
Seasonal Changes in Serum Metabolites in Multiple Sclerosis Relapse
 , , , , , and
, , , , , and
Abstract
:1. Introduction
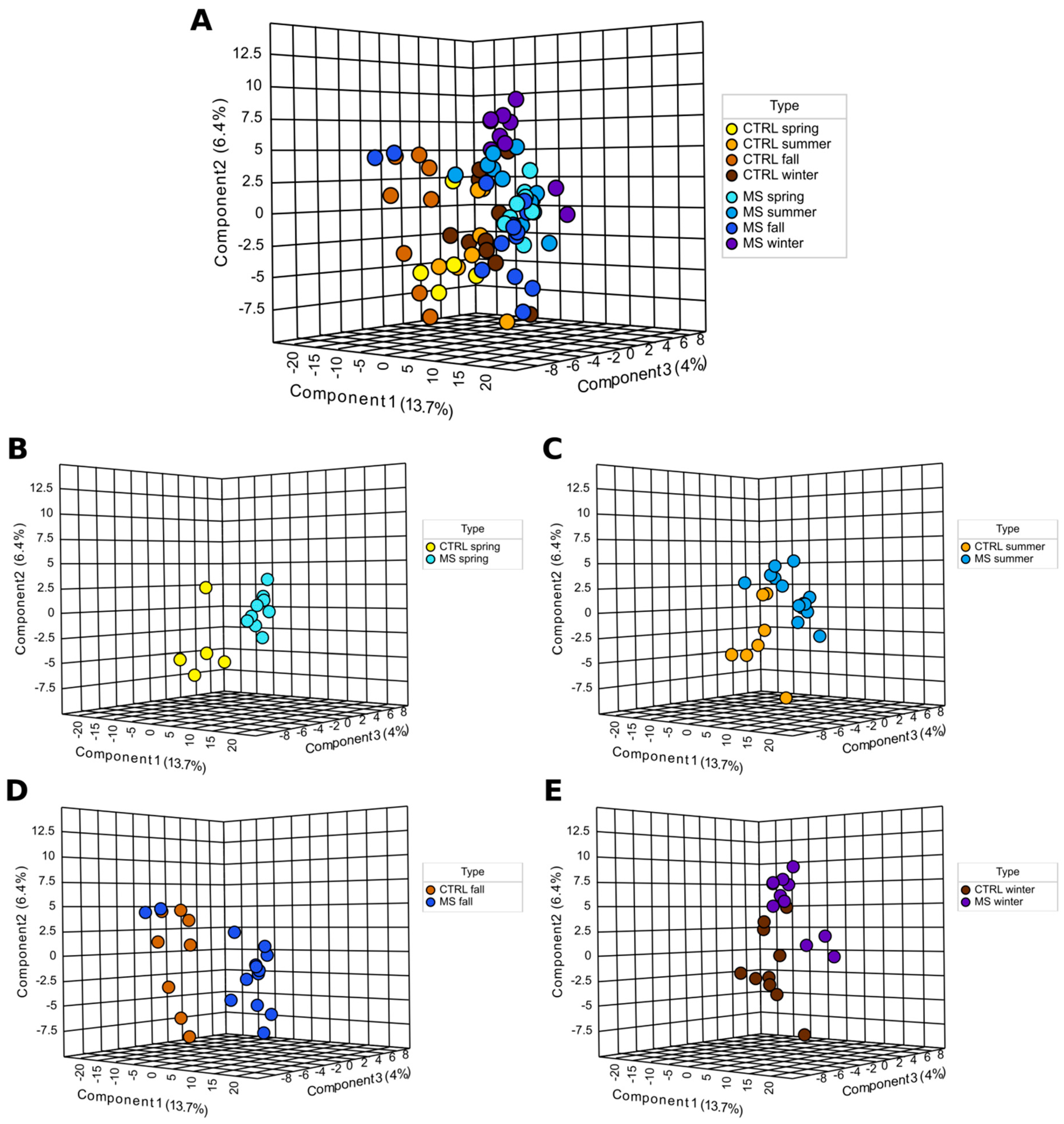
2. Results
2.1. Metabolites Analyses
Hierarchical Cluster Analysis
2.2. Analysis of Metabolite Levels in MS Based on the Season
2.3. Metabolic Pathways Affected in MS
3. Discussion
4. Materials and Methods
4.1. Study Subjects, Samples
4.2. Metabolome Analysis
4.3. Statistical Analysis
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Lublin, F.D.; Reingold, S.C.; Cohen, J.A.; Cutter, G.R.; Sorensen, P.S.; Thompson, A.J.; Wolinsky, J.S.; Balcer, L.J.; Banwell, B.; Barkhof, F.; et al. Defining the clinical course of multiple sclerosis: The 2013 revisions. Neurology 2014, 83, 278–286. [Google Scholar] [CrossRef] [PubMed]
- Goldenberg, M.M. Multiple sclerosis review. Pharm. Ther. 2012, 37, 175–184. [Google Scholar]
- Comi, G.; Radaelli, M.; Soelberg Sorensen, P. Evolving concepts in the treatment of relapsing multiple sclerosis. Lancet 2017, 389, 1347–1356. [Google Scholar] [CrossRef] [PubMed]
- Langer-Gould, A.; Popat, R.A.; Huang, S.M.; Cobb, K.; Fontoura, P.; Gould, M.K.; Nelson, L.M. Clinical and demographic predictors of long-term disability in patients with relapsing-remitting multiple sclerosis: A systematic review. Arch. Neurol. 2006, 63, 1686–1691. [Google Scholar] [CrossRef] [PubMed]
- Simpson, S., Jr.; Taylor, B.; Blizzard, L.; Ponsonby, A.L.; Pittas, F.; Tremlett, H.; Dwyer, T.; Gies, P.; van der Mei, I. Higher 25-hydroxyvitamin D is associated with lower relapse risk in multiple sclerosis. Ann. Neurol. 2010, 68, 193–203. [Google Scholar] [CrossRef] [PubMed]
- Stoessel, D.; Stellmann, J.P.; Willing, A.; Behrens, B.; Rosenkranz, S.C.; Hodecker, S.C.; Sturner, K.H.; Reinhardt, S.; Fleischer, S.; Deuschle, C.; et al. Metabolomic Profiles for Primary Progressive Multiple Sclerosis Stratification and Disease Course Monitoring. Front. Hum. Neurosci. 2018, 12, 226. [Google Scholar] [CrossRef]
- Lim, C.K.; Bilgin, A.; Lovejoy, D.B.; Tan, V.; Bustamante, S.; Taylor, B.V.; Bessede, A.; Brew, B.J.; Guillemin, G.J. Kynurenine pathway metabolomics predicts and provides mechanistic insight into multiple sclerosis progression. Sci. Rep. 2017, 7, 41473. [Google Scholar] [CrossRef]
- Meier, D.S.; Balashov, K.E.; Healy, B.; Weiner, H.L.; Guttmann, C.R. Seasonal prevalence of MS disease activity. Neurology 2010, 75, 799–806. [Google Scholar] [CrossRef]
- Salvi, F.; Bartolomei, I.; Smolensky, M.H.; Lorusso, A.; Barbarossa, E.; Malagoni, A.M.; Zamboni, P.; Manfredini, R. A seasonal periodicity in relapses of multiple sclerosis? A single-center, population-based, preliminary study conducted in Bologna, Italy. BMC Neurol. 2010, 10, 105. [Google Scholar] [CrossRef]
- Harding, K.; Tilling, K.; MacIver, C.; Willis, M.; Joseph, F.; Ingram, G.; Hirst, C.; Wardle, M.; Pickersgill, T.; Ben-Shlomo, Y.; et al. Seasonal variation in multiple sclerosis relapse. J. Neurol. 2017, 264, 1059–1067. [Google Scholar] [CrossRef]
- Ma, J.; Zhang, X. The relationship between season/latitude and multiple sclerosis. Chin. J. Intern. Med. 2015, 54, 945–948. [Google Scholar]
- O’Reilly, M.A.; O’Reilly, P.M. Temporal influences on relapses of multiple sclerosis. Eur. Neurol. 1991, 31, 391–395. [Google Scholar] [CrossRef] [PubMed]
- Fonseca, A.C.; Costa, J.; Cordeiro, C.; Geraldes, R.; de Sa, J. Influence of climatic factors in the incidence of multiple sclerosis relapses in a Portuguese population. Eur. J. Neurol. 2009, 16, 537–539. [Google Scholar] [CrossRef] [PubMed]
- Saaroni, H.; Sigal, A.; Lejbkowicz, I.; Miller, A. Mediterranean weather conditions and exacerbations of multiple sclerosis. Neuroepidemiology 2010, 35, 142–151. [Google Scholar] [CrossRef]
- Damasceno, A.; Von Glehn, F.; de Deus-Silva, L.; Damasceno, B.P. Monthly variation of multiple sclerosis activity in the southern hemisphere: Analysis from 996 relapses in Brazil. Eur. J. Neurol. 2012, 19, 660–662. [Google Scholar] [CrossRef] [PubMed]
- Cross, A.H.; Parks, B.J. The four seasons of multiple sclerosis. Neurology 2010, 75, 762–763. [Google Scholar] [CrossRef]
- Wurtman, R. Multiple Sclerosis, Melatonin, and Neurobehavioral Diseases. Front. Endocrinol. 2017, 8, 280. [Google Scholar] [CrossRef]
- Farez, M.F.; Mascanfroni, I.D.; Mendez-Huergo, S.P.; Yeste, A.; Murugaiyan, G.; Garo, L.P.; Balbuena Aguirre, M.E.; Patel, B.; Ysrraelit, M.C.; Zhu, C.; et al. Melatonin Contributes to the Seasonality of Multiple Sclerosis Relapses. Cell 2015, 162, 1338–1352. [Google Scholar] [CrossRef]
- Hartl, C.; Obermeier, V.; Gerdes, L.A.; Brugel, M.; von Kries, R.; Kumpfel, T. Seasonal variations of 25-OH vitamin D serum levels are associated with clinical disease activity in multiple sclerosis patients. J. Neurol. Sci. 2017, 375, 160–164. [Google Scholar] [CrossRef]
- Fiorini, A.; Koudriavtseva, T.; Bucaj, E.; Coccia, R.; Foppoli, C.; Giorgi, A.; Schinina, M.E.; Di Domenico, F.; De Marco, F.; Perluigi, M. Involvement of oxidative stress in occurrence of relapses in multiple sclerosis: The spectrum of oxidatively modified serum proteins detected by proteomics and redox proteomics analysis. PLoS ONE 2013, 8, e65184. [Google Scholar] [CrossRef]
- Tavazzi, B.; Batocchi, A.P.; Amorini, A.M.; Nociti, V.; D’Urso, S.; Longo, S.; Gullotta, S.; Picardi, M.; Lazzarino, G. Serum metabolic profile in multiple sclerosis patients. Mult. Scler. Int. 2011, 2011, 167156. [Google Scholar] [CrossRef] [PubMed]
- Tremlett, H.; Zhu, F.; Ascherio, A.; Munger, K.L. Sun exposure over the life course and associations with multiple sclerosis. Neurology 2018, 90, e1191–e1199. [Google Scholar] [CrossRef]
- Ostkamp, P.; Salmen, A.; Pignolet, B.; Gorlich, D.; Andlauer, T.F.M.; Schulte-Mecklenbeck, A.; Gonzalez-Escamilla, G.; Bucciarelli, F.; Gennero, I.; Breuer, J.; et al. Sunlight exposure exerts immunomodulatory effects to reduce multiple sclerosis severity. Proc. Natl. Acad. Sci. USA 2021, 118, e2018457118. [Google Scholar] [CrossRef] [PubMed]
- Spelman, T.; Gray, O.; Trojano, M.; Petersen, T.; Izquierdo, G.; Lugaresi, A.; Hupperts, R.; Bergamaschi, R.; Duquette, P.; Grammond, P.; et al. Seasonal variation of relapse rate in multiple sclerosis is latitude dependent. Ann. Neurol. 2014, 76, 880–890. [Google Scholar] [CrossRef] [PubMed]
- Zahoor, I.; Suhail, H.; Datta, I.; Ahmed, M.E.; Poisson, L.M.; Waters, J.; Rashid, F.; Bin, R.; Singh, J.; Cerghet, M.; et al. Blood-based untargeted metabolomics in relapsing-remitting multiple sclerosis revealed the testable therapeutic target. Proc. Natl. Acad. Sci. USA 2022, 119, e2123265119. [Google Scholar] [CrossRef] [PubMed]
- Loy, B.D.; Fling, B.W.; Sage, K.M.; Spain, R.I.; Horak, F.B. Serum Histidine is Lower in Fatigued Women with Multiple Sclerosis. Fatigue 2019, 7, 69–80. [Google Scholar] [CrossRef]
- Fitzgerald, K.C.; Smith, M.D.; Kim, S.; Sotirchos, E.S.; Kornberg, M.D.; Douglas, M.; Nourbakhsh, B.; Graves, J.; Rattan, R.; Poisson, L.; et al. Multi-omic evaluation of metabolic alterations in multiple sclerosis identifies shifts in aromatic amino acid metabolism. Cell Rep. Med. 2021, 2, 100424. [Google Scholar] [CrossRef]
- Haas, H.L.; Sergeeva, O.A.; Selbach, O. Histamine in the nervous system. Physiol. Rev. 2008, 88, 1183–1241. [Google Scholar] [CrossRef]
- Jutel, M.; Blaser, K.; Akdis, C.A. Histamine in allergic inflammation and immune modulation. Int. Arch. Allergy Immunol. 2005, 137, 82–92. [Google Scholar] [CrossRef]
- Pedotti, R.; De Voss, J.J.; Steinman, L.; Galli, S.J. Involvement of both ‘allergic’ and ‘autoimmune’ mechanisms in EAE, MS and other autoimmune diseases. Trends Immunol. 2003, 24, 479–484. [Google Scholar] [CrossRef]
- Teuscher, C.; Poynter, M.E.; Offner, H.; Zamora, A.; Watanabe, T.; Fillmore, P.D.; Zachary, J.F.; Blankenhorn, E.P. Attenuation of Th1 effector cell responses and susceptibility to experimental allergic encephalomyelitis in histamine H2 receptor knockout mice is due to dysregulation of cytokine production by antigen-presenting cells. Am. J. Pathol. 2004, 164, 883–892. [Google Scholar] [CrossRef] [PubMed]
- Niu, Y.C.; Feng, R.N.; Hou, Y.; Li, K.; Kang, Z.; Wang, J.; Sun, C.H.; Li, Y. Histidine and arginine are associated with inflammation and oxidative stress in obese women. Br. J. Nutr. 2012, 108, 57–61. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Watanabe, M.; Suliman, M.E.; Qureshi, A.R.; Garcia-Lopez, E.; Barany, P.; Heimburger, O.; Stenvinkel, P.; Lindholm, B. Consequences of low plasma histidine in chronic kidney disease patients: Associations with inflammation, oxidative stress, and mortality. Am. J. Clin. Nutr. 2008, 87, 1860–1866. [Google Scholar] [CrossRef] [PubMed]
- Teuscher, C.; Subramanian, M.; Noubade, R.; Gao, J.F.; Offner, H.; Zachary, J.F.; Blankenhorn, E.P. Central histamine H3 receptor signaling negatively regulates susceptibility to autoimmune inflammatory disease of the CNS. Proc. Natl. Acad. Sci. USA 2007, 104, 10146–10151. [Google Scholar] [CrossRef]
- Musio, S.; Gallo, B.; Scabeni, S.; Lapilla, M.; Poliani, P.L.; Matarese, G.; Ohtsu, H.; Galli, S.J.; Mantegazza, R.; Steinman, L.; et al. A key regulatory role for histamine in experimental autoimmune encephalomyelitis: Disease exacerbation in histidine decarboxylase-deficient mice. J. Immunol. 2006, 176, 17–26. [Google Scholar] [CrossRef]
- Liao, R.J.; Jiang, L.; Wang, R.R.; Zhao, H.W.; Chen, Y.; Li, Y.; Wang, L.; Jie, L.Y.; Zhou, Y.D.; Zhang, X.N.; et al. Histidine provides long-term neuroprotection after cerebral ischemia through promoting astrocyte migration. Sci. Rep. 2015, 5, 15356. [Google Scholar] [CrossRef]
- Rammohan, K.W. Histaminergic Basis of Fatigue in Multiple Sclerosis. Available online: https://clinicaltrials.gov/ct2/show/NCT04764383 (accessed on 8 November 2022).
- Rammohan, K.W. Histaminergic Basis of Central Fatigue in Multiple Sclerosis—A Novel Approach. Available online: https://clinicaltrials.gov/ct2/show/NCT03266965 (accessed on 8 November 2022).
- Noonan, F.P.; De Fabo, E.C. Immunosuppression by ultraviolet B radiation: Initiation by urocanic acid. Immunol. Today 1992, 13, 250–254. [Google Scholar] [CrossRef]
- Hart, P.H.; Gorman, S. Exposure to UV Wavelengths in Sunlight Suppresses Immunity. To What Extent is UV-induced Vitamin D3 the Mediator Responsible? Clin. Biochem. Rev. 2013, 34, 3–13. [Google Scholar]
- Norval, M.; El-Ghorr, A.A. Studies to determine the immunomodulating effects of cis-urocanic acid. Methods 2002, 28, 63–70. [Google Scholar] [CrossRef]
- Lopez-Alarcon, C.; Aspee, A.; Henriquez, C.; Campos, A.M.; Lissi, E.A. Interaction and reactivity of urocanic acid towards peroxyl radicals. Redox Rep. 2005, 10, 227–234. [Google Scholar] [CrossRef]
- Vidaurre, O.G.; Haines, J.D.; Katz Sand, I.; Adula, K.P.; Huynh, J.L.; McGraw, C.A.; Zhang, F.; Varghese, M.; Sotirchos, E.; Bhargava, P.; et al. Cerebrospinal fluid ceramides from patients with multiple sclerosis impair neuronal bioenergetics. Brain 2014, 137, 2271–2286. [Google Scholar] [CrossRef] [PubMed]
- Kurz, J.; Brunkhorst, R.; Foerch, C.; Blum, L.; Henke, M.; Gabriel, L.; Ulshofer, T.; Ferreiros, N.; Parnham, M.J.; Geisslinger, G.; et al. The relevance of ceramides and their synthesizing enzymes for multiple sclerosis. Clin. Sci. 2018, 132, 1963–1976. [Google Scholar] [CrossRef] [PubMed]
- Filippatou, A.G.; Moniruzzaman, M.; Sotirchos, E.S.; Fitzgerald, K.C.; Kalaitzidis, G.; Lambe, J.; Vasileiou, E.; Saidha, S.; Prince, J.L.; Haughey, N.; et al. Serum ceramide levels are altered in multiple sclerosis. Mult. Scler. J. 2021, 27, 1506–1519. [Google Scholar] [CrossRef]
- Checa, A.; Khademi, M.; Sar, D.G.; Haeggstrom, J.Z.; Lundberg, J.O.; Piehl, F.; Olsson, T.; Wheelock, C.E. Hexosylceramides as intrathecal markers of worsening disability in multiple sclerosis. Mult. Scler. J. 2015, 21, 1271–1279. [Google Scholar] [CrossRef]
- Mencarelli, C.; Martinez-Martinez, P. Ceramide function in the brain: When a slight tilt is enough. Cell. Mol. Life Sci. 2013, 70, 181–203. [Google Scholar] [CrossRef] [PubMed]
- Jana, A.; Hogan, E.L.; Pahan, K. Ceramide and neurodegeneration: Susceptibility of neurons and oligodendrocytes to cell damage and death. J. Neurol. Sci. 2009, 278, 5–15. [Google Scholar] [CrossRef]
- Jana, A.; Pahan, K. Oxidative stress kills human primary oligodendrocytes via neutral sphingomyelinase: Implications for multiple sclerosis. J. Neuroimmune Pharmacol. 2007, 2, 184–193. [Google Scholar] [CrossRef]
- Bikman, B.T.; Summers, S.A. Ceramides as modulators of cellular and whole-body metabolism. J. Clin. Investig. 2011, 121, 4222–4230. [Google Scholar] [CrossRef]
- Maceyka, M.; Spiegel, S. Sphingolipid metabolites in inflammatory disease. Nature 2014, 510, 58–67. [Google Scholar] [CrossRef]
- Wheeler, D.; Bandaru, V.V.; Calabresi, P.A.; Nath, A.; Haughey, N.J. A defect of sphingolipid metabolism modifies the properties of normal appearing white matter in multiple sclerosis. Brain 2008, 131, 3092–3102. [Google Scholar] [CrossRef]
- Jin, J.; Hou, Q.; Mullen, T.D.; Zeidan, Y.H.; Bielawski, J.; Kraveka, J.M.; Bielawska, A.; Obeid, L.M.; Hannun, Y.A.; Hsu, Y.T. Ceramide generated by sphingomyelin hydrolysis and the salvage pathway is involved in hypoxia/reoxygenation-induced Bax redistribution to mitochondria in NT-2 cells. J. Biol. Chem. 2008, 283, 26509–26517. [Google Scholar] [CrossRef]
- Mullen, T.D.; Hannun, Y.A.; Obeid, L.M. Ceramide synthases at the centre of sphingolipid metabolism and biology. Biochem. J. 2012, 441, 789–802. [Google Scholar] [CrossRef] [PubMed]
- Kwong, E.K.; Li, X.; Hylemon, P.B.; Zhou, H. Sphingosine Kinases/Sphingosine 1-Phosphate Signaling in Hepatic Lipid Metabolism. Curr. Pharmacol. Rep. 2017, 3, 176–183. [Google Scholar] [CrossRef] [PubMed]
- Mandala, S.M. Sphingosine-1-phosphate phosphatases. Prostaglandins Other Lipid Mediat. 2001, 64, 143–156. [Google Scholar] [CrossRef] [PubMed]
- Pyne, N.J.; Pyne, S. Sphingosine 1-phosphate and cancer. Nat. Rev. Cancer 2010, 10, 489–503. [Google Scholar] [CrossRef]
- Grbcic, P.; Sedic, M. Sphingosine 1-Phosphate Signaling and Metabolism in Chemoprevention and Chemoresistance in Colon Cancer. Molecules 2020, 25, 2436. [Google Scholar] [CrossRef]
- Movsesyan, V.A.; Yakovlev, A.G.; Dabaghyan, E.A.; Stoica, B.A.; Faden, A.I. Ceramide induces neuronal apoptosis through the caspase-9/caspase-3 pathway. Biochem. Biophys. Res. Commun. 2002, 299, 201–207. [Google Scholar] [CrossRef]
- Stoica, B.A.; Movsesyan, V.A.; Lea, P.M.t.; Faden, A.I. Ceramide-induced neuronal apoptosis is associated with dephosphorylation of Akt, BAD, FKHR, GSK-3beta, and induction of the mitochondrial-dependent intrinsic caspase pathway. Mol. Cell. Neurosci. 2003, 22, 365–382. [Google Scholar] [CrossRef]
- Scarlatti, F.; Bauvy, C.; Ventruti, A.; Sala, G.; Cluzeaud, F.; Vandewalle, A.; Ghidoni, R.; Codogno, P. Ceramide-mediated macroautophagy involves inhibition of protein kinase B and up-regulation of beclin 1. J. Biol. Chem. 2004, 279, 18384–18391. [Google Scholar] [CrossRef]
- Young, M.M.; Kester, M.; Wang, H.G. Sphingolipids: Regulators of crosstalk between apoptosis and autophagy. J. Lipid Res. 2013, 54, 5–19. [Google Scholar] [CrossRef]
- Cyster, J.G.; Schwab, S.R. Sphingosine-1-phosphate and lymphocyte egress from lymphoid organs. Annu. Rev. Immunol. 2012, 30, 69–94. [Google Scholar] [CrossRef] [PubMed]
- Pappu, R.; Schwab, S.R.; Cornelissen, I.; Pereira, J.P.; Regard, J.B.; Xu, Y.; Camerer, E.; Zheng, Y.W.; Huang, Y.; Cyster, J.G.; et al. Promotion of lymphocyte egress into blood and lymph by distinct sources of sphingosine-1-phosphate. Science 2007, 316, 295–298. [Google Scholar] [CrossRef] [PubMed]
- Baeyens, A.; Fang, V.; Chen, C.; Schwab, S.R. Exit Strategies: S1P Signaling and T Cell Migration. Trends Immunol. 2015, 36, 778–787. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Obinata, H.; Hla, T. Sphingosine 1-phosphate and inflammation. Int. Immunol. 2019, 31, 617–625. [Google Scholar] [CrossRef]
- McGinley, M.P.; Cohen, J.A. Sphingosine 1-phosphate receptor modulators in multiple sclerosis and other conditions. Lancet 2021, 398, 1184–1194. [Google Scholar] [CrossRef] [PubMed]
- Yelamanchi, S.D.; Jayaram, S.; Thomas, J.K.; Gundimeda, S.; Khan, A.A.; Singhal, A.; Keshava Prasad, T.S.; Pandey, A.; Somani, B.L.; Gowda, H. A pathway map of glutamate metabolism. J. Cell Commun. Signal. 2016, 10, 69–75. [Google Scholar] [CrossRef] [PubMed]
- Pitt, D.; Werner, P.; Raine, C.S. Glutamate excitotoxicity in a model of multiple sclerosis. Nat. Med. 2000, 6, 67–70. [Google Scholar] [CrossRef] [PubMed]
- Liu, J.; Zhang, X.; Chen, K.; Cheng, Y.; Liu, S.; Xia, M.; Chen, Y.; Zhu, H.; Li, Z.; Cao, X. CCR7 Chemokine Receptor-Inducible lnc-Dpf3 Restrains Dendritic Cell Migration by Inhibiting HIF-1alpha-Mediated Glycolysis. Immunity 2019, 50, 600–615. [Google Scholar] [CrossRef]
- Frauwirth, K.A.; Riley, J.L.; Harris, M.H.; Parry, R.V.; Rathmell, J.C.; Plas, D.R.; Elstrom, R.L.; June, C.H.; Thompson, C.B. The CD28 signaling pathway regulates glucose metabolism. Immunity 2002, 16, 769–777. [Google Scholar] [CrossRef]
- Cham, C.M.; Driessens, G.; O’Keefe, J.P.; Gajewski, T.F. Glucose deprivation inhibits multiple key gene expression events and effector functions in CD8+ T cells. Eur. J. Immunol. 2008, 38, 2438–2450. [Google Scholar] [CrossRef]
- Cham, C.M.; Gajewski, T.F. Glucose availability regulates IFN-gamma production and p70S6 kinase activation in CD8+ effector T cells. J. Immunol. 2005, 174, 4670–4677. [Google Scholar] [CrossRef] [PubMed]
- Cao, Y.; Rathmell, J.C.; Macintyre, A.N. Metabolic reprogramming towards aerobic glycolysis correlates with greater proliferative ability and resistance to metabolic inhibition in CD8 versus CD4 T cells. PLoS ONE 2014, 9, e104104. [Google Scholar] [CrossRef] [PubMed]
- Krawczyk, C.M.; Holowka, T.; Sun, J.; Blagih, J.; Amiel, E.; DeBerardinis, R.J.; Cross, J.R.; Jung, E.; Thompson, C.B.; Jones, R.G.; et al. Toll-like receptor-induced changes in glycolytic metabolism regulate dendritic cell activation. Blood 2010, 115, 4742–4749. [Google Scholar] [CrossRef] [Green Version]
- Everts, B.; Amiel, E.; van der Windt, G.J.; Freitas, T.C.; Chott, R.; Yarasheski, K.E.; Pearce, E.L.; Pearce, E.J. Commitment to glycolysis sustains survival of NO-producing inflammatory dendritic cells. Blood 2012, 120, 1422–1431. [Google Scholar] [CrossRef] [PubMed]
- O’Neill, L.A.; Kishton, R.J.; Rathmell, J. A guide to immunometabolism for immunologists. Nat. Rev. Immunol. 2016, 16, 553–565. [Google Scholar] [CrossRef]
- Guak, H.; Al Habyan, S.; Ma, E.H.; Aldossary, H.; Al-Masri, M.; Won, S.Y.; Ying, T.; Fixman, E.D.; Jones, R.G.; McCaffrey, L.M.; et al. Glycolytic metabolism is essential for CCR7 oligomerization and dendritic cell migration. Nat. Commun. 2018, 9, 2463. [Google Scholar] [CrossRef] [PubMed]
- Kornberg, M.D.; Bhargava, P.; Kim, P.M.; Putluri, V.; Snowman, A.M.; Putluri, N.; Calabresi, P.A.; Snyder, S.H. Dimethyl fumarate targets GAPDH and aerobic glycolysis to modulate immunity. Science 2018, 360, 449–453. [Google Scholar] [CrossRef]
- Negrotto, L.; Correale, J. Amino Acid Catabolism in Multiple Sclerosis Affects Immune Homeostasis. J. Immunol. 2017, 198, 1900–1909. [Google Scholar] [CrossRef]
- Moussallieh, F.M.; Elbayed, K.; Chanson, J.B.; Rudolf, G.; Piotto, M.; De Seze, J.; Namer, I.J. Serum analysis by 1H nuclear magnetic resonance spectroscopy: A new tool for distinguishing neuromyelitis optica from multiple sclerosis. Mult. Scler. J. 2014, 20, 558–565. [Google Scholar] [CrossRef]
- Schwarcz, R.; Bruno, J.P.; Muchowski, P.J.; Wu, H.Q. Kynurenines in the mammalian brain: When physiology meets pathology. Nat. Rev. Neurosci. 2012, 13, 465–477. [Google Scholar] [CrossRef]
- Schwarcz, R.; Whetsell, W.O., Jr.; Mangano, R.M. Quinolinic acid: An endogenous metabolite that produces axon-sparing lesions in rat brain. Science 1983, 219, 316–318. [Google Scholar] [CrossRef] [PubMed]
- Rahman, A.; Rao, M.S.; Khan, K.M. Intraventricular infusion of quinolinic acid impairs spatial learning and memory in young rats: A novel mechanism of lead-induced neurotoxicity. J. Neuroinflammation 2018, 15, 263. [Google Scholar] [CrossRef] [PubMed]
- Guillemin, G.J. Quinolinic acid, the inescapable neurotoxin. FEBS J. 2012, 279, 1356–1365. [Google Scholar] [CrossRef]
- Tavares, R.G.; Tasca, C.I.; Santos, C.E.; Wajner, M.; Souza, D.O.; Dutra-Filho, C.S. Quinolinic acid inhibits glutamate uptake into synaptic vesicles from rat brain. Neuroreport 2000, 11, 249–253. [Google Scholar] [CrossRef] [PubMed]
- Aeinehband, S.; Brenner, P.; Stahl, S.; Bhat, M.; Fidock, M.D.; Khademi, M.; Olsson, T.; Engberg, G.; Jokinen, J.; Erhardt, S.; et al. Cerebrospinal fluid kynurenines in multiple sclerosis; relation to disease course and neurocognitive symptoms. Brain Behav. Immun. 2016, 51, 47–55. [Google Scholar] [CrossRef] [PubMed]
- Sintzel, M.B.; Rametta, M.; Reder, A.T. Vitamin D and Multiple Sclerosis: A Comprehensive Review. Neurol. Ther. 2018, 7, 59–85. [Google Scholar] [CrossRef] [PubMed]
- Mieszkowski, J.; Brzezinska, P.; Stankiewicz, B.; Kochanowicz, A.; Niespodzinski, B.; Reczkowicz, J.; Waldzinski, T.; Kacprzak, B.; Siuba-Jarosz, N.; Petr, M.; et al. Direct Effects of Vitamin D Supplementation on Ultramarathon-Induced Changes in Kynurenine Metabolism. Nutrients 2022, 14, 4485. [Google Scholar] [CrossRef]
- Tohari, A.M.; Zhou, X.; Shu, X. Protection against oxidative stress by vitamin D in cone cells. Cell Biochem. Funct. 2016, 34, 82–94. [Google Scholar] [CrossRef]
- Bhat, M.; Ismail, A. Vitamin D treatment protects against and reverses oxidative stress induced muscle proteolysis. J. Steroid Biochem. Mol. Biol. 2015, 152, 171–179. [Google Scholar] [CrossRef]
- Jain, S.K.; Micinski, D. Vitamin D upregulates glutamate cysteine ligase and glutathione reductase, and GSH formation, and decreases ROS and MCP-1 and IL-8 secretion in high-glucose exposed U937 monocytes. Biochem. Biophys. Res. Commun. 2013, 437, 7–11. [Google Scholar] [CrossRef]
- Brewer, L.D.; Thibault, V.; Chen, K.C.; Langub, M.C.; Landfield, P.W.; Porter, N.M. Vitamin D hormone confers neuroprotection in parallel with downregulation of L-type calcium channel expression in hippocampal neurons. J. Neurosci. 2001, 21, 98–108. [Google Scholar] [CrossRef] [PubMed]
- Hur, J.; Lee, P.; Kim, M.J.; Cho, Y.W. Regulatory Effect of 25-hydroxyvitamin D3 on Nitric Oxide Production in Activated Microglia. Korean J. Physiol. Pharmacol. 2014, 18, 397–402. [Google Scholar] [CrossRef] [PubMed]
- Boontanrart, M.; Hall, S.D.; Spanier, J.A.; Hayes, C.E.; Olson, J.K. Vitamin D3 alters microglia immune activation by an IL-10 dependent SOCS3 mechanism. J. Neuroimmunol. 2016, 292, 126–136. [Google Scholar] [CrossRef] [PubMed]
- Li, K.; Naviaux, J.C.; Bright, A.T.; Wang, L.; Naviaux, R.K. A robust, single-injection method for targeted, broad-spectrum plasma metabolomics. Metabolomics 2017, 13, 122. [Google Scholar] [CrossRef] [PubMed]
- Naviaux, R.K.; Naviaux, J.C.; Li, K.; Wang, L.; Monk, J.M.; Bright, A.T.; Koslik, H.J.; Ritchie, J.B.; Golomb, B.A. Metabolic features of Gulf War illness. PLoS ONE 2019, 14, e0219531. [Google Scholar] [CrossRef]
- Li, K.; Naviaux, J.C.; Monk, J.M.; Wang, L.; Naviaux, R.K. Improved Dried Blood Spot-Based Metabolomics: A Targeted, Broad-Spectrum, Single-Injection Method. Metabolites 2020, 10, 82. [Google Scholar] [CrossRef]
- Spectrometry. Liquid Chromatography. Available online: https://www.sigmaaldrich.com/RU/ru/applications/analytical-chemistry/liquid-chromatography (accessed on 8 November 2022).
- Pang, Z. MetaboAnalyst 5.0: Narrowing the gap between raw spectra and functional insights. Nucleic Acids Res. 2021, 49, W388–W396. [Google Scholar] [CrossRef]











{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Decreased in MS Compared to CTRL | Increased in MS Compared to CTRL | ||||
|---|---|---|---|---|---|
| Seasons | Total Number of Shared Metabolites | Metabolite Names | Seasons | Total Number of Shared Metabolites | Metabolite Names |
| Fall Spring Summer Winter | 3 | Hexacosanoic acid | Fall Spring Summer Winter | 1 | Sphingosine |
| DHC (18:1/24:0) | Fall Spring Winter | 1 | Ceramide (d18:1/16:0) | ||
| Gentisic acid | Fall Spring Summer | 1 | Ceramide (d18:1/18:1 OH) | ||
| Fall Spring Winter | 1 | Glycylproline | Spring Summer Winter | 2 | Ceramide (d18:1/24:0) |
| Fall Spring Summer | 2 | Gamma-Aminobutyric acid | Deoxyguanosine | ||
| Docosahexaenoic acid (DHA) | Fall Winter | 1 | Ceramide (d18:1/18:1) | ||
| Spring Summer Winter | 1 | Vitamin D3 | Fall Spring | 1 | Deoxyguanosine 5’-monophosphate |
| Fall Winter | 3 | 4-Hydroxyphenylpyruvic acid | Summer Winter | 1 | Ceramide (d18:1/22:0 OH) |
| 4,5-Dihydroorotic acid | Spring Summer | 1 | Cytidine monophosphate | ||
| N-Acetylserine | Fall | 1 | Niacinamide | ||
| Fall Spring | 10 | Selenomethionine | Winter | 12 | Linoleic acid |
| L-Histidine | Alpha-Tocopherol | ||||
| Carnosine | Cer(d18:1/12:0) | ||||
| 4-Aminohippuric acid | N-Methyltyramine | ||||
| Agmatine | Eicosapentaenoic acid | ||||
| Hydroxykynurenine | Homovanillic acid | ||||
| LysoPE (P-18:0/0:0) | Biotin | ||||
| Methionine sulfoxide | 2-Arachidonylglycerol | ||||
| Glycerol | Glucosamine | ||||
| Plasmalogen (p18:0/22:6) | PE (20:1(11Z)/16:0) | ||||
| Fall Summer | 1 | CDP-choline | Quinolinic acid | ||
| Spring Winter | 2 | Allantoin | PC (18:0/18:2(9Z,12Z)) | ||
| Methylguanidine | Summer | 2 | L-Arginine | ||
| Summer Winter | 1 | L-Lactic acid | Guanosine | ||
| Fall | 38 | Glucose 6-phosphate | |||
| N-Acetylputrescine | |||||
| Gluconolactone | |||||
| Cer (d18:1/12:0) | |||||
| L-Isoleucine | |||||
| Picolinic acid | |||||
| N-a-Acetyl-L-arginine | |||||
| Anandamide | |||||
| L-Asparagine | |||||
| L-Valine | |||||
| Ornithine | |||||
| L-Arginine | |||||
| Glyceric acid | |||||
| Glucose | |||||
| 2-Keto-L-gluconate | |||||
| Myo-Inositol | |||||
| Homovanillic acid | |||||
| L-Cystathionine | |||||
| Glycolic acid | |||||
| S-Adenosylhomocysteine | |||||
| PE (18:0/16:1(9Z)) | |||||
| 2 Aminocaprylic acid | |||||
| Mevalonic acid | |||||
| Normetanephrine | |||||
| Homocysteine | |||||
| Methylcysteine | |||||
| Coenzyme Q9 | |||||
| L-Homoserine | |||||
| O-Acetylserine | |||||
| Ascorbic acid | |||||
| L-Leucine | |||||
| Ureidopropionic acid | |||||
| PG (16:0/16:0) | |||||
| Dimethyl-L-arginine | |||||
| L-Glutamic acid | |||||
| Hypotaurine | |||||
| L-Kynurenine | |||||
| L-Phenylalanine | |||||
| Winter | 1 | 5-Hydroxy-L-tryptophan | |||
| Spring | 2 | Riboflavin | |||
| Chenodeoxycholic acid | |||||
| Summer | 2 | O-Phosphorylethanolamine | |||
| DL-O-Phosphoserine | |||||
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Martynova, E.; Khaibullin, T.; Salafutdinov, I.; Markelova, M.; Laikov, A.; Lopukhov, L.; Liu, R.; Sahay, K.; Goyal, M.; Baranwal, M.; et al. Seasonal Changes in Serum Metabolites in Multiple Sclerosis Relapse. Int. J. Mol. Sci. 2023, 24, 3542. https://doi.org/10.3390/ijms24043542
Martynova E, Khaibullin T, Salafutdinov I, Markelova M, Laikov A, Lopukhov L, Liu R, Sahay K, Goyal M, Baranwal M, et al. Seasonal Changes in Serum Metabolites in Multiple Sclerosis Relapse. International Journal of Molecular Sciences. 2023; 24(4):3542. https://doi.org/10.3390/ijms24043542
Chicago/Turabian StyleMartynova, Ekaterina, Timur Khaibullin, Ilnur Salafutdinov, Maria Markelova, Alexander Laikov, Leonid Lopukhov, Rongzeng Liu, Kritika Sahay, Mehendi Goyal, Manoj Baranwal, and et al. 2023. "Seasonal Changes in Serum Metabolites in Multiple Sclerosis Relapse" International Journal of Molecular Sciences 24, no. 4: 3542. https://doi.org/10.3390/ijms24043542