

ERRα Up-Regulates Invadopodia Formation by Targeting HMGCS1 to Promote Endometrial Cancer Invasion and Metastasis
 , ,
, ,
Abstract
:1. Introduction
2. Results
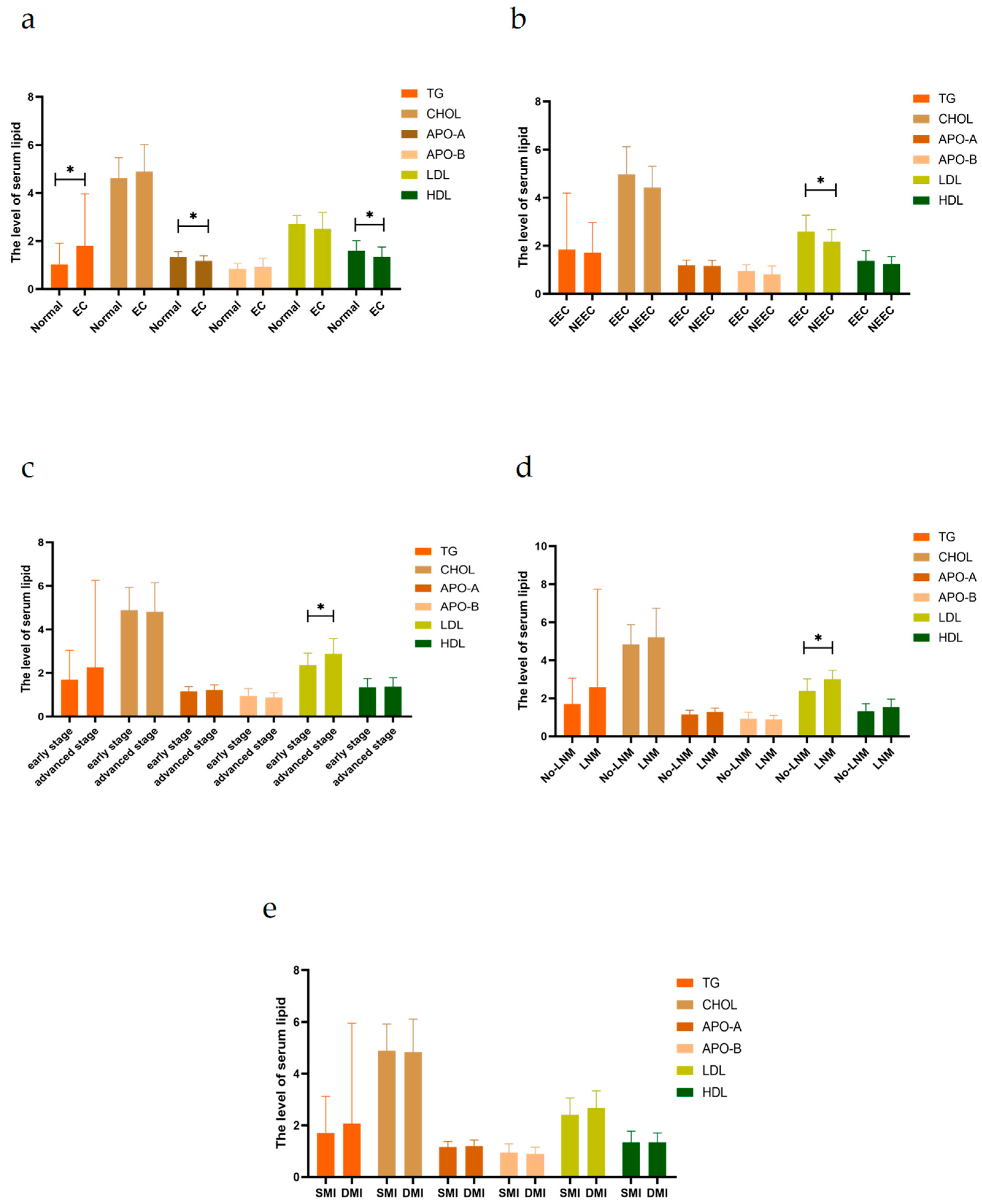
2.1. Serum Lipid Levels in Patients with EC Are Associated with the Progression of EC
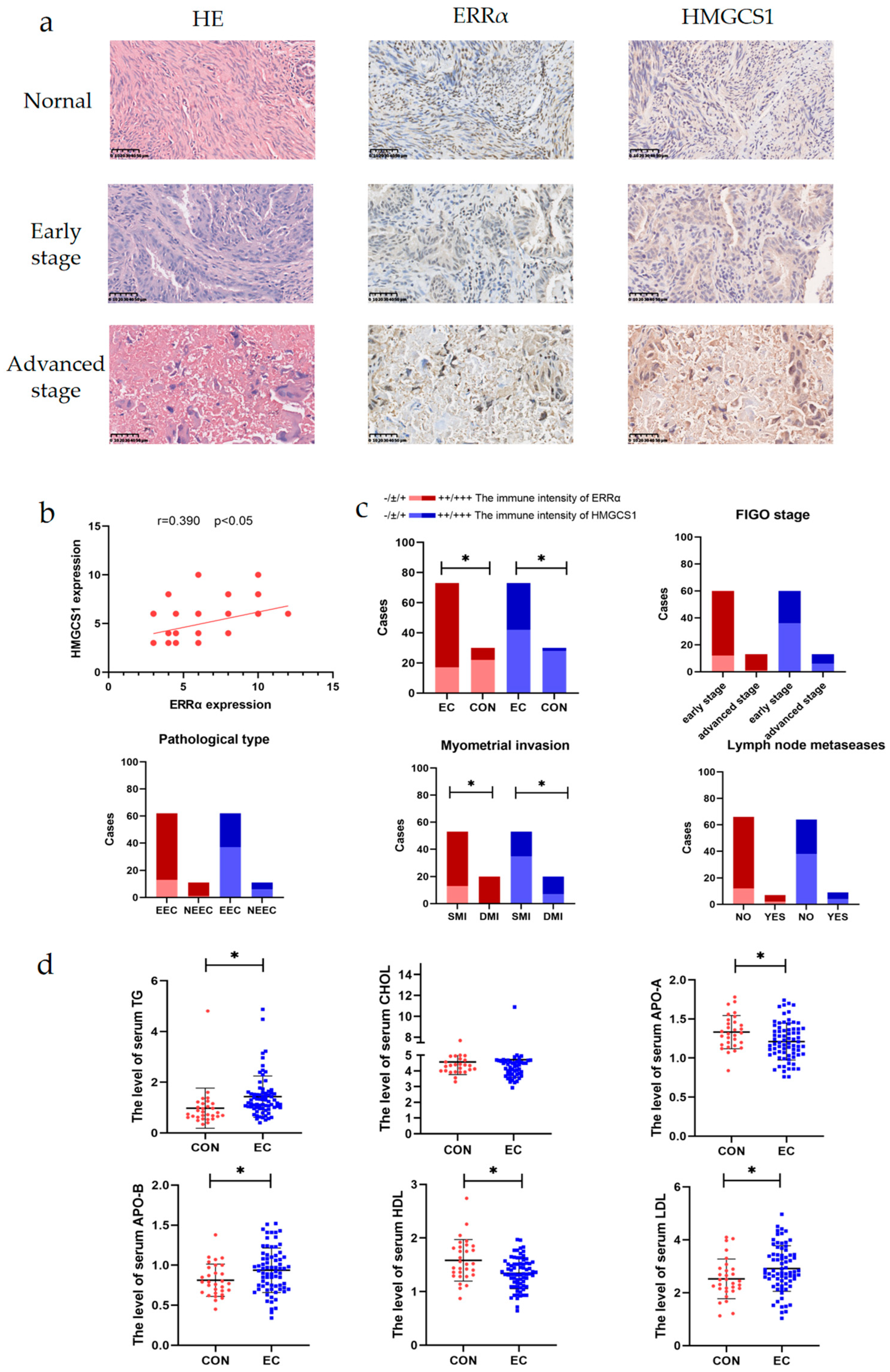
2.2. ERRα and HMGCS1 Are Highly Expressed in the Tissues of Patients with EC and Are Associated with Metastasis
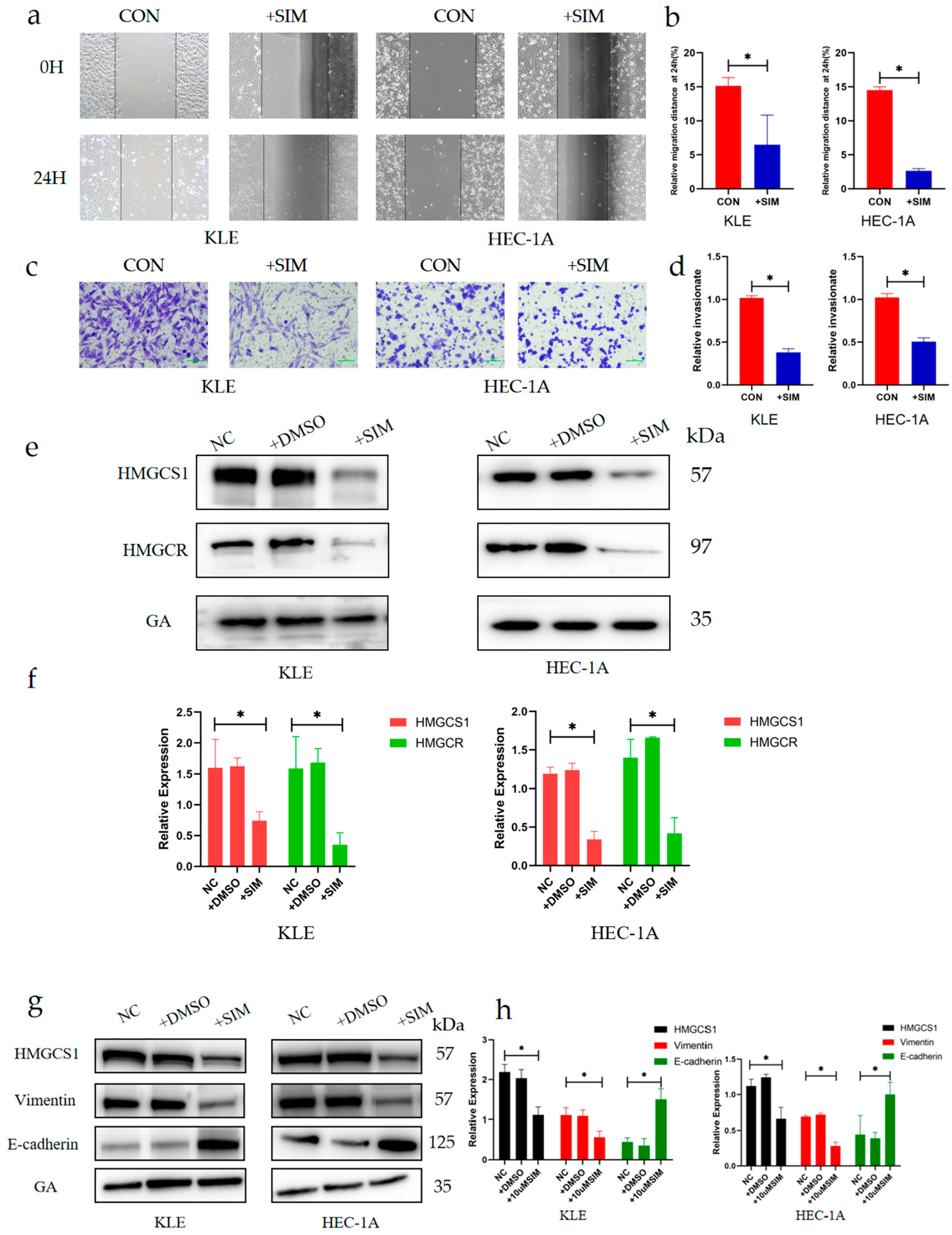
2.3. Simvastatin Inhibits Epithelial–Mesenchymal Transition in EC
2.4. ERRα Facilitates the Membrane Fluidity of EC Cells due to the Effects of HMGCS1 on EMT Signaling
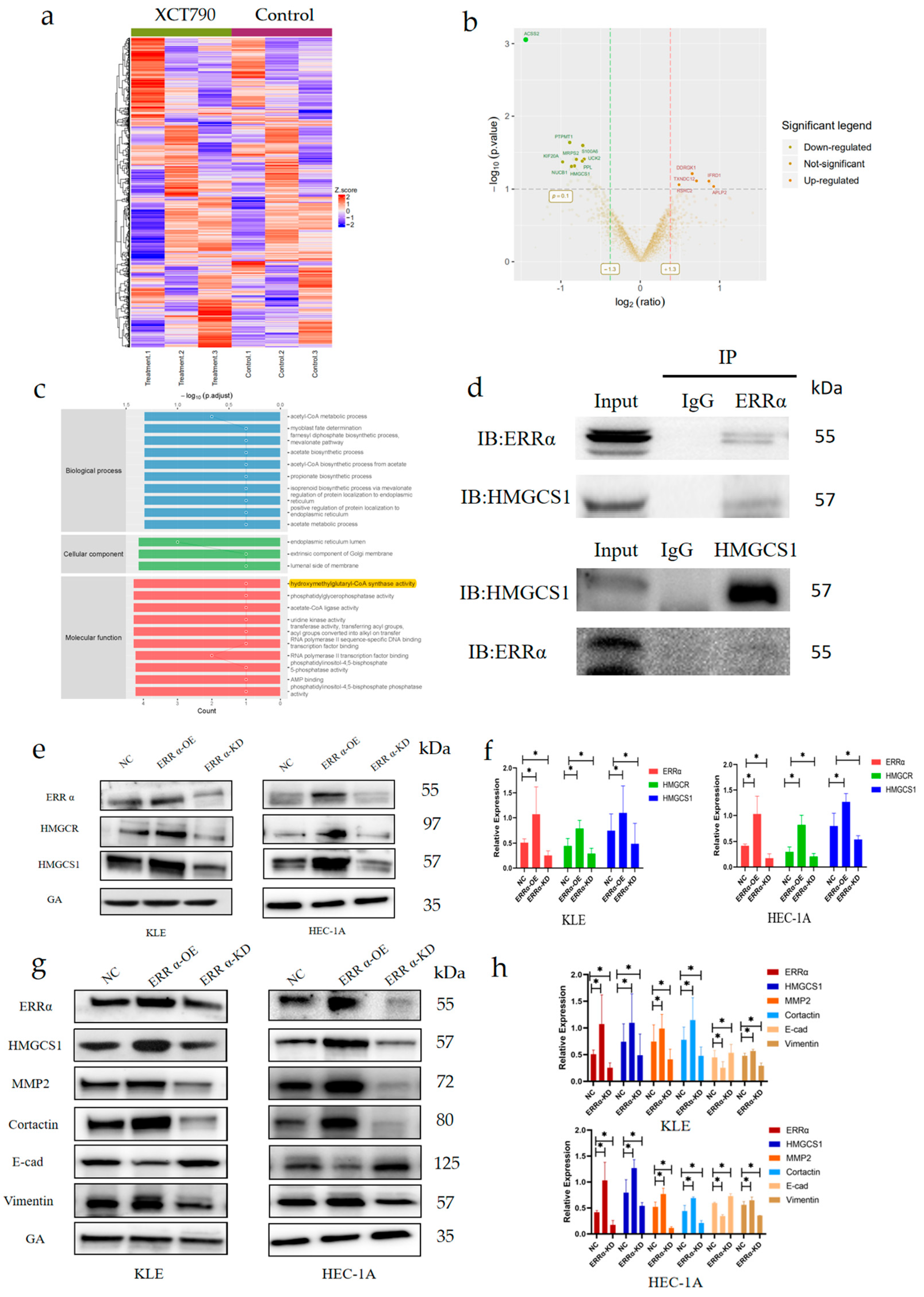
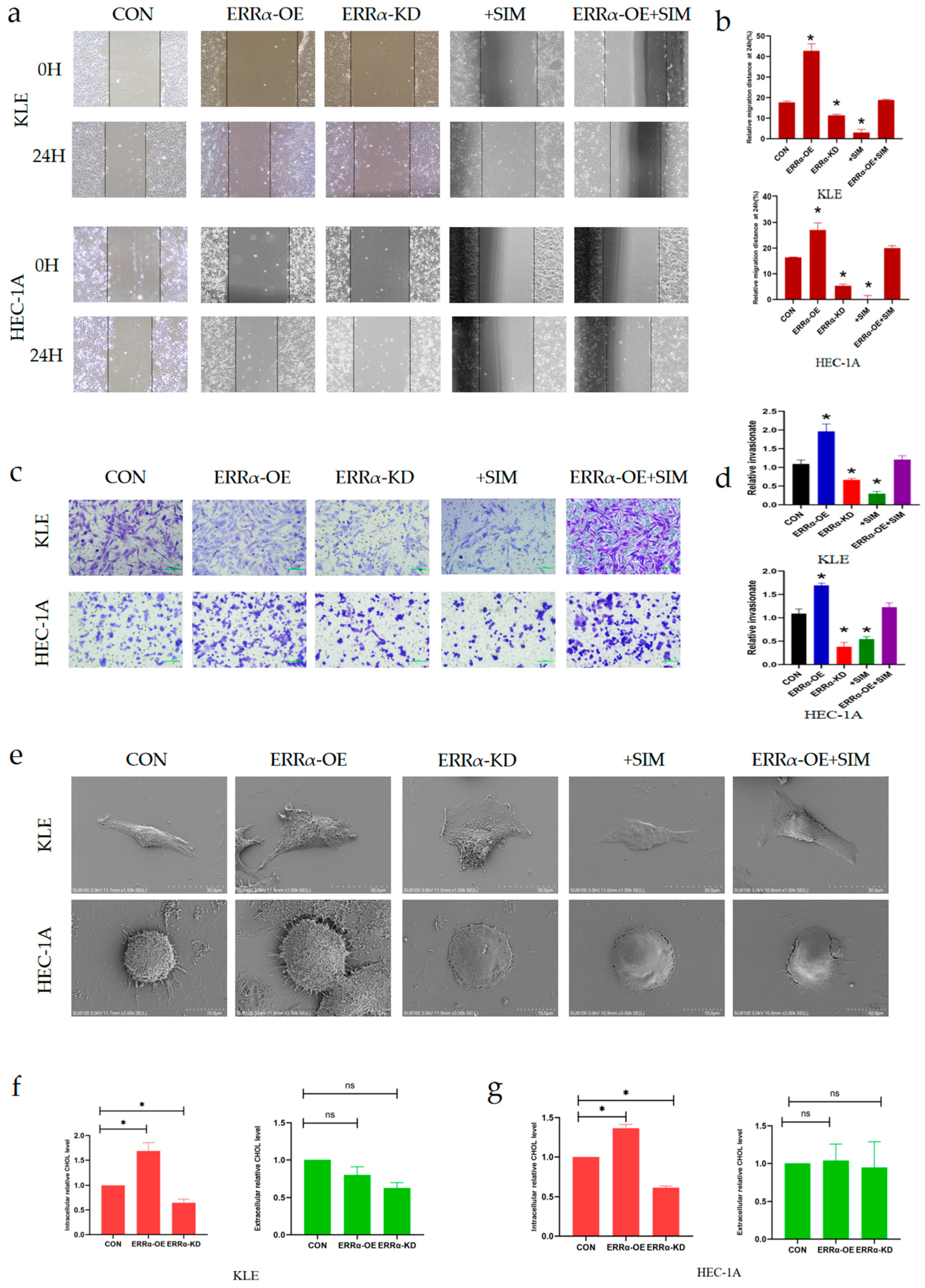
2.5. ERRα Promotes Intracellular Cholesterol Metabolism and Enhances the Formation of Invadopodia by Targeting HMGCS1
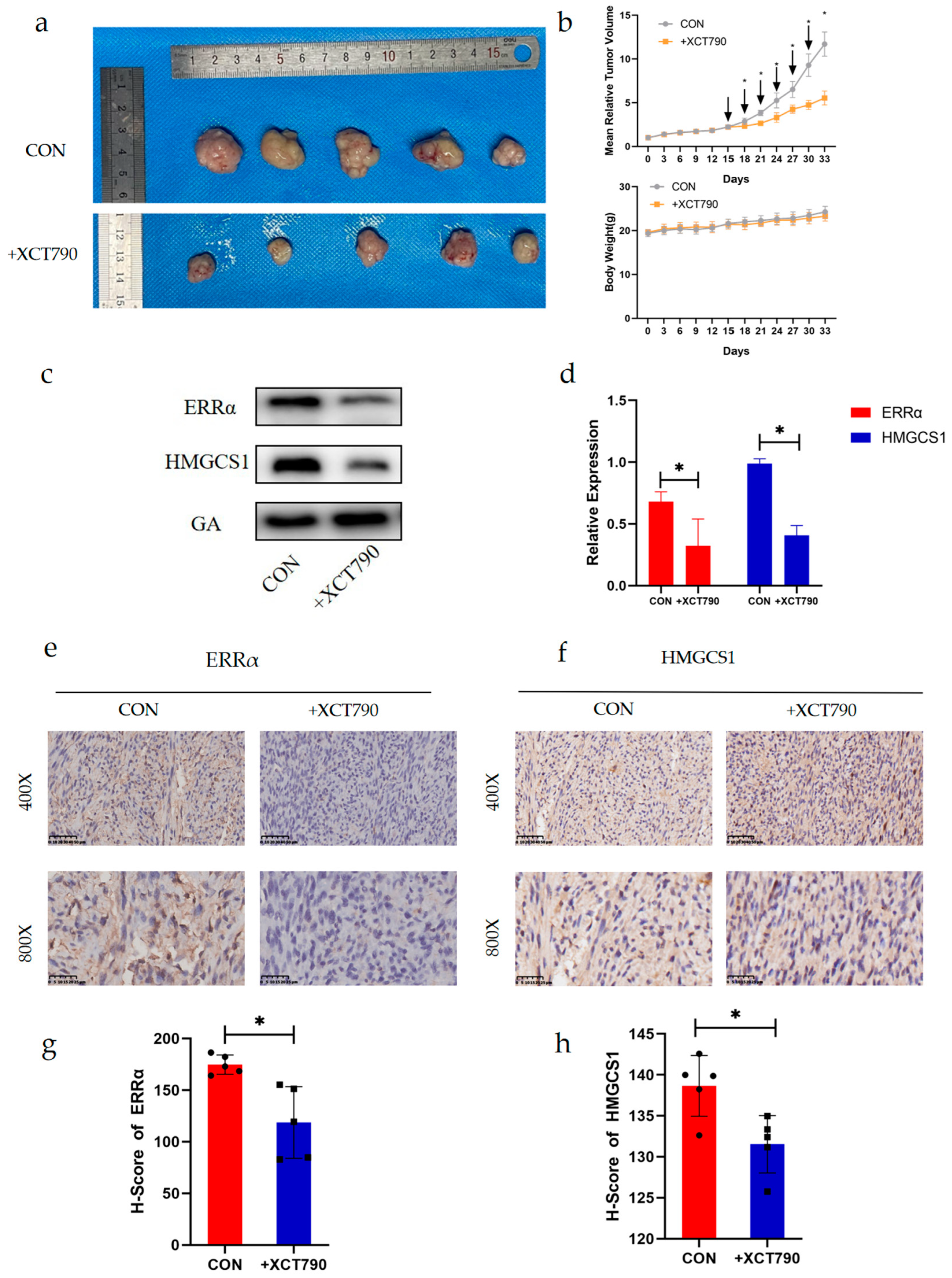
2.6. XCT790 Has an Inhibitory Effect on EC In Vivo
3. Discussion
4. Materials and Methods
4.1. Cell Lines and Cell Culture
4.2. Immunohistochemistry (IHC) on Patients Tissues
4.3. Western Blotting (WB)
4.4. Wound Healing Assay
4.5. Transwell Chamber Invasion Assay
4.6. Tandem Mass Tag (TMT) Labeling Proteomics
4.7. Co-Immunoprecipitation Assay (Co-IP)
4.8. Scanning Electron Microscope (SEM)
4.9. Cholesterol Quantitation Assay
4.10. Participants and Specimens
4.11. Animal Studies
4.12. Statistical Analysis
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Ferlay, J.; Colombet, M.; Soerjomataram, I.; Mathers, C.; Parkin, D.M.; Piñeros, M.; Znaor, A.; Bray, F. Estimating the global cancer incidence and mortality in 2018: GLOBOCAN sources and methods. Int. J. Cancer 2019, 144, 1941–1953. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Siegel, R.L.; Miller, K.D.; Jemal, A. Cancer statistics, 2019. CA Cancer J. Clin. 2019, 69, 7–34. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Siegel, R.L.; Miller, K.D.; Fuchs, H.E.; Jemal, A. Cancer Statistics, 2021. CA Cancer J. Clin. 2021, 71, 7–33. [Google Scholar] [CrossRef] [PubMed]
- Henley, S.J.; Ward, E.M.; Scott, S.; Ma, J.; Anderson, R.N.; Firth, A.U.; Thomas, C.C.; Islami, F.; Weir, H.K.; Lewis, D.R.; et al. Annual report to the nation on the status of cancer, part I: National cancer statistics. Cancer 2020, 126, 2225–2249. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Park, J.Y.; Nam, J.H.; Kim, Y.T.; Kim, Y.M.; Kim, J.H.; Kim, D.Y.; Sohn, I.; Lee, S.W.; Sung, C.O.; Kim, K.R. Poor prognosis of uterine serous carcinoma compared with grade 3 endometrioid carcinoma in early stage patients. Virchows Arch. 2013, 462, 289–296. [Google Scholar] [CrossRef] [PubMed]
- Kobori, T. New Insights into Immunotherapy for Gynecological Cancer. J. Clin. Med. 2022, 11, 4198. [Google Scholar] [CrossRef]
- Rizzo, A. Immune Checkpoint Inhibitors and Mismatch Repair Status in Advanced Endometrial Cancer: Elective Affinities. J. Clin. Med. 2022, 11, 3912. [Google Scholar] [CrossRef]
- Mc Auley, M.T. Effects of obesity on cholesterol metabolism and its implications for healthy ageing. Nutr. Res. Rev. 2020, 33, 121–133. [Google Scholar] [CrossRef]
- Guo, H.; Kong, W.; Zhang, L.; Han, J.; Clark, L.H.; Yin, Y.; Fang, Z.; Sun, W.; Wang, J.; Gilliam, T.P.; et al. Reversal of obesity-driven aggressiveness of endometrial cancer by metformin. Am. J. Cancer. Res. 2019, 9, 2170–2193. [Google Scholar]
- Kho, P.F.; Amant, F.; Annibali, D.; Ashton, K.; Attia, J.; Auer, P.L.; Beckmann, M.W.; Black, A.; Brinton, L.; Buchanan, D.D.; et al. Mendelian randomization analyses suggest a role for cholesterol in the development of endometrial cancer. Int. J. Cancer 2021, 148, 307–319. [Google Scholar] [CrossRef]
- Giguère, V. Orphan nuclear receptors: From gene to function. Endocr. Rev. 1999, 20, 689–725. [Google Scholar] [CrossRef] [PubMed]
- Liu, G.; Sun, P.; Dong, B.; Sehouli, J. Key regulator of cellular metabolism, estrogen-related receptor α, a new therapeutic target in endocrine-related gynecological tumor. Cancer Manag. Res. 2018, 10, 6887–6895. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tam, I.S.; Giguère, V. There and back again: The journey of the estrogen-related receptors in the cancer realm. J. Steroid Biochem. Mol. Biol. 2016, 157, 13–19. [Google Scholar] [CrossRef] [PubMed]
- Misawa, A.; Inoue, S. Estrogen-Related Receptors in Breast Cancer and Prostate Cancer. Front. Endocrinol. 2015, 6, 83. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- De Vitto, H.; Ryu, J.; Calderon-Aparicio, A.; Monts, J.; Dey, R.; Chakraborty, A.; Lee, M.H.; Bode, A.M.; Dong, Z. Estrogen-related receptor alpha directly binds to p53 and cooperatively controls colon cancer growth through the regulation of mitochondrial biogenesis and function. Cancer Metab. 2020, 8, 28. [Google Scholar] [CrossRef]
- Chen, L.; Mao, X.; Huang, M.; Lei, H.; Xue, L.; Sun, P. PGC-1α and ERRα in patients with endometrial cancer: A translational study for predicting myometrial invasion. Aging 2020, 12, 16963–16980. [Google Scholar] [CrossRef]
- Sharpe, L.; Brown, A. Controlling cholesterol synthesis beyond 3-hydroxy-3-methylglutaryl-CoA reductase (HMGCR). J. Biol. Chem. 2013, 288, 18707–18715. [Google Scholar] [CrossRef] [Green Version]
- Sapir, A.; Tsur, A.; Koorman, T.; Ching, K.; Mishra, P.; Bardenheier, A.; Podolsky, L.; Bening-Abu-Shach, U.; Boxem, M.; Chou, T.; et al. Controlled sumoylation of the mevalonate pathway enzyme HMGS-1 regulates metabolism during aging. Proc. Natl. Acad. Sci. USA 2014, 111, E3880–E3889. [Google Scholar] [CrossRef] [Green Version]
- Wang, I.; Huang, T.; Chen, J.; Chu, L.; Ping, Y.; Hsu, K.; Huang, K.; Fang, W.; Lee, H.; Chen, C.; et al. Mevalonate Pathway Enzyme HMGCS1 Contributes to Gastric Cancer Progression. Cancers 2020, 12, 1088. [Google Scholar] [CrossRef] [PubMed]
- Liu, Y.; Lu, L.L.; Wen, D.; Liu, D.L.; Dong, L.L.; Gao, D.M.; Bian, X.Y.; Zhou, J.; Fan, J.; Wu, W.Z. MiR-612 regulates invadopodia of hepatocellular carcinoma by HADHA-mediated lipid reprogramming. J. Hematol. Oncol. 2020, 13, 12. [Google Scholar] [CrossRef]
- Chimento, A.; Casaburi, I.; Avena, P.; Trotta, F.; De Luca, A.; Rago, V.; Pezzi, V.; Sirianni, R. Cholesterol and Its Metabolites in Tumor Growth: Therapeutic Potential of Statins in Cancer Treatment. Front. Endocrinol. 2018, 9, 807. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ding, X.; Zhang, W.; Li, S.; Yang, H. The role of cholesterol metabolism in cancer. Am. J. Cancer Res. 2019, 9, 219–227. [Google Scholar]
- Elustondo, P.; Martin, L.A.; Karten, B. Mitochondrial cholesterol import. Biochim. Biophys. Acta Mol. Cell Biol. Lipids 2017, 1862, 90–101. [Google Scholar] [CrossRef] [PubMed]
- Codini, M.; Garcia-Gil, M.; Albi, E. Cholesterol and Sphingolipid Enriched Lipid Rafts as Therapeutic Targets in Cancer. Int. J. Mol. Sci. 2021, 22, 726. [Google Scholar] [CrossRef] [PubMed]
- Leong, H.S.; Robertson, A.E.; Stoletov, K.; Leith, S.J.; Chin, C.A.; Chien, A.E.; Hague, M.N.; Ablack, A.; Carmine-Simmen, K.; McPherson, V.A.; et al. Invadopodia are required for cancer cell extravasation and are a therapeutic target for metastasis. Cell Rep. 2014, 8, 1558–1570. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kalluri, R.; Weinberg, R. The basics of epithelial-mesenchymal transition. J. Clin. Investig. 2009, 119, 1420–1428. [Google Scholar] [CrossRef] [Green Version]
- Schointuch, M.; Gilliam, T.; Stine, J.; Han, X.; Zhou, C.; Gehrig, P.; Kim, K.; Bae-Jump, V. Simvastatin, an HMG-CoA reductase inhibitor, exhibits anti-metastatic and anti-tumorigenic effects in endometrial cancer. Gynecol. Oncol. 2014, 134, 346–355. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Valcarcel-Jimenez, L.; Macchia, A.; Crosas-Molist, E.; Schaub-Clerigué, A.; Camacho, L.; Martín-Martín, N.; Cicogna, P.; Viera-Bardón, C.; Fernández-Ruiz, S.; Rodriguez-Hernandez, I.; et al. PGC1α Suppresses Prostate Cancer Cell Invasion through ERRα Transcriptional Control. Cancer Res. 2019, 79, 6153–6165. [Google Scholar] [CrossRef] [Green Version]
- Fielding, C.J.; Fielding, P.E. Cholesterol and caveolae: Structural and functional relationships. Biochim. Biophys. Acta 2000, 1529, 210–222. [Google Scholar] [CrossRef]
- Grundner, M.; Zemljič Jokhadar, S. Cytoskeleton modification and cholesterol depletion affect membrane properties and caveolae positioning of CHO cells. J. Membr. Biol. 2014, 247, 201–210. [Google Scholar] [CrossRef]
- Mao, X.; Lei, H.; Yi, T.; Su, P.; Tang, S.; Tong, Y.; Dong, B.; Ruan, G.; Mustea, A.; Sehouli, J.; et al. Lipid reprogramming induced by the TFEB-ERRα axis enhanced membrane fluidity to promote EC progression. J. Exp. Clin. Cancer Res. 2022, 41, 28. [Google Scholar] [CrossRef] [PubMed]
- Sun, P.; Mao, X.; Gao, M.; Huang, M.; Chen, L.; Ruan, G.; Huang, W.; Braicu, E.; Sehouli, J. Novel endocrine therapeutic strategy in endometrial carcinoma targeting estrogen-related receptor α by XCT790 and siRNA. Cancer Manag. Res. 2018, 10, 2521–2535. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, S.; Han, X. The Regulatory Role of Lipid Metabolism in Endometrial Cancer. J. Oncol. 2022, 2022, 6458877. [Google Scholar] [CrossRef] [PubMed]
- Onstad, M.; Schmandt, R.; Lu, K. Addressing the Role of Obesity in Endometrial Cancer Risk, Prevention, and Treatment. J. Clin. Oncol. Off. J. Am. Soc. Clin. Oncol. 2016, 34, 4225–4230. [Google Scholar] [CrossRef] [PubMed]
- Wang, S.; Huo, X. Comprehensive Analysis of ESRRA in Endometrial Cancer. Technol. Cancer Res. Treat. 2021, 20, 1533033821992083. [Google Scholar] [CrossRef] [PubMed]
- Ghanbari, F.; Fortier, A.; Park, M.; Philip, A. Cholesterol-Induced Metabolic Reprogramming in Breast Cancer Cells Is Mediated via the ERRα Pathway. Cancers 2021, 13, 2605. [Google Scholar] [CrossRef] [PubMed]
- Zhou, J.; Wang, Y.; Wu, D.; Wang, S.; Chen, Z.; Xiang, S.; Chan, F. Orphan nuclear receptors as regulators of intratumoral androgen biosynthesis in castration-resistant prostate cancer. Oncogene 2021, 40, 2625–2634. [Google Scholar] [CrossRef] [PubMed]
- Liu, C.; Chen, Y.; Chen, M.; Mao, X.; Dong, B.; Sun, P. A novel non-invasive molecular biomarker in ovarian endometriosis: Estrogen-related receptor α. Arch. Gynecol. Obstet. 2020, 302, 405–414. [Google Scholar] [CrossRef] [PubMed]
- Su, P.; Yu, L.; Mao, X.; Sun, P. Role of HIF-1α/ERRα in Enhancing Cancer Cell Metabolism and Promoting Resistance of Endometrial Cancer Cells to Pyroptosis. Front. Oncol. 2022, 12, 881252. [Google Scholar] [CrossRef] [PubMed]
- Tzing, S.F.; Bryant, M.P.; Wolfe, R.S. Factor 420-dependent pyridine nucleotide-linked formate metabolism of Methanobacterium ruminantium. J. Bacteriol. 1975, 121, 192–196. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Walsh, C.A.; Akrap, N.; Garre, E.; Magnusson, Y.; Harrison, H.; Andersson, D.; Jonasson, E.; Rafnsdottir, S.; Choudhry, H.; Buffa, F.; et al. The mevalonate precursor enzyme HMGCS1 is a novel marker and key mediator of cancer stem cell enrichment in luminal and basal models of breast cancer. PLoS ONE 2020, 15, e0236187. [Google Scholar] [CrossRef] [PubMed]
- Huang, B.; Song, B.L.; Xu, C. Cholesterol metabolism in cancer: Mechanisms and therapeutic opportunities. Nat. Metab. 2020, 2, 132–141. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yoriki, K.; Mori, T.; Kokabu, T.; Matsushima, H.; Umemura, S.; Tarumi, Y.; Kitawaki, J. Estrogen-related receptor alpha induces epithelial-mesenchymal transition through cancer-stromal interactions in endometrial cancer. Sci. Rep. 2019, 9, 6697. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Karamanou, K.; Franchi, M.; Vynios, D.; Brezillon, S. Epithelial-to-mesenchymal transition and invadopodia markers in breast cancer: Lumican a key regulator. Semin. Cancer Biol. 2020, 62, 125–133. [Google Scholar] [CrossRef] [PubMed]
- Alblazi, K.M.; Siar, C.H. Cellular protrusions-lamellipodia, filopodia, invadopodia and podosomes-and their roles in progression of orofacial tumours: Current understanding. Asian Pac. J. Cancer Prev. 2015, 16, 2187–2191. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Buttacavoli, M.; Di Cara, G.; Roz, E.; Pucci-Minafra, I.; Feo, S.; Cancemi, P. Integrated Multi-Omics Investigations of Metalloproteinases in Colon Cancer: Focus on MMP2 and MMP9. Int. J. Mol. Sci. 2021, 22, 12389. [Google Scholar] [CrossRef]
- Kessels, M.M.; Engqvist-Goldstein, A.E.; Drubin, D.G. Association of mouse actin-binding protein 1 (mAbp1/SH3P7), an Src kinase target, with dynamic regions of the cortical actin cytoskeleton in response to Rac1 activation. Mol. Biol. Cell 2000, 11, 393–412. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, Y.; Nolan, M.; Yamada, H.; Watanabe, M.; Nasu, Y.; Takei, K.; Takeda, T. Dynamin2 GTPase contributes to invadopodia formation in invasive bladder cancer cells. Biochem. Biophys. Res. Commun. 2016, 480, 409–414. [Google Scholar] [CrossRef] [Green Version]
- Caldieri, G.; Giacchetti, G.; Beznoussenko, G.; Attanasio, F.; Ayala, I.; Buccione, R. Invadopodia biogenesis is regulated by caveolin-mediated modulation of membrane cholesterol levels. J. Cell. Mol. Med. 2009, 13, 1728–1740. [Google Scholar] [CrossRef]
- Augoff, K.; Hryniewicz-Jankowska, A.; Tabola, R. Invadopodia: Clearing the way for cancer cell invasion. Ann. Transl. Med. 2020, 8, 902. [Google Scholar] [CrossRef]
- Kumar, N.; Mandal, C.C. Cholesterol-Lowering Drugs on Akt Signaling for Prevention of Tumorigenesis. Front. Genet. 2021, 12, 724149. [Google Scholar] [CrossRef] [PubMed]
- Mayengbam, S.S.; Singh, A.; Pillai, A.D.; Bhat, M.K. Influence of cholesterol on cancer progression and therapy. Transl. Oncol. 2021, 14, 101043. [Google Scholar] [CrossRef] [PubMed]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Number of Cases | Ratio (%) | ||
|---|---|---|---|
| FIGO Staging | Stage I | 47 | 64.38 |
| Stage II | 13 | 17.81 | |
| Stage III-IV | 13 | 17.81 | |
| Pathological Type | EEC | 61 | 83.56 |
| I | 31 | 50.82 | |
| II | 28 | 45.90 | |
| III | 5 | 8.20 | |
| NEEC | 16 | 21.92 | |
| Myometrial Invasion | <1/2 | 53 | 72.60 |
| ≥1/2 | 20 | 27.40 | |
| Lymph Node Metastasis | NO | 66 | 90.41 |
| YES | 7 | 9.6 | |
| Total | 73 | 100 |
| ERRα Expression | HMGCS1 Expression | ERRα/HMGCS1 Expression | |||||||
|---|---|---|---|---|---|---|---|---|---|
| −/±/+ | ++/+++ | p | −/±/+ | ++/+++ | p | Both ++/+++ | Either −/±/+ | p | |
| Nornal | 22 | 8 | p < 0.001 * | 28 | 2 | p < 0.001 * | 0 | 30 | p < 0.001 * |
| EC | 17 | 56 | 42 | 31 | 26 | 47 | |||
| FIGO staging | |||||||||
| Early stage | 12 | 48 | p = 0.515 | 36 | 24 | p = 0.360 | 19 | 41 | p = 0.232 |
| Advanced stage | 1 | 12 | 6 | 7 | 7 | 6 | |||
| Pathological Type | |||||||||
| EEC | 13 | 49 | p = 0.411 | 37 | 25 | p = 1.00 | 20 | 42 | p = 0.280 |
| NEEC | 1 | 10 | 6 | 5 | 6 | 5 | |||
| Myometrial Invasion | |||||||||
| <1/2 | 13 | 40 | p = 0.036 * | 35 | 18 | p = 0.017 * | 13 | 40 | p = 0.001 * |
| ≥1/2 | 0 | 20 | 7 | 13 | 13 | 7 | |||
| Lymph Node Metastasis | |||||||||
| NO | 12 | 54 | p = 0.392 | 38 | 26 | p = 0.625 | 21 | 45 | p = 0.096 |
| YES | 2 | 5 | 4 | 5 | 5 | 2 | |||
| Controls | EC | p Value | |
|---|---|---|---|
| TG | 0.789 ± 0.144 | 1.434 ± 0.095 | p = 0.011 * |
| CHOL | 4.569 ± 0.146 | 4.715 ± 0.132 | p = 0.523 |
| APO-A | 1.332 ± 0.039 | 1.211 ± 0.028 | p = 0.030 * |
| APO-B | 0.813 ± 0.037 | 0.937 ± 0.033 | p = 0.001 * |
| HDL | 1.584 ± 0.071 | 1.346 ± 0.034 | p = 0.001 * |
| LDL | 2.522 ± 0.137 | 2.915 ± 0.101 | p = 0.032 * |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Tang, S.; Ma, J.; Su, P.; Lei, H.; Tong, Y.; Cai, L.; Xu, S.; Mao, X.; Sun, P. ERRα Up-Regulates Invadopodia Formation by Targeting HMGCS1 to Promote Endometrial Cancer Invasion and Metastasis. Int. J. Mol. Sci. 2023, 24, 4010. https://doi.org/10.3390/ijms24044010
Tang S, Ma J, Su P, Lei H, Tong Y, Cai L, Xu S, Mao X, Sun P. ERRα Up-Regulates Invadopodia Formation by Targeting HMGCS1 to Promote Endometrial Cancer Invasion and Metastasis. International Journal of Molecular Sciences. 2023; 24(4):4010. https://doi.org/10.3390/ijms24044010
Chicago/Turabian StyleTang, Shuting, Jincheng Ma, Pingping Su, Huifang Lei, Yao Tong, Liangzhi Cai, Shuxia Xu, Xiaodan Mao, and Pengming Sun. 2023. "ERRα Up-Regulates Invadopodia Formation by Targeting HMGCS1 to Promote Endometrial Cancer Invasion and Metastasis" International Journal of Molecular Sciences 24, no. 4: 4010. https://doi.org/10.3390/ijms24044010