
Overexpression of Water-Responsive Genes Promoted by Elevated CO2 Reduces ROS and Enhances Drought Tolerance in Coffea Species

 ,
,  , ,
, ,  , , and
, , and 
Abstract
:1. Introduction
2. Results
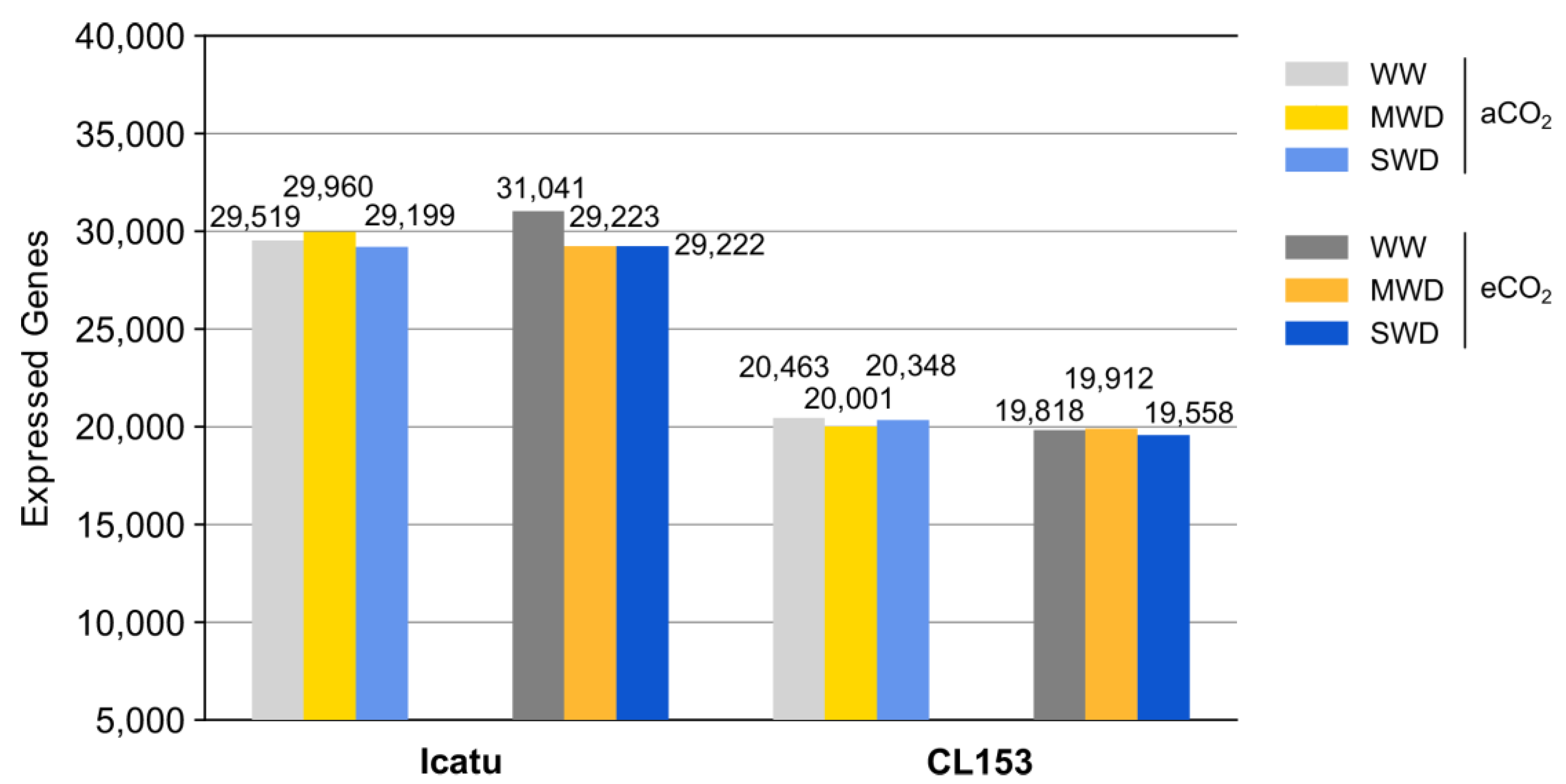
2.1. Overview of the RNA-Seq Data from the Two Coffee Genotypes
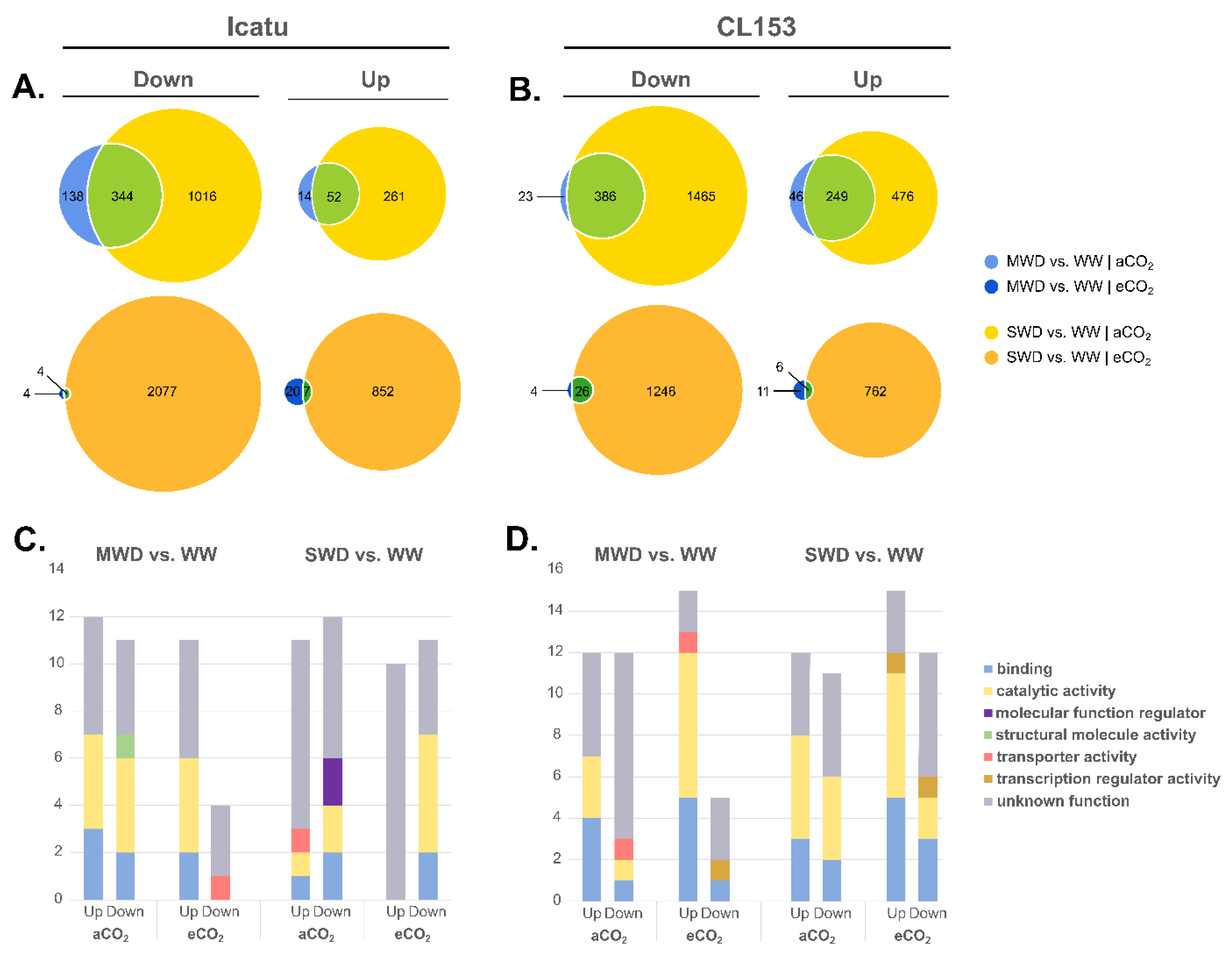
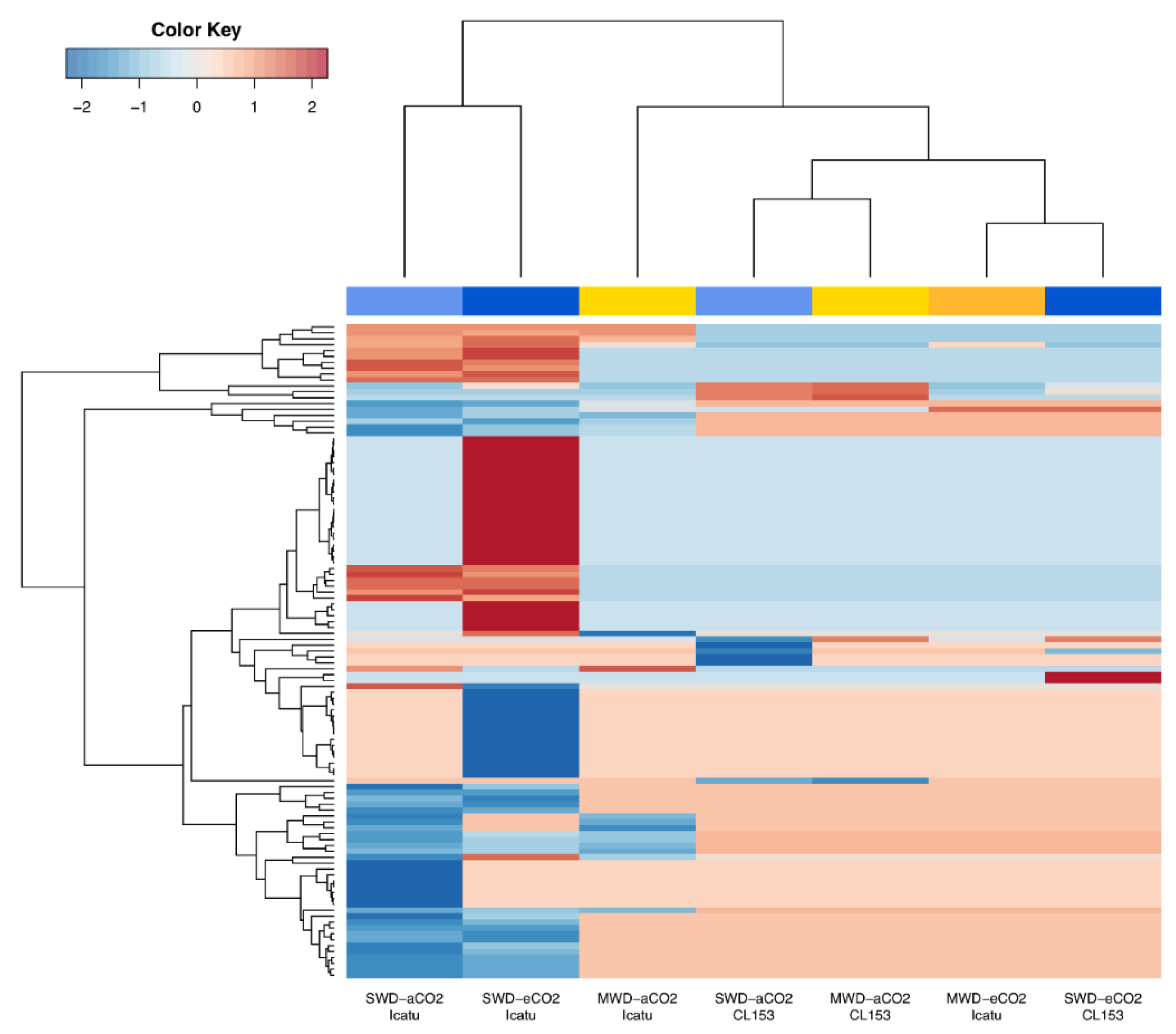
2.2. Response of Differentially Expressed Genes (DEGs) to Drought and eCO2
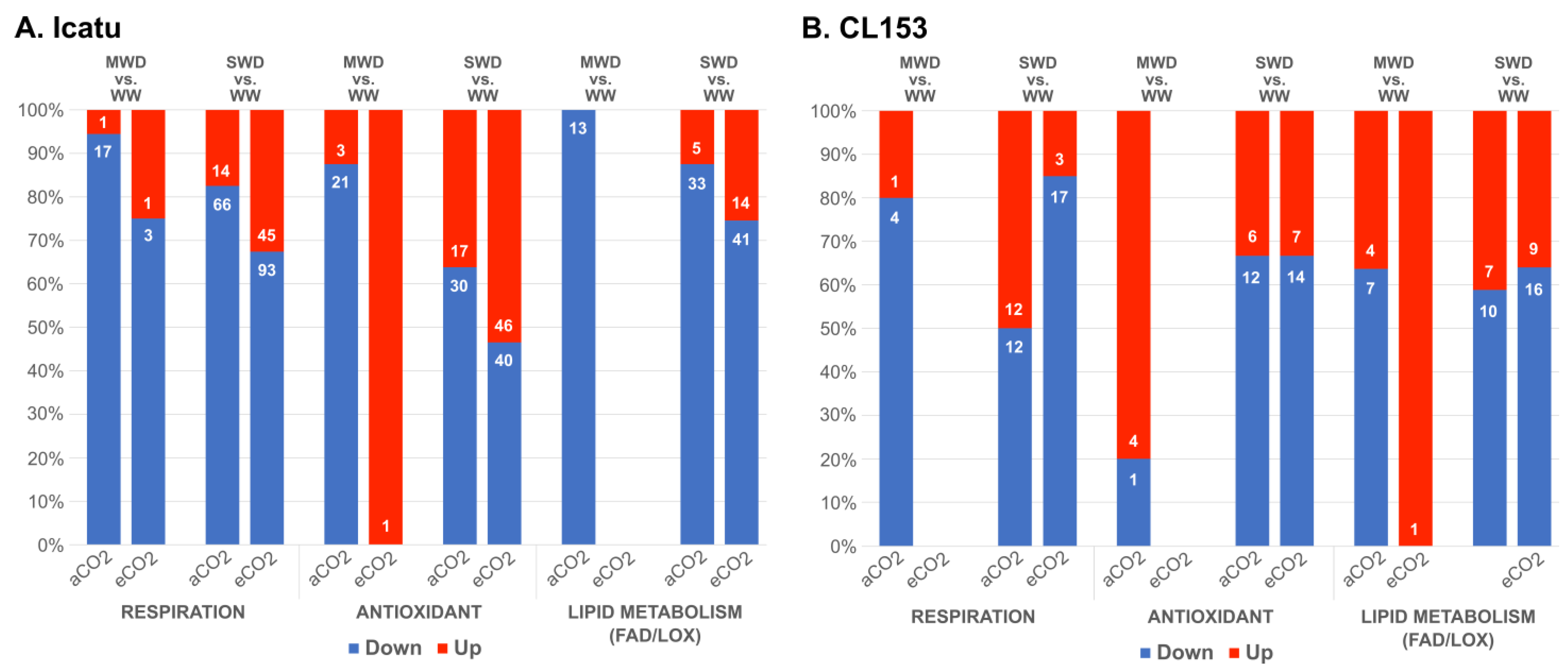
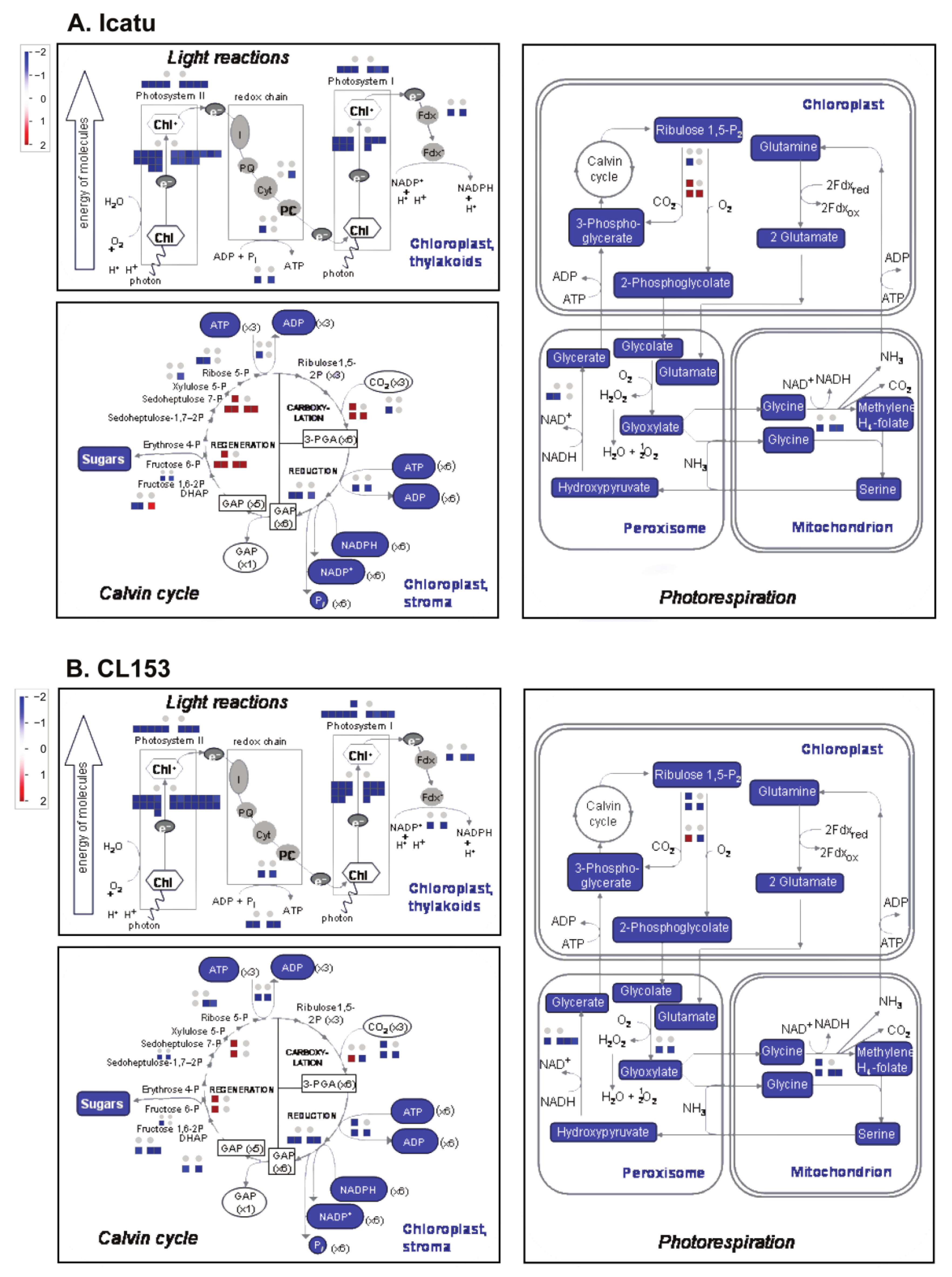
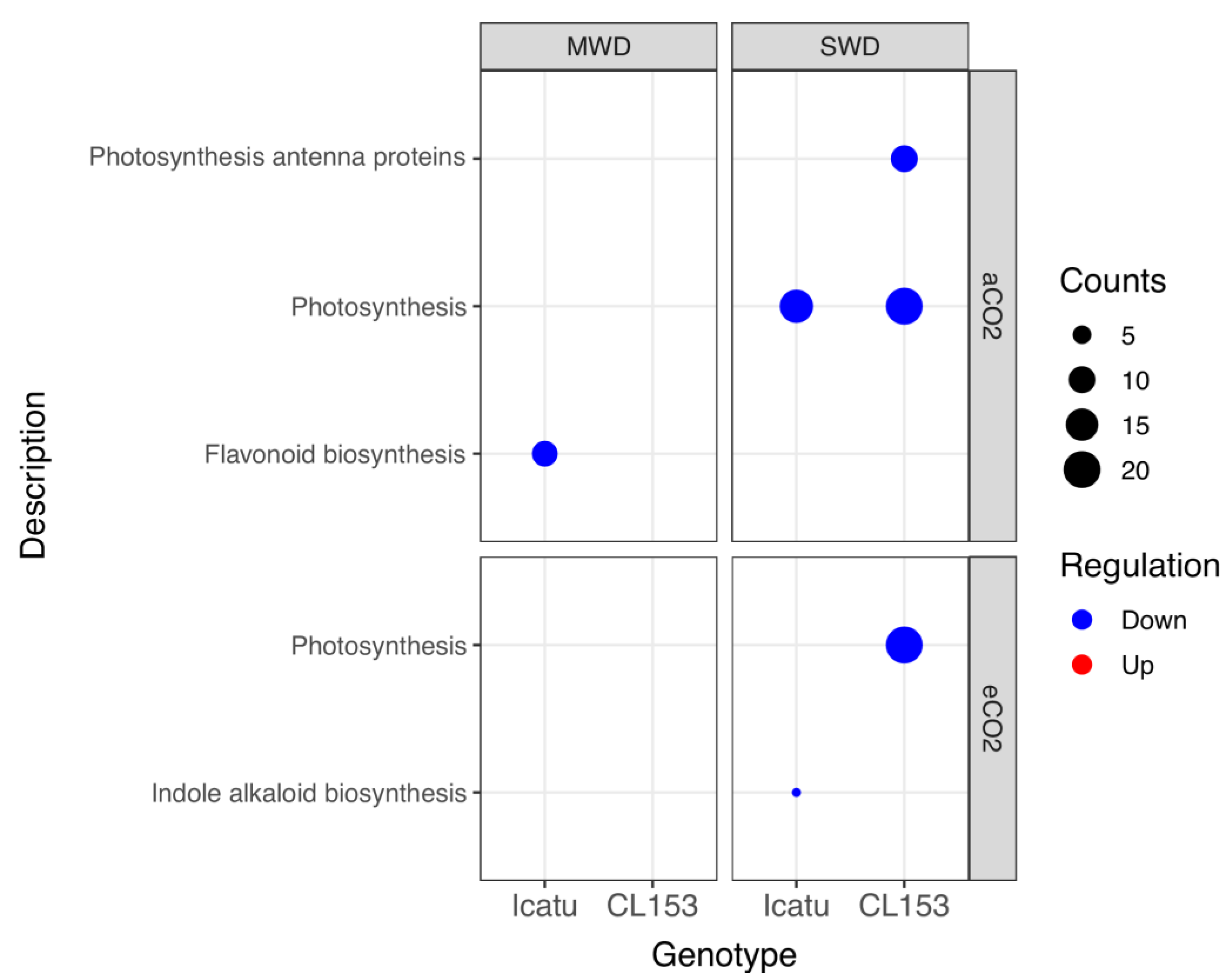
2.3. Drought and eCO2 Impact on DEGs Associated with Specific Biochemical Pathways
2.4. DEGs Involved in the Response to Water Deprivation and Desiccation
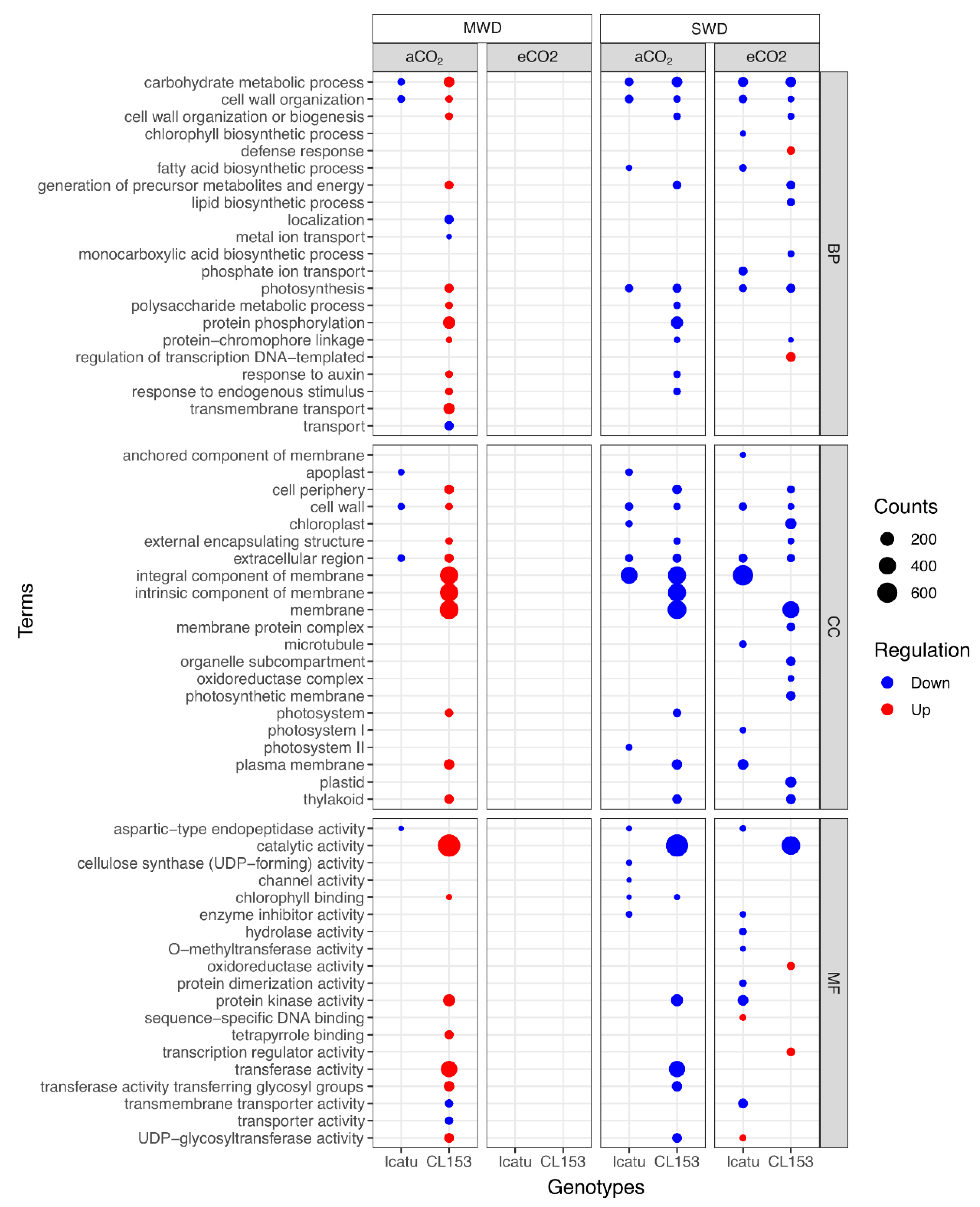
2.5. Enriched Gene Ontology (GO) Terms and Functional Pathways Responding to Drought and eCO2
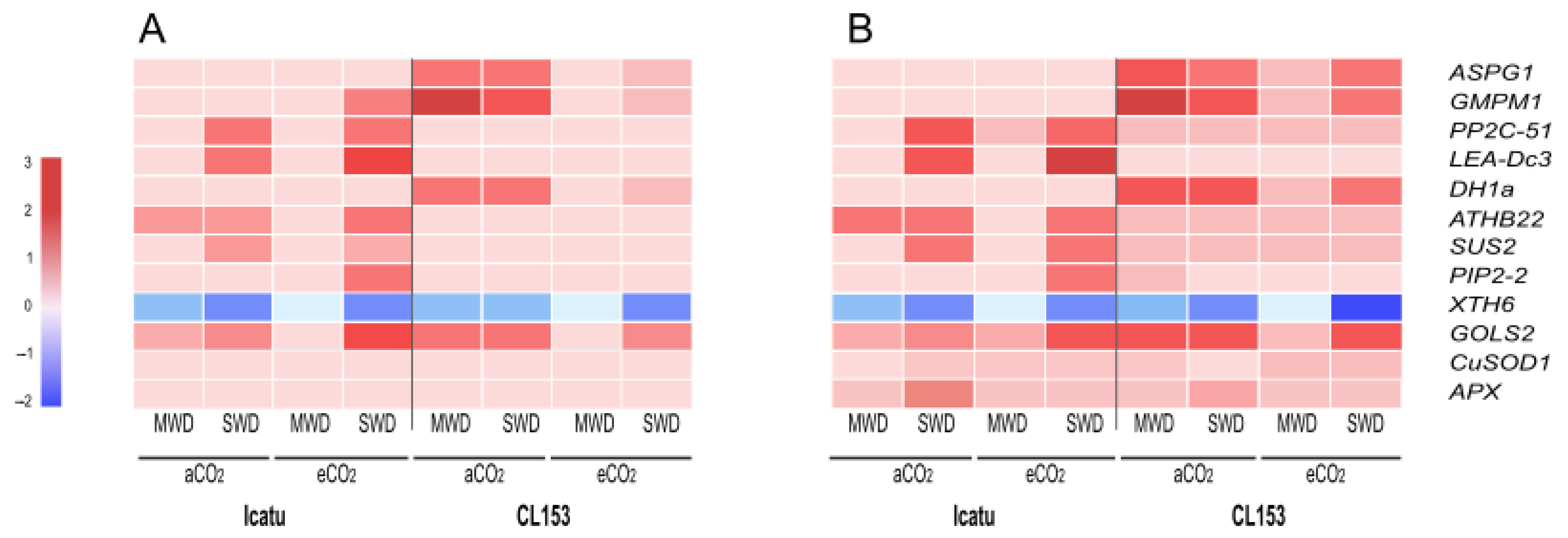
2.6. Validation of RNA-Seq Results by qRT-PCR
3. Discussion
3.1. Differential Transcriptional Drought Regulation Responses in the Two Coffea Genotypes
3.2. Influence of eCO2 in Drought-Responsive Genes
3.3. Overall Regulatory Mechanisms Involved in Coffea Responses
3.4. Evidence of Post-Transcriptional Regulatory Mechanisms in Coffea Responses to Stress
4. Materials and Methods
4.1. Plant Growth Conditions
4.2. Imposition and Monitoring of Water Deficit Conditions and Sampling
4.3. RNA Extraction and Illumina Sequencing
4.4. Quality Analysis of Sequencing Data
4.5. Reference-Based Mapping and Assembly
4.6. Identification of Differentially Expressed Genes (DEGs)
4.7. Drought and eCO2 Impact on DEGs Associated with Specific Biochemical Pathways
4.8. DEGs Involved in the Response to Water Deprivation and Desiccation
4.9. Functional Classification of Responsive DEGs
4.10. Quantitative RT-PCR
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Solomon, S.; Plattner, G.K.; Knutti, R.; Friedlingstein, P. Irreversible climate change due to carbon dioxide emissions. Proc. Natl. Acad. Sci. USA 2009, 106, 1704–1709. [Google Scholar] [CrossRef] [PubMed]
- Raza, A.; Mubarik, M.S.; Sharif, R.; Habib, M.; Jabeen, W.; Zhang, C.; Chen, H.; Chen, Z.-H.; Siddique, K.H.M.; Zhuang, W.; et al. The Plant Genome Developing drought-smart, ready-to-grow future crops. Plant Genome 2022, e20279. [Google Scholar] [CrossRef] [PubMed]
- Brodribb, T.J.; McAdam, S.A.M. Evolution of the Stomatal Regulation of Plant Water Content. Plant Physiol. 2017, 174, 639–649. [Google Scholar] [CrossRef]
- Chaves, M.M.; Maroco, J.P.; Pereira, J.S. Understanding plant responses to drought-From genes to the whole plant. Funct. Plant Biol. 2003, 30, 239–264. [Google Scholar] [CrossRef] [PubMed]
- Muller, B.; Pantin, F.; Génard, M.; Turc, O.; Freixes, S.; Piques, M.; Gibon, Y. Water deficits uncouple growth from photosynthesis, increase C content, and modify the relationships between C and growth in sink organs. J. Exp. Bot. 2011, 62, 1715–1729. [Google Scholar] [CrossRef]
- Ramalho, J.C.; DaMatta, F.M.; Rodrigues, A.P.; Scotti-Campos, P.; Pais, I.; Batista-Santos, P.; Partelli, F.L.; Ribeiro, A.; Lidon, F.C.; Leitão, A.E. Cold impact and acclimation response of Coffea spp. plants. Theor. Exp. Plant Physiol. 2014, 26, 5–18. [Google Scholar] [CrossRef]
- Ramalho, J.C.; Rodrigues, A.P.; Lidon, F.C.; Marques, L.M.C.; Leitão, A.E.; Fortunato, A.S.; Pais, I.P.; Silva, M.J.; Scotti-Campos, P.; Lopes, A.; et al. Stress cross-response of the antioxidative system promoted by superimposed drought and cold conditions in Coffea spp. PLoS ONE 2018, 13, e0198694. [Google Scholar] [CrossRef]
- Fahad, S.; Bajwa, A.A.; Nazir, U.; Anjum, S.A.; Farooq, A.; Zohaib, A.; Sadia, S.; Nasim, W.; Adkins, S.; Saud, S.; et al. Crop production under drought and heat stress: Plant responses and management options. Front. Plant Sci. 2017, 8, 1147. [Google Scholar] [CrossRef]
- Chaves, M.M.; Flexas, J.; Pinheiro, C. Photosynthesis under drought and salt stress: Regulation mechanisms from whole plant to cell. Ann. Bot. 2009, 103, 551–560. [Google Scholar] [CrossRef]
- Ainsworth, E.A.; Rogers, A. The response of photosynthesis and stomatal conductance to rising [CO2]: Mechanisms and environmental interactions. Plant Cell Environ. 2007, 30, 258–270. [Google Scholar] [CrossRef]
- Leakey, A.D.B.; Ainsworth, E.A.; Bernacchi, C.J.; Rogers, A.; Long, S.P.; Ort, D.R. Elevated CO2 effects on plant carbon, nitrogen, and water relations: Six important lessons from FACE. J. Exp. Bot. 2009, 60, 2859–2876. [Google Scholar] [CrossRef]
- Semedo, J.N.; Rodrigues, A.P.; Lidon, F.C.; Pais, I.P.; Marques, I.; Gouveia, D.; Armengaud, J.; Silva, M.J.; Martins, S.; Semedo, M.C.; et al. Intrinsic non-stomatal resilience to drought of the photosynthetic apparatus in Coffea spp. is strengthened by elevated air [CO2]. Tree Physiol. 2021, 41, 708–727. [Google Scholar] [CrossRef]
- Tausz-Posch, S.; Tausz, M.; Bourgault, M. Elevated [CO2] effects on crops: Advances in understanding acclimation, nitrogen dynamics and interactions with drought and other organisms. Plant Biol. 2020, 22, 38–51. [Google Scholar] [CrossRef]
- De Souza, A.P.; Cocuron, J.-C.; Garcia, A.C.; Alonso, A.P.; Buckeridge, M.S. Changes in Whole-Plant Metabolism during the Grain-Filling Stage in Sorghum Grown under Elevated CO2 and Drought. Plant Physiol. 2015, 169, 1755–1765. [Google Scholar] [CrossRef]
- Gray, S.B.; Dermody, O.; Klein, S.P.; Locke, A.M.; McGrath, J.M.; Paul, R.E.; Rosenthal, D.M.; Ruiz-Vera, U.M.; Siebers, M.H.; Strellner, R.; et al. Intensifying drought eliminates the expected benefits of elevated carbon dioxide for soybean. Nat. Plants 2016, 2, 16132. [Google Scholar] [CrossRef]
- ICO International Coffee Organization-What’s New. Available online: https://www.ico.org/ (accessed on 22 November 2022).
- DaMatta, F.M.; Rahn, E.; Läderach, P.; Ghini, R.; Ramalho, J.C. Why could the coffee crop endure climate change and global warming to a greater extent than previously estimated? Clim. Change 2019, 152, 167–178. [Google Scholar] [CrossRef]
- Cassamo, C.T.; Mangueze, A.V.J.; Leitão, A.E.; Pais, I.P.; Moreira, R.; Campa, C.; Chiulele, R.; Reis, F.O.; Marques, I.; Scotti-Campos, P.; et al. Shade and Altitude Implications on the Physical and Chemical Attributes of Green Coffee Beans from Gorongosa Mountain, Mozambique. Agronomy 2022, 12, 2540. [Google Scholar] [CrossRef]
- DaMatta, F.M.; Cochicho Ramalho, J.D. Impacts of drought and temperature stress on coffee physiology and production: A review. Braz. J. Plant Physiol. 2006, 18, 55–81. [Google Scholar] [CrossRef]
- Vinecky, F.; Davrieux, F.; Mera, A.C.; Alves, G.S.C.; Lavagnini, G.; Leroy, T.; Bonnot, F.; Rocha, O.C.; Bartholo, G.F.; Guerra, A.F.; et al. Controlled irrigation and nitrogen, phosphorous and potassium fertilization affect the biochemical composition and quality of Arabica coffee beans. J. Agric. Sci. 2017, 155, 902–918. [Google Scholar] [CrossRef]
- Avila, R.T.; Cardoso, A.A.; de Almeida, W.L.; Costa, L.C.; Machado, K.L.G.; Barbosa, M.L.; de Souza, R.P.B.; Oliveira, L.A.; Batista, D.S.; Martins, S.C.V.; et al. Coffee plants respond to drought and elevated [CO2] through changes in stomatal function, plant hydraulic conductance, and aquaporin expression. Environ. Exp. Bot. 2020, 177, 104148. [Google Scholar] [CrossRef]
- Avila, R.T.; de Almeida, W.L.; Costa, L.C.; Machado, K.L.G.; Barbosa, M.L.; de Souza, R.P.B.; Martino, P.B.; Juárez, M.A.T.; Marçal, D.M.S.; Martins, S.C.V.; et al. Elevated air [CO2] improves photosynthetic performance and alters biomass accumulation and partitioning in drought-stressed coffee plants. Environ. Exp. Bot. 2020, 177, 104137. [Google Scholar] [CrossRef]
- Sanches, R.F.E.; da Cruz Centeno, D.; Braga, M.R.; da Silva, E.A. Impact of high atmospheric CO2 concentrations on the seasonality of water-related processes, gas exchange, and carbohydrate metabolism in coffee trees under field conditions. Clim. Chang. 2020, 162, 1231–1248. [Google Scholar] [CrossRef]
- Catarino, I.C.A.; Monteiro, G.B.; Ferreira, M.J.P.; Torres, L.M.B.; Domingues, D.S.; Centeno, D.C.; Lobo, A.K.M.; Silva, E.A. Elevated [CO2] Mitigates Drought Effects and Increases Leaf 5-O-Caffeoylquinic Acid and Caffeine Concentrations during the Early Growth of Coffea Arabica Plants. Front. Sustain. Food Syst. 2021, 5, 266. [Google Scholar] [CrossRef]
- Rodrigues, A.M.; Jorge, T.; Osorio, S.; Pott, D.M.; Lidon, F.C.; DaMatta, F.M.; Marques, I.; Ribeiro-barros, A.I.; Ramalho, J.C.; António, C. Primary Metabolite Profile Changes in Coffea spp. Promoted by Single and Combined Exposure to Drought and Elevated CO2 Concentration. Metabolites 2021, 11, 427. [Google Scholar] [CrossRef]
- Dubberstein, D.; Lidon, F.C.; Rodrigues, A.P.; Semedo, J.N.; Marques, I.; Rodrigues, W.P.; Gouveia, D.; Armengaud, J.; Semedo, M.C.; Martins, S.; et al. Resilient and Sensitive Key Points of the Photosynthetic Machinery of Coffea spp. to the Single and Superimposed Exposure to Severe Drought and Heat Stresses. Front. Plant Sci. 2020, 11, 1049. [Google Scholar] [CrossRef]
- Marques, I.; Rodrigues, A.P.; Gouveia, D.; Lidon, F.C.; Martins, S.; Semedo, M.C.; Gaillard, J.C.; Pais, I.P.; Semedo, J.N.; Scotti-Campos, P.; et al. High-resolution shotgun proteomics reveals that increased air [CO2] amplifies the acclimation response of Coffea species to drought regarding antioxidative, energy, sugar, and lipid dynamics. J. Plant Physiol. 2022, 276, 153788. [Google Scholar] [CrossRef]
- Vranová, E.; Langebartels, C.; Van Montagu, M.; Inzé, D.; Van Camp, W. Oxidative stress, heat shock and drought differentially affect expression of a tobacco protein phosphatase 2C. J. Exp. Bot. 2000, 51, 1763–1764. [Google Scholar] [CrossRef]
- Merlot, S.; Gosti, F.; Guerrier, D.; Vavasseur, A.; Giraudat, J. The ABI1 and ABI2 protein phosphatases 2C act in a negative feedback regulatory loop of the abscisic acid signalling pathway. Plant J. 2001, 25, 295–303. [Google Scholar] [CrossRef]
- Komatsu, K.; Suzuki, N.; Kuwamura, M.; Nishikawa, Y.; Nakatani, M.; Ohtawa, H.; Takezawa, D.; Seki, M.; Tanaka, M.; Taji, T.; et al. Group A PP2Cs evolved in land plants as key regulators of intrinsic desiccation tolerance. Nat. Commun. 2013, 4, 2219. [Google Scholar] [CrossRef]
- Sun, L.; Wang, Y.P.; Chen, P.; Ren, J.; Ji, K.; Li, Q.; Li, P.; Dai, S.J.; Leng, P. Transcriptional regulation of SlPYL, SlPP2C, and SlSnRK2 gene families encoding ABA signal core components during tomato fruit development and drought stress. J. Exp. Bot. 2011, 62, 5659–5669. [Google Scholar] [CrossRef] [Green Version]
- Chen, J.; Zhang, D.; Zhang, C.; Xia, X.; Yin, W.; Tian, Q. A Putative PP2C-encoding gene negatively regulates ABA signaling in populus euphratica. PLoS ONE 2015, 10, e0139466. [Google Scholar] [CrossRef]
- Zhang, F.; Fu, X.; Lv, Z.; Shen, Q.; Yan, T.; Jiang, W.; Wang, G.; Sun, X.; Tang, K. Type 2C phosphatase 1 of Artemisia annua L. is a negative regulator of ABA signaling. Biomed Res. Int. 2014, 2014, 521794. [Google Scholar] [CrossRef]
- D’Ippólito, S.; Rey-Burusco, M.F.; Feingold, S.E.; Guevara, M.G. Role of proteases in the response of plants to drought. Plant Physiol. Biochem. 2021, 168, 1–9. [Google Scholar] [CrossRef]
- Meinhard, M.; Rodriguez, P.L.; Grill, E. The sensitivity of ABI2 to hydrogen peroxide links the abscisic acid-response regulator to redox signalling. Planta 2002, 214, 775–782. [Google Scholar] [CrossRef]
- Mittler, R. Oxidative stress, antioxidants and stress tolerance. Trends Plant Sci. 2002, 7, 405–410. [Google Scholar] [CrossRef]
- Liu, Y.; Song, Q.; Li, D.; Yang, X.; Li, D. Multifunctional roles of plant dehydrins in response to environmental stresses. Front. Plant Sci. 2017, 8, 1018. [Google Scholar] [CrossRef]
- Santos, A.B.; Mazzafera, P. Dehydrins Are Highly Expressed in Water-Stressed Plants of Two Coffee Species. Trop. Plant Biol. 2012, 5, 218–232. [Google Scholar] [CrossRef]
- Fernandes, I.; Marques, I.; Paulo, O.S.; Batista, D.; Partelli, F.L.; Lidon, F.C.; DaMatta, F.M.; Ramalho, J.C.; Ribeiro-Barros, A.I. Understanding the impact of drought in Coffea genotypes: Transcriptomic analysis supports a common high resilience to moderate water deficit but a genotype dependent sensitivity to severe water deficit. Agronomy 2021, 11, 2255. [Google Scholar] [CrossRef]
- Noctor, G.; Reichheld, J.P.; Foyer, C.H. ROS-related redox regulation and signaling in plants. Semin. Cell Dev. Biol. 2018, 80, 3–12. [Google Scholar] [CrossRef]
- Dumanović, J.; Nepovimova, E.; Natić, M.; Kuča, K.; Jaćević, V. The Significance of Reactive Oxygen Species and Antioxidant Defense System in Plants: A Concise Overview. Front. Plant Sci. 2021, 11, 552969. [Google Scholar] [CrossRef]
- Martins, M.Q.; Rodrigues, W.P.; Fortunato, A.S.; Leitão, A.E.; Rodrigues, A.P.; Pais, I.P.; Martins, L.D.; Silva, M.J.; Reboredo, F.H.; Partelli, F.L.; et al. Protective response mechanisms to heat stress in interaction with high [CO2] conditions in Coffea spp. Front. Plant Sci. 2016, 7, 947. [Google Scholar] [CrossRef] [PubMed]
- Rodrigues, W.P.; Martins, M.Q.; Fortunato, A.S.; Rodrigues, A.P.; Semedo, J.N.; Simões-Costa, M.C.; Pais, I.P.; Leitão, A.E.; Colwell, F.; Goulao, L.; et al. Long-term elevated air [CO2] strengthens photosynthetic functioning and mitigates the impact of supra-optimal temperatures in tropical Coffea arabica and C. canephora species. Glob. Chang. Biol. 2016, 22, 415–431. [Google Scholar] [CrossRef] [PubMed]
- Vinci, G.; Marques, I.; Rodrigues, A.P.; Martins, S.; Leitão, A.E.; Semedo, M.C.; Silva, M.J.; Lidon, F.C.; DaMatta, F.M.; Ribeiro-Barros, A.I.; et al. Protective Responses at the Biochemical and Molecular Level Differ between a Coffea arabica L. Hybrid and Its Parental Genotypes to Supra-Optimal Temperatures and Elevated Air [CO2]. Plants 2022, 11, 2702. [Google Scholar] [CrossRef]
- Fortunato, A.S.; Lidon, F.C.; Batista-Santos, P.; Eduardo Leitão, A.; Pais, I.P.; Ribeiro, A.I.; Cochicho Ramalho, J. Biochemical and molecular characterization of the antioxidative system of Coffea sp. under cold conditions in genotypes with contrasting tolerance. J. Plant Physiol. 2010, 167, 333–342. [Google Scholar] [CrossRef]
- Ramalho, J.C.; Fortunato, A.S.; Goulao, L.F.; Lidon, F.C. Cold-induced changes in mineral content in leaves of Coffea spp. Identification of descriptors for tolerance assessment. Biol. Plant. 2013, 57, 495–506. [Google Scholar] [CrossRef]
- Ramalho, J.C.; Campos, P.S.; Teixeira, M.; Nunes, M.A. Nitrogen dependent changes in antioxidant system and in fatty acid composition of chloroplast membranes from Coffea arabica L. plants submitted to high irradiance. Plant Sci. 1998, 135, 115–124. [Google Scholar] [CrossRef]
- Ramalho, J.C.; Pons, T.L.; Groeneveld, H.W.; Nunes, M.A. Photosynthetic responses of Coffea arabica leaves to a short-term high light exposure in relation to N availability. Physiol. Plant. 1997, 101, 229–239. [Google Scholar] [CrossRef]
- Miniussi, M.; Del Terra, L.; Savi, T.; Pallavicini, A.; Nardini, A. Aquaporins in Coffea arabica L.: Identification, expression, and impacts on plant water relations and hydraulics. Plant Physiol. Biochem. 2015, 95, 92–102. [Google Scholar] [CrossRef]
- Marques, I.; Fernandes, I.; Paulo, O.S.; Lidon, F.C.; DaMatta, F.M.; Ramalho, J.C.; Ribeiro-barros, A.I. A transcriptomic approach to understanding the combined impacts of supra-optimal temperatures and CO2 revealed different responses in the polyploid Coffea arabica and its diploid progenitor C. canephora. Int. J. Mol. Sci. 2021, 22, 3125. [Google Scholar] [CrossRef]
- Devnarain, N.; Crampton, B.G.; Olivier, N.; van der Westhuyzen, C.; Becker, J.V.W.; O’Kennedy, M.M. Transcriptomic analysis of a Sorghum bicolor landrace identifies a role for beta-alanine betaine biosynthesis in drought tolerance. S. Afr. J. Bot. 2019, 127, 244–255. [Google Scholar] [CrossRef]
- Aziz, M.A.; Sabeem, M.; Mullath, S.K.; Brini, F.; Masmoudi, K. Plant Group II LEA Proteins: Intrinsically Disordered Structure for Multiple Functions in Response to Environmental Stresses. Biomolecules 2021, 11, 1662. [Google Scholar] [CrossRef]
- Magwanga, R.O.; Lu, P.; Kirungu, J.N.; Lu, H.; Wang, X.; Cai, X.; Zhou, Z.; Zhang, Z.; Salih, H.; Wang, K.; et al. Characterization of the late embryogenesis abundant (LEA) proteins family and their role in drought stress tolerance in upland cotton. BMC Genet. 2018, 19, 6. [Google Scholar] [CrossRef]
- Buckley, T.N. How do stomata respond to water status? New Phytol. 2019, 224, 21–36. [Google Scholar] [CrossRef]
- Cui, L.; Yao, S.; Dai, X.; Yin, Q.; Liu, Y.; Jiang, X.; Wu, Y.; Qian, Y.; Pang, Y.; Gao, L.; et al. Identification of UDP-glycosyltransferases involved in the biosynthesis of astringent taste compounds in tea (Camellia sinensis). J. Exp. Bot. 2016, 67, 2285–2297. [Google Scholar] [CrossRef]
- Dong, N.-Q.; Sun, Y.; Guo, T.; Shi, C.-L.; Zhang, Y.-M.; Kan, Y.; Xiang, Y.-H.; Zhang, H.; Yang, Y.-B.; Li, Y.-C.; et al. UDP-glucosyltransferase regulates grain size and abiotic stress tolerance associated with metabolic flux redirection in rice. Nat. Commun. 2020, 11, 2629. [Google Scholar] [CrossRef]
- Jarzyniak, K.M.; Jasiński, M. Membrane transporters and drought resistance-A complex issue. Front. Plant Sci. 2014, 5, 687. [Google Scholar] [CrossRef]
- Ahmed, U.; Rao, M.J.; Qi, C.; Xie, Q.; Noushahi, H.A.; Yaseen, M.; Shi, X.; Zheng, B. Expression profiling of flavonoid biosynthesis genes and secondary metabolites accumulation in populus under drought stress. Molecules 2021, 26, 5546. [Google Scholar] [CrossRef]
- Watkins, J.M.; Hechler, P.J.; Muday, G.K. Ethylene-induced flavonol accumulation in guard cells suppresses reactive oxygen species and moderates stomatal aperture. Plant Physiol. 2014, 164, 1707–1717. [Google Scholar] [CrossRef]
- Takahama, U. Oxidation of vacuolar and apoplastic phenolic substrates by peroxidase: Physiological significance of the oxidation reactions. Phytochem. Rev. 2004, 3, 207–219. [Google Scholar] [CrossRef]
- Ferreres, F.; Figueiredo, R.; Bettencourt, S.; Carqueijeiro, I.; Oliveira, J.; Gil-Izquierdo, A.; Pereira, D.M.; Valentão, P.; Andrade, P.B.; Duarte, P.; et al. Identification of phenolic compounds in isolated vacuoles of the medicinal plant Catharanthus roseus and their interaction with vacuolar class III peroxidase: An H2O2 affair? J. Exp. Bot. 2011, 62, 2841–2854. [Google Scholar] [CrossRef] [Green Version]
- Marques, I.; Gouveia, D.; Gaillard, J.C.; Martins, S.; Semedo, M.C.; Lidon, F.C.; DaMatta, F.M.; Ribeiro-Barros, A.I.; Armengaud, J.; Ramalho, J.C. Next-Generation Proteomics Reveals a Greater Antioxidative Response to Drought in Coffea arabica than in Coffea Canephora. Agronomy 2022, 12, 148. [Google Scholar] [CrossRef]
- Gray, S.B.; Rodriguez-Medina, J.; Rusoff, S.; Toal, T.W.; Kajala, K.; Runcie, D.E.; Brady, S.M. Translational regulation contributes to the elevated CO2 response in two Solanum species. Plant J. 2020, 102, 383–397. [Google Scholar] [CrossRef] [PubMed]
- Shenton, D.; Smirnova, J.B.; Selley, J.N.; Carroll, K.; Hubbard, S.J.; Pavitt, G.D.; Ashe, M.P.; Grant, C.M. Global Translational Responses to Oxidative Stress Impact upon Multiple Levels of Protein Synthesis. J. Biol. Chem. 2006, 281, 29011–29021. [Google Scholar] [CrossRef]
- Ramalho, J.C.; Rodrigues, A.P.; Semedo, J.N.; Pais, I.P.; Martins, L.D.; Simões-Costa, M.C.; Leitão, A.E.; Fortunato, A.S.; Batista-Santos, P.; Palos, I.M.; et al. Sustained photosynthetic performance of Coffea spp. under long-term enhanced [CO2]. PLoS ONE 2013, 8, 82712. [Google Scholar] [CrossRef] [PubMed]
- Andrews, S. FastQC: A Quality Control Tool for High Throughput Sequence Data. 2010. Available online: http://www.bioinformatics.babraham.ac.uk/projects/fastqc (accessed on 4 April 2021).
- Wingett, S.W.; Andrews, S. FastQ Screen: A tool for multi-genome mapping and quality control. F1000Research 2018, 7, 1338. [Google Scholar] [CrossRef]
- Liao, Y.; Shi, W. Read trimming is not required for mapping and quantification of RNA-seq reads at the gene level. NAR Genom. Bioinforma. 2020, 2, lqaa068. [Google Scholar] [CrossRef]
- Denoeud, F.; Carretero-Paulet, L.; Dereeper, A.; Droc, G.; Guyot, R.; Pietrella, M.; Zheng, C.; Alberti, A.; Anthony, F.; Aprea, G.; et al. The coffee genome provides insight into the convergent evolution of caffeine biosynthesis. Science 2014, 345, 1181–1184. [Google Scholar] [CrossRef]
- Dobin, A.; Davis, C.A.; Schlesinger, F.; Drenkow, J.; Zaleski, C.; Jha, S.; Batut, P.; Chaisson, M.; Gingeras, T.R. STAR: Ultrafast universal RNA-seq aligner. Bioinformatics 2013, 29, 15–21. [Google Scholar] [CrossRef]
- Anders, S.; Pyl, P.T.; Huber, W. HTSeq—A Python framework to work with high-throughput sequencing data. Bioinformatics 2015, 31, 166. [Google Scholar] [CrossRef]
- Li, H.; Handsaker, B.; Wysoker, A.; Fennell, T.; Ruan, J.; Homer, N.; Marth, G.; Abecasis, G.; Durbin, R. The Sequence Alignment/Map format and SAMtools. Bioinformatics 2009, 25, 2078–2079. [Google Scholar] [CrossRef] [Green Version]
- Pertea, G.G. gffread: GFF/GTF Utility Providing Format Conversions, Region Filtering, FASTA Sequence Extraction and More. 2015. Available online: https://github.com/gpertea/gffread (accessed on 15 January 2021).
- Wickham, H. ggplot2: Elegant Graphics for Data Analysis; Springer: New York, NY, USA, 2016; ISBN 978-3-319-24277-4. [Google Scholar]
- Team, R.C. R: A Language and Environment for Statistical Computing; R Foundation for Statistical Computing: Vienna, Austria, 2018. [Google Scholar]
- Love, M.I.; Huber, W.; Anders, S. Moderated estimation of fold change and dispersion for RNA-seq data with DESeq2. Genome Biol. 2014, 15, 550. [Google Scholar] [CrossRef]
- Robinson, M.D.; McCarthy, D.J.; Smyth, G.K. edgeR: A Bioconductor package for differential expression analysis of digital gene expression data. Bioinformatics 2009, 26, 139–140. [Google Scholar] [CrossRef]
- Benjamini, Y.; Hochberg, Y. On the Adaptive Control of the False Discovery Rate in Multiple Testing With Independent Statistics. J. Educ. Behav. Stat. 2016, 25, 60–83. [Google Scholar] [CrossRef]
- Hunter, J.D. Matplotlib: A 2D graphics environment. Comput. Sci. Eng. 2007, 9, 90–95. [Google Scholar] [CrossRef]
- Thimm, O.; Bla È Sing, O.; Gibon, Y.; Nagel, A.; Meyer, S.; Kru È Ger, P.; Selbig, J.; Mu È Ller, L.A.; Rhee, S.Y.; Stitt, M. MAPMAN: A user-driven tool to display genomics data sets onto diagrams of metabolic pathways and other biological processes. Plant J. 2004, 37, 914–939. [Google Scholar] [CrossRef]
- Raudvere, U.; Kolberg, L.; Kuzmin, I.; Arak, T.; Adler, P.; Peterson, H.; Vilo, J. G: Profiler: A web server for functional enrichment analysis and conversions of gene lists (2019 update). Nucleic Acids Res. 2019, 47, W191–W198. [Google Scholar] [CrossRef]
- Supek, F.; Bošnjak, M.; Škunca, N.; Šmuc, T. Revigo summarizes and visualizes long lists of gene ontology terms. PLoS ONE 2011, 6, 21800. [Google Scholar] [CrossRef]
- Chen, C.; Huang, H.; Wu, C.H. Protein bioinformatics databases and resources. In Methods in Molecular Biology; Humana Press Inc.: Totowa, NJ, USA, 2017; Volume 1558, pp. 3–39. [Google Scholar] [CrossRef]
- Untergasser, A.; Cutcutache, I.; Koressaar, T.; Ye, J.; Faircloth, B.C.; Remm, M.; Rozen, S.G. Primer3-new capabilities and interfaces. Nucleic Acids Res. 2012, 40, e115. [Google Scholar] [CrossRef]
- Martins, M.Q.; Fortunato, A.S.; Rodrigues, W.P.; Partelli, F.L.; Campostrini, E.; Lidon, F.C.; DaMatta, F.M.; Ramalho, J.C.; Ribeiro-Barros, A.I. Selection and Validation of Reference Genes for Accurate RT-qPCR Data Normalization in Coffea spp. under a Climate Changes Context of Interacting Elevated [CO2] and Temperature. Front. Plant Sci. 2017, 8, 307. [Google Scholar] [CrossRef] [Green Version]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Gene ID | Protein Name | aCO2 | eCO2 | ||
|---|---|---|---|---|---|
| MWD | SWD | MWD | SWD | ||
| LOC113741996 | Late Embryogenesis Abundant protein Dc3-like | 5.51 | 7.19 | ||
| LOC113704200 | Homeobox-leucine zipper protein ATHB-12-like | 4.50 | 4.83 | 6.75 | |
| LOC113740436 | Late Embryogenesis Abundant protein Dc3-like | 3.58 | 6.10 | ||
| LOC113727829 | Galactinol synthase 2-like | 2.99 | 4.29 | 6.07 | |
| LOC113706564 | Protein phosphatase 2C 51-like | 4.43 | 6.06 | ||
| LOC113733000 | Galactinol synthase 2-like | 4.89 | 3.45 | 5.79 | |
| LOC113703008 | Protein phosphatase 2C 51-like | 5.62 | 5.54 | ||
| LOC113727830 | Galactinol synthase 2-like | 4.91 | 5.43 | ||
| LOC113743599 | NAC domain-containing protein 72-like | 3.84 | 5.41 | ||
| LOC113689826 | NAC domain-containing protein 72-like | 3.83 | 4.86 | ||
| LOC113740563 | HVA22-like protein e | 4.25 | 3.96 | 4.65 | |
| LOC113695700 | Protein LE25-like | 3.46 | 4.65 | ||
| LOC113692189 | Late Embryogenesis Abundant protein 46-like | 4.64 | |||
| LOC113740410 | HVA22-like protein e | 4.94 | 4.7 | 4.61 | |
| LOC113692190 | 18 kDa seed maturation protein-like | 4.53 | |||
| LOC113729974 | Acidic endochitinase SE2-like | 4.15 | |||
| LOC113740137 | Protein Aspartic Protease in Guard Cell 1-like | 4.05 | |||
| LOC113729880 | Homeobox-leucine zipper protein ATHB-12-like | 3.79 | 3.72 | 3.63 | |
| LOC113733488 | Late Embryogenesis Abundant protein-like | 3.62 | |||
| LOC113726525 | Zinc finger protein ZAT10-like | 4.46 | 3.54 | ||
| LOC113709223 | Aquaporin TIP1-3-like | −4.55 | −4.35 | −3.52 | |
| LOC113706149 | Probable LRR receptor-like serine/threonine-protein kinase At1g34110 | −4.01 | |||
| LOC113702113 | Serine/threonine-protein kinase SAPK1-like isoform X3 | −3.15 | −4.26 | ||
| LOC113742299 | Protein Aspartic Protease in Guard Cell 1-like | −4.16 | −7.11 | −4.32 | |
| LOC113733193 | Probable aquaporin PIP1-2 | −4.03 | |||
| LOC113716691 | Protein Aspartic Protease in Guard Cell 1-like | −2.96 | −4.37 | ||
| LOC113695827 | Pathogenesis-related protein PR-1-like | −4.83 | |||
| LOC113742753 | Aquaporin TIP2-1 | −3.25 | −5.97 | −4.38 | |
| LOC113741887 | Ethylene-responsive transcription factor WIN1-like | −4.46 | |||
| LOC113707187 | Aquaporin TIP4-1-like | −5.45 | −4.93 | ||
| LOC113740136 | Protein Aspartic Protease in Guard Cell 1-like | −3.8 | −6.26 | −4.93 | |
| LOC113732068 | Protein Eceriferum 1-like | −4.20 | −5.03 | ||
| LOC113735055 | Basic endochitinase-like | −4.17 | −7.54 | −5.58 | |
| LOC113742697 | Protein Aspartic Protease in Guard Cell 1-like | −3.81 | −6.63 | −5.62 | |
| LOC113716416 | Acidic endochitinase-like | −4.09 | −5.94 | ||
| LOC113701593 | PLAT domain-containing protein 3-like | −7.32 | −8.00 | −6.22 | |
| LOC113742441 | Protein Aspartic Protease in Guard Cell 1-like | −8.98 | |||
| LOC113737208 | Basic endochitinase-like | −4.41 | −7.12 | ||
| LOC113739398 | Ethylene-responsive transcription factor WIN1-like | −7.71 | |||
| LOC113697821 | Probable xyloglucan endotransglucosylase/hydrolase protein 6 | −4.72 | −12.31 | −11.17 | |
| Gene ID | Protein Name | aCO2 | eCO2 | ||
|---|---|---|---|---|---|
| MWD | SWD | MWD | SWD | ||
| Cc07_g07560 | Probable xyloglucan endotransglucosylase/hydrolase protein 6 | −6.90 | −5.85 | ||
| Cc04_g09640 | Protein Aspartic Protease in Guard Cell 1-like | −5.03 | −4.92 | ||
| Cc04_g07360 | Putative protein Aspartic Protease in Guard Cell 1-like | 4.89 | 4.07 | 3.96 | |
| Cc04_g07380 | Putative protein Aspartic Protease in Guard Cell 1-like | 4.27 | 3.17 | 2.80 | |
| Cc07_g10030 | Dehydrin DH1a | 6.05 | 5.47 | 2.37 | |
| Cc01_g11790 | Mitogen-activated protein kinase 3 | 1.45 | |||
| Cc06_g09540 | Multiprotein-bridging factor 1c | 2.34 | |||
| Cc04_g08280 | Putative movement protein binding protein 2C | −2.86 | −1.92 | ||
| Cc06_g15980 | 18 kDa seed maturation protein | 9.18 | 8.79 | 2.92 | |
| Cc04_g07350 | Putative protein Aspartic Protease in Guard Cell 1-like | −5.86 | |||
| Cc04_g07330 | Putative protein Aspartic Protease in Guard Cell 1-like | −7.93 | −8.16 | ||
| Cc02_g17270 | Putative Late Embryogenesis Abundant protein | 4.32 | 3.58 | ||
| Cc01_g08980 | Late Embryogenesis Abundant protein hydroxyproline-rich glycoprotein family | −3.25 | |||
| Cc04_g07370 | Putative protein Aspartic Protease in Guard Cell 1-like | −3.48 | |||
| Cc02_g15480 | Cellulose synthase A catalytic subunit 8 [UDP-forming] | −5.28 | |||
| Cc07_g15660 | C2 domain-containing protein | −1.73 | |||
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Marques, I.; Fernandes, I.; Paulo, O.S.; Batista, D.; Lidon, F.C.; Partelli, F.; DaMatta, F.M.; Ribeiro-Barros, A.I.; Ramalho, J.C. Overexpression of Water-Responsive Genes Promoted by Elevated CO2 Reduces ROS and Enhances Drought Tolerance in Coffea Species. Int. J. Mol. Sci. 2023, 24, 3210. https://doi.org/10.3390/ijms24043210
Marques I, Fernandes I, Paulo OS, Batista D, Lidon FC, Partelli F, DaMatta FM, Ribeiro-Barros AI, Ramalho JC. Overexpression of Water-Responsive Genes Promoted by Elevated CO2 Reduces ROS and Enhances Drought Tolerance in Coffea Species. International Journal of Molecular Sciences. 2023; 24(4):3210. https://doi.org/10.3390/ijms24043210
Chicago/Turabian StyleMarques, Isabel, Isabel Fernandes, Octávio S. Paulo, Dora Batista, Fernando C. Lidon, Fábio Partelli, Fábio M. DaMatta, Ana I. Ribeiro-Barros, and José C. Ramalho. 2023. "Overexpression of Water-Responsive Genes Promoted by Elevated CO2 Reduces ROS and Enhances Drought Tolerance in Coffea Species" International Journal of Molecular Sciences 24, no. 4: 3210. https://doi.org/10.3390/ijms24043210