
Maternal Donor and Genetic Variation of Lagerstroemia indica Cultivars
 and
and
Abstract
:1. Introduction
2. Results
2.1. Plastome and nrDNA Sequences of Lagerstroemia indica Cultivars
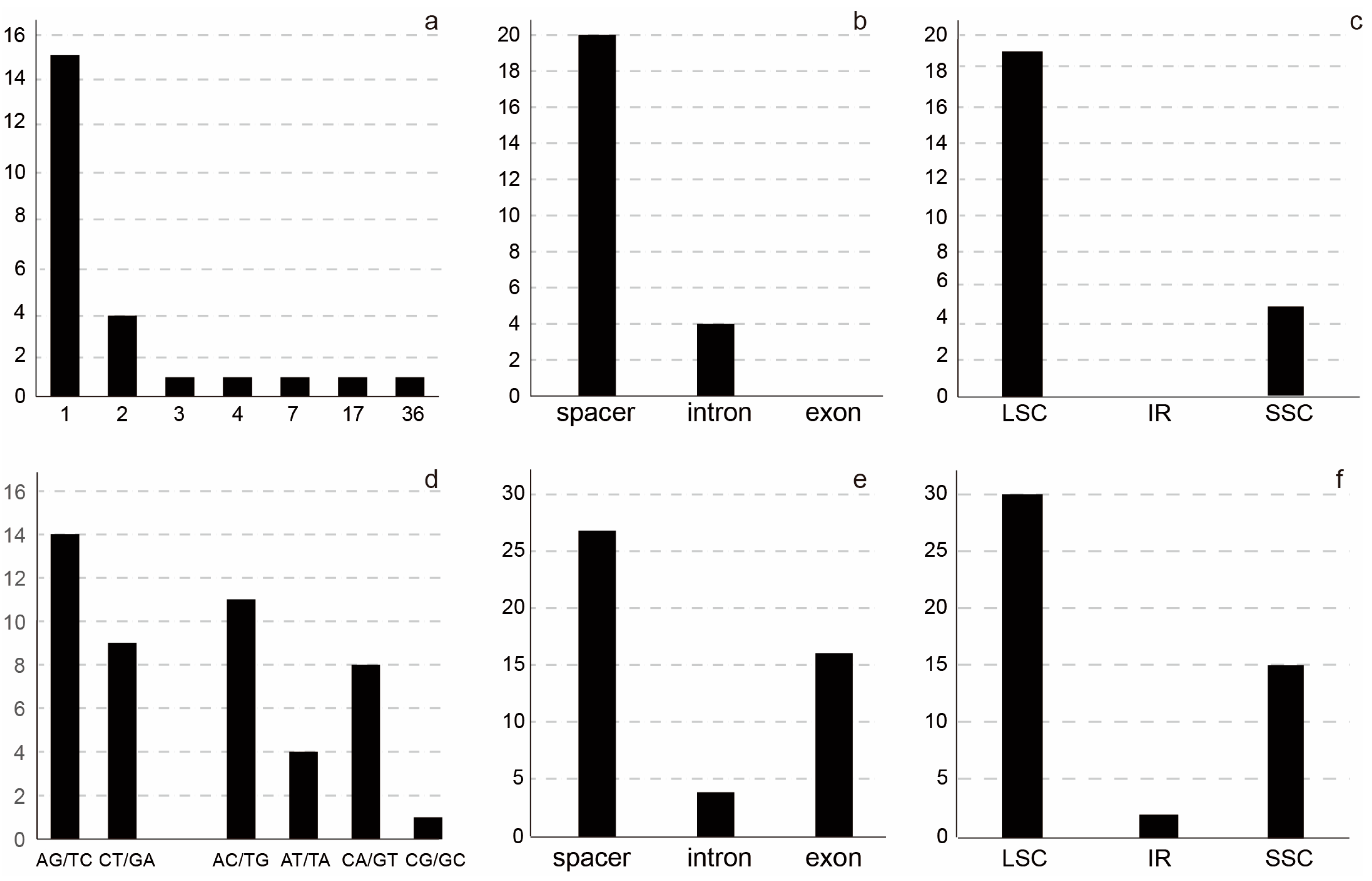
2.2. Plastome Variation in the Lagerstroemia indica Cultivars
2.3. Nuclear Ribosomal DNA Variability
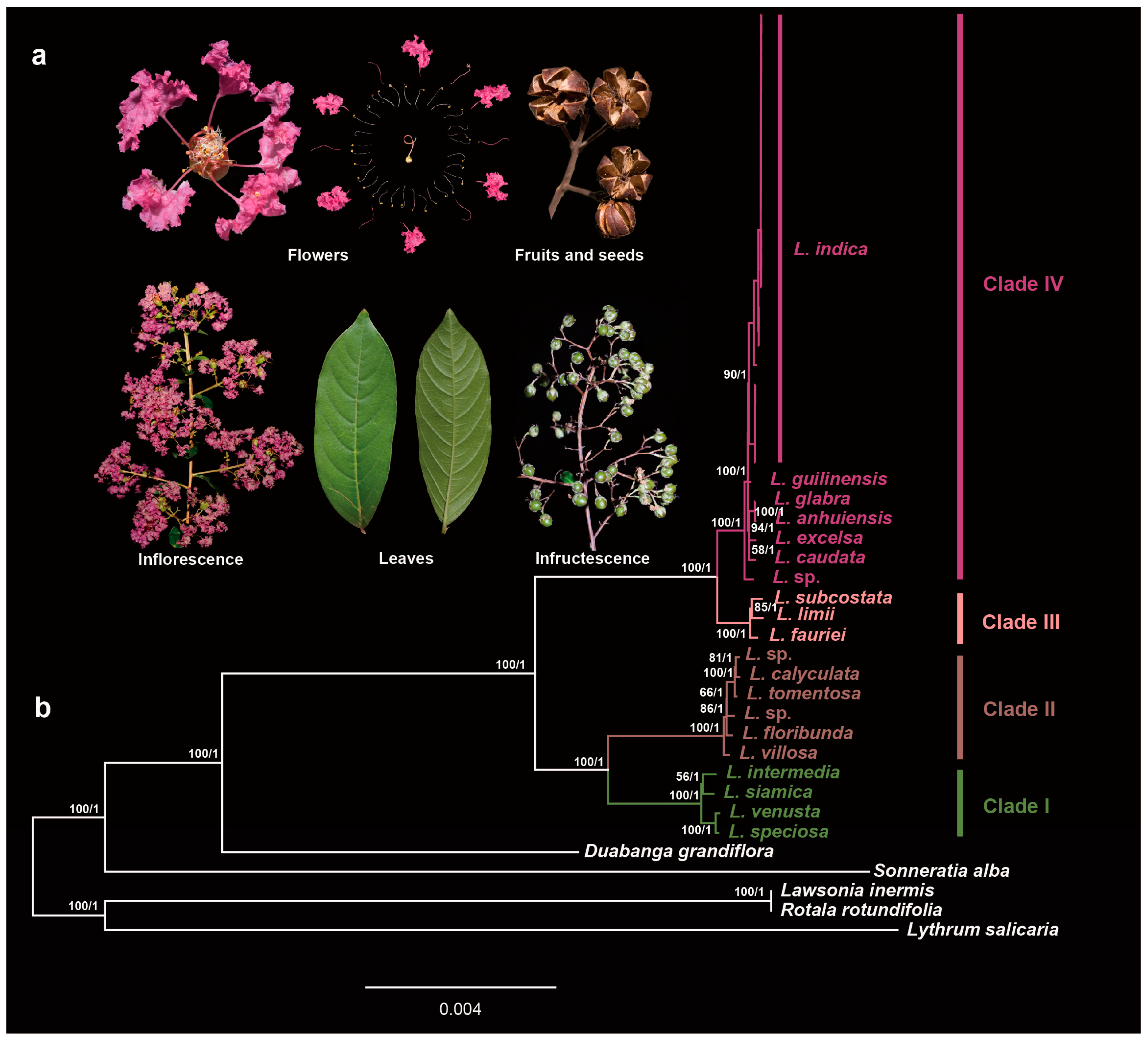
2.4. Maternal Origin of the Cultivars
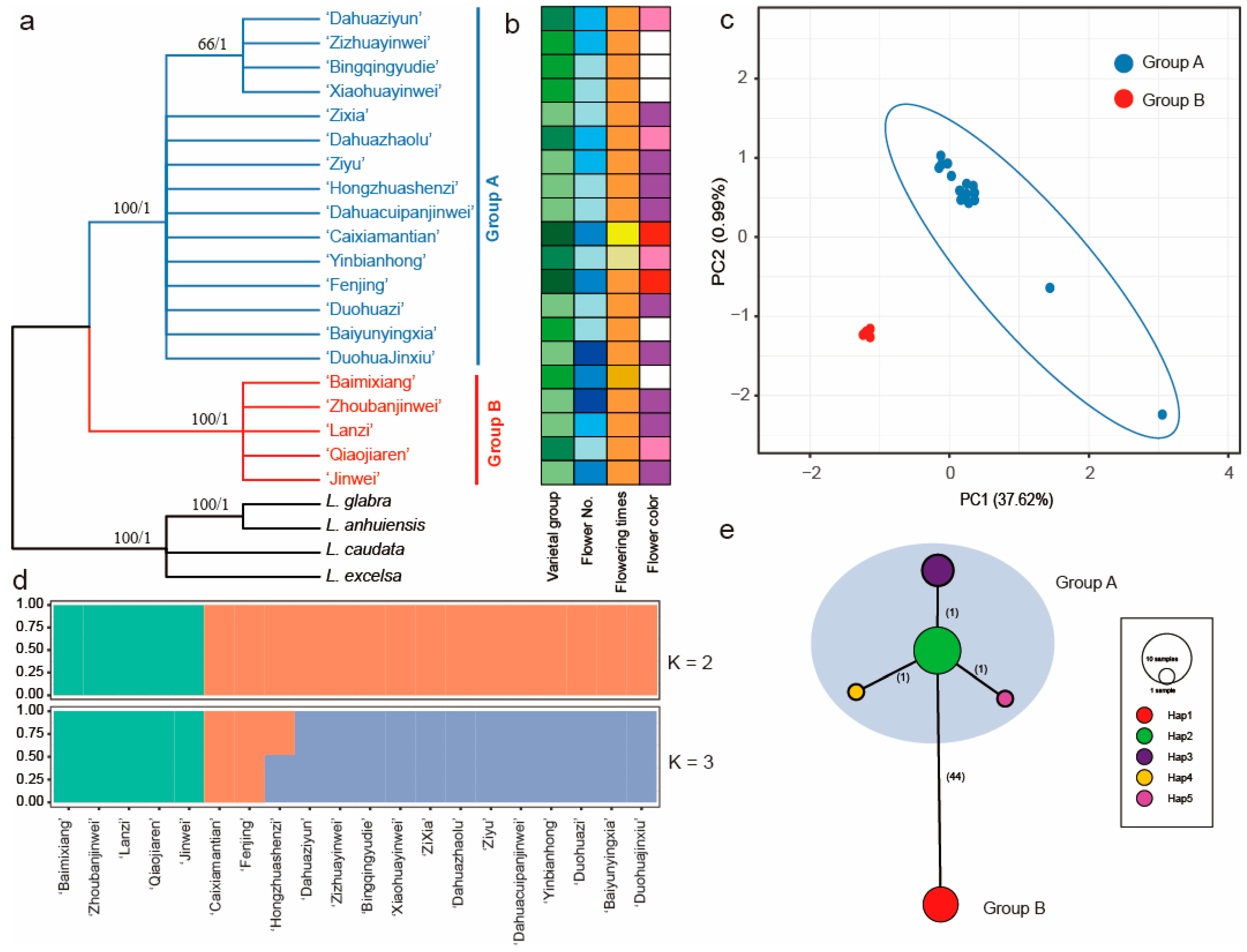
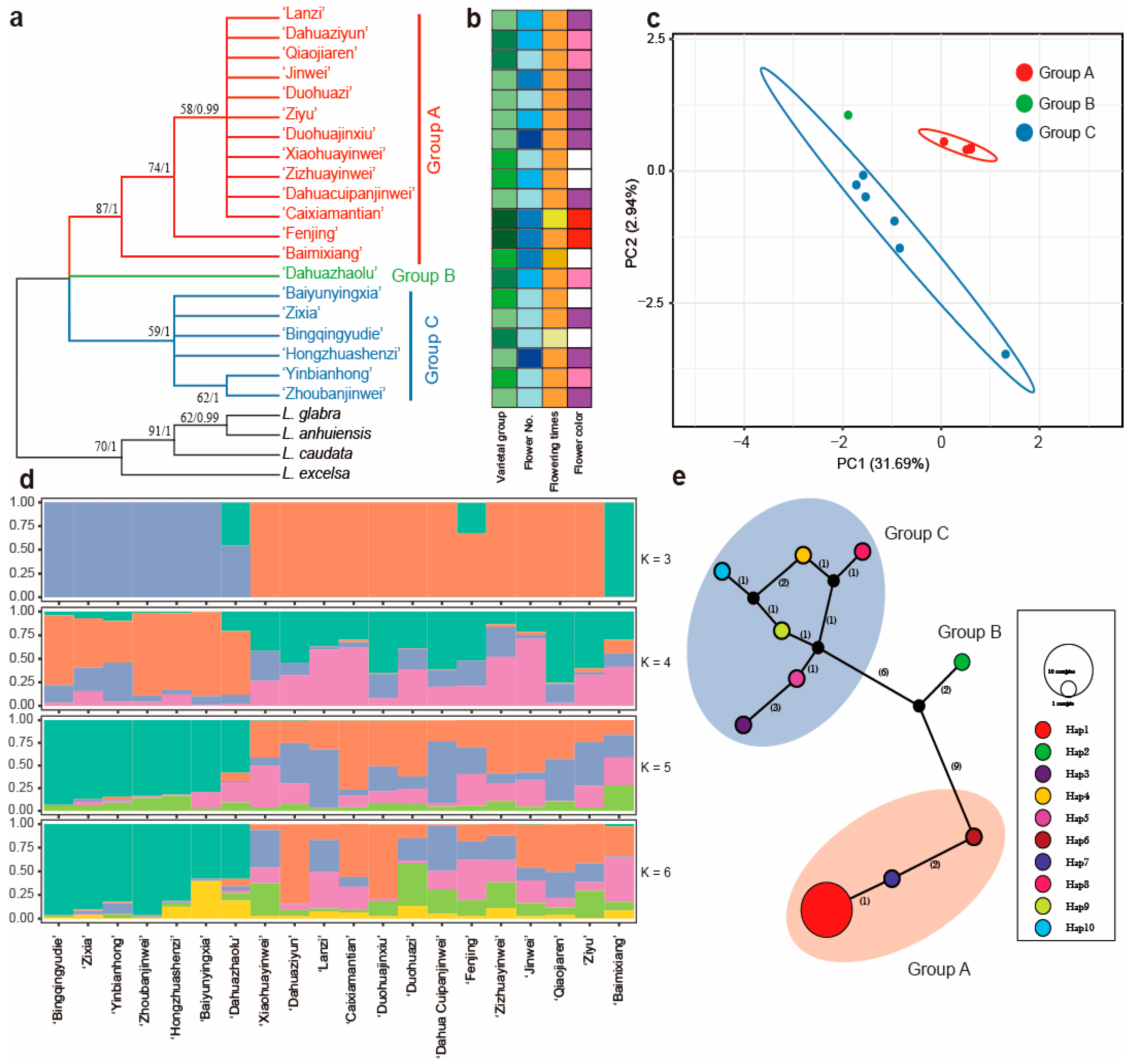
2.5. Genetic Variation Based on the Plastome Sequences
2.6. Genetic Variation Based on the nrDNA Sequences
2.7. Phenotypic Characterization
3. Discussion
3.1. Maternal Donor of Crape Myrtle Cultivars
3.2. Phenotypic Diversity and Genetic Variation of Lagerstroemia indica Cultivars
3.3. Utility of Plastome and nrDNA for Accessing Genetic Diversity of Cultivars
4. Materials and Methods
4.1. Sampling, DNA Extraction, and Sequencing
4.2. Plastome and nrDNA Assembly
4.3. Plastome and nrDNA Variation Analyses
4.4. Phylogenetic Analyses
4.5. Genetic Variation and Diversity Analyses
4.6. Phenotypic Analyses
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Zhang, Q. Studies on cultivars of crape-myrtles (Lagerstroemia indica) and their uses in urban greening. J. Beijing For. Univ. 1991, 13, 57–66. [Google Scholar]
- Cai, M.; Pan, H.-T.; Wang, X.-F.; He, D.; Wang, X.-Y.; Wang, X.-J.; Zhang, Q.-X. Development of novel microsatellites in Lagerstroemia indica and DNA fingerprinting in Chinese Lagerstroemia cultivars. Sci. Hortic. 2011, 131, 88–94. [Google Scholar] [CrossRef]
- Pounders, C.; Rinehart, T.; Sakhanokho, H. Evaluation of Interspecific Hybrids between Lagerstroemia indica and L. speciosa. HortSci. Horts 2007, 42, 1317–1322. [Google Scholar] [CrossRef]
- Ye, Y.M.; Tong, J.; Shi, X.P.; Yuan, W.; Li, G.R. Morphological and cytological studies of diploid and colchicine-induced tetraploid lines of crape myrtle (Lagerstroemia indica L.). Sci. Hortic. 2010, 124, 95–101. [Google Scholar] [CrossRef]
- Rinehart, T.A.; Pounders, C.T. Estimating Diversity among Lagerstroemia Species and Hybrids Using SSR Markers. Acta Hortic. 2010, 885, 285–290. [Google Scholar] [CrossRef]
- Wang, Y.-S.; Hou, B.-X.; Suo, Z.-L.; Liu, T.-H.; Chen, L.-J.; Yang, X.-J. Genetic Diversity of Lagerstroemia indica Germplasm Resources in Hunan Based on Morphological Characteristics. J. Plant Genet. Resour. 2015, 16, 71–79. [Google Scholar]
- Margaret, R.P. Molecular Genetic Diversity Among 12 Clones of Lagerstroemia fauriei Revealed by AFLP and RAPD Markers. HortScience 2003, 38, 256–259. [Google Scholar]
- Ming, C.; Miao, T.; Min, W.; Pan, H.-T.; Qi-Xiang, Z. Analysis of genetic diversity and relationship of Chinese Lagerstroemia indica cultivars based on AFLP and morphological markers. Acta Hortic. 2012, 938, 509–516. [Google Scholar] [CrossRef]
- Peng, C.; Li, Z.; Ma, L.; Huang, G.; Xu, H.; Yang, Y. Identification and genetic analysis in Lagerstroemia interspecific hybrids using SSR markers. North. Hortic. 2020, 13, 83–90. [Google Scholar]
- Liu, Y.; He, D.; Cai, M.; Tang, W.; Li, X.Y.; Pan, H.T.; Zhang, Q.X. Development of microsatellite markers for Lagerstroemia indica (Lythraceae) and related species. Appl. Plant Sci. 2013, 1, 1200203. [Google Scholar] [CrossRef]
- Suo, Z.; Li, W.; Jin, X.; Zhang, H. A new nuclear DNA marker revealing both microsatellite variations and single nucleotide polymorphic loci: A case study on classification of cultivars in Lagerstroemia indica L. J. Microb. Biochem. Technol. 2016, 8, 266–271. [Google Scholar] [CrossRef]
- He, D.; Liu, Y.; Cai, M.; Pan, H.; Zhang, Q.; Wang, X.; Wang, X. Genetic diversity of Lagerstroemia (Lythraceae) species assessed by simple sequence repeat markers. Genet. Mol. Res. 2012, 11, 3522–3533. [Google Scholar] [CrossRef] [PubMed]
- Dong, W.; Liu, Y.; Xu, C.; Gao, Y.; Yuan, Q.; Suo, Z.; Zhang, Z.; Sun, J. Chloroplast phylogenomic insights into the evolution of Distylium (Hamamelidaceae). BMC Genom. 2021, 22, 293. [Google Scholar] [CrossRef]
- Dong, W.; Li, E.; Liu, Y.; Xu, C.; Wang, Y.; Liu, K.; Cui, X.; Sun, J.; Suo, Z.; Zhang, Z.; et al. Phylogenomic approaches untangle early divergences and complex diversifications of the olive plant family. BMC Biol. 2022, 20, 92. [Google Scholar] [CrossRef] [PubMed]
- Dong, W.; Liu, Y.; Li, E.; Xu, C.; Sun, J.; Li, W.; Zhou, S.; Zhang, Z.; Suo, Z. Phylogenomics and biogeography of Catalpa (Bignoniaceae) reveal incomplete lineage sorting and three dispersal events. Mol. Phylogenet. Evol. 2022, 166, 107330. [Google Scholar] [CrossRef] [PubMed]
- Gao, Y.; Liu, K.; Li, E.; Wang, Y.; Xu, C.; Zhao, L.; Dong, W. Dynamic evolution of the plastome in the Elm family (Ulmaceae). Planta 2023, 257, 14. [Google Scholar] [CrossRef]
- Li, E.; Liu, K.; Deng, R.; Gao, Y.; Liu, X.; Dong, W.; Zhang, Z. Insights into the phylogeny and chloroplast genome evolution of Eriocaulon (Eriocaulaceae). BMC Plant Biol. 2023, 23, 32. [Google Scholar] [CrossRef] [PubMed]
- Sun, J.; Wang, S.; Wang, Y.; Wang, R.; Liu, K.; Li, E.; Qiao, P.; Shi, L.; Dong, W.; Huang, L.; et al. Phylogenomics and Genetic Diversity of Arnebiae Radix and Its Allies (Arnebia, Boraginaceae) in China. Front. Plant Sci. 2022, 13, 920826. [Google Scholar] [CrossRef] [PubMed]
- Dong, W.-P.; Sun, J.-H.; Liu, Y.-L.; Xu, C.; Wang, Y.-H.; Suo, Z.-L.; Zhou, S.-L.; Zhang, Z.-X.; Wen, J. Phylogenomic relationships and species identification of the olive genus Olea (Oleaceae). J. Syst. Evol. 2022, 60, 1263–1280. [Google Scholar] [CrossRef]
- Shang, C.; Li, E.; Yu, Z.; Lian, M.; Chen, Z.; Liu, K.; Xu, L.; Tong, Z.; Wang, M.; Dong, W. Chloroplast Genomic Resources and Genetic Divergence of Endangered Species Bretschneidera sinensis (Bretschneideraceae). Front. Ecol. Evol. 2022, 10, 873100. [Google Scholar] [CrossRef]
- Torre, S.; Sebastiani, F.; Burbui, G.; Pecori, F.; Pepori, A.L.; Passeri, I.; Ghelardini, L.; Selvaggi, A.; Santini, A. Novel Insights into Refugia at the Southern Margin of the Distribution Range of the Endangered Species Ulmus laevis. Front. Plant Sci. 2022, 13, 826158. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.; Sun, J.; Zhao, Z.; Xu, C.; Qiao, P.; Wang, S.; Wang, M.; Xu, Z.; Yuan, Q.; Guo, L.; et al. Multiplexed Massively Parallel Sequencing of Plastomes Provides Insights into the Genetic Diversity, Population Structure, and Phylogeography of Wild and Cultivated Coptis chinensis. Front. Plant Sci. 2022, 13, 923600. [Google Scholar] [CrossRef] [PubMed]
- Perdereau, A.; Klaas, M.; Barth, S.; Hodkinson, T.R. Plastid genome sequencing reveals biogeographical structure and extensive population genetic variation in wild populations of Phalaris arundinacea L. in north-western Europe. GCB Bioenergy 2017, 9, 46–56. [Google Scholar] [CrossRef]
- Xue, C.; Geng, F.D.; Li, J.J.; Zhang, D.Q.; Gao, F.; Huang, L.; Zhang, X.H.; Kang, J.Q.; Zhang, J.Q.; Ren, Y. Divergence in the Aquilegia ecalcarata complex is correlated with geography and climate oscillations: Evidence from plastid genome data. Mol. Ecol. 2021, 30, 5796–5813. [Google Scholar] [CrossRef]
- Huang, D.I.; Hefer, C.A.; Kolosova, N.; Douglas, C.J.; Cronk, Q.C. Whole plastome sequencing reveals deep plastid divergence and cytonuclear discordance between closely related balsam poplars, Populus balsamifera and P. trichocarpa (Salicaceae). New Phytol. 2014, 204, 693–703. [Google Scholar] [CrossRef]
- Cui, H.; Ding, Z.; Zhu, Q.; Wu, Y.; Gao, P. Population structure and genetic diversity of watermelon (Citrullus lanatus) based on SNP of chloroplast genome. 3 Biotech 2020, 10, 374. [Google Scholar] [CrossRef]
- Xu, Y.; Liao, B.; Ostevik, K.L.; Zhou, H.; Wang, F.; Wang, B.; Xia, H. The Maternal Donor of Chrysanthemum Cultivars Revealed by Comparative Analysis of the Chloroplast Genome. Front. Plant Sci. 2022, 13, 923442. [Google Scholar] [CrossRef]
- Nock, C.J.; Hardner, C.M.; Montenegro, J.D.; Ahmad Termizi, A.A.; Hayashi, S.; Playford, J.; Edwards, D.; Batley, J. Wild Origins of Macadamia Domestication Identified Through Intraspecific Chloroplast Genome Sequencing. Front. Plant Sci. 2019, 10, 334. [Google Scholar] [CrossRef] [PubMed]
- Tong, W.; Kim, T.-S.; Park, Y.-J. Rice Chloroplast Genome Variation Architecture and Phylogenetic Dissection in Diverse Oryza Species Assessed by Whole-Genome Resequencing. Rice 2016, 9, 57. [Google Scholar] [CrossRef] [PubMed]
- Dong, W.; Xu, C.; Liu, Y.; Shi, J.; Li, W.; Suo, Z. Chloroplast phylogenomics and divergence times of Lagerstroemia (Lythraceae). BMC Genom. 2021, 22, 434. [Google Scholar] [CrossRef]
- Wang, Y.; Hou, B.; Yang, Q.; Zhou, H.; Chen, L.; Yang, X. Investigation and analysis of application prospects of Lagerstroemia indica germplasm resources in Hunan Province. Acta Prataculturae Sin. 2014, 23, 77–91. [Google Scholar]
- Wang, J.; He, W.; Liao, X.; Ma, J.; Gao, W.; Wang, H.; Wu, D.; Tembrock, L.R.; Wu, Z.; Gu, C. Phylogeny, molecular evolution, and dating of divergences in Lagerstroemia using plastome sequences. Hortic. Plant J. 2022, in press. [Google Scholar] [CrossRef]
- Furtado, C.; Srisuko, M. A revision of Lagerstroemia L. (Lythraceae). Gard. Bull. 1969, 24, 185–334. [Google Scholar]
- Cai, M.; Meng, R.; Pan, H.-T.; Gao, Y.-K.; Sun, M.; Song, P.; Wang, X.-F.; Zhang, Q.-X. Isolation and characterization of microsatellite markers from Lagerstroemia caudata (Lythraceae) and cross-amplification in other related species. Conserv. Genet. Resour. 2010, 2, 89–91. [Google Scholar] [CrossRef]
- Xu, L.L.; Yu, R.M.; Lin, X.R.; Zhang, B.W.; Li, N.; Lin, K.; Zhang, D.Y.; Bai, W.N. Different rates of pollen and seed gene flow cause branch-length and geographic cytonuclear discordance within Asian butternuts. New Phytol. 2021, 232, 388–403. [Google Scholar] [CrossRef]
- Rose, J.P.; Toledo, C.A.P.; Lemmon, E.M.; Lemmon, A.R.; Sytsma, K.J. Out of sight, out of mind: Widespread nuclear and plastid-nuclear discordance in the flowering plant genus Polemonium (Polemoniaceae) suggests widespread historical gene flow despite limited nuclear signal. Syst. Biol. 2021, 70, 162–180. [Google Scholar] [CrossRef]
- Sarver, B.A.J.; Herrera, N.D.; Sneddon, D.; Hunter, S.S.; Settles, M.L.; Kronenberg, Z.; Demboski, J.R.; Good, J.M.; Sullivan, J. Diversification, Introgression, and Rampant Cytonuclear Discordance in Rocky Mountains Chipmunks (Sciuridae: Tamias). Syst. Biol. 2021, 70, 908–921. [Google Scholar] [CrossRef]
- Xiao, S.; Xu, P.; Deng, Y.; Dai, X.; Zhao, L.; Heider, B.; Zhang, A.; Zhou, Z.; Cao, Q. Comparative analysis of chloroplast genomes of cultivars and wild species of sweetpotato (Ipomoea batatas [L.] Lam). BMC Genom. 2021, 22, 262. [Google Scholar]
- Kim, K.; Lee, S.-C.; Lee, J.; Lee, H.O.; Joh, H.J.; Kim, N.-H.; Park, H.-S.; Yang, T.-J. Comprehensive Survey of Genetic Diversity in Chloroplast Genomes and 45S nrDNAs within Panax ginseng Species. PLoS ONE 2015, 10, e0117159. [Google Scholar] [CrossRef]
- Li, L.; Hu, Y.; He, M.; Zhang, B.; Wu, W.; Cai, P.; Huo, D.; Hong, Y. Comparative chloroplast genomes: Insights into the evolution of the chloroplast genome of Camellia sinensis and the phylogeny of Camellia. BMC Genom. 2021, 22, 138. [Google Scholar] [CrossRef]
- Zhao, F.; Chen, Y.-P.; Salmaki, Y.; Drew, B.T.; Wilson, T.C.; Scheen, A.-C.; Celep, F.; Bräuchler, C.; Bendiksby, M.; Wang, Q.; et al. An updated tribal classification of Lamiaceae based on plastome phylogenomics. BMC Biol. 2021, 19, 2. [Google Scholar] [CrossRef] [PubMed]
- Yang, L.; Abduraimov, O.; Tojibaev, K.; Shomurodov, K.; Zhang, Y.-M.; Li, W.-J. Analysis of complete chloroplast genome sequences and insight into the phylogenetic relationships of Ferula L. BMC Genom. 2022, 23, 643. [Google Scholar] [CrossRef] [PubMed]
- Dong, W.; Xu, C.; Li, C.; Sun, J.; Zuo, Y.; Shi, S.; Cheng, T.; Guo, J.; Zhou, S. ycf1, the most promising plastid DNA barcode of land plants. Sci. Rep. 2015, 5, 8348. [Google Scholar] [CrossRef] [PubMed]
- Dong, W.; Cheng, T.; Li, C.; Xu, C.; Long, P.; Chen, C.; Zhou, S. Discriminating plants using the DNA barcode rbcLb: An appraisal based on a large dataset. Mol. Ecol. Resour. 2014, 14, 336–343. [Google Scholar] [CrossRef]
- Dong, W.; Liu, J.; Yu, J.; Wang, L.; Zhou, S. Highly variable chloroplast markers for evaluating plant phylogeny at low taxonomic levels and for DNA barcoding. PLoS ONE 2012, 7, e35071. [Google Scholar] [CrossRef]
- Zheng, G.; Wei, L.; Ma, L.; Wu, Z.; Gu, C.; Chen, K. Comparative analyses of chloroplast genomes from 13 Lagerstroemia (Lythraceae) species: Identification of highly divergent regions and inference of phylogenetic relationships. Plant Mol. Biol. 2020, 102, 659–676. [Google Scholar] [CrossRef]
- Hu, G.; Wu, Y.; Guo, C.; Lu, D.; Dong, N.; Chen, B.; Qiao, Y.; Zhang, Y.; Pan, Q. Haplotype Analysis of Chloroplast Genomes for Jujube Breeding. Front. Plant Sci. 2022, 13, 841767. [Google Scholar] [CrossRef]
- Liu, H.; Zhao, W.; Hua, W.; Liu, J. A large-scale population based organelle pan-genomes construction and phylogeny analysis reveal the genetic diversity and the evolutionary origins of chloroplast and mitochondrion in Brassica napus L. BMC Genom. 2022, 23, 339. [Google Scholar] [CrossRef]
- Smith, S.A.; Donoghue, M.J. Rates of molecular evolution are linked to life history in flowering plants. Science 2008, 322, 86–89. [Google Scholar] [CrossRef]
- Schwarz, E.N.; Ruhlman, T.A.; Weng, M.-L.; Khiyami, M.A.; Sabir, J.S.M.; Hajarah, N.H.; Alharbi, N.S.; Rabah, S.O.; Jansen, R.K. Plastome-wide nucleotide substitution rates reveal accelerated rates in Papilionoideae and correlations with genome features across legume subfamilies. J. Mol. Evol. 2017, 84, 187–203. [Google Scholar] [CrossRef]
- Choi, K.; Weng, M.-L.; Ruhlman, T.A.; Jansen, R.K. Extensive variation in nucleotide substitution rate and gene/intron loss in mitochondrial genomes of Pelargonium. Mol. Phylogenet. Evol. 2021, 155, 106986. [Google Scholar] [CrossRef]
- Wang, X.; Wadl, P.A.; Pounders, C.; Trigiano, R.N.; Cabrera, R.I.; Scheffler, B.E.; Pooler, M.; Rinehart, T.A. Evaluation of Genetic Diversity and Pedigree within Crapemyrtle Cultivars Using Simple Sequence Repeat Markers. J. Am. Soc. Hort. Sci. 2011, 136, 116–128. [Google Scholar] [CrossRef]
- Cheng, T.; Xu, C.; Lei, L.; Li, C.; Zhang, Y.; Zhou, S. Barcoding the kingdom Plantae: New PCR primers for ITS regions of plants with improved universality and specificity. Mol. Ecol. Resour. 2016, 16, 138–149. [Google Scholar] [CrossRef] [PubMed]
- Kim, O.T.; Bang, K.H.; In, D.S.; Lee, J.W.; Kim, Y.C.; Shin, Y.S.; Hyun, D.Y.; Lee, S.S.; Cha, S.W.; Seong, N.S. Molecular authentication of ginseng cultivars by comparison of internal transcribed spacer and 5.8S rDNA sequences. Plant Biotechnol. Rep. 2007, 1, 163–167. [Google Scholar] [CrossRef]
- Hapsari, L.; Azrianingsih, R.; Arumingtyas, E.L. Genetic Variability and Relationship of Banana Cultivars (Musa L.) From East Java, Indonesia based on the Internal Transcribed Spacer Region nrDNA Sequences. J. Trop. Biol. Conserv. 2018, 15, 101–120. [Google Scholar] [CrossRef]
- Hidayat, T.; Abdullah, F.I.; Kuppusamy, C.; Samad, A.A.; Wagiran, A. Molecular Identification of Malaysian Pineapple Cultivar based on Internal Transcribed Spacer Region. APCBEE Procedia 2012, 4, 146–151. [Google Scholar] [CrossRef]
- Li, J.; Wang, S.; Jing, Y.; Wang, L.; Zhou, S. A modified CTAB protocol for plant DNA extraction. Chin. Bull. Bot. 2013, 48, 72–78. [Google Scholar]
- Bolger, A.M.; Lohse, M.; Usadel, B. Trimmomatic: A flexible trimmer for Illumina sequence data. Bioinformatics 2014, 30, 2114–2120. [Google Scholar] [CrossRef] [Green Version]
- Jin, J.-J.; Yu, W.-B.; Yang, J.-B.; Song, Y.; dePamphilis, C.W.; Yi, T.-S.; Li, D.-Z. GetOrganelle: A fast and versatile toolkit for accurate de novo assembly of organelle genomes. Genome Biol. 2020, 21, 241. [Google Scholar] [CrossRef]
- Bankevich, A.; Nurk, S.; Antipov, D.; Gurevich, A.A.; Dvorkin, M.; Kulikov, A.S.; Lesin, V.M.; Nikolenko, S.I.; Pham, S.; Prjibelski, A.D.; et al. SPAdes: A New Genome Assembly Algorithm and Its Applications to Single-Cell Sequencing. J. Comput. Biol. 2012, 19, 455–477. [Google Scholar] [CrossRef]
- Altschul, S.F.; Gish, W.; Miller, W.; Myers, E.W.; Lipman, D.J. Basic Local Alignment Search Tool. J. Mol. Biol. 1990, 215, 403–410. [Google Scholar] [CrossRef]
- Kearse, M.; Moir, R.; Wilson, A.; Stones-Havas, S.; Cheung, M.; Sturrock, S.; Buxton, S.; Cooper, A.; Markowitz, S.; Duran, C.; et al. Geneious Basic: An integrated and extendable desktop software platform for the organization and analysis of sequence data. Bioinformatics 2012, 28, 1647–1649. [Google Scholar] [CrossRef] [PubMed]
- Huang, D.I.; Cronk, Q.C.B. Plann: A command-line application for annotating plastome sequences. Appl. Plant Sci. 2015, 3, 1500026. [Google Scholar] [CrossRef] [PubMed]
- Katoh, K.; Standley, D.M. MAFFT multiple sequence alignment software version 7: Improvements in performance and usability. Mol. Biol. Evol. 2013, 30, 772–780. [Google Scholar] [CrossRef] [PubMed]
- Kumar, S.; Stecher, G.; Li, M.; Knyaz, C.; Tamura, K. MEGA X: Molecular Evolutionary Genetics Analysis across Computing Platforms. Mol. Biol. Evol. 2018, 35, 1547–1549. [Google Scholar] [CrossRef] [PubMed]
- Rozas, J.; Ferrer-Mata, A.; Sanchez-DelBarrio, J.C.; Guirao-Rico, S.; Librado, P.; Ramos-Onsins, S.E.; Sanchez-Gracia, A. DnaSP 6: DNA sequence polymorphism analysis of large data sets. Mol. Biol. Evol. 2017, 34, 3299–3302. [Google Scholar] [CrossRef]
- Kalyaanamoorthy, S.; Minh, B.Q.; Wong, T.K.F.; von Haeseler, A.; Jermiin, L.S. ModelFinder: Fast model selection for accurate phylogenetic estimates. Nat. Methods 2017, 14, 587–589. [Google Scholar] [CrossRef]
- Kozlov, A.M.; Darriba, D.; Flouri, T.; Morel, B.; Stamatakis, A. RAxML-NG: A fast, scalable and user-friendly tool for maximum likelihood phylogenetic inference. Bioinformatics 2019, 35, 4453–4455. [Google Scholar] [CrossRef] [Green Version]
- Ronquist, F.; Teslenko, M.; van der Mark, P.; Ayres, D.L.; Darling, A.; Hohna, S.; Larget, B.; Liu, L.; Suchard, M.A.; Huelsenbeck, J.P. MrBayes 3.2: Efficient Bayesian phylogenetic inference and model choice across a large model space. Syst. Biol. 2012, 61, 539–542. [Google Scholar] [CrossRef]
- Rambaut, A.; Suchard, M.; Xie, D.; Drummond, A. Tracer v1. 6. 2014. Available online: http://beast.community/tracer (accessed on 1 July 2022).
- Purcell, S.; Neale, B.; Todd-Brown, K.; Thomas, L.; Ferreira, M.A.R.; Bender, D.; Maller, J.; Sklar, P.; de Bakker, P.I.W.; Daly, M.J.; et al. PLINK: A Tool Set for Whole-Genome Association and Population-Based Linkage Analyses. Am. J. Hum. Genet. 2007, 81, 559–575. [Google Scholar] [CrossRef]
- Wickham, H. (Ed.) Data Analysis. In ggplot2: Elegant Graphics for Data Analysis; Springer International Publishing: Cham, Switzerland, 2016; pp. 189–201. [Google Scholar]
- Excoffier, L.; Laval, G.; Schneider, S. Arlequin (version 3.0): An integrated software package for population genetics data analysis. Evol. Bioinform. 2005, 1, 47–50. [Google Scholar] [CrossRef]
- Leigh, J.W.; Bryant, D. POPART: Full-feature software for haplotype network construction. Methods Ecol. Evol. 2015, 6, 1110–1116. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Samples | Varietal Group | Flower Number | Flowering Times | Flower Color | Origin | GenBank Accession Number of Plastome | GenBank Accession Number of nrDNA |
|---|---|---|---|---|---|---|---|
| ‘Dahuazhaolu’ | ‘Bicolor’ Group | 43–98 | Middle period (July and August) | Bicolor | Jishou, Hunan | OP613198 | OP723643 |
| ‘Ziyu’ | ‘Amabilis’ Group | 62–112 | Middle period (July and August) | Purple | Changde, Hunan | OP613199 | OP723644 |
| ‘Duohuazi’ | ‘Amabilis’ Group | 64–172 | Middle period (July and August) | Purple | Liuyang, Hunan | OP613200 | OP723645 |
| ‘Qiaojiaren’ | ‘Bicolor’ Group | 61–138 | Middle period (July and August) | Bicolor | Wuhan, Hubei | OP613201 | OP723646 |
| ‘Zizhuayinwei’ | ‘Alba’ Group | 69–102 | Middle period (July and August) | White | Guiyang, Guizhou | OP613202 | OP723647 |
| ‘Yinbianhong’ | ‘Bicolor’ Group | 86–113 | Early flowering period (before July) | Bicolor | Jishou, Hunan | OP613203 | OP723648 |
| ‘Bingqingyudie’ | ‘Alba’ Group | 76–158 | Middle period (July and August) | White | Guiyang, Guizhou | OP613204 | OP723649 |
| ‘JinWei’ | ‘Amabilis’ Group | 83–185 | Middle period (July and August) | Purple | Changsha, Hunan | OP613205 | OP723650 |
| ‘Zixia’ | ‘Amabilis’ Group | 66–119 | Middle period (July and August) | Purple | Changde, Hunan | OP613206 | OP723651 |
| ‘Zhoubanjinwei’ | ‘Amabilis’ Group | 153–225 | Middle period (July and August) | Purple | Changsha, Hunan | OP613207 | OP723652 |
| ‘Caixiamantian’ | ‘Rubra’ Group | 98–198 | Late flowering (September) | Red | Chengdu, Sichuan | OP613208 | OP723653 |
| ‘Baiyunyingxia’ | ‘Alba’ Group | 57–125 | Middle period (July and August) | White | Changsha, Hunan | OP613209 | OP723654 |
| ‘Baimixiang’ | ‘Alba’ Group | 100–198 | Flowering long | White | Changsha, Hunan | OP613210 | OP723655 |
| ‘Lanzi’ | ‘Amabilis’ Group | 59–106 | Middle period (July and August) | Purple | Changde, Hunan | OP613211 | OP723656 |
| ‘Fenjing’ | ‘Rubra’ Group | 97–198 | Middle period (July and August) | Red | Chengdu, Sichuan | OP613212 | OP723657 |
| ‘Dahuaziyun’ | ‘Bicolor’ Group | 43–122 | Middle period (July and August) | Bicolor | Shaoyang, Hunan | OP613213 | OP723658 |
| ‘Xiaohuayinwei’ | ‘Alba’ Group | 69–145 | Middle period (July and August) | White | Chengdu, Sichuan | OP613214 | OP723659 |
| ‘Duohuajinxiu’ | ‘Amabilis’ Group | 138–222 | Middle period (July and August) | Purple | Shaoyang, Hunan | OP613215 | OP723660 |
| ‘Hongzhuashenzi’ | ‘Amabilis’ Group | 64–164 | Middle period (July and August) | Purple | Shaoyang, Hunan | OP613216 | OP723661 |
| ‘DahuacuipanJinwei’ | ‘Amabilis’ Group | 75–156 | Middle period (July and August) | Purple | Shaoyang, Hunan | OP613217 | OP723662 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Guo, C.; Liu, K.; Li, E.; Chen, Y.; He, J.; Li, W.; Dong, W.; Suo, Z. Maternal Donor and Genetic Variation of Lagerstroemia indica Cultivars. Int. J. Mol. Sci. 2023, 24, 3606. https://doi.org/10.3390/ijms24043606
Guo C, Liu K, Li E, Chen Y, He J, Li W, Dong W, Suo Z. Maternal Donor and Genetic Variation of Lagerstroemia indica Cultivars. International Journal of Molecular Sciences. 2023; 24(4):3606. https://doi.org/10.3390/ijms24043606
Chicago/Turabian StyleGuo, Chang, Kangjia Liu, Enze Li, Yifeng Chen, Jiayao He, Wenying Li, Wenpan Dong, and Zhili Suo. 2023. "Maternal Donor and Genetic Variation of Lagerstroemia indica Cultivars" International Journal of Molecular Sciences 24, no. 4: 3606. https://doi.org/10.3390/ijms24043606