
CBL-Interacting Protein Kinases 18 (CIPK18) Gene Positively Regulates Drought Resistance in Potato
 ,
,
Abstract
:1. Introduction
2. Results
2.1. Bioinformatics and Expression Analysis of StCIPK18
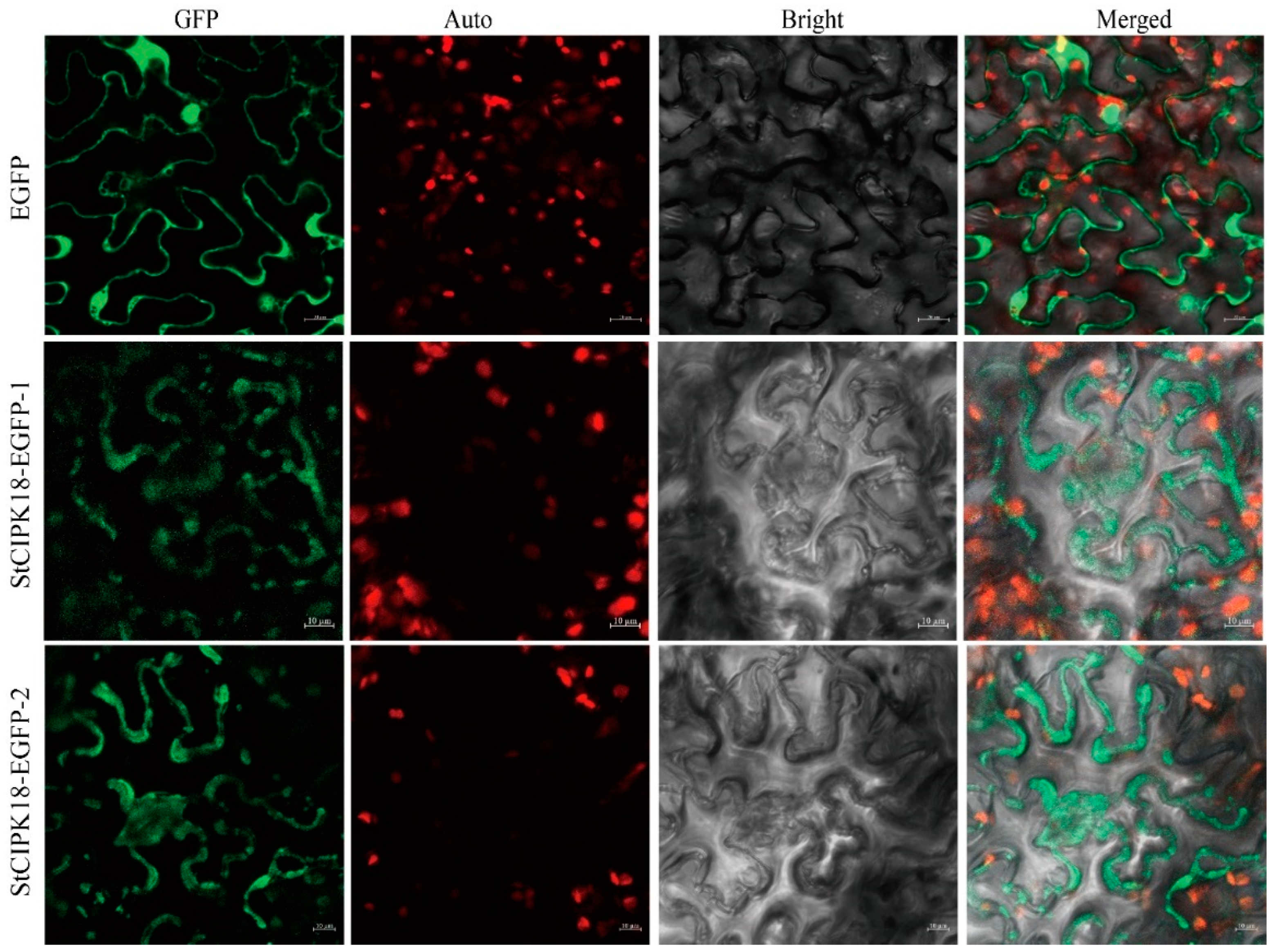
2.2. Cloning and Subcellular Localization of StCIPK18 in Potatoes
2.3. Interaction between StCIPK18 and StCBLs
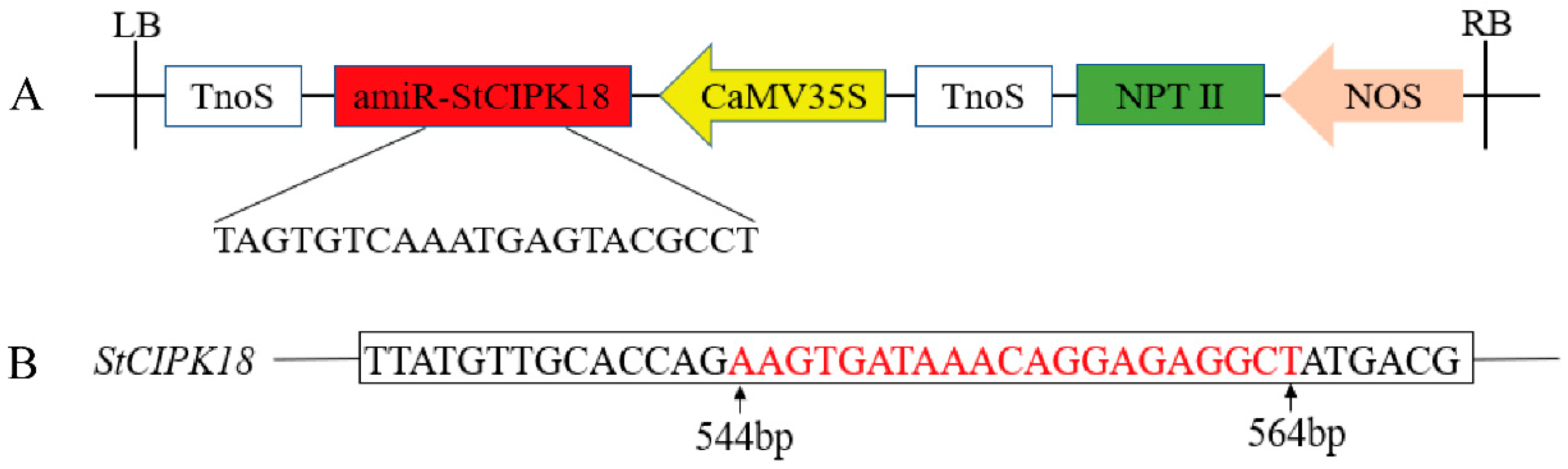
2.4. Genetic Transformation of StCIPK18 Gene in Potatoes and Identification of Transgenic Plants
2.5. StCIPK18 Positively Regulates Potato Drought Stress Resistance
3. Discussion
4. Materials and Methods
4.1. Growth Conditions and Treatment of Plant Materials
4.2. Bioinformatics Analysis
4.3. Cloning of StCIPK18
4.4. StCIPK18 Expression Analysis by qRT-PCR
4.5. Subcellular Localization of StCIPK18
4.6. Yeast Two-Hybrid Assay
4.7. Bimolecular Fluorescence Complementarity Assay
4.8. Construction of Plant Expression Vectors
4.9. Genetic Transformation of Potatoes and Identification of Transgenic Plants
4.10. Drought Stress Treatment
4.11. Statistical Analyses
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Ma, X.; Li, Q.; Yu, Y.; Qiao, Y.; Saeed, U.H.; Gong, Z. The CBL–CIPK pathway in plant response to stress signals. Int. J. Mol. Sci. 2020, 21, 5668. [Google Scholar] [CrossRef]
- Batistic, O.; Kudla, J. Integration and channeling of calcium signaling through the CBL calcium sensor/CIPK protein kinase network. Planta 2004, 219, 915–924. [Google Scholar] [CrossRef]
- Sheng, J.; Zheng, Z.; Pan, W.; Pan, J. Functions and action Mechanisms of CBL-CIPK signaling system in plants. Plant Physiol. J. 2014, 50, 641–650. [Google Scholar]
- Qiu, Q.; Guo, Y.; Dietrich, M.A.; Schumaker, K.S.; Zhu, J. Regulation of SOS1, a plasma membrane Na+/H+ exchanger in Arabidopsis thaliana, by SOS2 and SOS3. Proc. Natl. Acad. Sci. USA 2002, 99, 8436–8441. [Google Scholar] [CrossRef]
- Ma, R.; Li, S.; Liu, W.; Wang, Z.; Yang, J.; Tang, X.; Zhang, N.; Si, H. Functions and progress in mechanism research of CBL-CIPK signaling system in plants. Plant Physiol. J. 2021, 57, 521–530. [Google Scholar]
- Qiu, Q.; Guo, Y.; Quintero, F.J.; Pardo, J.M.; Schumaker, K.S.; Zhu, J. Regulation of vacuolar Na+/H+ exchange in Arabidopsis thaliana by the salt-overly-sensitive (SOS) pathway. J. Biol. Chem. 2004, 279, 207–215. [Google Scholar] [CrossRef]
- Xu, J.; Li, H.; Chen, L.; Wang, Y.; Liu, L.; Wu, W. A protein kinase, interacting with two calcineurin B-like proteins, regulates K+ transporter AKT1 in Arabidopsis. Cell 2006, 125, 1347–1360. [Google Scholar] [CrossRef]
- Behera, S.; Long, Y.; Schmitz, T.I.; Wang, X.; Zhang, C.; Li, H.; Steinhorst, L.; Manishankar, P.; Ren, X.; Offenborn, J.N.; et al. Two spatially and temporally distinct Ca2+ signals convey Arabidopsis thaliana responses to K+ deficiency. New Phytol. 2017, 213, 739–750. [Google Scholar] [CrossRef]
- Chen, L.; Wang, Q.; Zhou, L.; Ren, F.; Li, D.; Li, X. Arabidopsis CBL-interacting protein kinase (CIPK6) is involved in plant response to salt/osmotic stress and ABA. Mol. Biol. Rep. 2013, 40, 4759–4767. [Google Scholar] [CrossRef]
- Cheong, Y.; Pandey, G.; Grant, J.; Batistic, O.; Li, L.; Kim, B.G.; Lee, S.C.; Kudla, J.; Luan, S. Two calcineurin B-like calcium sensors, interacting with protein kinase CIPK23, regulate leaf transpiration and root potassium uptake in Arabidopsis. Plant J. 2007, 52, 223–239. [Google Scholar] [CrossRef]
- Li, J.; Long, Y.; Qi, G.; Xu, Z.; Wu, W.; Wang, Y. The Os-AKT1 channel is critical for K+ uptake in rice roots and is modulated by the rice CBL1-CIPK23 complex. Plant Cell 2014, 26, 3387–3402. [Google Scholar] [CrossRef]
- Chen, X.; Wang, D.; Guan, H.; Guo, J.; Sha, X.; Li, Y.; Zhang, D.; Liu, X.; He, G.; Shi, Y.; et al. Identification of CIPK gene family members and investigation of the drought tolerance of ZmCIPK3 in Maize. J. Plant Genet. Resour. 2020, 23, 1064–1075. [Google Scholar]
- Xiang, Y.; Huang, Y.; Xiong, L. Characterization of stress-responsive CIPK genes in rice for stress tolerance improvement. Plant Physiol. 2007, 144, 1416–1428. [Google Scholar] [CrossRef] [Green Version]
- Cui, X.; Du, Y.; Fu, J.; Yu, T.; Wang, C.; Chen, M.; Chen, J.; Ma, Y.; Xu, Z. Wheat CBL-interacting protein kinase 23 positively regulates drought stress and ABA responses. BMC Plant Biol. 2018, 18, 93. [Google Scholar] [CrossRef]
- Ma, R.; Liu, W.; Li, S.; Zhu, X.; Yang, J.; Zhang, N.; Si, H. Genome-wide identification, characterization and expression analysis of the CIPK gene family in potato (Solanum tuberosum L.) and the role of StCIPK10 in response to drought and osmotic stress. Int. J. Mol. Sci. 2021, 22, 13535. [Google Scholar] [CrossRef]
- Rudack, K.; Seddig, S.; Sprenger, H.; Köhl, K.; Uptmoor, R.; Ordon, F. Drought stress-induced changes in starch yield and physiological traits in potato. J. Agron. Crop Sci. 2017, 203, 494–505. [Google Scholar] [CrossRef]
- D’Angelo, C.; Weinl, S.; Batistic, O.; Pandey, G.K.; Cheong, Y.H.; Schultke, S.; A1brecht, V.; Ehlert, B.; Schulz, B.H.; Arter, K.; et al. Alternative complex formation of the Ca2+-regulated protein kinase CIPK1 controls abscisic acid- dependent and independent stress response s in Arabidopsis. Plant J. 2006, 48, 857–872. [Google Scholar] [CrossRef]
- Quintero, F.J.; Ohta, M.; Shi, H.; Zhu, J.; Pardo, J.M. Reconstitution in yeast of the Arabidopsis SOS signaling pathway for Na+ homeostasis. Proc. Natl. Acad. Sci. USA 2002, 99, 9061–9066. [Google Scholar] [CrossRef]
- Kim, B.; Waadt, R.; Cheong, Y.; Pandey, G.; Dominguez Solis, J.R.; Schültke, S.; Lee, S.C.; Kudla, J.; Luan, S. The calcium sensor CBL10 mediates salt tolerance by regulating ion homeostasis in Arabidopsis. Plant J. 2007, 52, 473–484. [Google Scholar] [CrossRef]
- Quan, R.; Lin, H.; Mendoza, I.; Zhang, Y.; Cao, W.; Yang, Y.; Shang, M.; Chen, S.; Pardo, J.; Guo, Y. SCaBP8/CBL10, a putative calcium sensor, interacts with the protein kinase SOS2 to protect Arabidopsis shoots from salt stress. Plant Cell 2007, 19, 1415–1431. [Google Scholar] [CrossRef]
- Cheong, Y.; Kim, K.; Pandey, G.; Gupta, R.; Grant, J.; Luan, S. CBL1, a calcium sensor that differentially regulates salt, drought, and cold responses in Arabidopsis. Plant Cell 2003, 15, 1833–1845. [Google Scholar] [CrossRef]
- Pandey, G.; Cheong, Y.; Kim, K.; Grant, J.; Li, L.; Hung, W.; D’Angelo, C.; Weinl, S.; Kudla, J.; Luan, S. The calcium sensor calcineurin B-like 9 modulates abscisic acid sensitivity and biosynthesis in Arabidopsis. Plant Cell 2004, 16, 1912–1924. [Google Scholar] [CrossRef]
- Rajashekar, C.B.; Panda, M. Water stress is a component of cold acclimation process essential for inducing full freezing tolerance in strawberry. Sci. Hortic. 2014, 174, 54–59. [Google Scholar] [CrossRef] [Green Version]
- Griffiths, C.A.; Gaff, D.F.; Neale, A.D. Drying without senescence in resurrection plants. Front. Plant Sci. 2014, 5, 36. [Google Scholar] [CrossRef]
- Fichman, Y.; Mittler, R. Rapid systemic signaling during abiotic and biotic stresses: Is the ROS wave master of all trades. Plant J. 2020, 102, 887–896. [Google Scholar] [CrossRef]
- Madhulika, S.; Jitendra, K.; Samiksha, S.; Singh, V.P.; Prasad, S.M. Roles of osmoprotectants in improving salinity and drought tolerance in plants: A review. Rev. Environ. Sci. Biotechnol. 2015, 14, 407–426. [Google Scholar]
- Chun, S.C.; Paramasivan, M.; Chandrasekaran, M. Proline accumulation influenced by osmotic stress in arbuscular mycorrhizal symbiotic plants. Front. Microbiol. 2018, 9, 2525. [Google Scholar] [CrossRef]
- Zhang, J.; Yang, K.; Zhang, Y.; Yan, Y.; Zhao, Z.; Fang, R.; Sun, Z.; Fu, Y.; Chen, X. Identification and characterization of rice OsCIPK10 gene. Chin. J. Biotechnol. 2009, 25, 1394–1401. [Google Scholar]
- Wang, Y.; Sun, T.; Li, T.; Wang, M.; Yang, G.; He, G. A CBL-interacting protein kinase TaCIPK2 confers drought tolerance in transgenic tobacco plants through regulating the stomatal movement. PLoS ONE 2016, 11, e0167962. [Google Scholar] [CrossRef]
- Nieves, C.M.; Caballero, F.; Martinez, V. Disruption of the Arabidopsis thaliana inward-rectifier K+ channel AKT1 improves plant responses to water stress. Plant Cell Physiol. 2012, 53, 423–432. [Google Scholar] [CrossRef]
- Deng, X.; Hu, W.; Wei, S. TaCIPK29, a CBL-interacting protein kinase gene from wheat, confers salt stress tolerance in transgenic tobacco. PLoS ONE 2013, 8, e69881. [Google Scholar] [CrossRef] [PubMed]
- Zhang, N.; Si, H.; Wang, D. Cloning of rd29A gene promoter from Arabidopsis thaliana and its application in stress-resistance transgenic potato. Acta Agron. Sin. 2005, 31, 159–164. [Google Scholar]
- Li, S.; Zhang, N.; Zhu, X.; Ma, R.; Liu, S.; Wang, X.; Yang, J.; Si, H. Genome-wide analysis of NF-Y genes in potato and functional identification of StNF-YC9 in drought tolerance. Front. Plant Sci. 2021, 12, 749688. [Google Scholar] [CrossRef] [PubMed]
- Zhu, X.; Zhang, N.; Liu, X.; Li, S.; Yang, J.; Hong, X.; Wang, F.; Si, H. Mitogen-activated protein kinase 11 (MAPK11) maintains growth and photosynthesis of potato plant under drought condition. Plant Cell Rep. 2021, 40, 491–506. [Google Scholar] [CrossRef] [PubMed]
- Tang, X.; Zhang, N.; Si, H.; Calderón-Urrea, A. Selection and validation of reference genes for RT-qPCR analysis in potato under abiotic stress. Plant Methods 2017, 13, 85. [Google Scholar] [CrossRef]
- Cai, C.; Wang, W.; Zhang, J.; Deng, M.; Huang, T.; Yu, L.; Li, L.; Wang, X. Identification and sequence analysis of CBL family genes in potato. Mol. Plant Breed. 2019, 17, 2778–2784. [Google Scholar]
- Li, S.; Zhang, N.; Zhu, X.; Ma, R.; Yang, J.; Tang, X.; Si, H. Enhanced drought tolerance with artificial microRNA-mediated StProDH1 gene silencing in potato. Crop Sci. 2020, 60, 1462–1471. [Google Scholar] [CrossRef]
- Hayta, S.; Smedley, M.A.; Li, J.; Harwood, W.A.; Gilmartin, P.M. Agrobacterium-mediated transformation systems of Primula vulgaris. Plant Methods 2018, 14, 93. [Google Scholar] [CrossRef]
- Si, H.; Xie, C.; Liu, J. An efficient protocol for Agrobacteriu mediated transformation with microtuber and the introduction of an antisense class i patatin gene into potato. Acta Agron. Sin. 2003, 29, 801–805. [Google Scholar]
- Zhu, X.; Wang, F.; Li, S.; Feng, Y.; Yang, J.; Zhang, N.; Si, H. Calcium-dependent protein kinase 28 maintains potato photosynthesis and its tolerance under water deficiency and osmotic Stress. Int. J. Mol. Sci. 2022, 23, 8795. [Google Scholar] [CrossRef]
- You, L.; Song, Q.; Wu, Y.; Li, S.; Jiang, C.; Chang, L. Accumulation of glycine betaine in transplastomic potato plants expressing choline oxidase confers improved drought tolerance. Planta 2019, 249, 1963–1975. [Google Scholar] [CrossRef] [PubMed]
- Heath, R.L.; Packer, L. Photoperoxidation in isolated chloroplasts I kinetics and stoichiometry of fatty acid peroxidation. Arch. Biochem. Biophys. 1968, 125, 189–198. [Google Scholar] [CrossRef] [PubMed]
- Bates, L.S.; Waldren, R.P.; Teare, I.D. Rapid determination of free proline for water-stress studies. Plant Soil 1973, 39, 205–207. [Google Scholar] [CrossRef]
- Giannopolitis, C.N.; Ries, S.K. Superoxide dismutases: I. Occurrence in higher plants. Plant Physiol. 1977, 59, 309–314. [Google Scholar] [CrossRef] [PubMed]
- Maehly, A.C.; Chance, B. The assay of catalases and peroxidases. Methods Biochem. Anal. 1954, 1, 357–424. [Google Scholar] [PubMed]
- Aebi, H. Catalase in vitro. Methods Enzymol. 1984, 105, 121–126. [Google Scholar] [PubMed]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Name | Sequence | Number | Function |
|---|---|---|---|
| ABRE | ACGTG | 1 | cis-acting element involved in the abscisic acid responsiveness |
| ARE | AAACCA | 2 | cis-acting regulatory element essential for the anaerobic induction |
| CAAT-box | CAAAT | 13 | common cis-acting element in promoter and enhancer regions |
| MBS | CAACTG | 1 | MYB binding site involved in drought-inducibility |
| TC-rich repeats | GTTTTCTTAC | 1 | cis-acting element involved in defense and stress responsiveness |
| TCA-element | CCATCTTTTT | 1 | cis-acting element involved in salicylic acid responsiveness |
| TCCC-motif | TCTCCCT | 1 | part of a light responsive element |
| TCT-motif | TCTTAC | 1 | part of a light responsive element |
| ATC-motif | AGCTATCCA | 1 | part of a conserved DNA module involved in light responsiveness |
| Box 4 | ATTAAT | 2 | part of a conserved DNA module involved in light responsiveness |
| G-Box | CACGTT | 1 | cis-acting regulatory element involved in light responsiveness |
| Sp1 | GGGCGG | 1 | light responsive element |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Yang, L.; Zhang, N.; Wang, K.; Zheng, Z.; Wei, J.; Si, H. CBL-Interacting Protein Kinases 18 (CIPK18) Gene Positively Regulates Drought Resistance in Potato. Int. J. Mol. Sci. 2023, 24, 3613. https://doi.org/10.3390/ijms24043613
Yang L, Zhang N, Wang K, Zheng Z, Wei J, Si H. CBL-Interacting Protein Kinases 18 (CIPK18) Gene Positively Regulates Drought Resistance in Potato. International Journal of Molecular Sciences. 2023; 24(4):3613. https://doi.org/10.3390/ijms24043613
Chicago/Turabian StyleYang, Liang, Ning Zhang, Kaitong Wang, Zhiyong Zheng, Jingjing Wei, and Huaijun Si. 2023. "CBL-Interacting Protein Kinases 18 (CIPK18) Gene Positively Regulates Drought Resistance in Potato" International Journal of Molecular Sciences 24, no. 4: 3613. https://doi.org/10.3390/ijms24043613