
Antibacterial Activity of Juglone Revealed in a Wound Model of Staphylococcus aureus Infection
Abstract
:1. Introduction
2. Results
2.1. Minimum Inhibitory Concentration (MIC) and Minimum Bactericidal Concentration (MBC) of Juglone against S. aureus
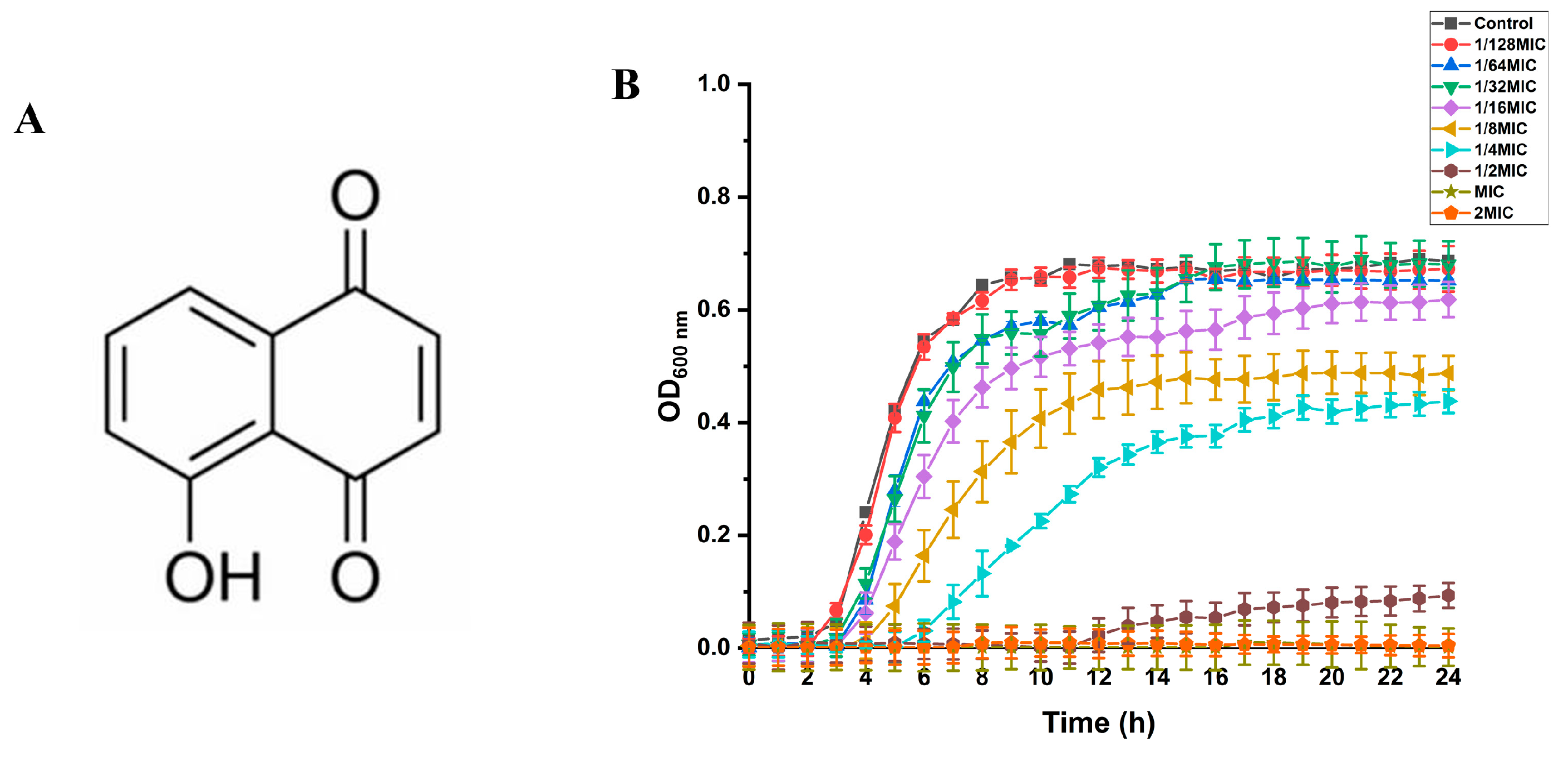
2.2. Growth Curve of S. aureus and SIC of Juglone on S. aureus
2.3. The Effect of Juglone on the Membrane Potential of S. aureus
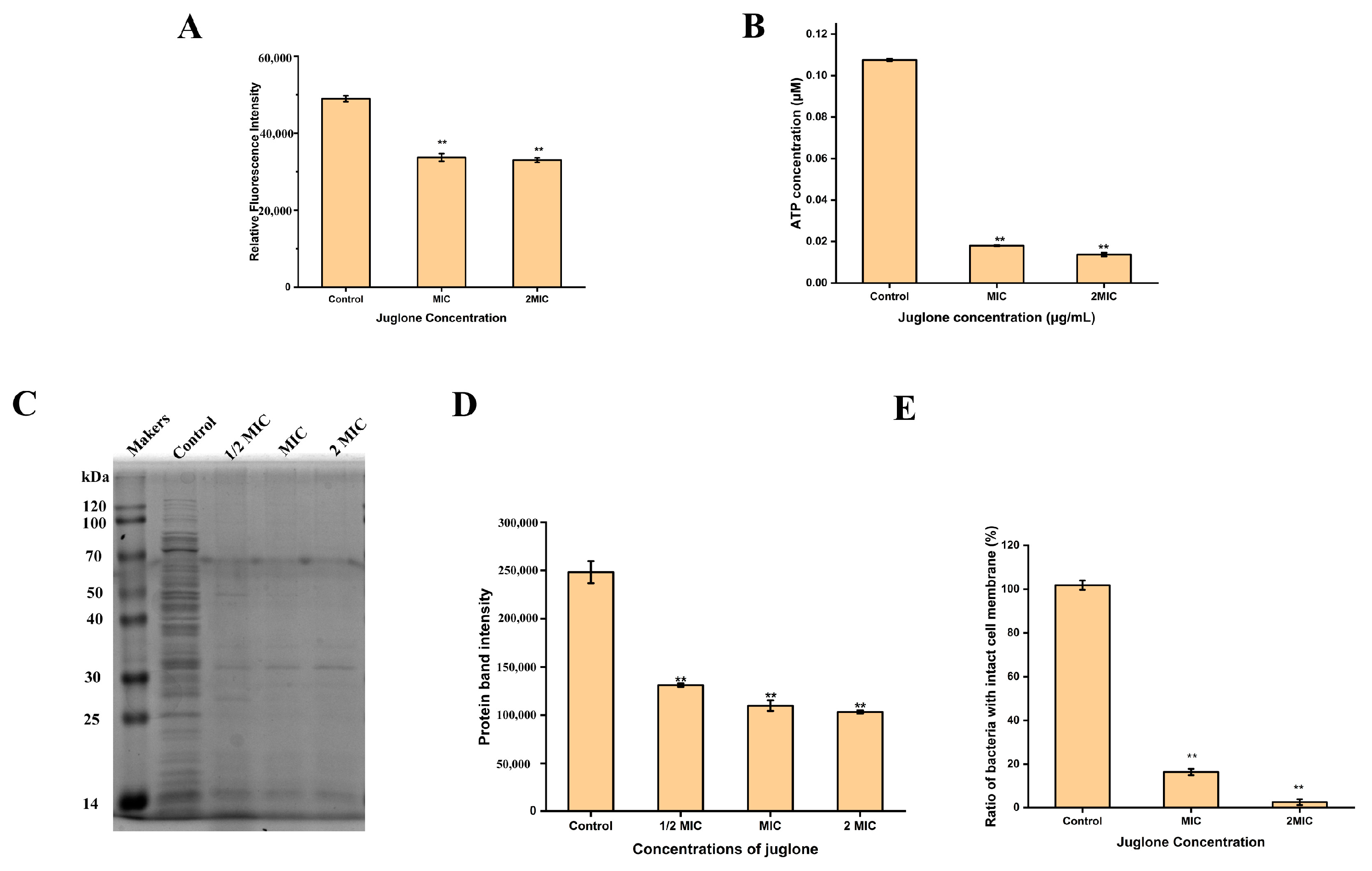
2.4. The Effect of Juglone on the Intracellular ATP Concentration of S. aureus
2.5. The Effect of Juglone on Total Protein Expression in S. aureus
2.6. The Effect of Juglone on the Cell Membrane Integrity of S. aureus
2.7. The Effect of Juglone on the Cell Morphology of S. aureus
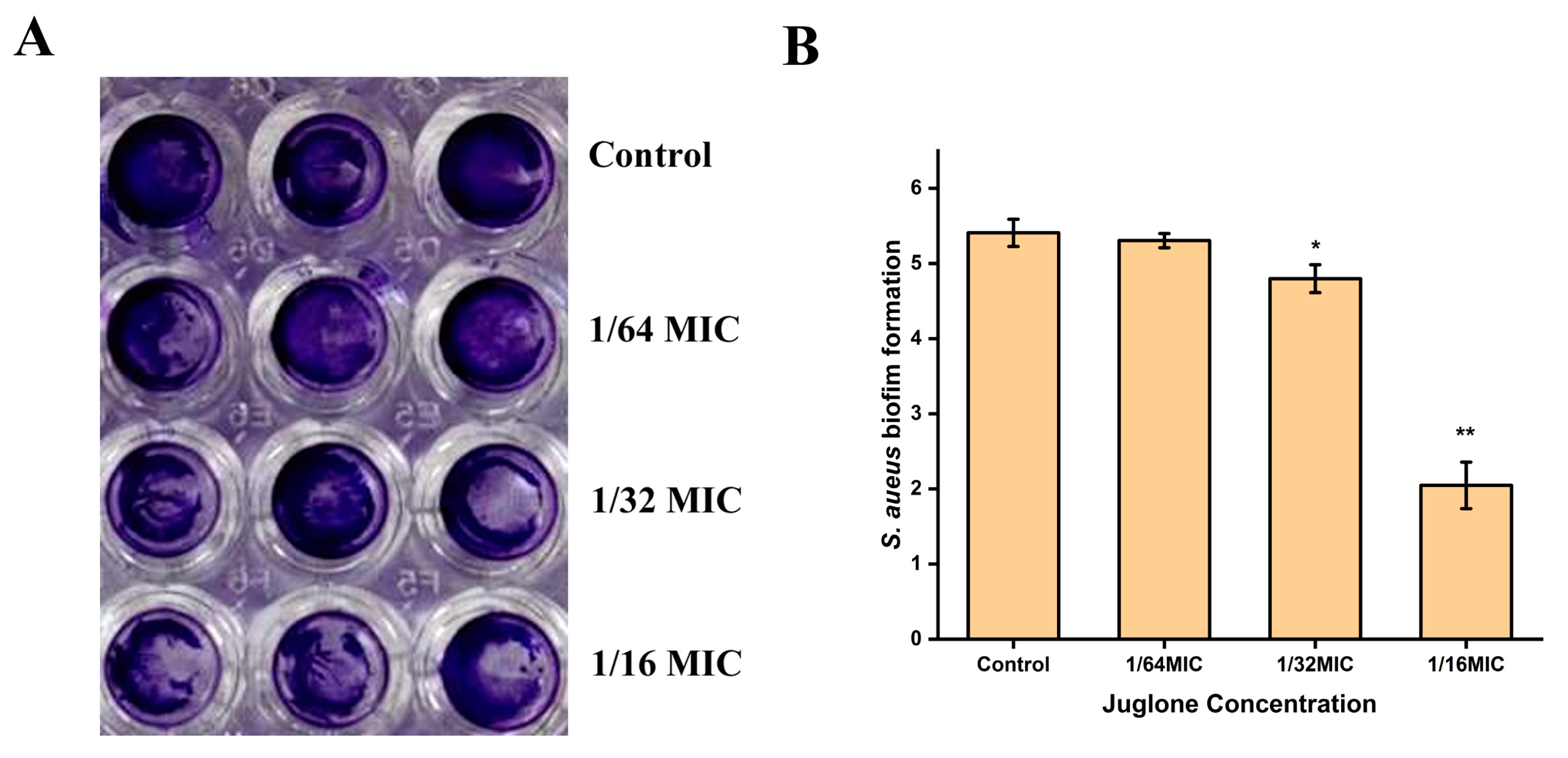
2.8. Inhibition of S. aureus Biofilm Formation by Juglone
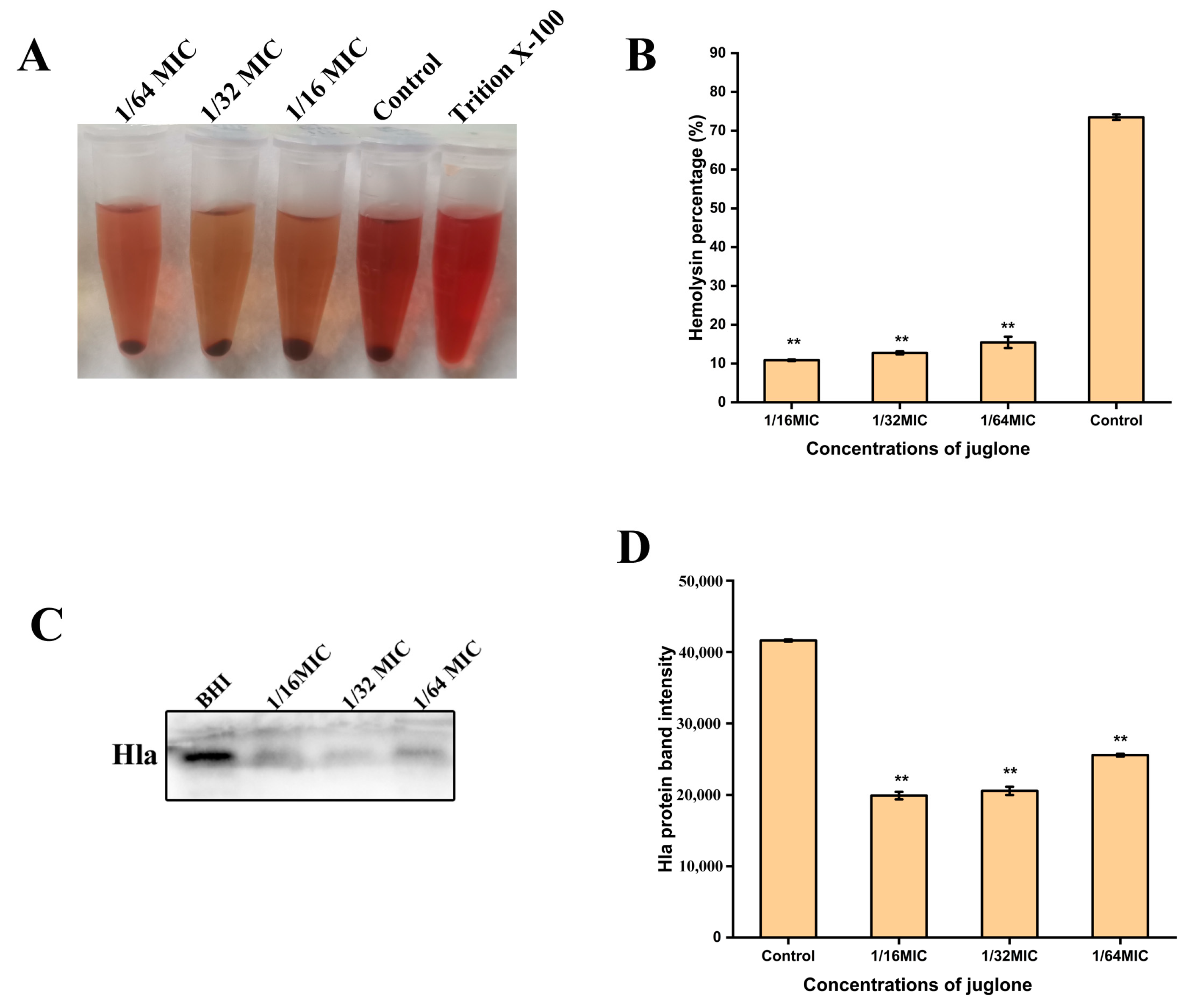
2.9. Inhibition of Hemolytic Activity of S. aureus and Expression of Toxins (Hla) by Juglone
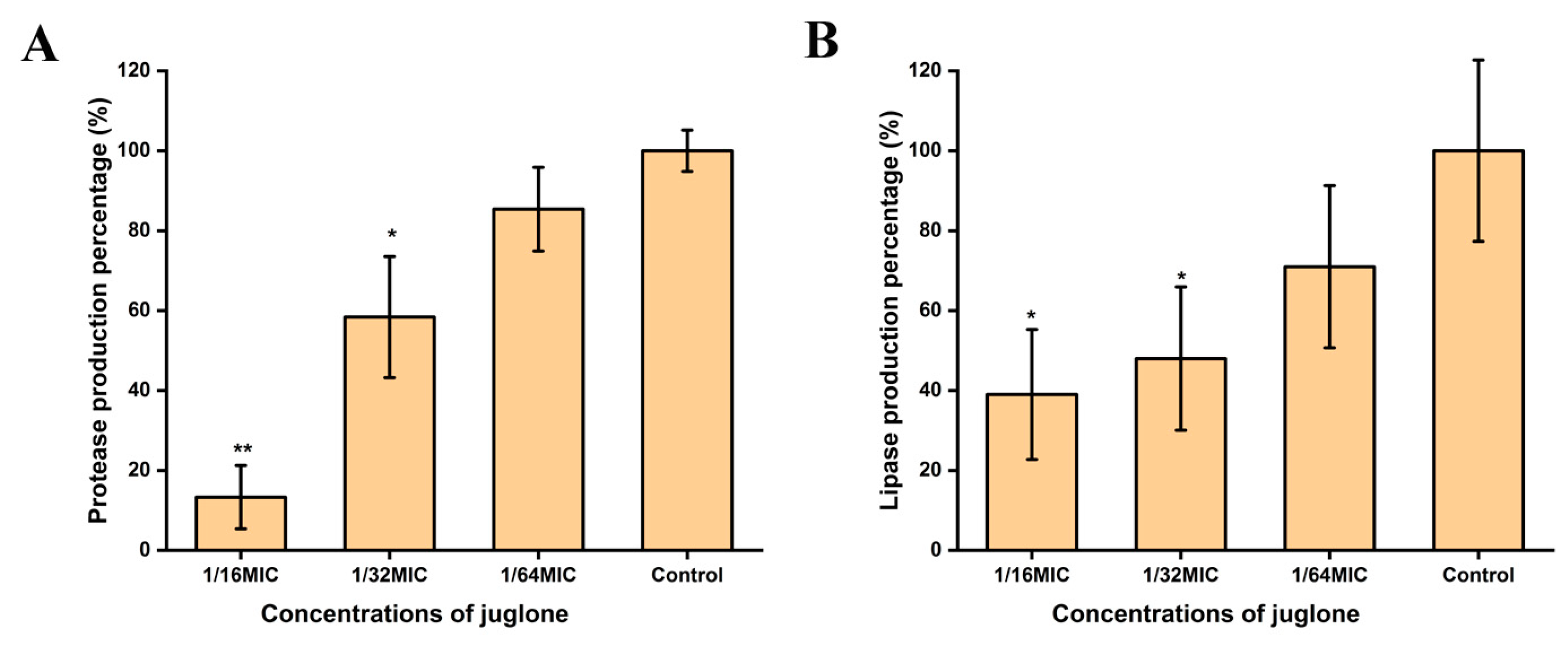
2.10. Juglone Inhibited the Expression of Protease and Lipase of S. aureus
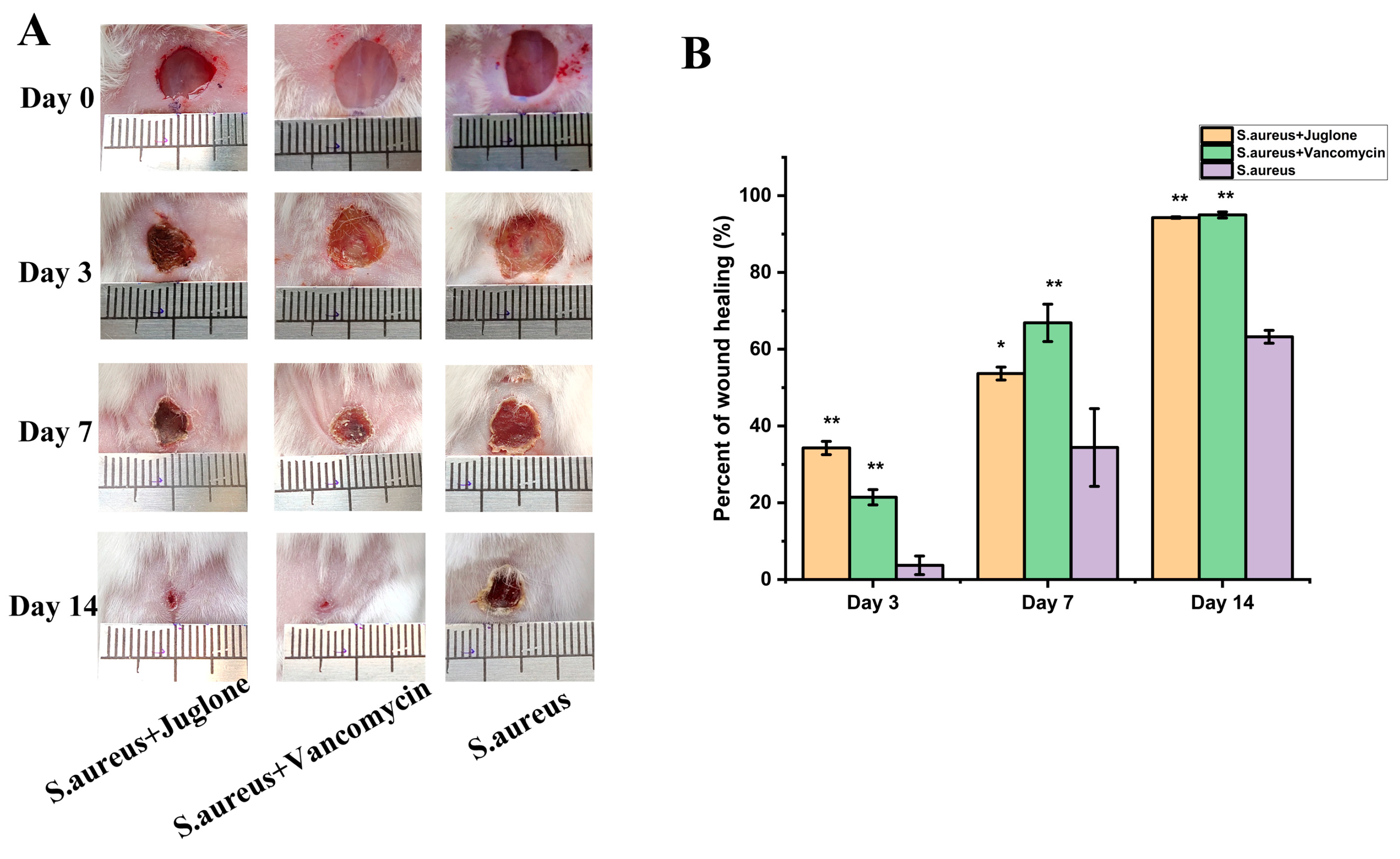
2.11. Topical Juglone Treatment Accelerates the Shrinkage of Wounds Infected with S. aureus
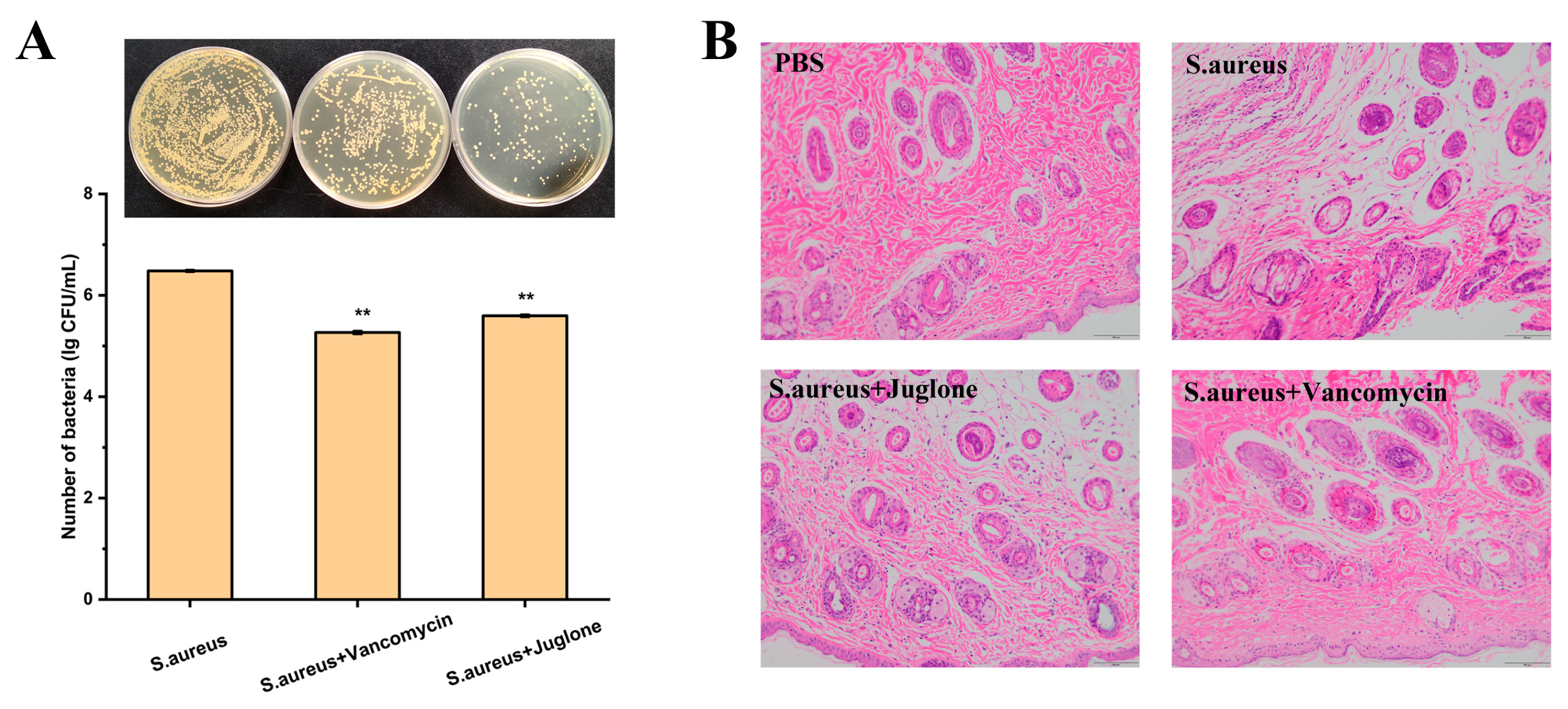
2.12. Topical Treatment with Juglone Reduces Bacterial Burden in S. aureus-Infected Wounds and Improves Histological Evaluation
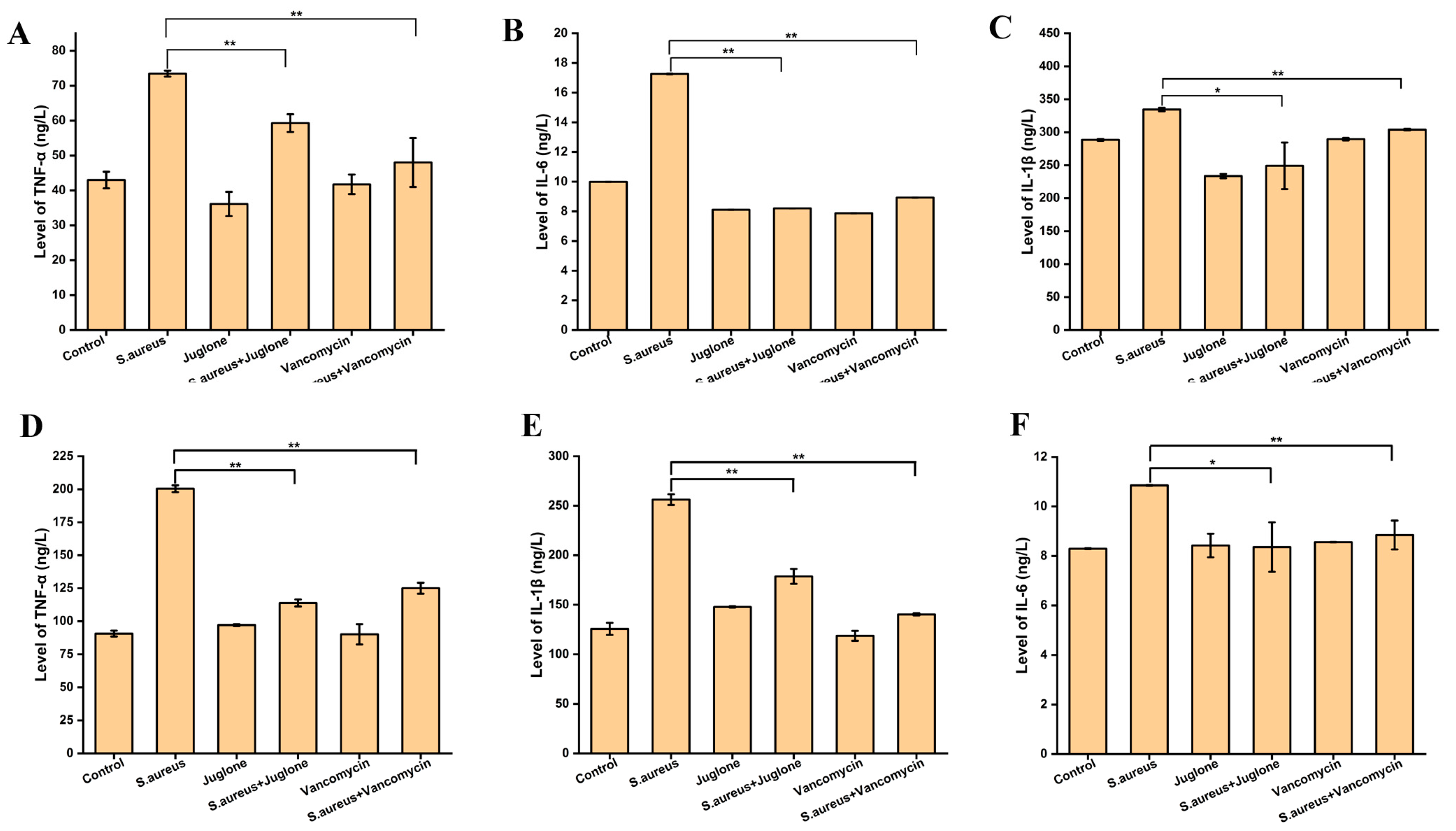
2.13. Topical Treatment of Juglone Modulated the Levels of Inflammatory Factors of Mice e with S. aureus-Infected Wounds
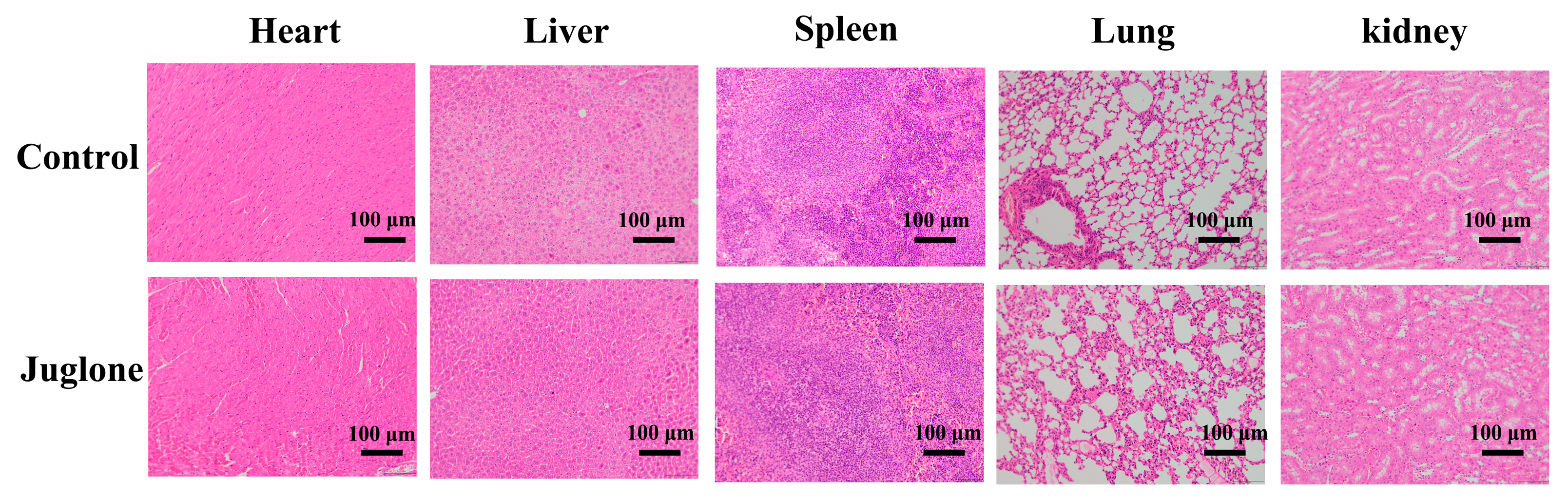
2.14. Biosafety Evaluation of Juglone
3. Discussion
4. Materials and Methods
4.1. Strains and Culture
4.2. MIC and MBC
4.3. Growth Curve and Subinhibitory Concentration (SIC)
4.4. Cell Membrane Potential
4.5. Intracellular ATP Concentration
4.6. Intracellular Proteins
4.7. Cell Membrane Integrity
4.8. Confocal Laser Scanning Microscopy (CLSM)
4.9. Field Emission Scanning Electron Microscopy (FE-SEM)
4.10. S. aureus Biofilm Formation
4.11. Hemolytic Activity
4.12. Hla Expression
4.13. Protease Production Assay
4.14. Lipase Production Assay
4.15. Mouse Wound Model and S. aureus Infection
4.16. Biocompatibility Analysis of Juglone
4.17. Macroscopic Assessment of Lesions
4.18. Histological Evaluation
4.19. Bacterial Loads of Wound Tissue
4.20. Cytokines in Wound Tissue
4.21. Statistical Analysis
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Leroy, S.; Lebert, I.; Andant, C.; Talon, R. Interaction in dual species biofilms between Staphylococcus xylosus and Staphylococcus aureus—ScienceDirect. Int. J. Food Microbiol. 2020, 326, 108653. [Google Scholar] [CrossRef] [PubMed]
- Guest, J.F.; Ayoub, N.; Mcilwraith, T.; Uchegbu, I.; Gerrish, A.; Weidlich, D.; Vowden, K.; Vowden, P. Health economic burden that different wound types impose on the UK’s National Health Service. Int. Wound J. 2017, 14, 322–330. [Google Scholar] [CrossRef] [PubMed]
- Foster, T.J. Antibiotic resistance in Staphylococcus aureus. Current status and future prospects. FEMS Microbiol. Rev. 2017, 41, 430–449. [Google Scholar] [CrossRef]
- Idrees, M.; Sawant, S.; Karodia, N.; Rahman, A. Staphylococcus aureus Biofilm: Morphology, Genetics, Pathogenesis and Treatment Strategies. Int. J. Environ. Res. Public Health 2021, 18, 7602. [Google Scholar] [CrossRef] [PubMed]
- Wang, T.; Zhang, P.; Lv, H.; Deng, X.; Wang, J. A Natural Dietary Flavone Myricetin as an α-Hemolysin Inhibitor for Controlling Staphylococcus aureus Infection. Front. Cell. Infect. Microbiol. 2020, 10, 330. [Google Scholar] [CrossRef]
- Biswas, P.P.; Dey, S.; Sen, A.; Adhikari, L. Virulence markers of vancomycin resistant enterococci isolated from infected and colonized patients. J. Glob. Infect. Dis. 2014, 6, 157–163. [Google Scholar] [CrossRef]
- Seetha, A.; Devaraj, H.; Sudhandiran, G. Indomethacin and juglone inhibit inflammatory molecules to induce apoptosis in colon cancer cells. J. Biochem. Mol. Toxicol. 2020, 34, e22433. [Google Scholar] [CrossRef]
- Wang, J.; Cheng, Y.; Wu, R.; Jiang, D.; Bing, B.; Tan, D.; Yan, T.; Sun, X.; Zhang, Q.; Wu, Z. Antibacterial Activity of Juglone against Staphylococcus aureus: From Apparent to Proteomic. Int. J. Mol. Sci. 2016, 17, 965. [Google Scholar] [CrossRef] [Green Version]
- Montenegro, R.C.; Araújo, A.; Molina, M.T.; Filho, J.; Rocha, D.D.; Lopéz-Montero, E.; Goulart, M.; Bento, E.S.; Alves, A.; Pessoa, C. Cytotoxic activity of naphthoquinones with special emphasis on juglone and its 5-O-methyl derivative. Chem.-Biol. Interact. 2010, 184, 439–448. [Google Scholar] [CrossRef]
- Bonjar, G.; Aghighi, S.; Nik, A.K. Antibacterial and Antifungal Survey in Plants used in Indigenous Herbal-Medicine of South East Regions of Iran. J. Biol. Sci. 2004, 4, 405–412. [Google Scholar]
- Bai, J.R.; Zhong, K.; Wu, Y.P.; Elena, G.; Gao, H. Antibiofilm activity of shikimic acid against Staphylococcus aureus—ScienceDirect. Food Control 2019, 95, 327–333. [Google Scholar] [CrossRef]
- Hindy, J.R.; Haddad, S.F.; Kanj, S.S. New drugs for methicillin-resistant Staphylococcus aureus skin and soft tissue infections. Curr. Opin. Infect. Dis. 2022, 35, 112–119. [Google Scholar] [CrossRef] [PubMed]
- Babula, P.; Adam, V.; Havel, L.; Kizek, R. Noteworthy Secondary Metabolites Naphthoquinones-their Occurrence, Pharmacological Properties and Analysis. Curr. Pharm. Anal. 2009, 5, 47–68. [Google Scholar] [CrossRef]
- Envelope, W.; Cwa, B.; Jya, B. Decolorizing shellac incorporated with natural antibacterial juglone from walnut green husk extract for preserving the postharvest quality of Wichita pecans (Carya illinoinensis [Wangenh.] K. Koch) during storage. Sci. Hortic. 2022, 304, 111313. [Google Scholar]
- Strugstad, M.P.; Despotovski, S. A summary of extraction, properties, and potential uses of juglone: A literature review. J. Ecosyst. Manag. 2012, 13, 72–86. [Google Scholar] [CrossRef]
- Duvauchelle, V.; Majdi, C.; Bénimélis, D.; Dunyach-Remy, C.; Meffre, P.; Benfodda, Z. Synthesis, Structure Elucidation, Antibacterial Activities, and Synergistic Effects of Novel Juglone and Naphthazarin Derivatives Against Clinical Methicillin-Resistant Staphylococcus aureus Strains. Front. Chem. 2021, 863. [Google Scholar] [CrossRef]
- Wan, Y.; Wang, X.; Zhang, P.; Zhang, M.; Kou, M.; Shi, C.; Peng, X.; Wang, X. Control of Foodborne Staphylococcus aureus by Shikonin, a Natural Extract. Foods 2021, 10, 2954. [Google Scholar] [CrossRef]
- Dissanayake, D.M.I.H.; Perera, D.D.B.D.; Keerthirathna, L.R.; Heendeniya, S.; Peiris, L.D.C. Antimicrobial activity of Plumbago indica and ligand screening of plumbagin against methicillin-resistant Staphylococcus aureus. J. Biomol. Struct. Dyn. 2022, 40, 3273–3284. [Google Scholar] [CrossRef]
- Strahl, H.; Errington, J. Bacterial Membranes: Structure, Domains, and Function. Annu. Rev. Microbiol. 2017, 71, 519–538. [Google Scholar] [CrossRef]
- Zw, A.; Qy, A.; Xw, A.; Rl, A.; Hq, A.; Pm, A.; Qi, S.B.; Hz, A. Antibacterial activity of xanthan-oligosaccharide against Staphylococcus aureus via targeting biofilm and cell membrane. Int. J. Biol. Macromol. 2020, 153, 539–544. [Google Scholar]
- Khemakhem, I.; Abdelhedi, O.; Trigui, I.; Ayadi, M.A.; Bouaziz, M. Structural, antioxidant and antibacterial activities of polysaccharides extracted from olive leaves. Int. J. Biol. Macromol. Struct. Funct. Interact. 2018, 106, 425–432. [Google Scholar] [CrossRef] [PubMed]
- Luther, A.; Urfer, M.; Zahn, M.; Muller, M.; Wang, S.Y.; Mondal, M.; Vitale, A.; Hartmann, J.B.; Sharpe, T.; Monte, F.L.; et al. Chimeric peptidomimetic antibiotics against Gram-negative bacteria. Nature 2019, 576, 452–458. [Google Scholar] [CrossRef] [PubMed]
- Shu, Q.; Niu, Y.; Zhao, W.; Chen, Q. Antibacterial activity and mannosylerythritol lipids against vegetative cells and spores of Bacillus cereus. Food Control 2019, 106, 106711. [Google Scholar] [CrossRef]
- Zhang, L.L.; Zhang, L.F.; Hu, Q.P.; Hao, D.L.; Xu, J.G. Chemical composition, antibacterial activity of Cyperus rotundus rhizomes essential oil against Staphylococcus aureus via membrane disruption and apoptosis pathway. Food Control 2017, 80, 290–296. [Google Scholar] [CrossRef]
- Lu, C.H.; Shiau, C.W.; Chang, Y.C.; Kung, H.N.; Chiu, H.C. SC5005 dissipates the membrane potential to kill Staphylococcus aureus persisters without detectable resistance. J. Antimicrob. Chemother. 2021, 76, 2049–2056. [Google Scholar] [CrossRef] [PubMed]
- Kramer, B.; Thielmann, J. Monitoring the live to dead transition of bacteria during thermal stress by a multi-method approach. J. Microbiol. Methods 2016, 123, 24–30. [Google Scholar] [CrossRef]
- Wang, L.; Hu, W.; Deng, J.; Liu, X.; Zhou, J.; Li, X. Antibacterial activity of Litsea cubeba essential oil and its mechanism against Botrytis cinerea. RSC Adv. 2019, 9, 28987–28995. [Google Scholar] [CrossRef] [Green Version]
- Meira, N.; Holley, R.A.; Bordin, K.; Macedo, R.; Luciano, F.B. Combination of essential oil compounds and phenolic acids against Escherichia coli O157:H7 in vitro and in dry-fermented sausage production. Int. J. Food Microbiol. 2017, 260, 59–64. [Google Scholar] [CrossRef]
- Vestergaard, M.; Roshanak, S.; Ingmer, H. Targeting the ATP Synthase in Staphylococcus aureus Small Colony Variants, Streptococcus pyogenes and Pathogenic Fungi. Antibiotics 2021, 10, 376. [Google Scholar] [CrossRef]
- Song, Y.J.; Yu, H.H.; Kim, Y.J.; Lee, N.K.; Paik, H.D. Anti-Biofilm Activity of Grapefruit Seed Extract against Staphylococcus aureus and Escherichia coli. J. Microbiol. Biotechnol. 2019, 29, 1177–1183. [Google Scholar] [CrossRef]
- Chen, J.; Chen, J.; Wang, Z.; Chen, C.; Zheng, J.; Yu, Z.; Deng, Q.; Zhao, Y.; Wen, Z. 20S-ginsenoside Rg3 inhibits the biofilm formation and haemolytic activity of Staphylococcus aureus by inhibiting the SaeR/SaeS two-component system. J. Med. Microbiol. 2022, 71, 001587. [Google Scholar] [CrossRef] [PubMed]
- Yang, Y.; Li, J.; Yin, Y.; Guo, D.; Shi, C. Antibiofilm activity of coenzyme Q0 against Salmonella Typhimurium and its effect on adhesion–invasion and survival–replication. Appl. Microbiol. Biotechnol. 2019, 103, 8545–8557. [Google Scholar] [CrossRef] [PubMed]
- Tang, C.; Li, Q.; Lin, T. Lycopene attenuates Staphylococcus aureus -induced inflammation via inhibiting α-hemolysin expression. Microbes Infect. 2021, 23. [Google Scholar] [CrossRef] [PubMed]
- Sirobhushanam, S.; Parsa, N.; Reed, T.J.; Berthier, C.C.; Sarkar, M.K.; Hile, G.A.; Tsoi, L.C.; Banfield, J.; Dobry, C.; Horswill, A.R. Staphylococcus aureus Colonization Is Increased on Lupus Skin Lesions and Is Promoted by IFN-Mediated Barrier Disruption—ScienceDirect. J. Investig. Dermatol. 2020, 140, 1066–1074. [Google Scholar] [CrossRef] [PubMed]
- Suarez Carneiro, M.A.M.; Silva, L.D.S.; Diniz, R.M.; Saminez, W.; Oliveira, P.V.; Pereira Mendonca, J.S.; Colasso, A.H.M.; Soeiro Silva, I.S.; Jandu, J.J.B.; Sa, J.C.; et al. Immunomodulatory and anti-infective effects of Cratylia mollis lectin (Cramoll) in a model of wound infection induced by Staphylococcus aureus. Int. Immunopharmacol. 2021, 100, 108094. [Google Scholar] [CrossRef]
- Ceriotti, F.; Zakowski, J.; Sine, H.; Altaie, S.; Horowitz, G.; Pesce, A.J.; Boyd, J.; Horn, P.; Gard, U.; Horowitz, G. Clinical and Laboratory Standards Institute (CLSI). 2012. Available online: https://www.scienceopen.com/document?vid=1d021ccc-8583-4e6b-a47c-8735734df154 (accessed on 30 December 2022).
- Guo, D.; Wang, S.; Li, J.; Bai, F.; Shi, C. The antimicrobial activity of coenzyme Q0 against planktonic and biofilm forms of Cronobacter sakazakii. Food Microbiol. 2019, 86, 103337. [Google Scholar] [CrossRef]
- Han, Q.; Yan, X.; Zhang, R.; Wang, G.; Zhang, Y. Juglone Inactivates Pseudomonas aeruginosa through Cell Membrane Damage, Biofilm Blockage, and Inhibition of Gene Expression. Molecules 2021, 26, 5854. [Google Scholar] [CrossRef]
- Kang, J.; Liu, L.; Liu, Y.; Wang, X. Ferulic Acid Inactivates Shigella flexneri through Cell Membrane Destructieon, Biofilm Retardation, and Altered Gene Expression. J. Agric. Food Chem. 2020, 68, 7121–7131. [Google Scholar] [CrossRef]
- Shi, C.; Sun, Y.; Liu, Z.; Guo, D.; Sun, H.; Sun, Z.; Chen, S.; Zhang, W.; Wen, Q.; Peng, X. Inhibition of Cronobacter sakazakii Virulence Factors by Citral. Sci. Rep. 2017, 7, 43243. [Google Scholar] [CrossRef] [Green Version]
- Li, J.; Li, S.; Li, H.; Guo, X.; Guo, D.; Yang, Y.; Wang, X.; Zhang, C.; Shan, Z.; Xia, X. Antibiofilm activity of shikonin against Listeria monocytogenes and inhibition of key virulence factors. Food Control 2021, 120, 107558. [Google Scholar] [CrossRef]
- Teng, Z.; Shi, D.; Liu, H.; Shen, Z.; Zha, Y.; Li, W.; Deng, X.; Wang, J. Lysionotin attenuates Staphylococcus aureus pathogenicity by inhibiting α-toxin expression. Appl. Microbiol. Biotechnol. 2017, 101, 6697–6703. [Google Scholar] [CrossRef] [PubMed]
- Wang, R.; Zhang, B.; Liang, Z.; He, Y.; Wang, Z.; Ma, X.; Yao, X.; Sun, J.; Wang, J. Insights into rapid photodynamic inactivation mechanism of Staphylococcus aureus via rational design of multifunctional nitrogen-rich carbon-coated bismuth/cobalt nanoparticles. Appl. Catal. B Environ. 2018, 241, 167–177. [Google Scholar] [CrossRef]












{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Strain | MIC (μg/mL) | MBC (μg/mL) | Original Source of Strain |
|---|---|---|---|
| ATCC29213 (MSSA) | 1000 | 2000 | American type culture collection |
| ATCC43300 (MRSA) | 1000 | 2000 | American type culture collection |
| ST97 (MRSA) | 1000 | 2000 | Cow mastitis wound |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Wan, Y.; Wang, X.; Yang, L.; Li, Q.; Zheng, X.; Bai, T.; Wang, X. Antibacterial Activity of Juglone Revealed in a Wound Model of Staphylococcus aureus Infection. Int. J. Mol. Sci. 2023, 24, 3931. https://doi.org/10.3390/ijms24043931
Wan Y, Wang X, Yang L, Li Q, Zheng X, Bai T, Wang X. Antibacterial Activity of Juglone Revealed in a Wound Model of Staphylococcus aureus Infection. International Journal of Molecular Sciences. 2023; 24(4):3931. https://doi.org/10.3390/ijms24043931
Chicago/Turabian StyleWan, Yangli, Xiaowen Wang, Liu Yang, Qianhong Li, Xuting Zheng, Tianyi Bai, and Xin Wang. 2023. "Antibacterial Activity of Juglone Revealed in a Wound Model of Staphylococcus aureus Infection" International Journal of Molecular Sciences 24, no. 4: 3931. https://doi.org/10.3390/ijms24043931