
Phylogeny and Historical Biogeography of the East Asian Clematis Group, Sect. Tubulosae, Inferred from Phylogenomic Data
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results
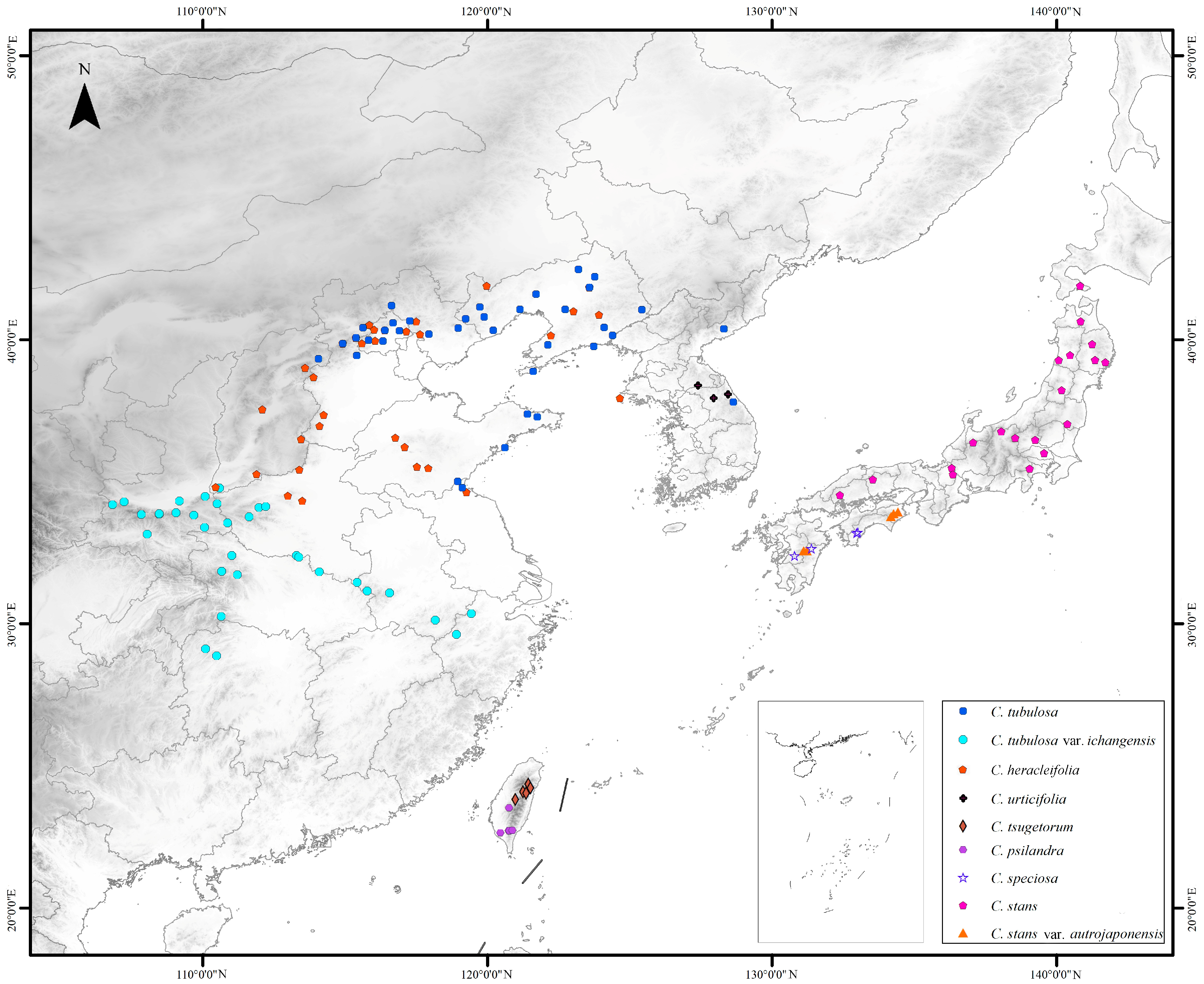
2.1. Features of the Complete Plastid Genome, nrDNA, and SNPs Sequences
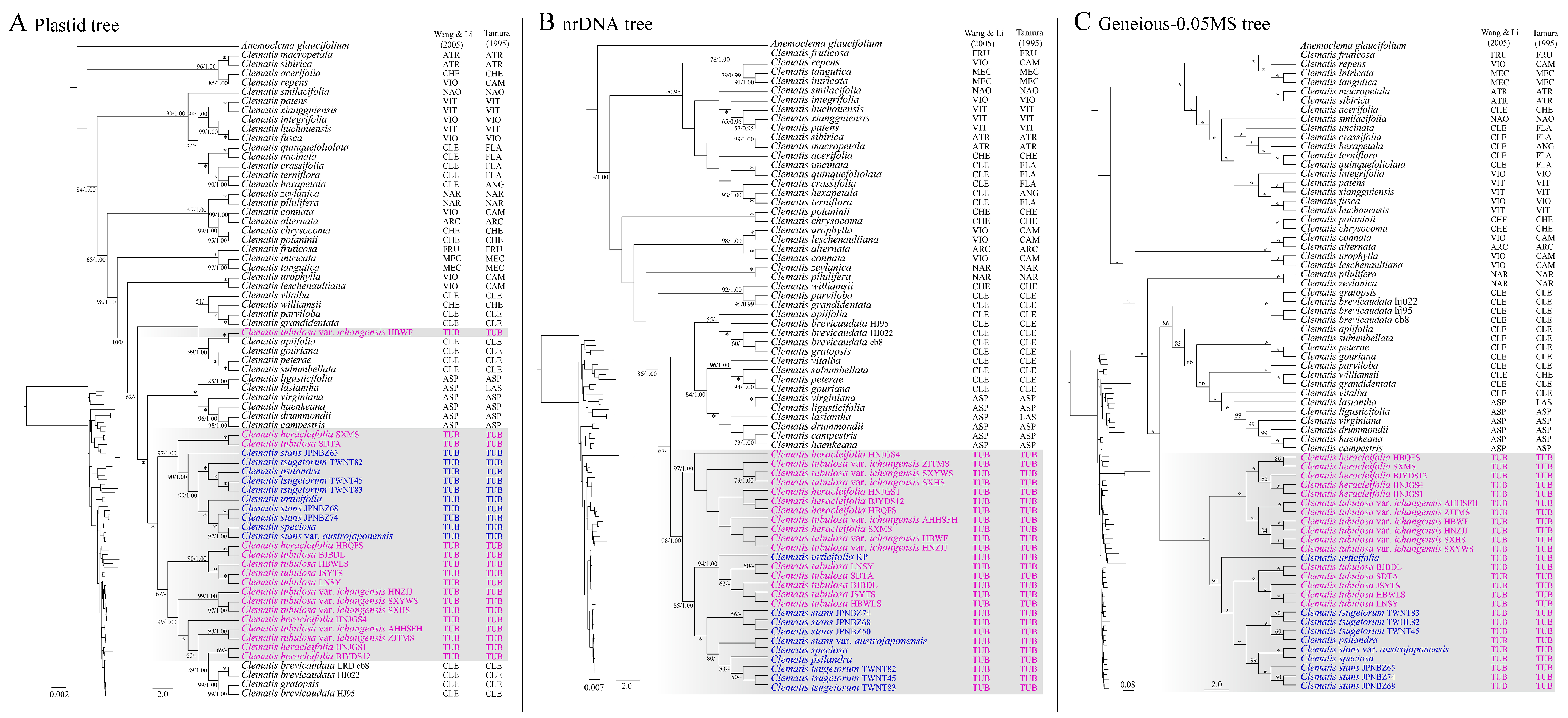
2.2. Phylogenetic Analyses
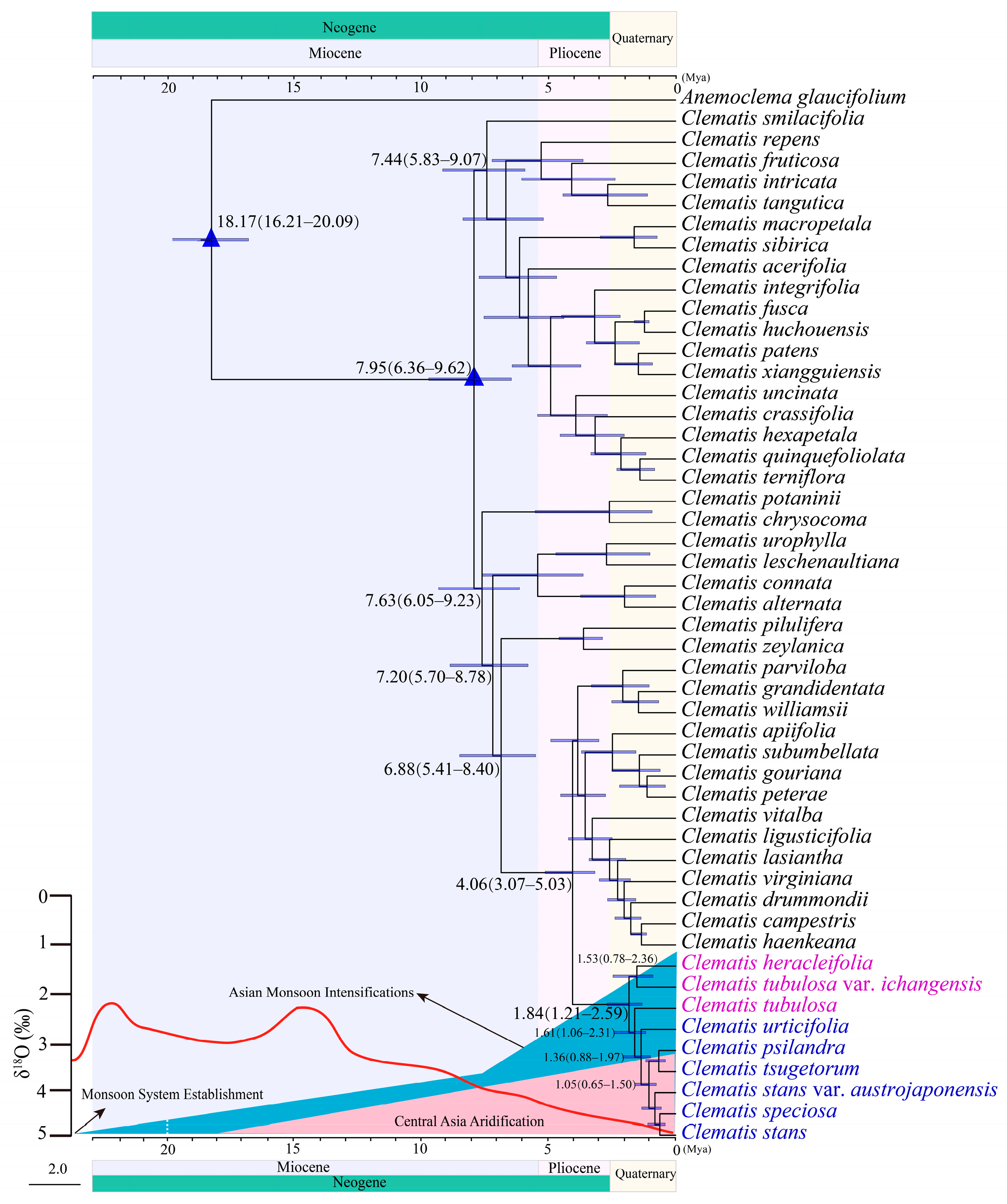
2.3. Divergence Time Estimation
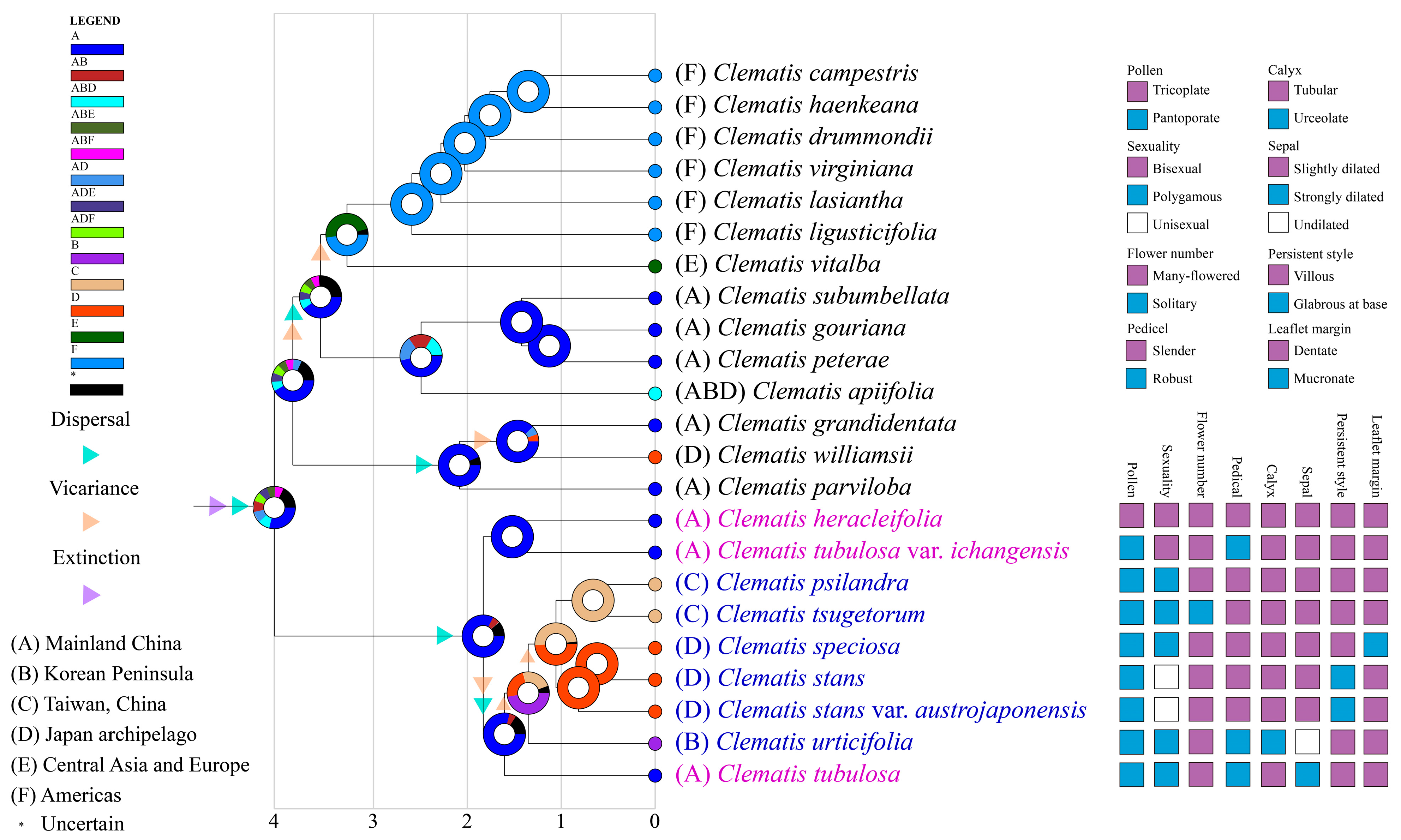
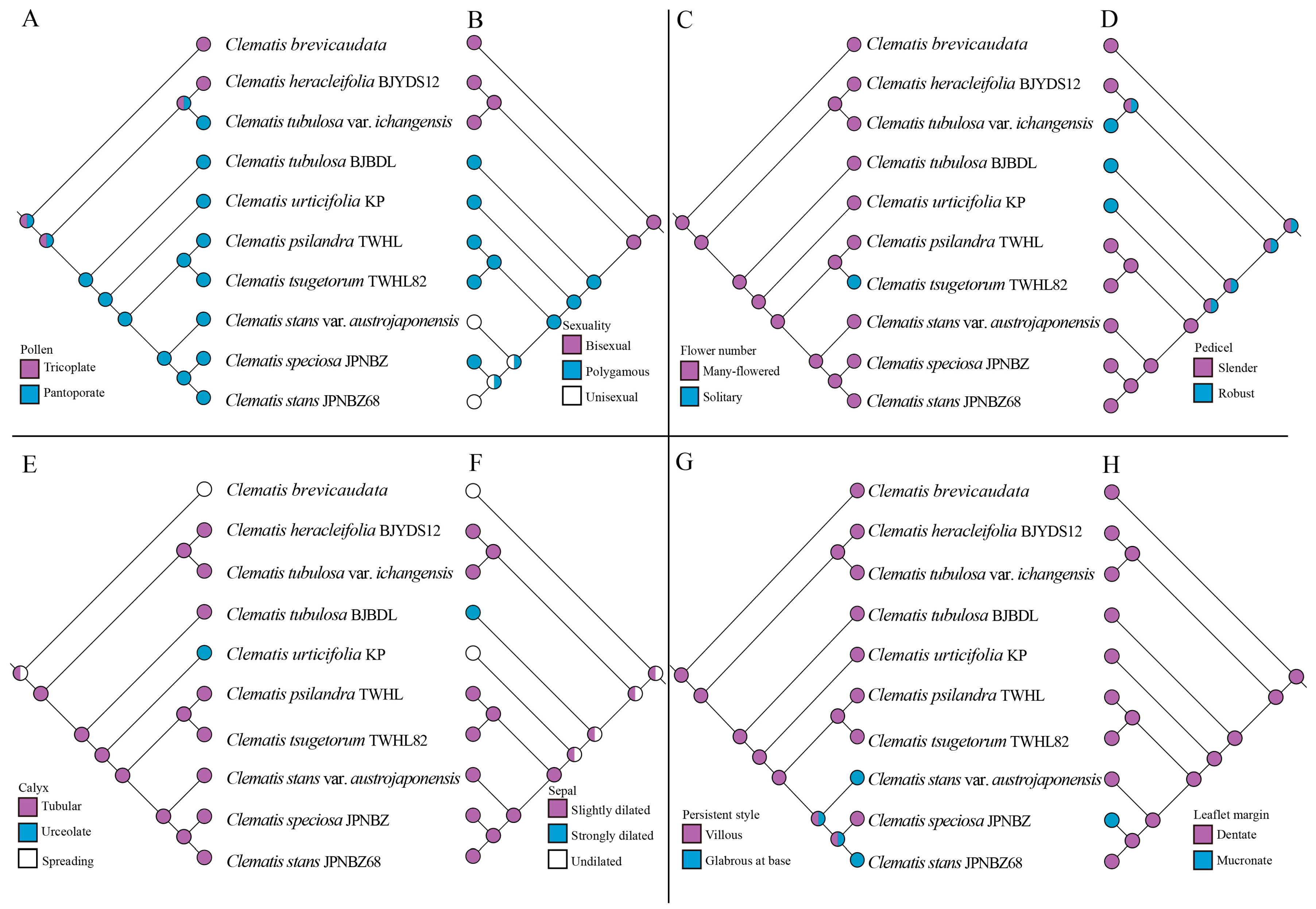
2.4. Ancestral Area and Morphological Character Reconstruction
3. Discussion
3.1. Phylogenetic Position of Sect. Tubulosae
3.2. Species Divergence of Sect. Tubulosae
3.3. Historical Biogeography of Sect. Tubulosae in the Sino-Japanese Floristic Region
4. Materials and Methods
4.1. Plant Material and Taxon Sampling
4.2. DNA Extraction and Sequencing
4.3. Plastid Genome, nrDNA, and Nuclear SNPs Assembly and Annotation
4.4. Phylogenetic Analysis
4.5. Molecular Dating
4.6. Ancestral Area and Morphological Character Reconstruction
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Takhtajan, A. Flowering Plants: Origin and Dispersal; Smithsonian Institution Press: Washington, DC, USA, 1969. [Google Scholar]
- Wu, C.Y.; Wu, S.K. A proposal for a new floristic kingdom (realm)-the E. Asiatic kingdom, its delineation and characteristics. In Floristic Characteristics and Diversity of East Asian Plants; Zhang, A.L., Wu, S.K., Eds.; China Higher Education Press: Beijing, China, 1996; pp. 3–42. [Google Scholar]
- Qiu, Y.X.; Fu, C.X.; Comes, H.P. Plant molecular phylogeography in China and adjacent regions: Tracing the genetic imprints of Quaternary climate and environmental change in the world’s most diverse temperate flora. Mol. Phylogenet. Evol. 2011, 59, 225–244. [Google Scholar] [CrossRef] [PubMed]
- Lu, R.S.; Chen, Y.; Tamaki, I.; Sakaguchic, S.; Ding, Y.Q.; Takahashi, D.; Li, P.; Isaji, Y.; Chen, J.; Qiu, Y.X. Pre-quaternary diversification and glacial demographic expansions of Cardiocrinum (Liliaceae) in temperate forest biomes of Sino-Japanese Floristic Region. Mol. Phylogenet. Evol. 2020, 143, 106693. [Google Scholar] [CrossRef] [PubMed]
- Qian, H.; Ricklefs, R.E. Large-scale processes and the Asian bias in species diversity of temperate plants. Nature 2000, 407, 180–182. [Google Scholar] [CrossRef]
- Sun, H. Evolution of Arctic-Tertiary Flora in Himalayan-Hengduan Mountains. Plant Divers. 2002, 24, 1–3. [Google Scholar]
- Favre, A.; Päckert, M.; Pauls, S.U.; Jähnig, S.C.; Uhl, D.; Michalak, I.; Muellner-Riehl, A.N. The role of the uplift of the Qinghai-Tibetan Plateau for the evolution of Tibetan biotas. Biol. Rev. 2015, 90, 236–253. [Google Scholar] [CrossRef]
- Liao, Y.Y.; Gichira, A.W.; Wang, Q.F.; Chen, J.M. Molecular phylogeography of four endemic Sagittaria species (Alismataceae) in the Sino-Japanese floristic region of East Asia. Bot. J. Linn. Soc. 2016, 180, 6–20. [Google Scholar] [CrossRef]
- Sun, H.; Zhang, J.; Deng, T.; Boufford, D.E. Origins and evolution of plant diversity in the Hengduan Mountains, China. Plant Divers. 2017, 39, 161–166. [Google Scholar] [CrossRef]
- Xing, Y.W.; Ree, R.H. Uplift-driven diversification in the Hengduan Mountains, a temperate biodiversity hotspot. Proc. Natl. Acad. Sci. USA 2017, 114, 3444–3451. [Google Scholar] [CrossRef]
- He, X.; Burgess, K.S.; Gao, L.M.; Li, D.Z. Distributional responses to climate change for alpine species of Cyananthus and Primula endemic to the Himalaya-Hengduan Mountains. Plant Divers. 2019, 41, 26–32. [Google Scholar] [CrossRef]
- He, X.; Burgess, K.S.; Yang, X.F.; Ahrends, A.; Gao, L.M.; Li, D.Z. Upward elevation and northwest range shifts for alpine Meconopsis species in the Himalaya-Hengduan Mountains region. Ecol. Evol. 2019, 9, 4055–4064. [Google Scholar] [CrossRef]
- Salick, J.; Fang, Z.; Hart, R. Rapid changes in eastern Himalayan alpine flora with climate change. Am. J. Bot. 2019, 106, 520–530. [Google Scholar] [CrossRef] [PubMed]
- Yu, H.B.; Miao, S.Y.; Xie, G.W.; Guo, X.Y.; Chen, Z.; Favre, A. Contrasting floristic diversity of the Hengduan Mountains, the Himalayas and the Qinghai-Tibet Plateau sensu stricto in China. Front. Ecol. Evol. 2020, 8, 136. [Google Scholar] [CrossRef]
- Liu, B.B.; Ma, Z.Y.; Ren, C.; Hodel, R.G.J.; Sun, M.; Liu, X.Q.; Liu, G.N.; Hong, D.Y.; Zimmer, E.A.; Wen, J. Capturing single-copy nuclear genes, organellar genomes, and nuclear ribosomal DNA from deep genome skimming data for plant phylogenetics: A case study in Vitaceae. J. Syst. Evol. 2021, 59, 1124–1138. [Google Scholar] [CrossRef]
- Mu, Q.Y.; Yu, C.C.; Wang, Y.; Han, T.S.; Wang, H.; Ding, W.N.; Zhang, Q.Y.; Low, S.L.; Zheng, Q.J.; Peng, C.; et al. Comparative phylogeography of Acanthocalyx (Caprifoliaceae) reveals distinct genetic structures in the Himalaya-Hengduan Mountains. Alp. Bot. 2021, 132, 153–168. [Google Scholar] [CrossRef]
- Yang, M.X.; Werth, S.; Wang, L.S.; Scheidegger, C. Phylogeographic analyses of an epiphytic foliose lichen show multiple dispersal events westward from the Hengduan Mountains of Yunnan into the Himalayas. Ecol. Evol. 2022, 12, e9308. [Google Scholar] [CrossRef] [PubMed]
- Wang, Q.; Hong, D.Y. Understanding the plant diversity on the roof of the world—A brief review of flora of Pan-Himalaya. Innovation 2022, 3, 100215. [Google Scholar] [CrossRef]
- Cheng, M. The floristic relationship between Eastern China and Japan. Acta Phytotax. Sin. 1984, 22, 1–5. [Google Scholar]
- Mitsui, Y.; Chen, S.T.; Zhou, Z.K.; Peng, C.I.; Deng, Y.F.; Setoguchi, H. Phylogeny and biogeography of the genus Ainsliaea (Asteraceae) in the Sino-Japanese region based on nuclear rDNA and plastid DNA sequence data. Ann. Bot. 2008, 101, 111–124. [Google Scholar] [CrossRef]
- Bai, W.N.; Wang, W.T.; Zhang, D.Y. Phylogeographic breaks within Asian butternuts indicate the existence of a phytogeographic divide in East Asia. New Phytol. 2016, 209, 1757–1772. [Google Scholar] [CrossRef]
- Kubota, Y.; Kusumoto, B.; Shiono, T.; Tanaka, T. Phylogenetic properties of Tertiary relict flora in the East Asian continental islands: Imprint of climatic niche conservatism and in situ diversification. Ecography 2017, 40, 436–447. [Google Scholar] [CrossRef]
- Deng, M.; Jiang, X.L.; Hipp, A.L.; Manos, P.S.; Hahn, M. Phylogeny and biogeography of East Asian evergreen oaks (Quercus section Cyclobalanopsis; Fagaceae): Insights into the Cenozoic history of evergreen broad-leaved forests in subtropical Asia. Mol. Phylogenet. Evol. 2018, 119, 170–181. [Google Scholar] [CrossRef] [PubMed]
- Naciri, Y.; Christe, C.; Bétrisey, S.; Song, Y.G.; Deng, M.; Garfì, G.; Kozlowski, G. Species delimitation in the East Asian species of the relict tree genus Zelkova (Ulmaceae): A complex history of diversification and admixture among species. Mol. Phylogenet. Evol. 2019, 134, 172–185. [Google Scholar] [CrossRef] [PubMed]
- Zhao, K.K.; Landrein, S.; Barrett, R.L.; Sakaguchi, S.; Maki, M.; Mu, W.X.; Yang, T.; Zhu, Z.X.; Liu, H.; Wang, H.F. Phylogeographic analysis and genetic structure of an endemic Sino-Japanese disjunctive genus Diabelia (Caprifoliaceae). Front. Plant Sci. 2019, 10, 913. [Google Scholar] [CrossRef] [PubMed]
- Wang, H.X.; Moore, M.J.; Barrett, R.L.; Landrein, S.; Sakaguchi, S.; Maki, M.; Wen, J.; Wang, H.F. Plastome phylogenomic insights into the Sino-Japanese biogeography of Diabelia (Caprifoliaceae). J. Syst. Evol. 2020, 58, 972–987. [Google Scholar] [CrossRef]
- Takahashi, D.; Sakaguchi, S.; Feng, Y.; Isagi, Y.; Qiu, Y.X.; Li, P.; Lu, R.S.; Lu, C.T.; Chung, S.W.; Lin, Y.S.; et al. Geographic and subsequent biotic isolations led to a diversity anomaly of section Heterotropa (genus Asarum: Aristolochiaceae) in insular versus continental regions of the Sino-Japanese Floristic Region. J. Biogeogr. 2021, 48, 1917–1929. [Google Scholar] [CrossRef]
- Ota, H. Geographic patterns of endemism and speciation in amphibians and reptiles of the Ryukyu Archipelago, Japan, with special reference to their paleogeographical implications. Res. Pop. Ecol. 1998, 40, 189–204. [Google Scholar] [CrossRef]
- Axelrod, D.I.; Al-Shehbaz, I.A.; Raven, P.H. History of the modern flora of China. In Floristic Characteristics and Diversity of East Asian Plants; Zhang, A.L., Wu, S.K., Eds.; China Higher Education Press: Beijing, China, 1996; pp. 43–55. [Google Scholar]
- Manchester, S.R.; Chen, Z.D.; Lu, A.M.; Uemura, K. Eastern Asian endemic seed plant genera and their paleogeographic history throughout the Northern Hemisphere. J. Syst. Evol. 2009, 47, 1–42. [Google Scholar] [CrossRef]
- Irizuki, T.; Kusumoto, M.; Ishida, K.; Tanaka, Y. Sea-level changes and water structures between 3.5 and 2.8 Ma in the central part of the Japan Sea Borderland: Analyses of fossil Ostracoda from the Pliocene Kuwae Formation, central Japan. Palaeogeogr. Palaeoclimatol. Palaeoecol. 2007, 245, 421–443. [Google Scholar] [CrossRef]
- Cao, Y.N.; Wang, I.J.; Chen, L.Y.; Ding, Y.Q.; Liu, L.X.; Qiu, Y.X. Inferring spatial patterns and drivers of population divergence of Neolitsea sericea (Lauraceae), based on molecular phylogeography and landscape genomics. Mol. Phylogenet. Evol. 2018, 126, 162–172. [Google Scholar] [CrossRef]
- Ujiie, H. Nature of Okinawa—Geomorphology and Geology; Hirugisha: Naha-shi, Japan, 1990. [Google Scholar]
- Qiu, Y.X.; Guan, B.C.; Fu, C.X.; Comes, H.P. Did glacials and/or interglacials promote allopatric incipient speciation in East Asian temperate plants? Phylogeographic and coalescent analyses on refugial isolation and divergence in Dysosma versipellis. Mol. Phylogenet. Evol. 2009, 51, 281–293. [Google Scholar] [CrossRef]
- Sakaguchi, S.; Qiu, Y.X.; Liu, Y.H.; Qi, X.S.; Kim, S.H.; Han, J.; Takeuchi, Y.; Worth, J.R.P.; Yamasaki, M.; Sakurai, S.; et al. Climate oscillation during the Quaternary associated with landscape heterogeneity promoted allopatric lineage divergence of a temperate tree Kalopanax septemlobus (Araliaceae) in East Asia. Mol. Ecol. 2012, 21, 3823–3838. [Google Scholar] [CrossRef] [PubMed]
- Harris, N. The elevation history of the Tibetan Plateau and its implications for the Asian monsoon. Palaeogeogr. Palaeoclimatol. Palaeoecol. 2006, 241, 4–15. [Google Scholar] [CrossRef]
- An, Z.S.; Kutzbach, J.E.; Prell, W.L.; Porter, S.C. Evolution of Asian monsoons and phased uplift of the Himalayan-Tibetan Plateau since Late Miocene times. Nature 2001, 411, 62–66. [Google Scholar] [CrossRef]
- Li, F.J.; Rousseau, D.D.; Wu, N.Q.; Hao, Q.Z.; Pei, Y.P. Late Neogene evolution of the East Asian monsoon revealed by terrestrial mollusk record in Western Chinese Loess Plateau: From winter to summer dominated sub-regime. Earth Planet. Sci. Lett. 2008, 274, 439–447. [Google Scholar] [CrossRef]
- Hewitt, G.M. Genetic consequences of climatic oscillations in the Quaternary. Philos. Trans. Roy. Soc. Lond. Ser. B: Biol. Sci. 2004, 359, 183–195. [Google Scholar] [CrossRef]
- Tamura, M. Clematis L. In Die Natürlichen Pflanzenfamilien, 2nd ed.; Heipko, P., Ed.; Duncker & Humblot: Berlin, Germany, 1995; Volume 17a, pp. 368–387. [Google Scholar]
- Wang, W.T.; Li, L.Q. A new system of classification of the genus Clematis (Ranunculaceae). Acta Phytotax. Sin. 2005, 43, 431–488. [Google Scholar]
- Xie, L.; Wen, J.; Li, L.Q. Phylogenetic analyses of Clematis (Ranunculaceae) based on sequences of nuclear ribosomal ITS and three plastid regions. Syst. Bot. 2011, 36, 907–921. [Google Scholar] [CrossRef]
- He, J.; Lyu, R.D.; Luo, Y.K.; Yao, M.; Xiao, J.M.; Xie, L.; Wen, J.; Pei, L.Y.; Yan, S.X.; Cheng, J.; et al. An updated phylogenetic and biogeographic analysis based on genome skimming data reveals convergent evolution of shrubby habit in Clematis in the Pliocene and Pleistocene. Mol. Phylogenet. Evol. 2021, 164, 107259. [Google Scholar] [CrossRef]
- Wang, W.T.; Xie, L. A revision of Clematis sect. Tubulosae (Ranunculaceae). Acta Phytotax. Sin. 2007, 45, 425–457. [Google Scholar] [CrossRef]
- Makino, T. Obserbations on the Flora of Japan. Bot. Mag. (Tokyo) 1907, 21, 86–88. [Google Scholar] [CrossRef]
- Lyu, R.D.; He, J.; Luo, Y.K.; Lin, L.L.; Yao, M.; Cheng, J.; Xie, L.; Pei, L.Y.; Yan, S.X.; Li, L.Q. Natural hybrid origin of the controversial “species” Clematis × pinnata (Ranunculaceae) based on multidisciplinary evidence. Front. Plant Sci. 2021, 12, 745988. [Google Scholar] [CrossRef]
- Xie, L.; Shi, J.H.; Li, L.Q. Identity of Clematis tatarinowii and C. pinnata var. ternatifolia (Ranunculaceae). Ann. Bot. Fenn. 2005, 42, 305–308. [Google Scholar]
- Miikeda, O.; Kita, K.; Handa, T.; Yukawa, T. Phylogenetic relationships of Clematis (Ranunculaceae) based on chloroplast and nuclear DNA sequences. Bot. J. Linn. Soc. 2006, 152, 153–168. [Google Scholar] [CrossRef]
- Lehtonen, S.; Christenhusz, M.J.; Falck, D. Sensitive phylogenetics of Clematis and its position in Ranunculaceae. Bot. J. Linn. Soc. 2016, 182, 825–867. [Google Scholar] [CrossRef] [Green Version]
- Jiang, N.; Zhou, Z.; Yang, J.B.; Zhang, S.D.; Guan, K.Y.; Tan, Y.H.; Yu, W.B. Phylogenetic reassessment of tribe Anemoneae (Ranunculaceae): Non-monophyly of Anemone s. l. revealed by plastid datasets. PLoS ONE 2017, 12, e0174792. [Google Scholar] [CrossRef] [PubMed]
- Zimmer, E.A.; Wen, J. Using nuclear gene data for plant phylogenetics: Progress and prospects II. Next-gen approaches. J. Syst. Evol. 2015, 53, 371–379. [Google Scholar] [CrossRef]
- Wen, J.; Harris, A.J.; Ickert-Bond, S.M.; Dikow, R.; Wurdack, K.; Zimmer, E.A. Developing integrative systematics in the informatics and genomic era, and calling for a global Biodiversity Cyberbank. J. Syst. Evol. 2017, 55, 308–321. [Google Scholar] [CrossRef]
- McKain, M.R.; Johnson, M.G.; Uribe-Convers, S.; Eaton, D.; Yang, Y. Practical considerations for plant phylogenomics. Appl. Plant Sci. 2018, 6, e1038. [Google Scholar] [CrossRef]
- Yu, X.Q.; Yang, D.; Guo, C.; Gao, L.M. Plant phylogenomics based on genome-partitioning strategies: Progress and prospects. Plant Divers. 2018, 40, 158–164. [Google Scholar] [CrossRef]
- Valcárcel, V.; Wen, J. Chloroplast phylogenomic data support Eocene amphi-Pacific early radiation for the Asian Palmate core Araliaceae. J. Syst. Evol. 2019, 57, 547–560. [Google Scholar] [CrossRef]
- Watson, L.E.; Siniscalchi, C.M.; Mandel, J. Phylogenomics of the hyperdiverse daisy tribes: Anthemideae, Astereae, Calenduleae, Gnaphalieae, and Senecioneae. J. Syst. Evol. 2020, 58, 841–852. [Google Scholar] [CrossRef]
- Thode, V.A.; Lohmann, L.G.; Sanmartín, I. Evaluating character partitioning and molecular models in plastid phylogenomics at low taxonomic levels: A case study using Amphilophium (Bignonieae, Bignoniaceae). J. Syst. Evol. 2020, 58, 1071–1089. [Google Scholar] [CrossRef]
- Wikström, N.; Bremer, B.; Rydin, C. Conflicting phylogenetic signals in genomic data of the coffee family (Rubiaceae). J. Syst. Evol. 2020, 58, 440–460. [Google Scholar] [CrossRef]
- Olofsson, J.K.; Cantera, I.; Van de Paer, C.; Hong-Wa, C.; Zedane, L.; Dunning, L.T.; Alberti, A.; Christin, P.; Besnard, G. Phylogenomics using low-depth whole genome sequencing: A case study with the olive tribe. Mol. Ecol. Resour. 2019, 19, 877–892. [Google Scholar] [CrossRef] [PubMed]
- Xiao, J.M.; Lyu, R.D.; He, J.; Li, M.Y.; Cheng, J.; Xie, L. Genome-partitioning strategy, plastid and nuclear phylogenomic discordance, and its evolutionary implications of Clematis (Ranunculaceae). Front. Plant Sci. 2022, 13, 1059379. [Google Scholar] [CrossRef]
- Yuan, T.; Wang, L.Y.; Roh, M.S. Confirmation of Clematis hybrids using molecular markers. Sci. Hort. 2010, 125, 136–145. [Google Scholar] [CrossRef]
- Zachos, J.C.; Dickens, G.R.; Zeebe, R.E. An early Cenozoic perspective on greenhouse warming and carbon-cycle dynamics. Nature 2008, 451, 279–283. [Google Scholar] [CrossRef]
- Zachos, J.; Pagani, M.; Sloan, L.; Thomas, E.; Billups, K. Trends, rhythms, and aberrations in global climate 65 Ma to present. Science 2001, 292, 686–693. [Google Scholar] [CrossRef]
- Matzke, N.J. Model selection in historical biogeography reveals that founder-event speciation is a crucial process in island clades. Syst. Biol. 2014, 63, 951–970. [Google Scholar] [CrossRef]
- Handel-Mazzetti, H. Plantae Sinenses a Dre. H. Smith annis 1921–1922, 1924 et 1934 lectae. 33. Handel-Mazzetti, Ranunculaceae. Acta Horti Gotob. 1939, 13, 37–219. [Google Scholar]
- Chang, M.C.; Fang, M.Y.; Ting, C.T.; Ling, P.P. Archiclematis, Clematis. In Flora Reipublicae Popularis Sinicae; Science Press: Beijing, China, 1980; Volume 28, pp. 74–235. [Google Scholar]
- Johnson, M. Slaktet Klematis; M. Johnsons Plantskola AB: Södertälje, Swedish, 1997. [Google Scholar]
- Yan, S.X.; Liu, H.J.; Lin, L.L.; Liao, S.; Li, J.Y.; Pei, L.Y.; Xie, L. Taxonomic status of Clematis acerifolia var. elobata, based on molecular evidence. Phytotaxa 2016, 268, 209–219. [Google Scholar] [CrossRef]
- Liu, B.B.; Campbell, C.S.; Hong, D.Y.; Wen, J. Phylogenetic relationships and chloroplast capture in the Amelanchier-Malacomeles-Peraphyllum clade (Maleae, Rosaceae): Evidence from chloroplast genome and nuclear ribosomal DNA data using genome skimming. Mol. Phylogenet. Evol. 2020, 147, 106784. [Google Scholar] [CrossRef] [PubMed]
- Wang, H.X.; Morales-Briones, D.F.; Moore, M.J.; Wen, J.; Wang, H.F. A phylogenomic perspective on gene tree conflict and character evolution in Caprifoliaceae using target enrichment data, with Zabelioideae recognized as a new subfamily. J. Syst. Evol. 2021, 59, 897–914. [Google Scholar] [CrossRef]
- He, J.; Lyu, R.D.; Luo, Y.K.; Xiao, J.M.; Xie, L.; Wen, J.; Li, W.H.; Pei, L.Y.; Cheng, J. A phylotranscriptome study using silica gel-dried leaf tissues produces an updated robust phylogeny of Ranunculaceae. Mol. Phylogenet. Evol. 2022, 174, 107545. [Google Scholar] [CrossRef] [PubMed]
- Rehder, A.; Wilson, E.H. Clematis. In Plantae Wilsonianae; Sargent, C.S., Ed.; The University Press: Cambridge, UK, 1913; Volume 1, pp. 319–343. [Google Scholar]
- Xie, L.; Li, L.Q. Variation of pollen morphology, and its implications in the phylogeny of Clematis (Ranunculaceae). Plant Syst. Evol. 2012, 298, 1437–1453. [Google Scholar] [CrossRef]
- Tiffney, B.H. Perspectives on the origin of the floristic similarity between eastern Asia and eastern North America. J. Arnold Arbor. 1985, 66, 73–94. [Google Scholar] [CrossRef]
- Wang, H.L.; Lu, H.Y.; Zhao, L.; Zhang, H.Y.; Lei, F.; Wang, Y.C. Asian monsoon rainfall variation during the Pliocene forced by global temperature change. Nat. Commun. 2019, 10, 5272. [Google Scholar] [CrossRef]
- Sun, X.; Wang, P. How old is the Asian monsoon system?—Palaeobotanical records from China. Palaeogeogr. Palaeoclim. Palaeoeco. 2005, 222, 181–222. [Google Scholar] [CrossRef]
- Willeit, M.; Ganopolski, A.; Calov, R.; Brovkin, V. Mid-Pleistocene transition in glacial cycles explained by declining CO2 and regolith removal. Sci. Adv. 2019, 5, eaav7337. [Google Scholar] [CrossRef]
- Momohara, A. Floral and paleoenvironmental history from the late Pliocene to middle Pleistocene in and around central Japan. Palaeogeogr. Palaeoclim. Palaeoeco. 1994, 108, 281–293. [Google Scholar] [CrossRef]
- Svenning, J.C. Deterministic Plio-Pleistocene extinctions in the European cool-temperate tree flora. Ecol. Lett. 2003, 6, 646–653. [Google Scholar] [CrossRef]
- Martinetto, E.; Momohara, A.; Bizzarri, R.; Baldanza, A.; Delfino, M.; Esu, D.; Sardella, R. Late persistence and deterministic extinction of “humid thermophilous plant taxa of East Asian affinity” (HUTEA) in southern Europe. Palaeogeogr. Palaeoclim. Palaeoeco. 2017, 467, 211–231. [Google Scholar] [CrossRef]
- Petit, J.R.; Duminil, J.; Fineschi, S.; Hampe, A.; Salvini, D.; Vendramin, G.G. Comparative organization of chloroplast, mitochondrial and nuclear diversity in plant populations. Mol. Ecol. 2005, 14, 689–701. [Google Scholar] [CrossRef] [PubMed]
- Xiang, Q.Y.; Zhang, W.H.; Ricklefs, R.E.; Qian, H.; Chen, Z.D.; Wen, J.; Li, J.H. Regional differences in rates of plant speciation and molecular evolution: A comparison between eastern Asia and eastern North America. Evolution 2004, 58, 2175–2184. [Google Scholar] [CrossRef]
- Kubota, Y.; Shiono, T.; Kusumoto, B. Role of climate and geohistorical factors in driving plant richness patterns and endemicity on the east Asian continental islands. Ecography 2015, 38, 639–648. [Google Scholar] [CrossRef]
- Yang, L.Q.; Hu, H.Y.; Xie, C.; Lai, S.P.; Yang, M.; He, X.J.; Zhou, S.D. Molecular phylogeny, biogeography and ecological niche modelling of Cardiocrinum (Liliaceae): Insights into the evolutionary history of endemic genera distributed across the Sino-Japanese floristic region. Ann. Bot. 2017, 119, 59–72. [Google Scholar] [CrossRef]
- Ying, T.S.; Hsu, K.S. An analysis of the flora of seed plants of Taiwan, China: Its nature, characteristics, and relations with the flora of the mainland. Acta Phytotax. Sin. 2002, 40, 1–51. [Google Scholar]
- Ye, J.F.; Chen, Z.D.; Liu, B.; Qin, H.; Yang, Y. Disjunct distribution of vascular plants between southwestern area and Taiwan area in China. Biodivers. Sci. 2012, 20, 482–494. [Google Scholar] [CrossRef]
- Dan, T.; Mitsui, Y.; Ikeda, H.; Isagi, Y.; Setoguchi, H. Isolation and characterization of microsatellite loci in Shortia rotundifolia (Diapensiaceae), an endangered relict plant on the Ryukyu Islands and Taiwan. Conserv. Genet. 2009, 10, 507–509. [Google Scholar] [CrossRef]
- Chiang, T.Y.; Schaal, B.A. Phylogeography of plants in Taiwan and the Ryukyu Archipelago. Taxon 2006, 55, 31–41. [Google Scholar] [CrossRef]
- Li, J.L.; Wang, S.; Yu, J.; Wang, L.; Zhou, S.L. A modified CTAB protocol for plant DNA extraction. Chin. Bull. Bot. 2013, 48, 72–78. [Google Scholar] [CrossRef]
- He, J.; Yao, M.; Lyu, R.D.; Lin, L.L.; Liu, H.J.; Pei, L.Y.; Yan, S.X.; Xie, L.; Cheng, J. Structural variation of the complete chloroplast genome and plastid phylogenomics of the genus Asteropyrum (Ranunculaceae). Sci. Rep. 2019, 9, 15285. [Google Scholar] [CrossRef] [PubMed]
- Kearse, M.; Moir, R.; Wilson, A.; Stones-Havas, S.; Cheung, M.; Sturrock, S.; Buxton, S.; Cooper, A.; Markowitz, S.; Duran, C.; et al. Geneious Basic: An integrated and extendable desktop software platform for the organization and analysis of sequence data. Bioinformatics 2012, 28, 1647–1649. [Google Scholar] [CrossRef] [PubMed]
- Qu, X.J.; Moore, M.J.; Li, D.Z.; Yi, T.S. PGA: A software package for rapid, accurate, and flexible batch annotation of plastomes. Plant Methods 2019, 15, 50. [Google Scholar] [CrossRef]
- Lohse, M.; Drechsel, O.; Kahlau, S.; Bock, R. OrganellarGenomeDRAW—A suite of tools for generating physical maps of plastid and mitochondrial genomes and visualizing expression data sets. Nucleic Acids Res. 2013, 41, W575–W581. [Google Scholar] [CrossRef]
- Drezen, E.; Rizk, G.; Chikhi, R.; Deltel, C.; Lemaitre, C.; Peterlongo, P.; Lavenier, D. GATB: Genome assembly & analysis tool box. Bioinformatics 2014, 30, 2959–2961. [Google Scholar] [CrossRef] [Green Version]
- Chen, N. Using Repeat Masker to identify repetitive elements in genomic sequences. Current Protoc. Bioinform. 2004, 5, 4.10.1–4.10.14. [Google Scholar] [CrossRef]
- Duvall, M.R.; Burke, S.V.; Clark, D.C. Plastome phylogenomics of Poaceae: Alternate topologies depend on alignment gaps. Bot. J. Linn. Soc. 2020, 192, 9–20. [Google Scholar] [CrossRef]
- Liu, H.J.; Ding, C.H.; He, J.; Chen, J.; Pei, L.Y.; Xie, L. Complete chloroplast genomes of Archiclematis, Naravelia and Clematis (Ranunculaceae), and their phylogenetic implications. Phytotaxa 2018, 343, 214–226. [Google Scholar] [CrossRef]
- Katoh, K.; Standley, D.M. MAFFT multiple sequence alignment software version 7: Improvements in performance and usability. Mol. Biol. Evol. 2013, 30, 772–780. [Google Scholar] [CrossRef]
- Darriba, D.; Taboada, G.L.; Doallo, R.; Posada, D. jModelTest 2: More models, new heuristics and parallel computing. Nat. Methods 2012, 9, 772. [Google Scholar] [CrossRef] [PubMed]
- Stamatakis, A. RAxML version 8: A tool for phylogenetic analysis and post-analysis of large phylogenies. Bioinformatics 2014, 30, 1312–1313. [Google Scholar] [CrossRef] [PubMed]
- Ronquist, F.; Huelsenbeck, J.P. MrBayes 3: Bayesian phylogenetic inference under mixed models. Bioinformatics 2003, 19, 1572–1574. [Google Scholar] [CrossRef] [PubMed]
- Page, A.J.; Taylor, B.; Delaney, A.J.; Soares, J.; Seemann, T.; Keane, J.A.; Harris, S.R. SNP-sites: Rapid efficient extraction of SNPs from multi-FASTA alignments. Microb. Genom. 2016, 2, e000056. [Google Scholar] [CrossRef] [PubMed]
- Bouckaert, R.; Heled, J.; Kühnert, D.; Vaughan, T.; Wu, C.H.; Xie, D.; Suchard, M.A.; Rambaut, A.; Drummond, A.J. BEAST 2: A software platform for Bayesian evolutionary analysis. PLoS Comput. Biol. 2014, 10, e1003537. [Google Scholar] [CrossRef] [PubMed]
- Rambaut, A.; Drummond, A.J.; Xie, D.; Baele, G.; Suchard, M.A. Posterior summarization in Bayesian phylogenetics using Tracer 1.7. Syst. Biol. 2018, 67, 901–904. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yu, Y.; Harris, A.J.; Blair, C.; He, X.J. RASP (Reconstruct Ancestral State in Phylogenies): A tool for historical biogeography. Mol. Phylogenet. Evol. 2015, 87, 46–49. [Google Scholar] [CrossRef]
- Maddison, W.P.; Maddison, D.R. Mesquite: A Modular System for Evolutionary Analysis. Available online: http://mesquiteproject.org (accessed on 12 January 2023).





Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Lyu, R.; Xiao, J.; Li, M.; Luo, Y.; He, J.; Cheng, J.; Xie, L. Phylogeny and Historical Biogeography of the East Asian Clematis Group, Sect. Tubulosae, Inferred from Phylogenomic Data. Int. J. Mol. Sci. 2023, 24, 3056. https://doi.org/10.3390/ijms24033056
Lyu R, Xiao J, Li M, Luo Y, He J, Cheng J, Xie L. Phylogeny and Historical Biogeography of the East Asian Clematis Group, Sect. Tubulosae, Inferred from Phylogenomic Data. International Journal of Molecular Sciences. 2023; 24(3):3056. https://doi.org/10.3390/ijms24033056
Chicago/Turabian StyleLyu, Rudan, Jiamin Xiao, Mingyang Li, Yike Luo, Jian He, Jin Cheng, and Lei Xie. 2023. "Phylogeny and Historical Biogeography of the East Asian Clematis Group, Sect. Tubulosae, Inferred from Phylogenomic Data" International Journal of Molecular Sciences 24, no. 3: 3056. https://doi.org/10.3390/ijms24033056