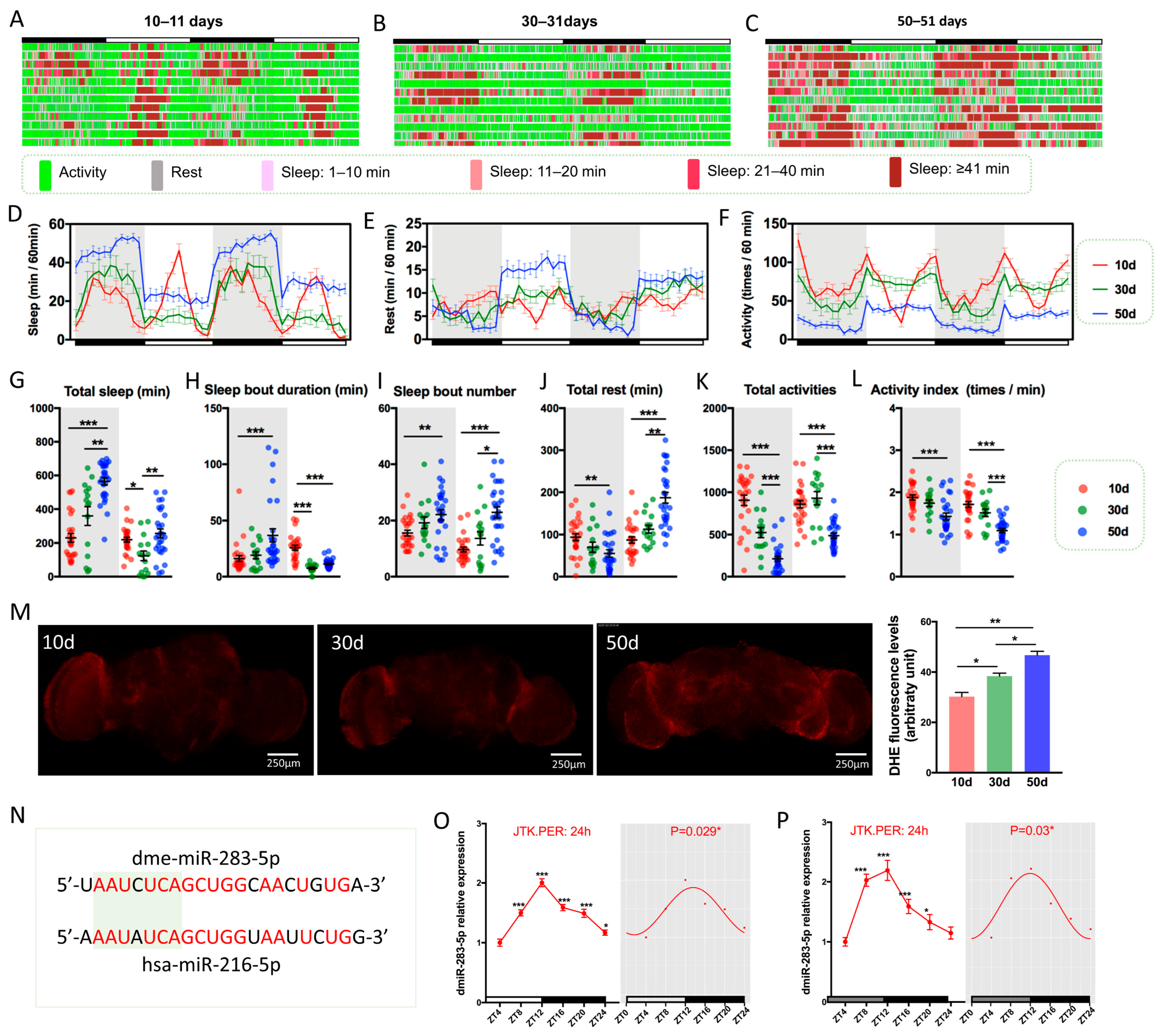
2.1. Drosophila Exhibits Age-Dependent Decline in Sleep–Wake Behavior during Aging
2.2. dmiR-283 Is Rhythmically Expressed in Drosophila Brains
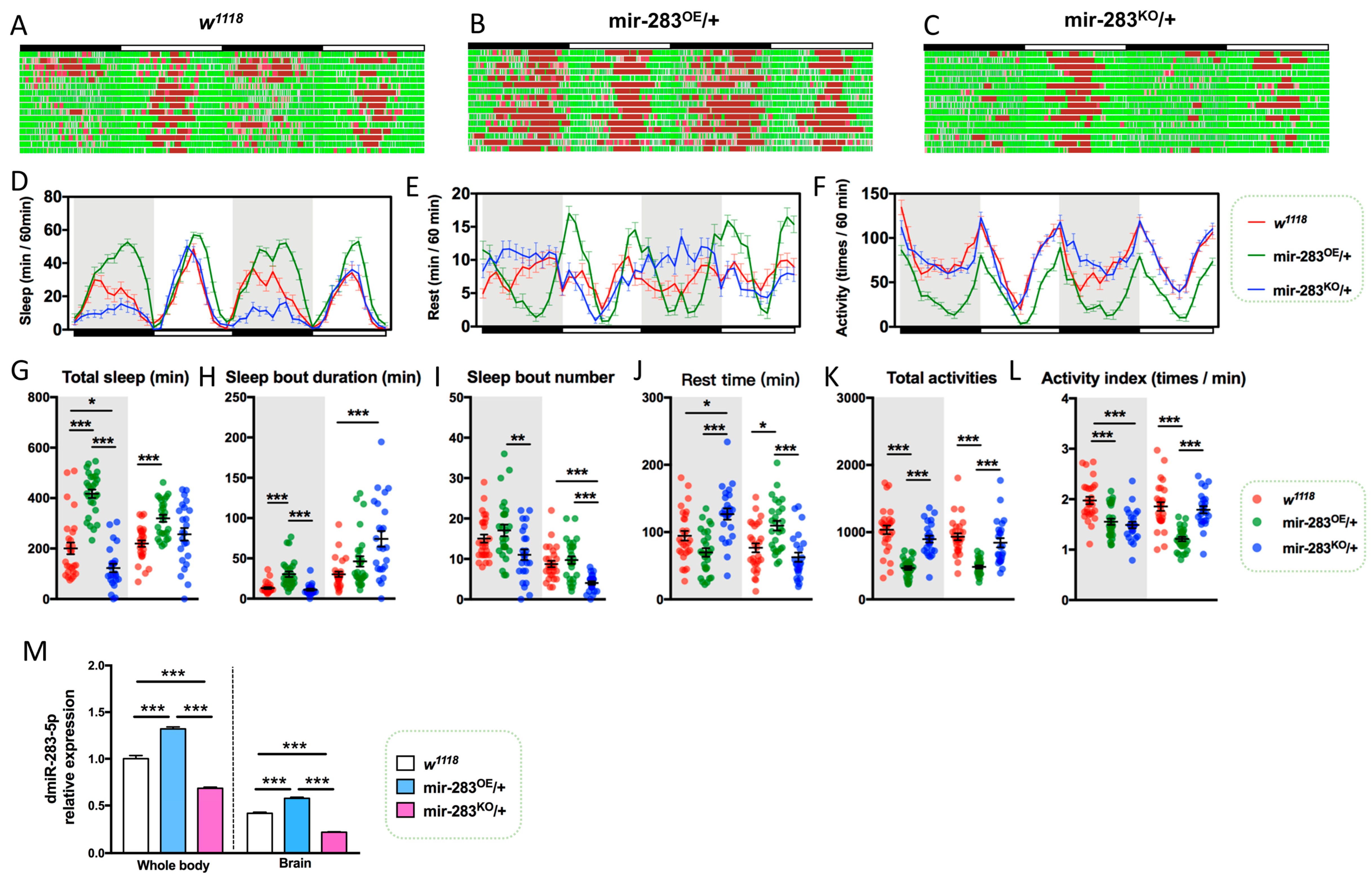
2.3. LNvs-Specific Knockdown of dmiR-283 Induces the Aging Sleep–Wake Behavior
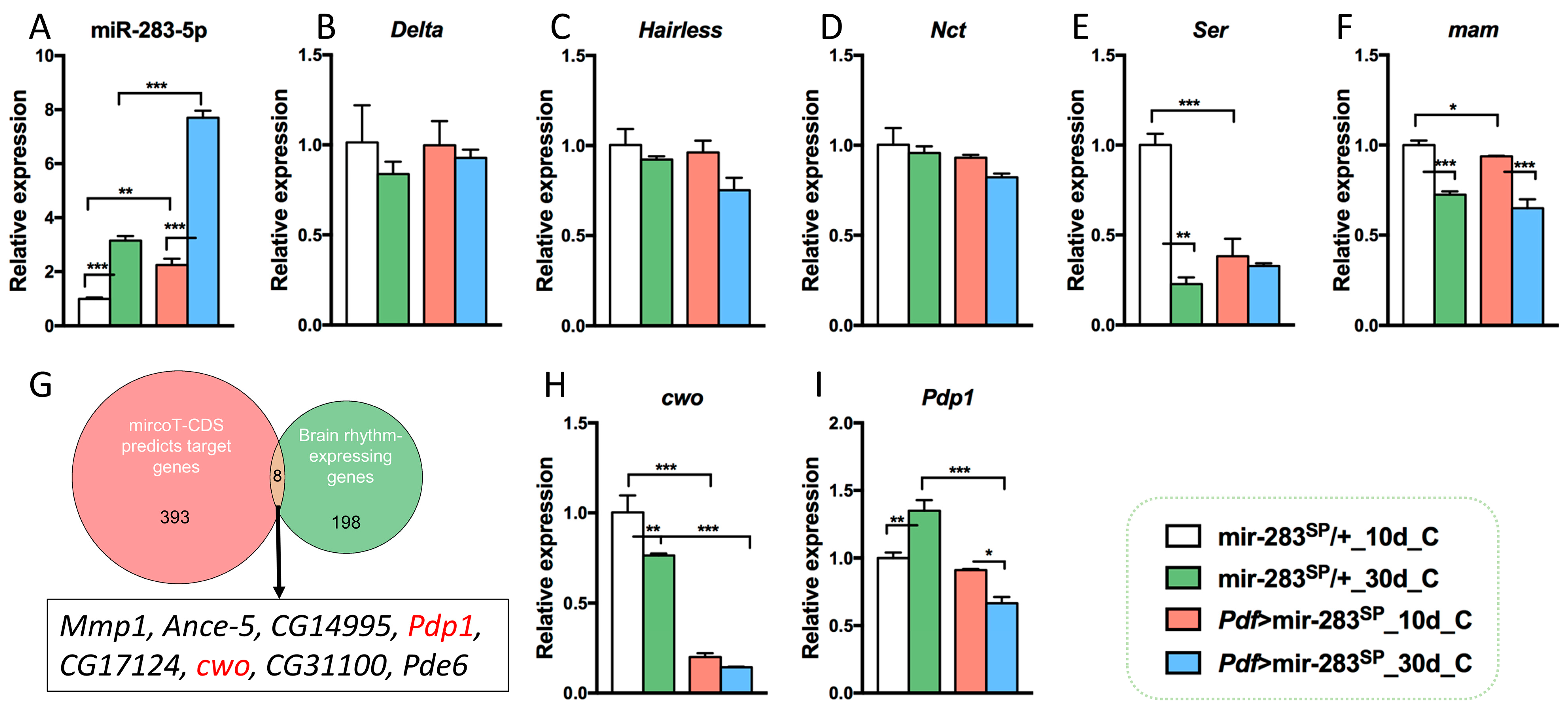
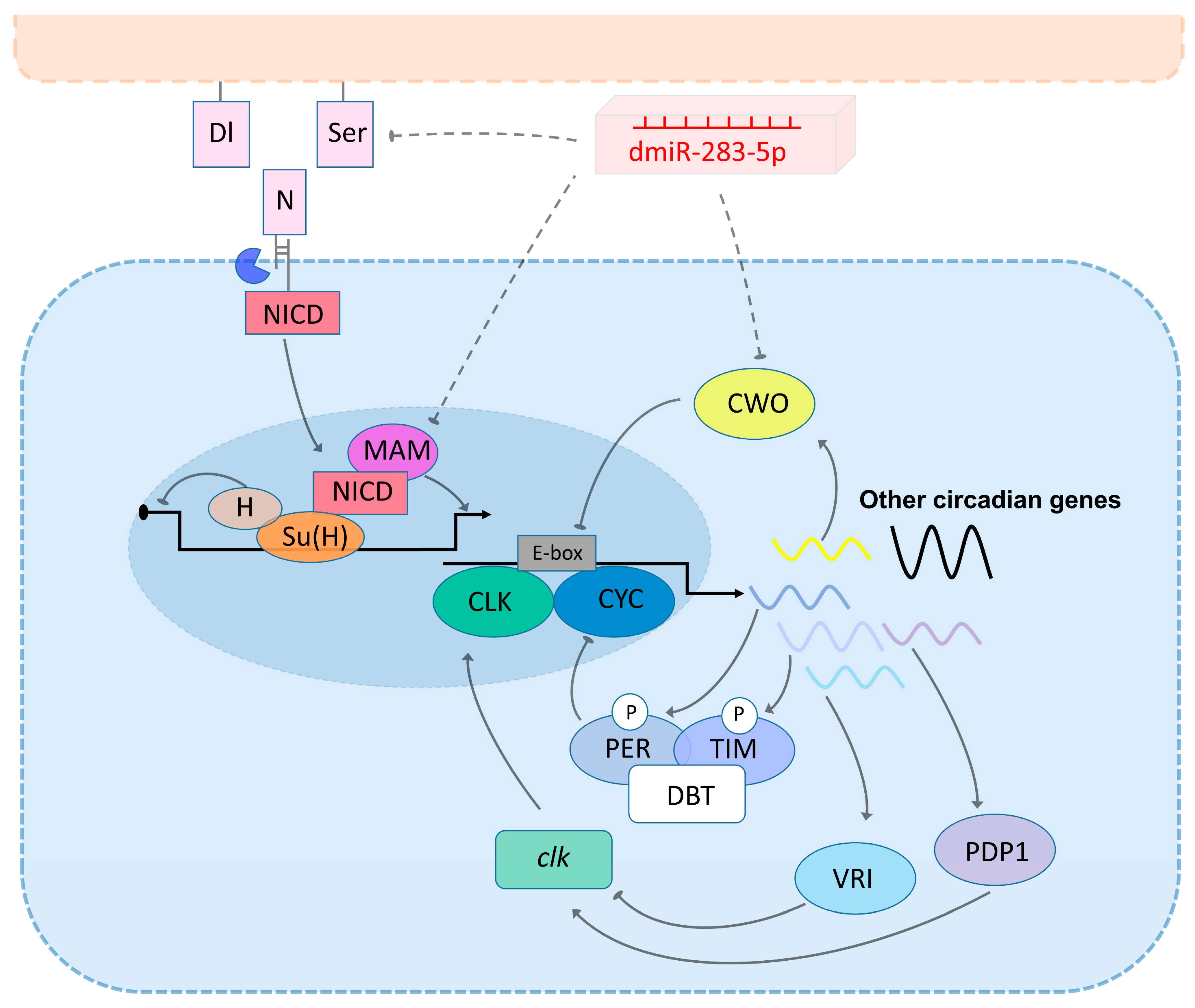
2.4. Prospective Mechanisms of dmiR-283 to Regulate age-Dependent Circadian Rhythm Decline and Sleep Prolongation
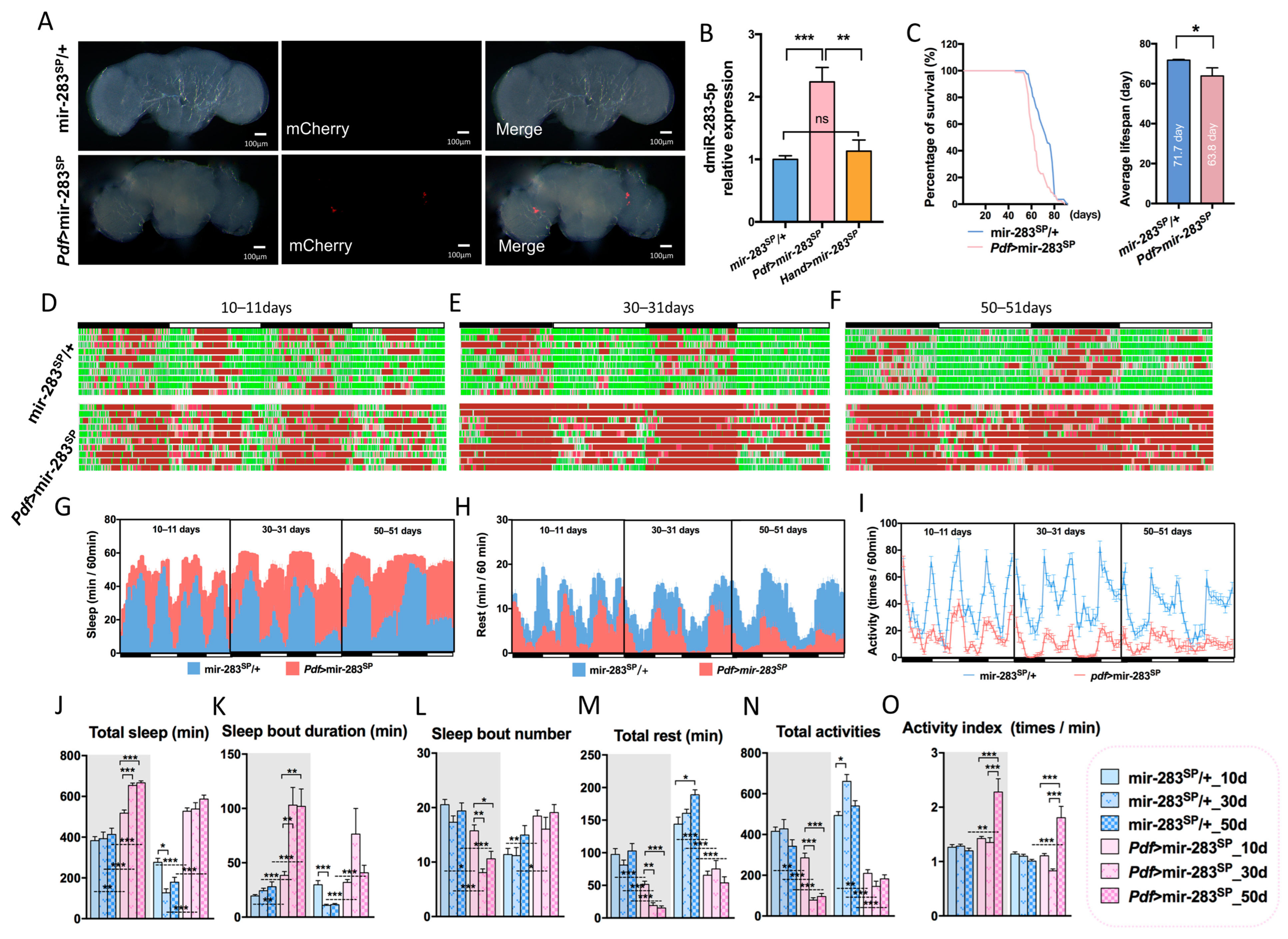
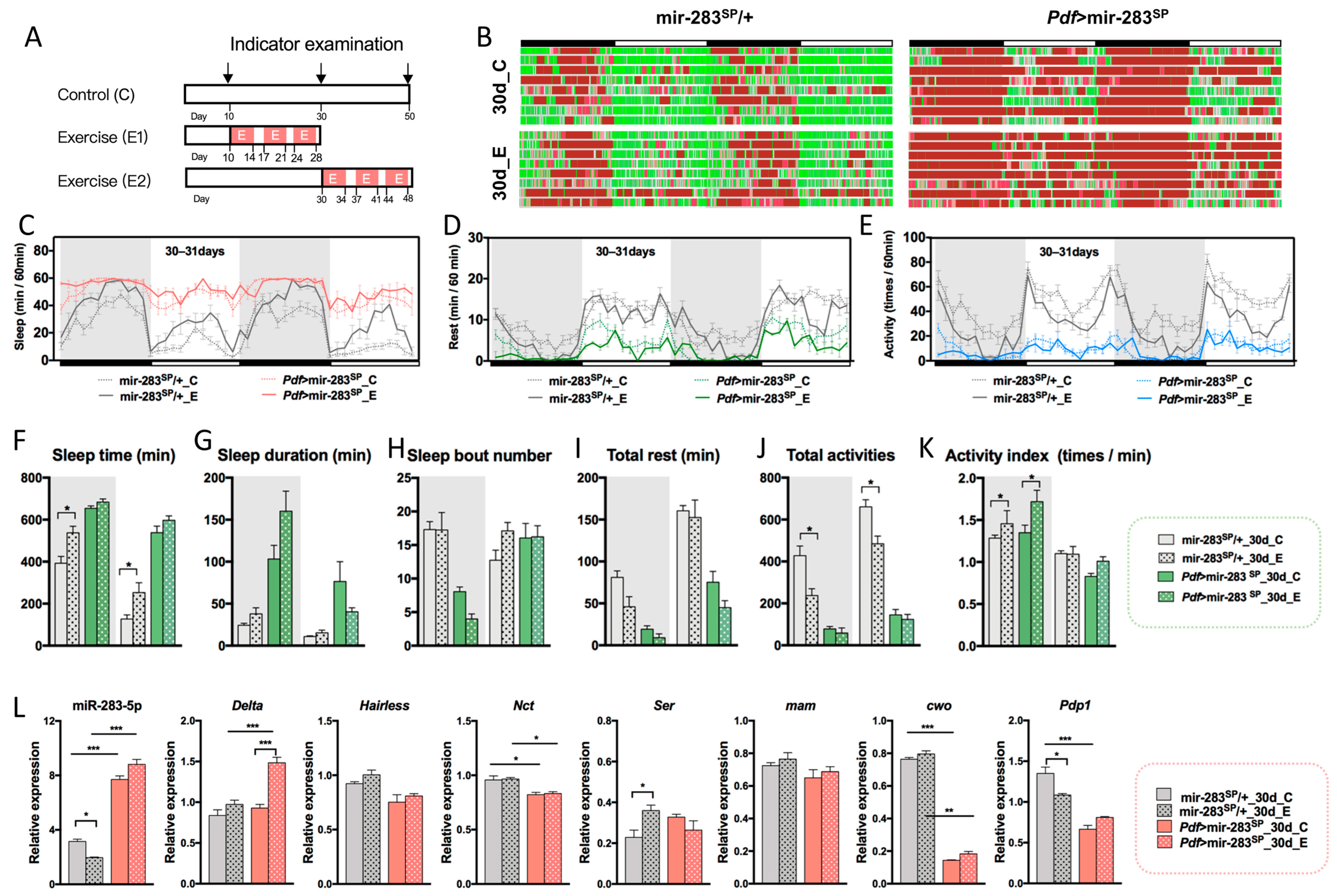
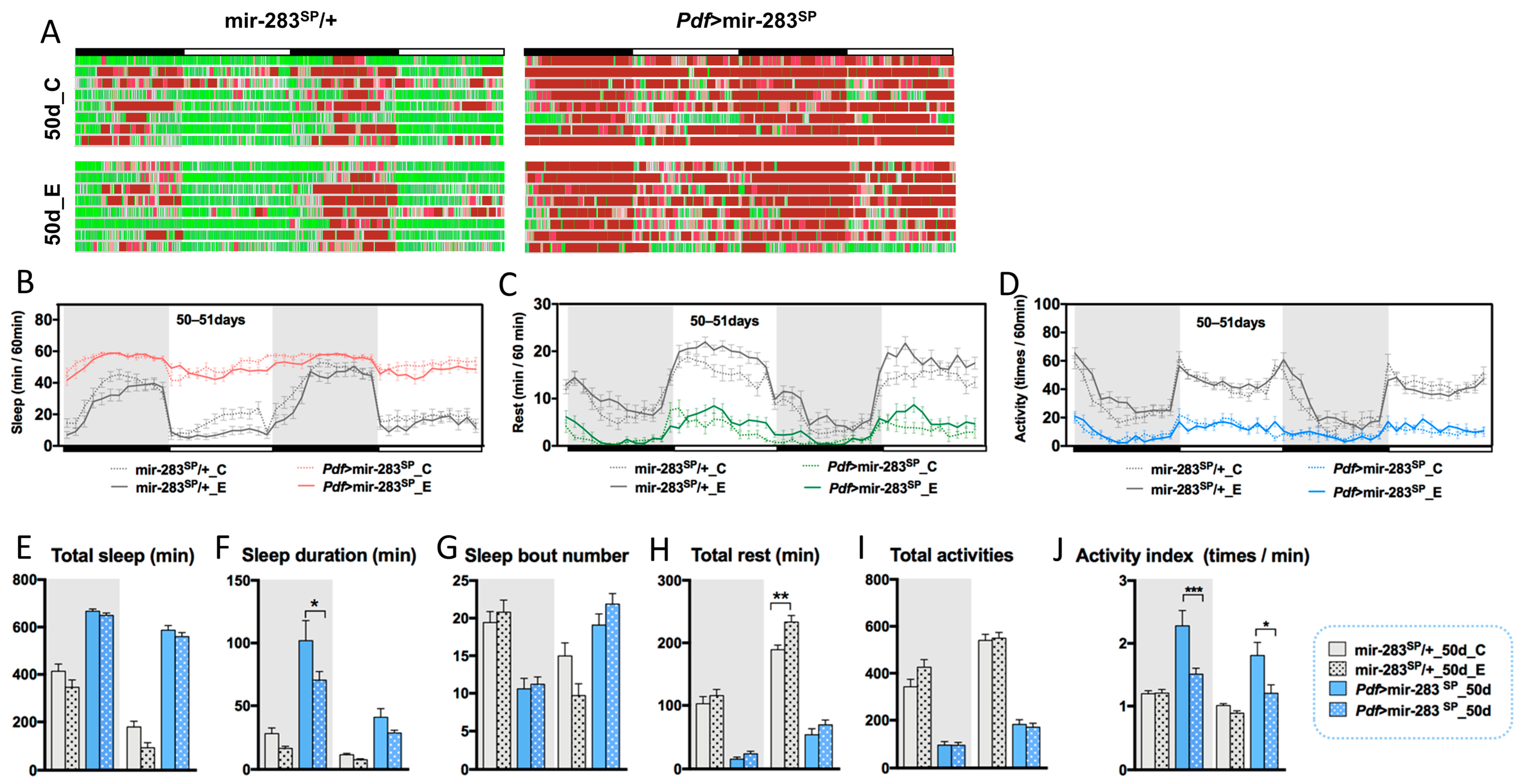
2.5. Expression Levels of dmiR-283 in the Brain Play a Key Role in Response to Exercise Effects
3. Discussion
4. Materials and Methods
4.1. Drosophila Strains, Crosses, and Rearing
4.2. Exercise Training Program
4.3. Life Expectancy Statistics
4.4. Sleep–Wake Behavior Analysis
4.5. Circadian Rhythm Analysis
4.6. Tissue Sample Collection, Total RNA Extraction, and RT-qPCR
4.7. Reactive Oxygen (ROS) Testing
4.8. Fluorescence Imaging
4.9. Bioinformatics Analysis
4.10. Data Analysis and Processing
5. Conclusions
6. Patents
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Musiek, E.S.; Holtzman, D.M. Mechanisms linking circadian clocks, sleep, and neurodegeneration. Science 2016, 354, 1004–1008. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Panagiotou, M.; Michel, S.; Meijer, J.H.; Deboer, T. The aging brain: Sleep, the circadian clock and exercise. Biochem. Pharmacol. 2021, 191, 114563. [Google Scholar] [CrossRef] [PubMed]
- Kondratov, R.V.; Kondratova, A.A.; Gorbacheva, V.Y.; Vykhovanets, O.V.; Antoch, M.P. Early aging and age-related pathologies in mice deficient in BMAL1, the core componentof the circadian clock. Genes Dev. 2006, 20, 1868–1873. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sterniczuk, R.; Theou, O.; Rusak, B.; Rockwood, K. Sleep Disturbance is Associated with Incident Dementia and Mortality. Curr. Alzheimer Res. 2013, 10, 767–775. [Google Scholar] [CrossRef]
- Lim, A.S.; Kowgier, M.; Yu, L.; Buchman, A.S.; Bennett, D.A. Sleep Fragmentation and the Risk of Incident Alzheimer’s Disease and Cognitive Decline in Older Persons. Sleep 2013, 36, 1027–1032. [Google Scholar] [CrossRef] [Green Version]
- Sowell, E.R.; Peterson, B.S.; Thompson, P.M.; Welcome, S.E.; Henkenius, A.L.; Toga, A.W. Mapping cortical change across the human life span. Nat. Neurosci. 2003, 6, 309–315. [Google Scholar] [CrossRef]
- Kinser, H.E.; Pincus, Z. MicroRNAs as modulators of longevity and the aging process. Hum. Genet. 2020, 139, 291–308. [Google Scholar] [CrossRef]
- Garg, D.; Cohen, S.M. miRNAs and aging: A genetic perspective. Ageing Res. Rev. 2014, 17, 3–8. [Google Scholar] [CrossRef]
- Kumar, S.; Vijayan, M.; Bhatti, J.S.; Reddy, P.H. MicroRNAs as Peripheral Biomarkers in Aging and Age-Related Diseases. Prog. Mol. Biol. Transl. 2017, 146, 47–94. [Google Scholar] [CrossRef]
- Wagner, K.H.; Cameron-Smith, D.; Wessner, B.; Franzke, B. Biomarkers of Aging: From Function to Molecular Biology. Nutrients 2016, 8, 338. [Google Scholar] [CrossRef] [Green Version]
- Chen, X.; Rosbash, M. MicroRNA-92a is a circadian modulator of neuronal excitability in Drosophila. Nat. Commun. 2017, 8, 14707. [Google Scholar] [CrossRef] [Green Version]
- Sha, N.; Wang, H.W.; Sun, B.; Gong, M.; Miao, P.; Jiang, X.L.; Yang, X.F.; Li, M.; Xu, L.X.; Feng, C.X.; et al. The role of pineal microRNA-325 in regulating circadian rhythms after neonatal hypoxic-ischemic brain damage. Neural Regen. Res. 2021, 16, 2071. [Google Scholar] [CrossRef]
- Davis, C.J.; Bohnet, S.G.; Meyerson, J.M.; Krueger, J.M. Sleep loss changes microRNA levels in the brain: A possible mechanism for state-dependent translational regulation. Neurosci. Lett. 2007, 422, 68–73. [Google Scholar] [CrossRef] [Green Version]
- Davis, C.J.; Clinton, J.M.; Krueger, J.M. MicroRNA 138, let-7b, and 125a inhibitors differentially alter sleep and EEG delta-wave activity in rats. J. Appl. Physiol. 2012, 113, 1756–1762. [Google Scholar] [CrossRef] [Green Version]
- Holm, A.; Bang-Berthelsen, C.H.; Knudsen, S.; Kornum, B.R.; Modvig, S.; Jennum, P.; Gammeltoft, S. miRNA Profiles in Plasma from Patients with Sleep Disorders Reveal Dysregulation of miRNAs in Narcolepsy and Other Central Hypersomnias. Sleep 2014, 37, 1525–1533. [Google Scholar] [CrossRef] [Green Version]
- Shafer, O.T.; Keene, A.C. The Regulation of Drosophila Sleep. Curr. Biol. 2021, 31, R38–R49. [Google Scholar] [CrossRef]
- Vienne, J.; Spann, R.; Guo, F.; Rosbash, M. Age-Related Reduction of Recovery Sleep and Arousal Threshold in Drosophila. Sleep 2016, 39, 1613–1624. [Google Scholar] [CrossRef] [Green Version]
- De Nobrega, A.K.; Lyons, L.C. Aging and the clock: Perspective from flies to humans. Eur. J. Neurosci. 2020, 51, 454–481. [Google Scholar] [CrossRef] [Green Version]
- Gabriel, B.M.; Zierath, J.R. Circadian rhythms and exercise—Re-setting the clock in metabolic disease. Nat. Rev. Endocrinol. 2019, 15, 197–206. [Google Scholar] [CrossRef]
- Koh, K.; Evans, J.M.; Hendricks, J.C.; Sehgal, A. A Drosophila model for age-associated changes in sleep:wake cycles. Proc. Natl. Acad. Sci. USA 2006, 103, 13843–13847. [Google Scholar] [CrossRef] [Green Version]
- Grandner, M.A.; Drummond, S.P.A. Who are the long sleepers? Towards an understanding of the mortality relationship. Sleep Med. Rev. 2007, 11, 341–360. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Andretic, R.; Shaw, P.J. Essentials of sleep recordings in Drosophila: Moving beyond sleep time. Method Enzymol. 2005, 393, 759–772. [Google Scholar] [CrossRef]
- Chiu, J.C.; Low, K.H.; Pike, D.H.; Yildirim, E.; Edery, I. Assaying locomotor activity to study circadian rhythms and sleep parameters in Drosophila. J. Vis. Exp. 2010, 43, e2157. [Google Scholar] [CrossRef] [Green Version]
- Vaccaro, A.; Issa, A.R.; Seugnet, L.; Birman, S.; Kiarsfeid, A. Drosophila Clock Is Required in Brain Pacemaker Neurons to Prevent Premature Locomotor Aging Independently of Its Circadian Function. PLoS Genet. 2017, 13, e1006507. [Google Scholar] [CrossRef] [Green Version]
- Mendoza-Viveros, L.; Bouchard-Cannon, P.; Hegazi, S.; Cheng, A.H.; Pastore, S.; Cheng, H.M. Molecular modulators of the circadian clock: Lessons from flies and mice. Cell. Mol. Life Sci. 2017, 74, 1035–1059. [Google Scholar] [CrossRef]
- Mehta, N.; Cheng, H.Y. Micro-managing the circadian clock: The role of microRNAs in biological timekeeping. J. Mol. Biol. 2013, 425, 3609–3624. [Google Scholar] [CrossRef]
- Aboobaker, A.A.; Tomancak, P.; Patel, N.; Rubin, G.M.; Lai, E.C. Drosophila microRNAs exhibit diverse spatial expression patterns during embryonic development. Proc. Natl. Acad. Sci. USA 2005, 102, 18017–18022. [Google Scholar] [CrossRef] [Green Version]
- Xue, Y.B.; Zhang, Y. Emerging roles for microRNA in the regulation of Drosophila circadian clock. BMC Neurosci. 2018, 19. [Google Scholar] [CrossRef] [Green Version]
- Kula-Eversole, E.; Nagoshi, E.; Shang, Y.; Rodriguez, J.; Allada, R.; Rosbash, M. Surprising gene expression patterns within and between PDF-containing circadian neurons in Drosophila. Proc. Natl. Acad. Sci. USA 2010, 107, 13497–13502. [Google Scholar] [CrossRef] [Green Version]
- Klarsfeld, A.; Rouyer, F. Effects of circadian mutations and LD periodicity on the life span of Drosophila melanogaster. J. Biol. Rhythms 1998, 13, 471–478. [Google Scholar] [CrossRef]
- Fernandes, J.; Varghese, J. Sexually dimorphic microRNA miR-190 regulates lifespan in male Drosophila. RNA Biol. 2022, 19, 1085–1093. [Google Scholar] [CrossRef]
- Esslinger, S.M.; Schwalb, B.; Helfer, S.; Michalik, K.M.; Witte, H.; Maier, K.C.; Martin, D.; Michalke, B.; Tresch, A.; Cramer, P.; et al. Drosophila miR-277 controls branched-chain amino acid catabolism and affects lifespan. RNA Biol. 2013, 10, 1042–1056. [Google Scholar] [CrossRef] [Green Version]
- Pandey, M.; Bansal, S.; Bar, S.; Yadav, A.K.; Sokol, N.S.; Tennessen, J.M.; Kapahi, P.; Chawla, G. miR-125-chinmo pathway regulates dietary restriction-dependent enhancement of lifespan in Drosophila. Elife 2021, 10, e62621. [Google Scholar] [CrossRef]
- Pittendrigh, C.S.; Minis, D.H. Circadian systems: Longevity as a function of circadian resonance in Drosophila melanogaster. Proc. Natl. Acad. Sci. USA 1972, 69, 1537–1539. [Google Scholar] [CrossRef] [Green Version]
- Hendricks, J.C.; Lu, S.; Kume, K.; Yin, J.C.; Yang, Z.; Sehgal, A. Gender dimorphism in the role of cycle (BMAL1) in rest, rest regulation, and longevity in Drosophila melanogaster. J. Biol. Rhythms 2003, 18, 12–25. [Google Scholar] [CrossRef]
- Krishnan, N.; Kretzschmar, D.; Rakshit, K.; Chow, E.; Giebultowicz, J.M. The circadian clock gene period extends healthspan in aging Drosophila melanogaster. Aging 2009, 1, 937–948. [Google Scholar] [CrossRef] [Green Version]
- Krishnan, N.; Rakshit, K.; Chow, E.S.; Wentzell, J.S.; Kretzschmar, D.; Giebultowicz, J.M. Loss of circadian clock accelerates aging in neurodegeneration-prone mutants. Neurobiol. Dis. 2012, 45, 1129–1135. [Google Scholar] [CrossRef] [Green Version]
- Katewa, S.D.; Akagi, K.; Bose, N.; Rakshit, K.; Camarella, T.; Zheng, X.; Hall, D.; Davis, S.; Nelson, C.S.; Brem, R.B.; et al. Peripheral Circadian Clocks Mediate Dietary Restriction-Dependent Changes in Lifespan and Fat Metabolism in Drosophila. Cell. Metab. 2016, 23, 143–154. [Google Scholar] [CrossRef] [Green Version]
- Menzel, P.; McCorkindale, A.L.; Stefanov, S.R.; Zinzen, R.P.; Meyer, I.M. Transcriptional dynamics of microRNAs and their targets during Drosophila neurogenesis. RNA Biol. 2019, 16, 69–81. [Google Scholar] [CrossRef] [Green Version]
- Salazar, J.L.; Yang, S.A.; Yamamoto, S. Post-Developmental Roles of Notch Signaling in the Nervous System. Biomolecules 2020, 10, 985. [Google Scholar] [CrossRef]
- Huang, H.; Zhu, C.T.; Skuja, L.L.; Hayden, D.J.; Hart, A.C. Genome-Wide Screen for Genes Involved in Caenorhabditis elegans Developmentally Timed Sleep. G3 Genes Genom. Genet. 2017, 7, 2907–2917. [Google Scholar] [CrossRef] [Green Version]
- Litovchenko, M.; Meireles-Filho, A.C.A.; Frochaux, M.V.; Bevers, R.P.J.; Prunotto, A.; Anduaga, A.M.; Hollis, B.; Gardeux, V.; Braman, V.S.; Russeil, J.M.C.; et al. Extensive tissue-specific expression variation and novel regulators underlying circadian behavior. Sci. Adv. 2021, 7, eabc3781. [Google Scholar] [CrossRef] [PubMed]
- Lim, C.; Chung, B.Y.; Pitman, J.L.; McGill, J.J.; Pradhan, S.; Lee, J.; Keegan, K.P.; Choe, J.; Allada, R. Clockwork orange encodes a transcriptional repressor important for circadian-clock amplitude in Drosophila. Curr. Biol. 2007, 17, 1082–1089. [Google Scholar] [CrossRef] [Green Version]
- Cyran, S.A.; Buchsbaum, A.M.; Reddy, K.L.; Lin, M.C.; Glossop, N.R.; Hardin, P.E.; Young, M.W.; Storti, R.V.; Blau, J. vrille, Pdp1, and dClock form a second feedback loop in the Drosophila circadian clock. Cell 2003, 112, 329–341. [Google Scholar] [CrossRef] [PubMed]
- Benito, J.; Zheng, H.; Hardin, P.E. PDP1epsilon functions downstream of the circadian oscillator to mediate behavioral rhythms. J. Neurosci. 2007, 27, 2539–2547. [Google Scholar] [CrossRef] [Green Version]
- Zheng, L.; Li, Q.F.; Ni, L.; Wang, H.; Ruan, X.C.; Wu, X.S. Lifetime regular exercise affects the incident of different arrhythmias and improves organismal health in aging female Drosophila melanogaster. Biogerontology 2017, 18, 97–108. [Google Scholar] [CrossRef]
- Zheng, L.; Feng, Y.; Wen, D.T.; Wang, H.; Wu, X.S. Fatiguing exercise initiated later in life reduces incidence of fibrillation and improves sleep quality in Drosophila. Age 2015, 37, 77. [Google Scholar] [CrossRef] [Green Version]
- Li, Q.F.; Wang, H.; Zheng, L.; Yang, F.; Li, H.Z.; Li, J.X.; Cheng, D.; Lu, K.; Liu, Y. Effects of Modest Hypoxia and Exercise on Cardiac Function, Sleep-Activity, Negative Geotaxis Behavior of Aged Female Drosophila. Front. Physiol. 2019, 10, 1610. [Google Scholar] [CrossRef] [Green Version]
- Ferreiro, M.J.; Perez, C.; Marchesano, M.; Ruiz, S.; Caputi, A.; Aguilera, P.; Barrio, R.; Cantera, R. Drosophila melanogaster White Mutant w(1118) Undergo Retinal Degeneration. Front. Neurosci. 2017, 11, 732. [Google Scholar] [CrossRef] [Green Version]
- Majumder, S.; Crabtree, J.S.; Golde, T.E.; Minter, L.M.; Osborne, B.A.; Miele, L. Targeting Notch in oncology: The path forward. Nat. Rev. Drug Discov. 2021, 20, 125–144. [Google Scholar] [CrossRef]
- Yamamoto, S.; Schulze, K.L.; Bellen, H.J. Introduction to Notch signaling. Methods Mol. Biol. 2014, 1187, 1–14. [Google Scholar] [CrossRef]
- Yang, S.A.; Deng, W.M. Serrate/Notch Signaling Regulates the Size of the Progenitor Cell Pool in Drosophila Imaginal Rings. Genetics 2018, 209, 829–843. [Google Scholar] [CrossRef] [Green Version]
- Maier, D. Hairless: The ignored antagonist of the Notch signalling pathway. Hereditas 2006, 143, 212–221. [Google Scholar] [CrossRef]
- Seugnet, L.; Suzuki, Y.; Merlin, G.; Gottschalk, L.; Duntley, S.P.; Shaw, P.J. Notch signaling modulates sleep homeostasis and learning after sleep deprivation in Drosophila. Curr. Biol. 2011, 21, 835–840. [Google Scholar] [CrossRef] [Green Version]
- Bennett, H.L.; Khoruzhik, Y.; Hayden, D.; Huang, H.; Sanders, J.; Walsh, M.B.; Biron, D.; Hart, A.C. Normal sleep bouts are not essential for C. elegans survival and FoxO is important for compensatory changes in sleep. BMC Neurosci. 2018, 19, 10. [Google Scholar] [CrossRef] [Green Version]
- Wadhwa, M.; Prabhakar, A.; Anand, J.P.; Ray, K.; Prasad, D.; Kumar, B.; Panjwani, U. Complement activation sustains neuroinflammation and deteriorates adult neurogenesis and spatial memory impairment in rat hippocampus following sleep deprivation. Brain Behav. Immun. 2019, 82, 129–144. [Google Scholar] [CrossRef]
- Ding, M.; Li, P.; Wen, Y.; Zhao, Y.; Cheng, B.; Zhang, L.; Ma, M.; Cheng, S.; Liu, L.; Du, Y.; et al. Integrative analysis of genome-wide association study and brain region related enhancer maps identifies biological pathways for insomnia. Prog. Neuropsychopharmacol. Biol. Psychiatry 2018, 86, 180–185. [Google Scholar] [CrossRef]
- Fathallah-Shaykh, H.M.; Bona, J.L.; Kadener, S. Mathematical model of the Drosophila circadian clock: Loop regulation and transcriptional integration. Biophys. J. 2009, 97, 2399–2408. [Google Scholar] [CrossRef] [Green Version]
- Kadener, S.; Stoleru, D.; McDonald, M.; Nawathean, P.; Rosbash, M. Clockwork Orange is a transcriptional repressor and a new Drosophila circadian pacemaker component. Gene Dev. 2007, 21, 1675–1686. [Google Scholar] [CrossRef] [Green Version]
- Zhao, J.; Kilman, V.L.; Keegan, K.P.; Peng, Y.; Emery, P.; Rosbash, M.; Allada, R. Drosophila clock can generate ectopic circadian clocks. Cell 2003, 113, 755–766. [Google Scholar] [CrossRef] [Green Version]
- Matsumoto, A.; Ukai-Tadenuma, M.; Yamada, R.G.; Houl, J.; Uno, K.D.; Kasukawa, T.; Dauwalder, B.; Itoh, T.Q.; Takahashi, K.; Ueda, R.; et al. A functional genomics strategy reveals clockwork orange as a transcriptional regulator in the Drosophila circadian clock. Genes Dev. 2007, 21, 1687–1700. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rivas, G.B.S.; Zhou, J.; Merlin, C.; Hardin, P.E. Clockwork orange promotes CLOCK-CYCLE activation via the putative Drosophila ortholog of clock interacting protein circadian. Curr. Biol. 2021, 31, 4207–4218. [Google Scholar] [CrossRef] [PubMed]
- Jensen, L.D.; Cao, Z.; Nakamura, M.; Yang, Y.; Brautigam, L.; Andersson, P.; Zhang, Y.; Wahlberg, E.; Lanne, T.; Hosaka, K.; et al. Opposing effects of circadian clock genes bmal1 and period2 in regulation of VEGF-dependent angiogenesis in developing zebrafish. Cell. Rep. 2012, 2, 231–241. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hillman, C.H.; Erickson, K.I.; Kramer, A.F. Be smart, exercise your heart: Exercise effects on brain and cognition. Nat. Rev. Neurosci. 2008, 9, 58–65. [Google Scholar] [CrossRef] [PubMed]
- Yang, P.Y.; Ho, K.H.; Chen, H.C.; Chien, M.Y. Exercise training improves sleep quality in middle-aged and older adults with sleep problems: A systematic review. J. Physiother. 2012, 58, 157–163. [Google Scholar] [CrossRef] [Green Version]
- Kredlow, M.A.; Capozzoli, M.C.; Hearon, B.A.; Calkins, A.W.; Otto, M.W. The effects of physical activity on sleep: A meta-analytic review. J. Behav. Med. 2015, 38, 427–449. [Google Scholar] [CrossRef]
- Aschoff, J. Exogenous and endogenous components in circadian rhythms. Cold Spring Harb. Symp. Quant. Biol. 1960, 25, 11–28. [Google Scholar] [CrossRef]
- Van Reeth, O.; Sturis, J.; Byrne, M.M.; Blackman, J.D.; L’Hermite-Baleriaux, M.; Leproult, R.; Oliner, C.; Refetoff, S.; Turek, F.W.; Van Cauter, E. Nocturnal exercise phase delays circadian rhythms of melatonin and thyrotropin secretion in normal men. Am. J. Physiol. 1994, 266, E964–E974. [Google Scholar] [CrossRef]
- Buxton, O.M.; Frank, S.A.; L’Hermite-Baleriaux, M.; Leproult, R.; Turek, F.W.; Van Cauter, E. Roles of intensity and duration of nocturnal exercise in causing phase delays of human circadian rhythms. Am. J. Physiol. 1997, 273, E536–E542. [Google Scholar] [CrossRef]
- Cassim, T.Z.; McGregor, K.M.; Nocera, J.R.; Garcia, V.V.; Sinon, C.G.; Kreuzer, M.; Garcia, P.S. Effects of exercise on the sleep microarchitecture in the aging brain: A study on a sedentary sample. Front. Syst. Neurosci. 2022, 16, 855107. [Google Scholar] [CrossRef]
- Park, I.; Diaz, J.; Matsumoto, S.; Iwayama, K.; Nabekura, Y.; Ogata, H.; Kayaba, M.; Aoyagi, A.; Yajima, K.; Satoh, M.; et al. Exercise improves the quality of slow-wave sleep by increasing slow-wave stability. Sci. Rep. 2021, 11, 4410. [Google Scholar] [CrossRef]
- Zheng, X.; Koh, K.; Sowcik, M.; Smith, C.J.; Chen, D.; Wu, M.N.; Sehgal, A. An isoform-specific mutant reveals a role of PDP1 epsilon in the circadian oscillator. J. Neurosci. 2009, 29, 10920–10927. [Google Scholar] [CrossRef] [Green Version]
- Weigelt, C.M.; Sehgal, R.; Tain, L.S.; Cheng, J.; Esser, J.; Pahl, A.; Dieterich, C.; Gronke, S.; Partridge, L. An Insulin-Sensitive Circular RNA that Regulates Lifespan in Drosophila. Mol. Cell. 2020, 79, 268–279. [Google Scholar] [CrossRef]
- Linford, N.J.; Chan, T.P.; Pletcher, S.D. Re-patterning sleep architecture in Drosophila through gustatory perception and nutritional quality. PLoS Genet. 2012, 8, e1002668. [Google Scholar] [CrossRef] [Green Version]
- Chung, B.Y.; Ro, J.; Hutter, S.A.; Miller, K.M.; Guduguntla, L.S.; Kondo, S.; Pletcher, S.D. Drosophila Neuropeptide F Signaling Independently Regulates Feeding and Sleep-Wake Behavior. Cell. Rep. 2017, 19, 2441–2450. [Google Scholar] [CrossRef] [Green Version]
- Hendricks, J.C.; Finn, S.M.; Panckeri, K.A.; Chavkin, J.; Williams, J.A.; Sehgal, A.; Pack, A.I. Rest in Drosophila is a sleep-like state. Neuron 2000, 25, 129–138. [Google Scholar] [CrossRef] [Green Version]
- Shaw, P.J.; Cirelli, C.; Greenspan, R.J.; Tononi, G. Correlates of sleep and waking in Drosophila melanogaster. Science 2000, 287, 1834–1837. [Google Scholar] [CrossRef] [Green Version]
- van Alphen, B.; Yap, M.H.; Kirszenblat, L.; Kottler, B.; van Swinderen, B. A dynamic deep sleep stage in Drosophila. J. Neurosci. 2013, 33, 6917–6927. [Google Scholar] [CrossRef] [Green Version]
- Parsons, R.; Parsons, R.; Garner, N.; Oster, H.; Rawashdeh, O. CircaCompare: A method to estimate and statistically support differences in mesor, amplitude and phase, between circadian rhythms. Bioinformatics 2020, 36, 1208–1212. [Google Scholar] [CrossRef]
- Williamson, W.R.; Hiesinger, P.R. Preparation of developing and adult Drosophila brains and retinae for live imaging. J. Vis. Exp. 2010, 37, e1936. [Google Scholar] [CrossRef] [Green Version]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Gene | Forward Primer | Backward Primer |
|---|---|---|
| U6 snRNA | 5′-TGGCCCCTGCGCAAGGATG-3′ | —— |
| dmiR-283-5p | 5′-TCTCGAATAGCGTTGTGACTGA-3′ | —— |
| rp49 | 5′-CTAAGCTGTCGCACAAATGG-3′ | 5′-AACTTCTTGAATCCGGTGGG-3′ |
| cwo | 5′-GCCGTATCGAGAAGACGGAG-3′ | 5′-TCCATGTAGCCACTCCGGTA-3′ |
| pdp1 | 5′-CGTCCCCAACACTGATCGAA-3′ | 5′-TGTGTTACCTTGAGGTCGGC-3′ |
| Delta | 5′-GGGTACCTTCTCGCTGATCG-3′ | 5′-GTCCAAATGAATCGTCGCGG-3′ |
| Hairless | 5′-TTGGTGGCGGTCTAAGTCAC-3′ | 5′-GACTCCGTTTTCCTCCAGCA-3′ |
| Nct | 5′-GAAAGTGCGCAACGTTTCCT-3′ | 5′-GGCCTTAAAGAGTGGGCAGT-3′ |
| Ser | 5′-CACAGCCACCGCGATTATTG-3′ | 5′-GCAATCGCGACCCTTGAATC-3′ |
| mam | 5′-CGATCTCGGCTCATTGGACA-3′ | 5′-AAGCCATCGAGGAAACTGGG-3′ |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Li, Q.; Wang, L.; Cao, Y.; Wang, X.; Tang, C.; Zheng, L. Stable Expression of dmiR-283 in the Brain Promises Positive Effects in Endurance Exercise on Sleep–Wake Behavior in Aging Drosophila. Int. J. Mol. Sci. 2023, 24, 4180. https://doi.org/10.3390/ijms24044180
Li Q, Wang L, Cao Y, Wang X, Tang C, Zheng L. Stable Expression of dmiR-283 in the Brain Promises Positive Effects in Endurance Exercise on Sleep–Wake Behavior in Aging Drosophila. International Journal of Molecular Sciences. 2023; 24(4):4180. https://doi.org/10.3390/ijms24044180
Chicago/Turabian StyleLi, Qiufang, Lingxiao Wang, Yurou Cao, Xiaoya Wang, Chao Tang, and Lan Zheng. 2023. "Stable Expression of dmiR-283 in the Brain Promises Positive Effects in Endurance Exercise on Sleep–Wake Behavior in Aging Drosophila" International Journal of Molecular Sciences 24, no. 4: 4180. https://doi.org/10.3390/ijms24044180