
Kinesins Modify ERR1-Dependent Transcription Using a Conserved Nuclear Receptor Box Motif
Abstract
:1. Introduction
2. Results
2.1. Identification of Kinesin Family Motor Transcriptional Regulator Candidates
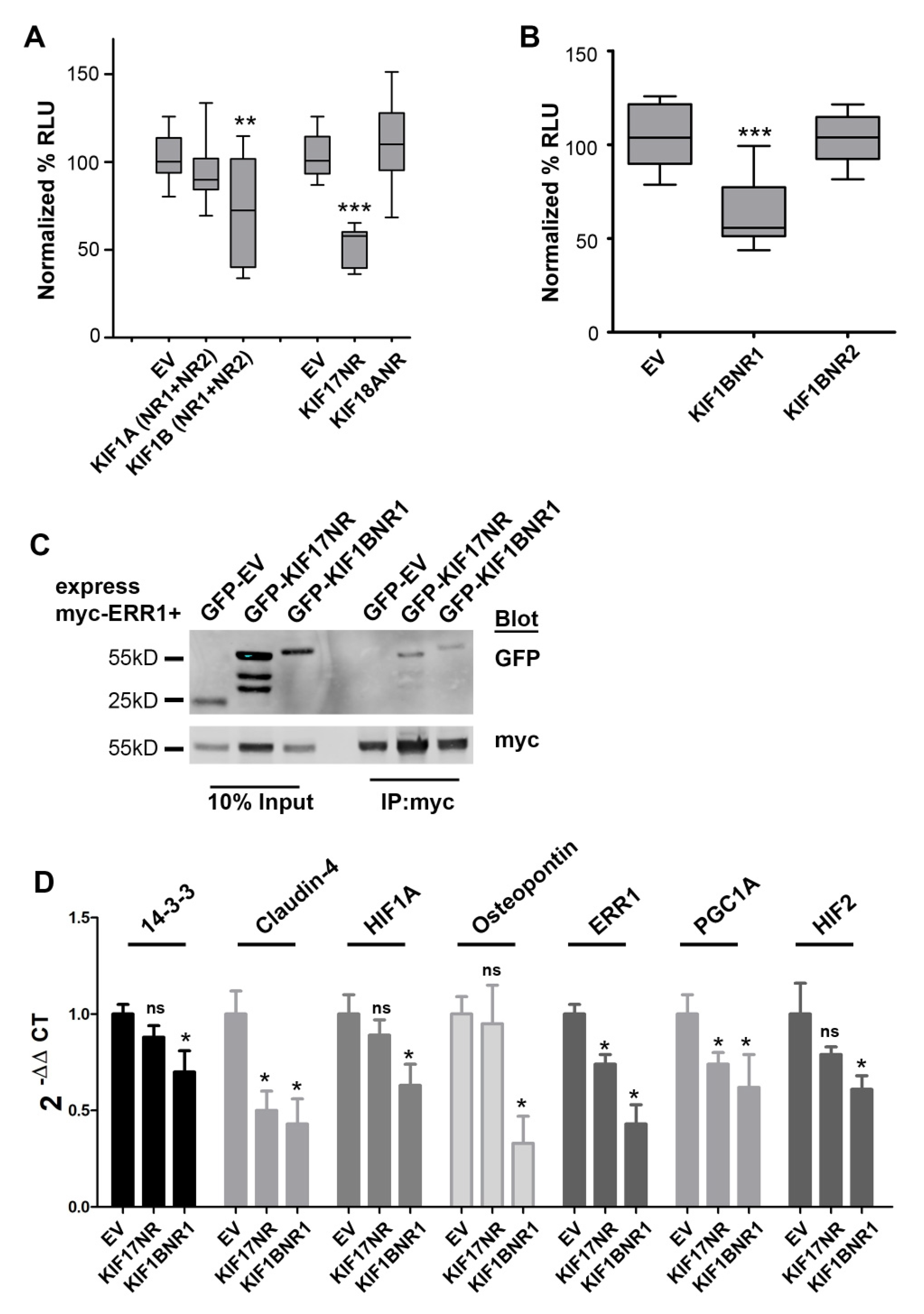
2.2. KIF1B-NR1 Inhibits ERR1-Dependent Transcription
2.3. KIF1B-NR1 Interacts with ERR1
2.4. KIF1B-NR1 Globally Inhibits ERR1 Target Genes
2.5. KIF1B-NR1 Inhibits Nuclear Import of ERR1 by Sequestering ERR1 in the Cytoplasm
3. Discussion
4. Materials and Methods
4.1. Cell Culture, Transfection, and Microinjection
4.2. Expression Constructs
- KIF1A (NR1+NR2): forward, 5′-GAC ATG AAG CAG GAG ATG GAG and reverse, 5′-CAG GAG GTG GAG AAG ACT AGG.
- KIF1B (NR1+NR2): forward, 5′-CGC TCA GGA AAC CGT ATC ATC ATG and reverse, 5′-GTA GAC TCT AGG AGC AAC TCT CTG.
- KIF1B-NR1: forward, 5′-CCT GTG GAC TGG ACA TTT GCC and reverse, 5′-AGA CCA ATA GTG TGT TGC TCC.
- KIF1B-NR2: forward, 5′-GAT CTC TTC AGT GAC GGG and reverse, 5′-CAA GGA CAG ACC AGA ACG.
- KIF18: forward, 5′-GCT TGT CTT CAG GAA CAG CAA CAC AGG and reverse, 5′-GGA CCA GTT CAG CCT ATT CCT TGT TGC.
4.3. Co-Immunoprecipitation and Immunoblotting
4.4. Luciferase Reporter Assays
4.5. Time-Lapse Imaging and Analysis
4.6. qRT-PCR
- Tbp1: forward, 5′–TGT ATC CAC AGT GAA TCT TGG TTG and reverse, 5′–GGT TCG TGG CTC TCT TAT CCT C.
- 14-3-3: forward, 5′–GCATGAAGTCTGTAACTGAGCA and reverse, 5′–GCACCTTCCGTCTTTTGTTC.
- Claudin-4: forward, 5′–CCATATAACTGCTCAACCTGTCC and reverse, 5′–AGATAAAGCCAGTCCTGATGC.
- ERR1: forward, 5′–TCTCCGCTTGGTGATCTCA and reverse, 5′–CTATGGTGTGGCATCCTGTG.
- HIF1A: forward, 5′–CAACCCAGACATATCCACCTC and reverse, 5′–CTCTGATCATCTGACCAAAACTCA.
- HIF2: forward, 5′–CTTTGCGAGCATCCGGTA and reverse, 5′–AGCCTATGAATTCTACCATGCG.
- Osteopontin: forward, 5′–CCCCACAGTAGACACATATGATG and reverse, 5′–TTCAACTCCTCGCTTTCCAT.
- PGC1A: forward, 5′–GTC CTT TTC TCG ACA CAG GT and reverse, 5′–GTC TGT AGT GGC TTG ACT CAT AG.
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Hirokawa, N.; Noda, Y.; Tanaka, Y.; Niwa, S. Kinesin superfamily motor proteins and intracellular transport. Nat. Rev. Mol. Cell. Biol. 2009, 10, 682–696. [Google Scholar] [CrossRef] [PubMed]
- Sisson, J.C.; Ho, K.S.; Suyama, K.; Scott, M.P. Costal2, a novel kinesin-related protein in the Hedgehog signaling pathway. Cell 1997, 90, 235–245. [Google Scholar] [CrossRef] [PubMed]
- Robbins, D.J.; Nybakken, K.E.; Kobayashi, R.; Sisson, J.C.; Bishop, J.M.; Thérond, P.P. Hedgehog elicits signal transduction by means of a large complex containing the kinesin-related protein costal2. Cell 1997, 90, 225–234. [Google Scholar] [CrossRef] [PubMed]
- He, M.; Subramanian, R.; Bangs, F.; Omelchenko, T.; Liem, K.F., Jr.; Kapoor, T.M.; Anderson, K.V. The kinesin-4 protein Kif7 regulates mammalian Hedgehog signalling by organizing the cilium tip compartment. Nat. Cell Biol. 2014, 16, 663–672. [Google Scholar] [CrossRef] [PubMed]
- Marks, S.A.; Kalderon, D. Regulation of mammalian Gli proteins by Costal 2 and PKA in Drosophila reveals Hedgehog pathway conservation. Development 2011, 138, 2533–2542. [Google Scholar] [CrossRef] [PubMed]
- Cheung HO, L.; Zhang, X.; Ribeiro, A.; Mo, R.; Makino, S.; Puviindran, V.; Law, K.K.L.; Briscoe, J.; Hui, C.C. The kinesin protein Kif7 is a critical regulator of Gli transcription factors in mammalian hedgehog signaling. Sci. Signal. 2009, 2, ra29. [Google Scholar] [CrossRef]
- Endoh-Yamagami, S.; Evangelista, M.; Wilson, D.; Wen, X.; Theunissen, J.W.; Phamluong, K.; Davis, M.; Scales, S.J.; Solloway, M.J.; de Sauvage, F.J.; et al. The mammalian Cos2 homolog Kif7 plays an essential role in modulating Hh signal transduction during development. Curr. Biol. 2009, 19, 1320–1326. [Google Scholar] [CrossRef] [PubMed]
- Liem, K.F., Jr.; He, M.; Ocbina PJ, R.; Anderson, K.V. Mouse Kif7/Costal2 is a cilia-associated protein that regulates Sonic hedgehog signaling. Proc. Natl. Acad. Sci. USA 2009, 106, 13377–13382. [Google Scholar] [CrossRef]
- Haque, F.; Freniere, C.; Ye, Q.; Mani, N.; Wilson-Kubalek, E.M.; Ku, P.I.; Milligan, R.A.; Subramanian, R. Cytoskeletal regulation of a transcription factor by DNA mimicry via coiled-coil interactions. Nat. Cell Biol. 2022, 24, 1088–1098. [Google Scholar] [CrossRef]
- Midorikawa, R.; Takei, Y.; Hirokawa, N. KIF4 motor regulates activity-dependent neuronal survival by suppressing PARP-1 enzymatic activity. Cell 2006, 125, 371–383. [Google Scholar] [CrossRef] [Green Version]
- Li, J.; Jiang, J.; Qian, Q.; Xu, Y.; Zhang, C.; Xiao, J.; Du, C.; Luo, W.; Zou, G.; Chen, M.; et al. Mutation of rice BC12/GDD1, which encodes a kinesin-like protein that binds to a GA biosynthesis gene promoter, leads to dwarfism with impaired cell elongation. Plant. Cell 2011, 23, 628–640. [Google Scholar] [CrossRef]
- Macho, B.; Brancorsini, S.; Fimia, G.M.; Setou, M.; Hirokawa, N.; Sassone-Corsi, P. CREM-dependent transcription in male germ cells controlled by a kinesin. Science 2002, 298, 2388–2390. [Google Scholar] [CrossRef]
- Kotaja, N.; Macho, B.; Sassone-Corsi, P. Microtubule-independent and protein kinase A-mediated function of kinesin KIF17b controls the intracellular transport of activator of CREM in testis (ACT). J. Biol. Chem. 2005, 280, 31739–31745. [Google Scholar] [CrossRef] [PubMed]
- Seneviratne, A.P.B.; Turan, Z.; Hermant, A.; Lecine, P.; Smith, W.O.; Borg, J.P.; Jaulin, F.; Kreitzer, G. Modulation of estrogen related receptor alpha activity by the kinesin KIF17. Oncotarget 2017, 8, 50359–50375. [Google Scholar] [CrossRef] [PubMed]
- Bevan, C.; Parker, M. The role of coactivators in steroid hormone action. Exp. Cell Res. 1999, 253, 349–356. [Google Scholar] [CrossRef]
- Plevin, M.J.; Mills, M.M.; Ikura, M. The LxxLL motif: A multifunctional binding sequence in transcriptional regulation. Trends Biochem. Sci. 2005, 30, 66–69. [Google Scholar] [CrossRef] [PubMed]
- Alaynick, W.A. Nuclear receptors, mitochondria and lipid metabolism. Mitochondrion 2008, 8, 329–337. [Google Scholar] [CrossRef]
- Audet-Walsh, E.; Giguere, V. The multiple universes of estrogen-related receptor alpha and gamma in metabolic control and related diseases. Acta Pharmacol. Sin. 2015, 36, 51–61. [Google Scholar] [CrossRef]
- Kim, S.Y.; Yang, C.S.; Lee, H.M.; Kim, J.K.; Kim, Y.S.; Kim, Y.R.; Kim, J.S.; Kim, T.S.; Yuk, J.M.; Dufour, C.R.; et al. ESRRA (estrogen-related receptor alpha) is a key coordinator of transcriptional and post-translational activation of autophagy to promote innate host defense. Autophagy 2018, 14, 152–168. [Google Scholar] [CrossRef]
- Kim, S.; Lee, J.Y.; Shin, S.G.; Kim, J.K.; Silwal, P.; Kim, Y.J.; Shin, N.R.; Kim, P.S.; Won, M.; Lee, S.H.; et al. ESRRA (estrogen related receptor alpha) is a critical regulator of intestinal homeostasis through activation of autophagic flux via gut microbiota. Autophagy 2021, 17, 2856–2875. [Google Scholar] [CrossRef]
- Tripathi, M.; Yen, P.M.; Singh, B.K. Estrogen-Related Receptor Alpha: An Under-Appreciated Potential Target for the Treatment of Metabolic Diseases. Int. J. Mol. Sci. 2020, 21, 1645. [Google Scholar] [CrossRef] [PubMed]
- Gaillard, S.; Dwyer, M.A.; McDonnell, D.P. Definition of the molecular basis for estrogen receptor-related receptor-alpha-cofactor interactions. Mol. Endocrinol. 2007, 21, 62–76. [Google Scholar] [CrossRef] [PubMed]
- Lu, N.; Wang, W.; Liu, J.; Wong, C.W. Protein kinase C epsilon affects mitochondrial function through estrogen-related receptor alpha. Cell. Signal. 2011, 23, 1473–1478. [Google Scholar] [CrossRef] [PubMed]
- Huss, J.M.; Torra, I.P.; Staels, B.; Giguere, V.; Kelly, D.P. Estrogen-related receptor alpha directs peroxisome proliferator-activated receptor alpha signaling in the transcriptional control of energy metabolism in cardiac and skeletal muscle. Mol. Cell. Biol. 2004, 24, 9079–9091. [Google Scholar] [CrossRef]
- Schreiber, S.N.; Knutti, D.; Brogli, K.; Uhlmann, T.; Kralli, A. The transcriptional coactivator PGC-1 regulates the expression and activity of the orphan nuclear receptor estrogen-related receptor alpha (ERRalpha). J. Biol. Chem. 2003, 278, 9013–9018. [Google Scholar] [CrossRef]
- Kamei, Y.; Ohizumi, H.; Fujitani, Y.; Nemoto, T.; Tanaka, T.; Takahashi, N.; Kawada, T.; Miyoshi, M.; Ezaki, O.; Kakizuka, A. PPARgamma coactivator 1beta/ERR ligand 1 is an ERR protein ligand, whose expression induces a high-energy expenditure and antagonizes obesity. Proc. Natl. Acad. Sci. USA 2003, 100, 12378–12383. [Google Scholar] [CrossRef]
- Giguere, V.; Dufour, C.R.; Eichner, L.J.; Deblois, G.; Cermakian, N. Estrogen-related receptor alpha, the molecular clock, and transcriptional control of metabolic outputs. Cold Spring Harb. Symp. Quant. Biol. 2011, 76, 57–61. [Google Scholar] [CrossRef]
- Wei, W.; Schwaid, A.G.; Wang, X.; Wang, X.; Chen, S.; Chu, Q.; Saghatelian, A.; Wan, Y. Ligand Activation of ERRalpha by Cholesterol Mediates Statin and Bisphosphonate Effects. Cell Metab. 2016, 23, 479–491. [Google Scholar] [CrossRef]
- Casaburi, I.; Chimento, A.; De Luca, A.; Nocito, M.; Sculco, S.; Avena, P.; Trotta, F.; Rago, V.; Sirianni, R.; Pezzi, V. Cholesterol as an Endogenous ERRalpha Agonist: A New Perspective to Cancer Treatment. Front. Endocrinol. 2018, 9, 525. [Google Scholar] [CrossRef]
- Ranhotra, H.S. Estrogen-related receptor alpha in select host functions and cancer: New frontiers. Mol. Cell. Biochem. 2022, 477, 1349–1359. [Google Scholar] [CrossRef]
- Deblois, G.; St-Pierre, J.; Giguere, V. The PGC-1/ERR signaling axis in cancer. Oncogene 2013, 32, 3483–3490. [Google Scholar] [CrossRef] [PubMed]
- Ranhotra, H.S. Estrogen-related receptor alpha and cancer: Axis of evil. J. Recept. Signal Transduct. Res. 2015, 35, 505–508. [Google Scholar] [CrossRef] [PubMed]
- Kosugi, S.; Hasebe, M.; Tomita, M.; Yanagawa, H. Systematic identification of cell cycle-dependent yeast nucleocytoplasmic shuttling proteins by prediction of composite motifs. Proc. Natl. Acad. Sci. USA 2009, 106, 10171–10176. [Google Scholar] [CrossRef] [PubMed]
- Kosugi, S.; Hasebe, M.; Matsumura, N.; Takashima, H.; Miyamoto-Sato, E.; Tomita, M.; Yanagawa, H. Six classes of nuclear localization signals specific to different binding grooves of importin alpha. J. Biol. Chem. 2009, 284, 478–485. [Google Scholar] [CrossRef] [PubMed]
- Lu, J.; Wu, T.; Zhang, B.; Liu, S.; Song, W.; Qiao, J.; Ruan, H. Types of nuclear localization signals and mechanisms of protein import into the nucleus. Cell Commun. Signal. 2021, 19, 60. [Google Scholar] [CrossRef]
- Yang, N.; Shigeta, H.; Shi, H.; Teng, C.T. Estrogen-related receptor, hERR1, modulates estrogen receptor-mediated response of human lactoferrin gene promoter. J. Biol. Chem. 1996, 271, 5795–5804. [Google Scholar] [CrossRef]
- Parikh, I.; Rajendran, K.G.; Su, J.L.; Lopez, T.; Sar, M. Are estrogen receptors cytoplasmic or nuclear? Some immunocytochemical and biochemical studies. J. Steroid Biochem. 1987, 27, 185–192. [Google Scholar] [CrossRef]
- Lee, S.J.; Chae, C.; Wang, M.M. p150/glued modifies nuclear estrogen receptor function. Mol. Endocrinol. 2009, 23, 620–629. [Google Scholar] [CrossRef]
- Rayala, S.K.; den Hollander, P.; Balasenthil, S.; Yang, Z.; Broaddus, R.R.; Kumar, R. Functional regulation of oestrogen receptor pathway by the dynein light chain 1. EMBO Rep. 2005, 6, 538–544. [Google Scholar] [CrossRef]
- Hirokawa, N.; Takemura, R. Kinesin superfamily proteins and their various functions and dynamics. Exp. Cell Res. 2004, 301, 50–59. [Google Scholar] [CrossRef]
- Kapitein, L.C.; Peterman, E.J.; Kwok, B.H.; Kim, J.H.; Kapoor, T.M.; Schmidt, C.F. The bipolar mitotic kinesin Eg5 moves on both microtubules that it crosslinks. Nature 2005, 435, 114–118. [Google Scholar] [CrossRef] [PubMed]
- Fozzatti, L.; Lu, C.; Kim, D.W.; Cheng, S.Y. Differential recruitment of nuclear coregulators directs the isoform-dependent action of mutant thyroid hormone receptors. Mol. Endocrinol. 2011, 25, 908–921. [Google Scholar] [CrossRef] [PubMed]
- Darimont, B.D.; Wagner, R.L.; Apriletti, J.W.; Stallcup, M.R.; Kushner, P.J.; Baxter, J.D.; Fletterick, R.J.; Yamamoto, K.R. Structure and specificity of nuclear receptor-coactivator interactions. Genes Dev. 1998, 12, 3343–3356. [Google Scholar] [CrossRef]
- Ko, L.; Cardona, G.R.; Iwasaki, T.; Bramlett, K.S.; Burris, T.P.; Chin, W.W. Ser-884 adjacent to the LXXLL motif of coactivator TRBP defines selectivity for ERs and TRs. Mol. Endocrinol. 2002, 16, 128–140. [Google Scholar] [CrossRef] [PubMed]
- Heery, D.M.; Hoare, S.; Hussain, S.; Parker, M.G.; Sheppard, H. Core LXXLL motif sequences in CREB-binding protein, SRC1, and RIP140 define affinity and selectivity for steroid and retinoid receptors. J. Biol. Chem. 2001, 276, 6695–6702. [Google Scholar] [CrossRef] [PubMed]
- Chang, C.; Norris, J.D.; Gron, H.; Paige, L.A.; Hamilton, P.T.; Kenan, D.J.; Fowlkes, D.; McDonnell, D.P. Dissection of the LXXLL nuclear receptor-coactivator interaction motif using combinatorial peptide libraries: Discovery of peptide antagonists of estrogen receptors alpha and beta. Mol. Cell. Biol. 1999, 19, 8226–8239. [Google Scholar] [CrossRef]
- Ren, Y.; Behre, E.; Ren, Z.; Zhang, J.; Wang, Q.; Fondell, J.D. Specific structural motifs determine TRAP220 interactions with nuclear hormone receptors. Mol. Cell. Biol. 2000, 20, 5433–5446. [Google Scholar] [CrossRef] [PubMed]
- Jumper, J.; Evans, R.; Pritzel, A.; Green, T.; Figurnov, M.; Ronneberger, O.; Tunyasuvunakool, K.; Bates, R.; Zidek, A.; Potapenko, A.; et al. Highly accurate protein structure prediction with AlphaFold. Nature 2021, 596, 583–589. [Google Scholar] [CrossRef]
- Zusev, M.; Benayahu, D. The Regulation of MS-KIF18A Expression and Cross Talk with Estrogen Receptor. PLoS ONE 2009, 4, e6407. [Google Scholar] [CrossRef]
- Vanacker, J.M.; Pettersson, K.; Gustafsson, J.A.; Laudet, V. Transcriptional targets shared by estrogen receptor- related receptors (ERRs) and estrogen receptor (ER) alpha, but not by ERbeta. EMBO J. 1999, 18, 4270–4279. [Google Scholar] [CrossRef] [Green Version]
- Kraus, R.J.; Ariazi, E.A.; Farrell, M.L.; Mertz, J.E. Estrogen-related receptor alpha 1 actively antagonizes estrogen receptor-regulated transcription in MCF-7 mammary cells. J. Biol. Chem. 2002, 277, 24826–24834. [Google Scholar] [CrossRef] [PubMed]
- Acharya, B.R.; Espenel, C.; Kreitzer, G. Direct regulation of microtubule dynamics by KIF17 motor and tail domains. J. Biol. Chem. 2013, 288, 32302–32313. [Google Scholar] [CrossRef] [PubMed]
- Weaver, L.N.; Ems-McClung, S.C.; Stout, J.R.; LeBlanc, C.; Shaw, S.L.; Gardner, M.K.; Walczak, C.E. Kif18A uses a microtubule binding site in the tail for plus-end localization and spindle length regulation. Curr. Biol. 2011, 21, 1500–1506. [Google Scholar] [CrossRef] [PubMed] [Green Version]




{kind=link}
{kind=link}
{kind=link}
| Scan Kinesins for LxxLL Motif --> Define Kinesin Domain Containing LxxLL --> Scan for Monopartite, Classical NLS --> Include/Exclude | |||||||||
|---|---|---|---|---|---|---|---|---|---|
| Kinesin Family | LxxLL (+/− 6 amino acids) | Position in Sequence | Kinesin Domain | NLS | Kinesin Family | LxxLL (+/− 6 amino acids) | Position in Sequence | Kinesin Domain | NLS |
| Kinesin-1 family | Kinesin-6 family | ||||||||
| KIF5A | None | N/A | N/A | + | KIF20A | LHPTPDLKPLLSNEVIW | 201–205 | motor | − |
| KIF5B | None | N/A | N/A | − | EIYNELLYDLLEPPSQQ | 313–317 | motor | ||
| KIF5C | QEREMKLEKLLLLNDKR | 767–771 | coiled-coil | − | DEKIEELEALLQEARQQ | 651–655 | coiled-coil | ||
| PGKKPFLRNLLPRTPTC | 850–854 | tail | |||||||
| Kinesin-2 family | KIF20B | None | N/A | N/A | − | ||||
| KIF23 | None | N/A | N/A | + | |||||
| KIF3A | PYRNSKLTRLLQDSLGG | 302–306 | motor | + | |||||
| KIF3B | PYRDSKLTRLLQDSLGG | 297–301 | motor | + | Kinesin-7 family | ||||
| KIF3C | PYRDSKLTRLLQDSLGG | 322–326 | motor | − | |||||
| KIF17 | PYRDSKLTRLLQDSLGG | 287–301 | motor | + | KIF10/CENPE | AMEKDQLAQLLEEKDLL | 376–380 | coiled-coil | + |
| QMSPSSLSALLSRQVPP | 368–382 | coiled-coil | |||||||
| EPQEVPLQGLLGLQDPF | 601–615 | stalk | Kinesin-8 family | ||||||
| ERDSMLLQQLLEQVQPL | 852–866 | tail | |||||||
| KIF18A | PYRNSKLTRLLKDSLGG | 312–316 | motor | + | |||||
| Kinesin-3 family | RQEYLKLEMLLKENELK | 443–447 | coiled-coil | ||||||
| RQTEAVLNALLPTLRKQ | 573–577 | stalk | |||||||
| KIF1A | PYRDSVLTWLLRENLGG | 311–315 | motor | + | KIF18B | PYRDSKLTRLLKDSLGG | 308–312 | motor | + |
| KDEVTRLRDLLYAQGLG | 379–383 | motor | KIF19 | NYRDSKLTRLLKDSLGG | 303–307 | motor | + | ||
| WYQFTSLRDLLWGNAIF | 722–726 | stalk | |||||||
| GRAFVYLSNLLYPVPLV | 857–861 | tail | Kinesin-9 family | ||||||
| KIF1B | PYRDSVLTWLLRENLGG | 311–315 | motor | + | |||||
| KEEVTRLKDLLRAQGLG | 379–383 | neck-coil or coiled-coil | KIF6 | HHCFHHLKKLLNDKKIL | 428–432 | stalk | − | ||
| SHQFTSLRDLLWGNAVY | 762–766 | stalk | KIF9 | EIYNESLFDLLSTLPYV | 149–153 | motor | − | ||
| GRAFVYLSNLLYPVPLI | 980–984 | tail | |||||||
| KIF1C | PYRDSVLTWLLKENLGG | 305–309 | motor | + | Kinesin-10 family | ||||
| QEEVARLRELLMAQGLS | 373–377 | motor | |||||||
| KIF13A | PYRDSVLTWLLKDNLGG | 309–313 | motor | + | KIF22 | GVIPRALMDLLQLTREE | 151–155 | motor | + |
| LNADPALNELLVYYLKD | 458–462 | stalk | PYRDSKLTRLLQDSLGG | 325–329 | motor | ||||
| KIF13B | PYRDSVLTWLLKDSLGG | 310–314 | motor | + | SMDPAMLERLLSLDRLL | 440–444 | stalk | ||
| LNADPALNELLVYYLKE | 459–463 | stalk | LERLLSLDRLLASQGSQ | 446–450 | stalk | ||||
| KIF14 | PYRESVLTWLLKESLGG | 658–662 | motor | − | |||||
| VLMKHWLSDLLPCTNIA | 1325–1329 | stalk | Kinesin-11 family | ||||||
| KIF16A | PYRDSVLTWLLKDSLGG | 341–345 | motor | + | |||||
| REEIERLKALLLSFELR | 409–413 | coiled-coil | KIF26A | TLRDPCLSALLLDKLPA | 100–104 | N-terminal region | + | ||
| AASRLGLSPLLWKERRA | 617–621 | stalk | CGRDQSLRDLLAEVAPG | 530–534 | motor | ||||
| AYLKNNLPVLLQNQNSK | 1798–1802 | stalk | PYRDHRLTMLLRESLAT | 681–685 | motor | ||||
| HNLSLHLSQLLHSTSEL | 3684–3688 | stalk | KIF26B | WGKEENLRDLLSEVATG | 607–611 | motor | + | ||
| KIF16B | PYRDSVLTWLLKDSLGG | 315–319 | motor | − | PYKESKLAMLLRESLGN | 757–761 | motor | ||
| RAEIARLKTLLAQGNQI | 383–387 | coiled-coil | |||||||
| FHIENKLKDLLAEKEKF | 669–673 | coiled-coil | Kinesin-12 family | ||||||
| QYKERQLQYLLQNHLPT | 909–913 | stalk | |||||||
| YLLQNHLPTLLEEKQRA | 917–921 | stalk | KIF12 | FGSLEALMELLQTGLSR | 204–208 | motor | − | ||
| KIF28 | PYRDSVLTKLLQSALGG | 312–316 | motor | − | PFRDSKLTKLLADSLGG | 317–321 | motor | ||
| KIF15 | CYRDSKLTFLLRDSLGG | 320–324 | motor | − | |||||
| Kinesin-4 family | QKANLNLENLLEATKAC | 630–634 | coiled-coil | ||||||
| KSDLNNLMELLEAEKER | 902–906 | coiled-coil | |||||||
| KIF4A | PYRDSKLTRLLQDSLGG | 293–297 | motor | + | PHFQTHLAKLLETQEQE | 1152–1156 | coiled-coil | ||
| KQQVQQLQVLLLQAHGG | 361–365 | motor | |||||||
| EEAKRHLNDLLEDRKIL | 764–768 | coiled-coil | Kinesin-13 family | ||||||
| KIF4B | PYRDSKLTRLLQDSLGG | 293–297 | motor | + | |||||
| KQQVQQLQVLLLQAHGGT | 361–365 | coiled-coil | KIF2A | None | N/A | N/A | − | ||
| EEAKRHLNDLLEDRKIL | 764–768 | coiled-coil | KIF2B | IEEVETLPTLLGKDTTI | 601–605 | C-terminal domain | + | ||
| KIF7 | None | N/A | N/A | + | KIF2C | AAINPELLQLLPLHPKD | 68–72 | N-terminal domain | − |
| KIF21A | PYRDSKLTRLLQDSLGG | 328–332 | motor | + | EIYNGKLFDLLNKKAKL | 409–413 | motor | ||
| AELNPELDALLGHALQD | 1097–1101 | tail | KIF24 | EIYCGQLYDLLNRRKRL | 367–371 | motor | − | ||
| KIF21B | PYRDSKLTRLLQDSLGG | 327–331 | motor | + | |||||
| KIF27 | EVYKEDLRDLLELETSM | 143–147 | motor | + | Kinesin-14 family | ||||
| KIFC1 | PYRNSKLTYLLQNSLGG | 620–624 | motor | + | |||||
| Kinesin-5 family | KIFC2 | LTVTSQLLALLAWLRSP | 124–128 | N terminal domain | − | ||||
| PFRDSQLTRLLQPALGP | 687–691 | motor | |||||||
| KIF11 | IYNEELFDLLNPSSDV | 168–172 | motor | − | KIFC3 | EIYNEVLRDLLGKEPQE | 582–586 | motor | + |
| NEREQELHNLLEVVSQC | 844–848 | tail | PFRNSKLTYLLQDSLSG | 725–729 | motor | ||||
| KIF25 | GDVCPLLTSLLDGYNVC | 50–54 | motor | − | |||||
| AALAGVLGALLEHRGHA | 303–307 | motor | |||||||
| PYRNSRLTHLLQDCLGG | 320–324 | motor | |||||||
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Seneviratne, A.M.P.B.; Lidagoster, S.; Valbuena-Castor, S.; Lashley, K.; Saha, S.; Alimova, A.; Kreitzer, G. Kinesins Modify ERR1-Dependent Transcription Using a Conserved Nuclear Receptor Box Motif. Int. J. Mol. Sci. 2023, 24, 3795. https://doi.org/10.3390/ijms24043795
Seneviratne AMPB, Lidagoster S, Valbuena-Castor S, Lashley K, Saha S, Alimova A, Kreitzer G. Kinesins Modify ERR1-Dependent Transcription Using a Conserved Nuclear Receptor Box Motif. International Journal of Molecular Sciences. 2023; 24(4):3795. https://doi.org/10.3390/ijms24043795
Chicago/Turabian StyleSeneviratne, A. M. Pramodh Bandara, Sarah Lidagoster, Sofia Valbuena-Castor, Kareena Lashley, Sumit Saha, Aleksandra Alimova, and Geri Kreitzer. 2023. "Kinesins Modify ERR1-Dependent Transcription Using a Conserved Nuclear Receptor Box Motif" International Journal of Molecular Sciences 24, no. 4: 3795. https://doi.org/10.3390/ijms24043795