


Mechanisms of Kale (Brassica oleracea var. acephala) Tolerance to Individual and Combined Stresses of Drought and Elevated Temperature
 , , and
, , and 
Abstract
:1. Introduction
2. Results
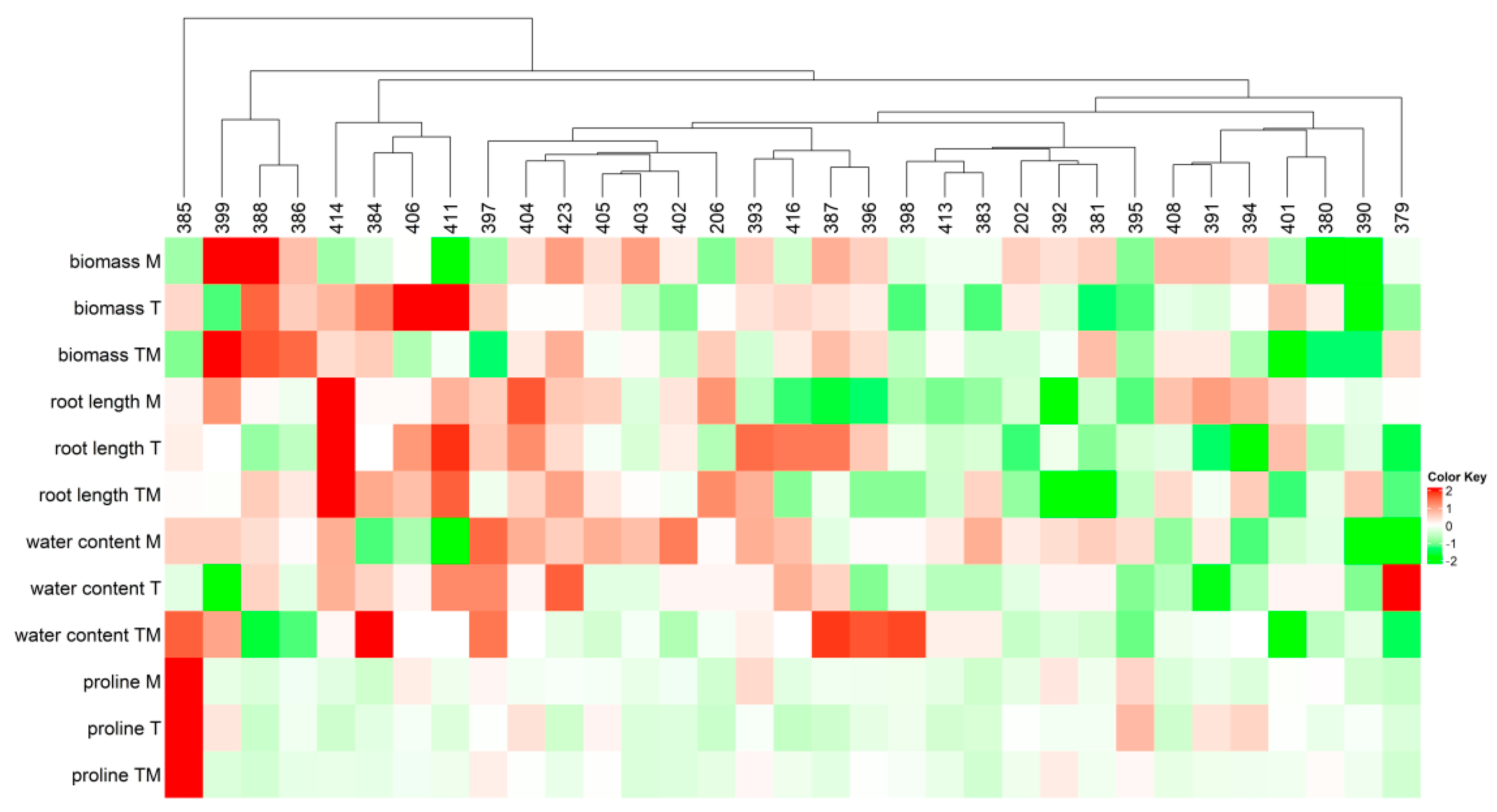
2.1. Screening of Kale Accessions to Abiotic Stresses
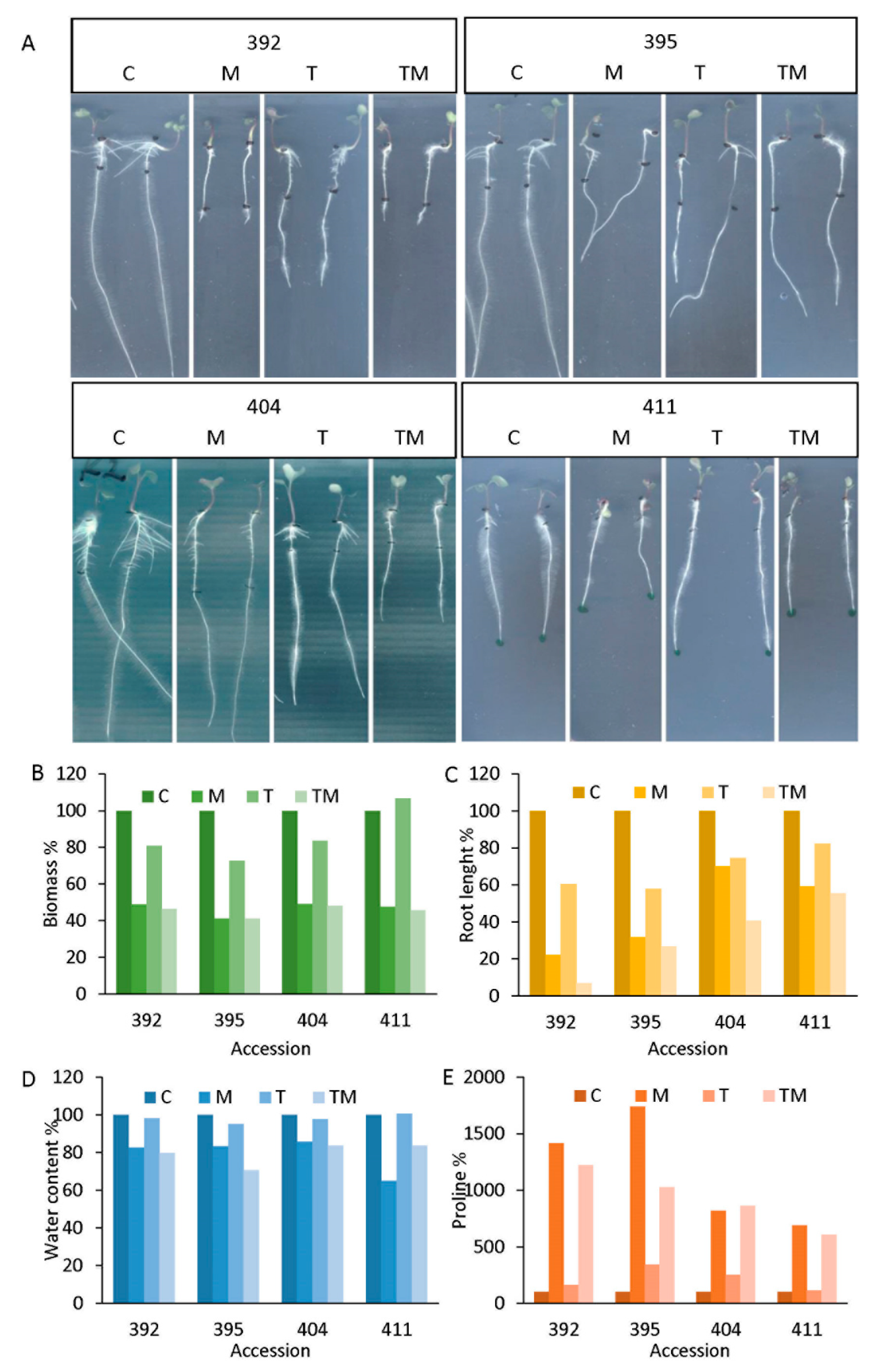
2.2. Selection of Tolerant and Sensitive Accessions
2.3. Selection of Tolerant and Sensitive Accessions
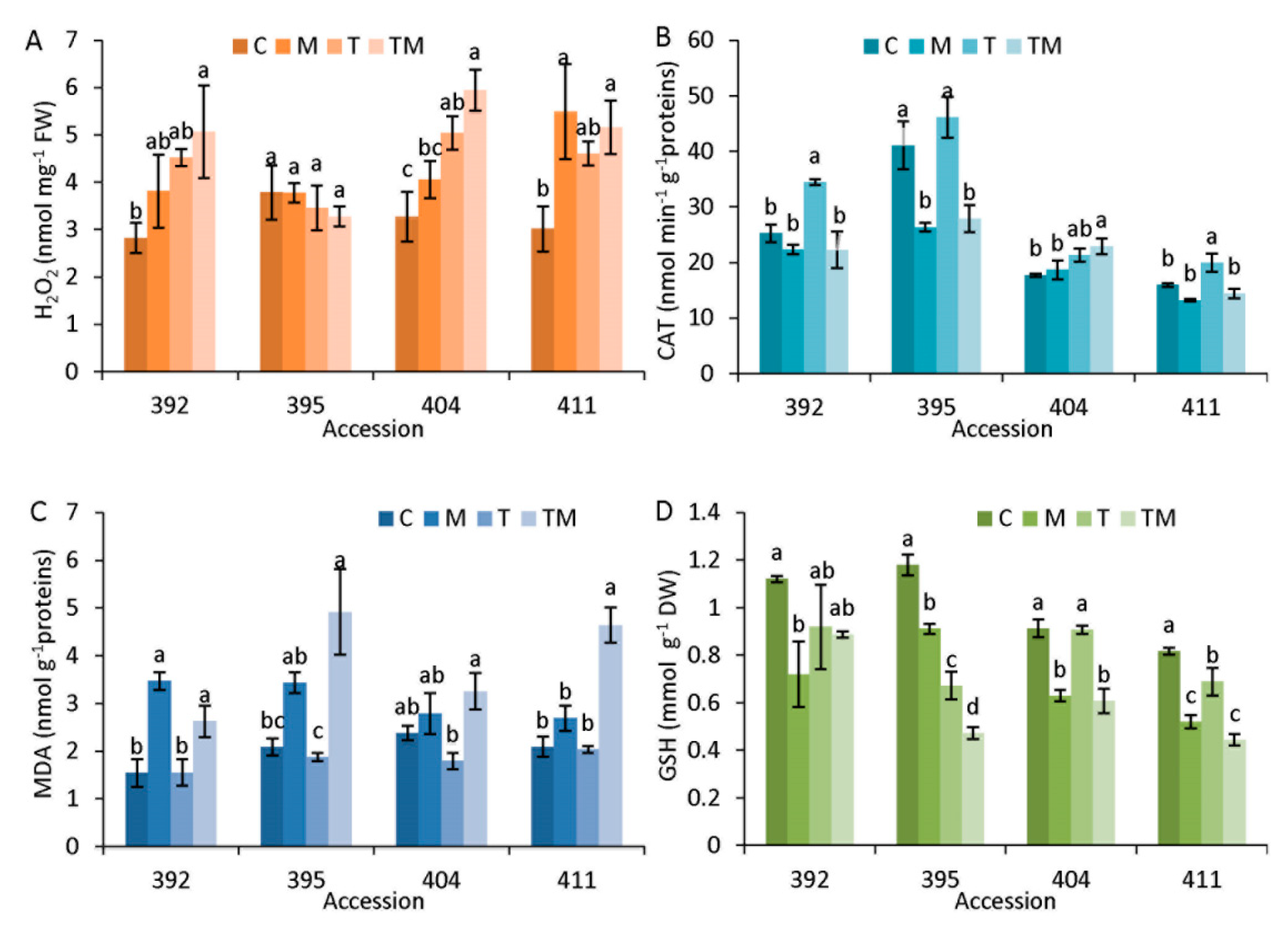
2.4. Stress Markers
2.5. Specialized Metabolites and Antioxidant Activity
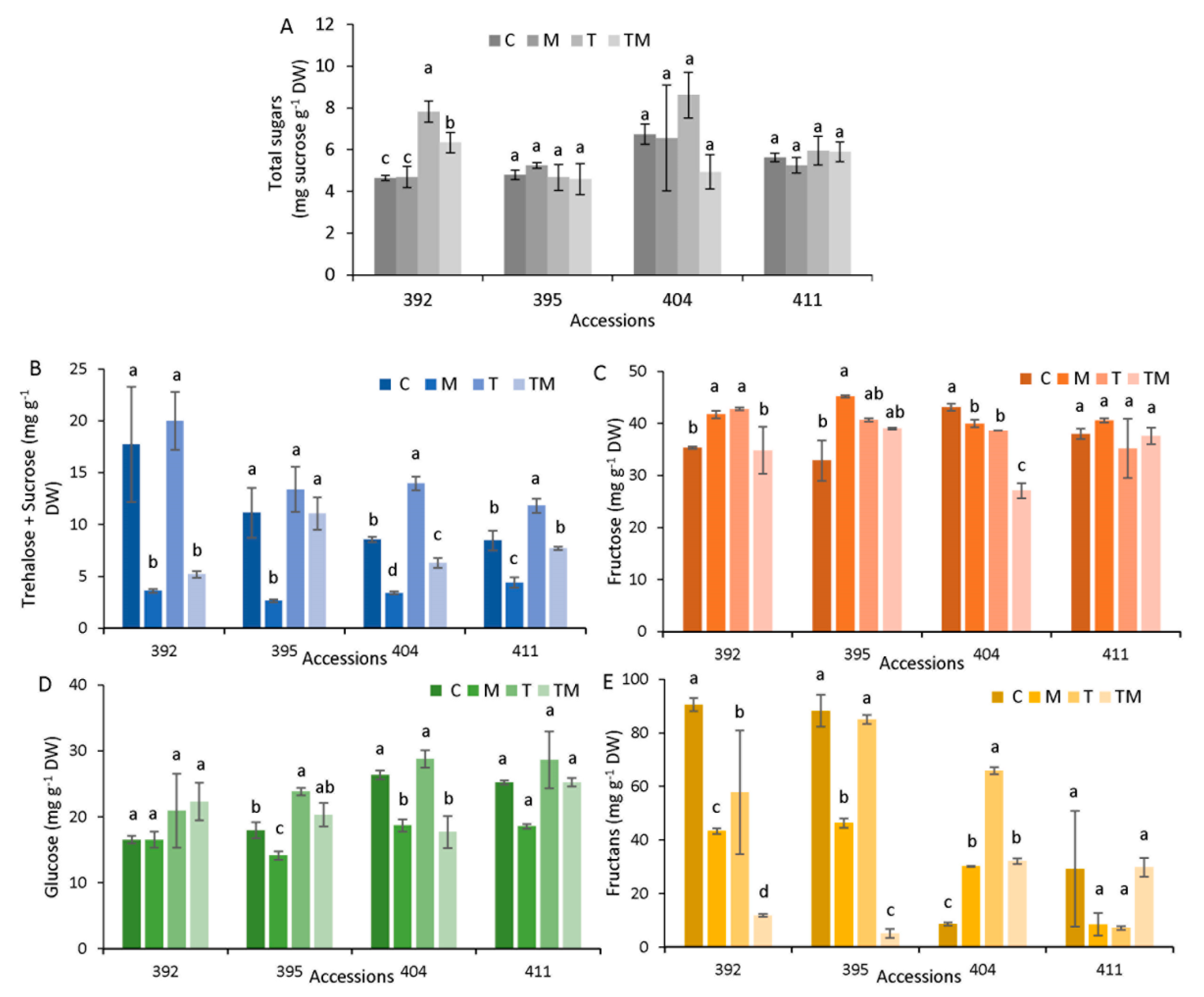
2.6. Sugar Analysis
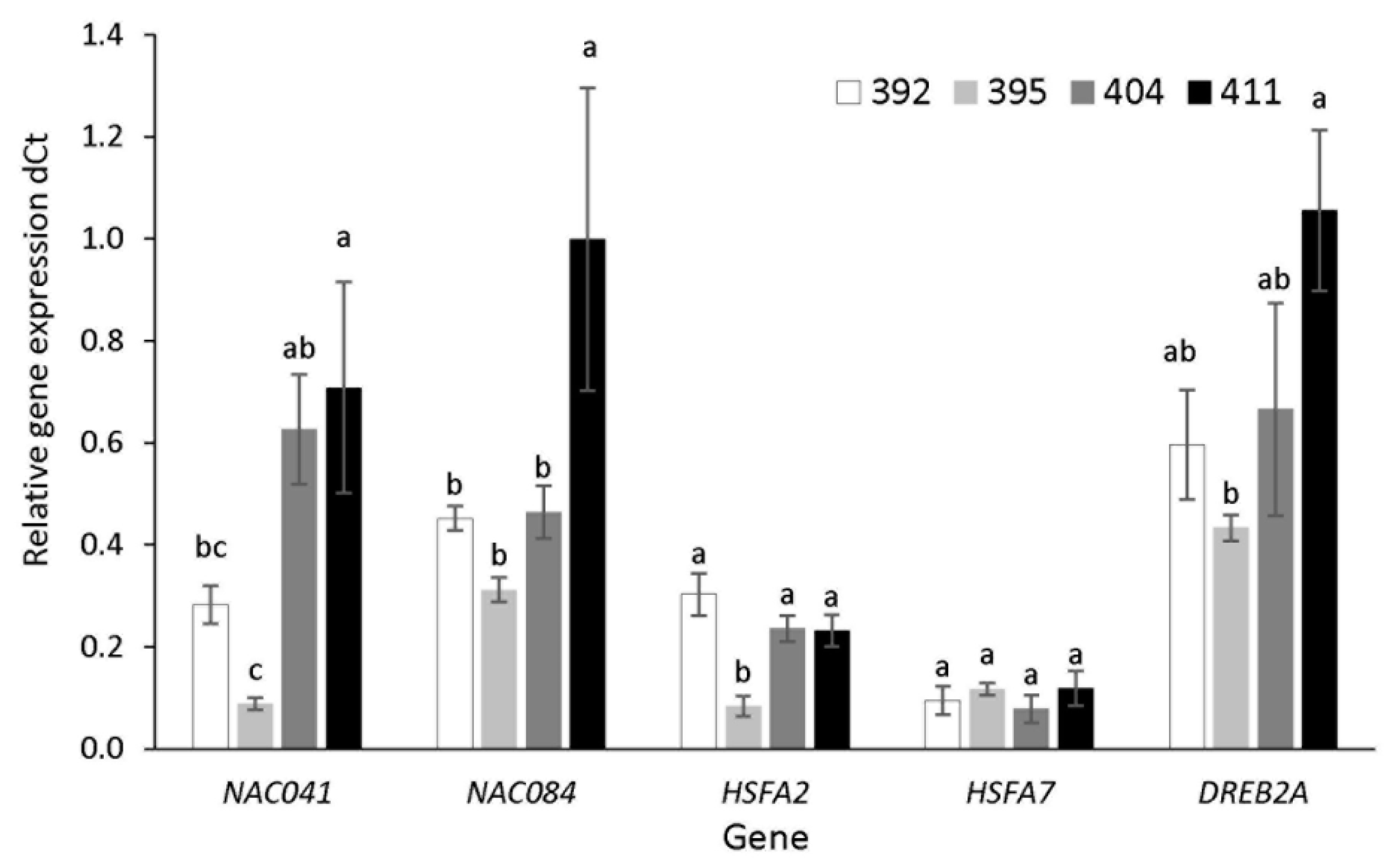
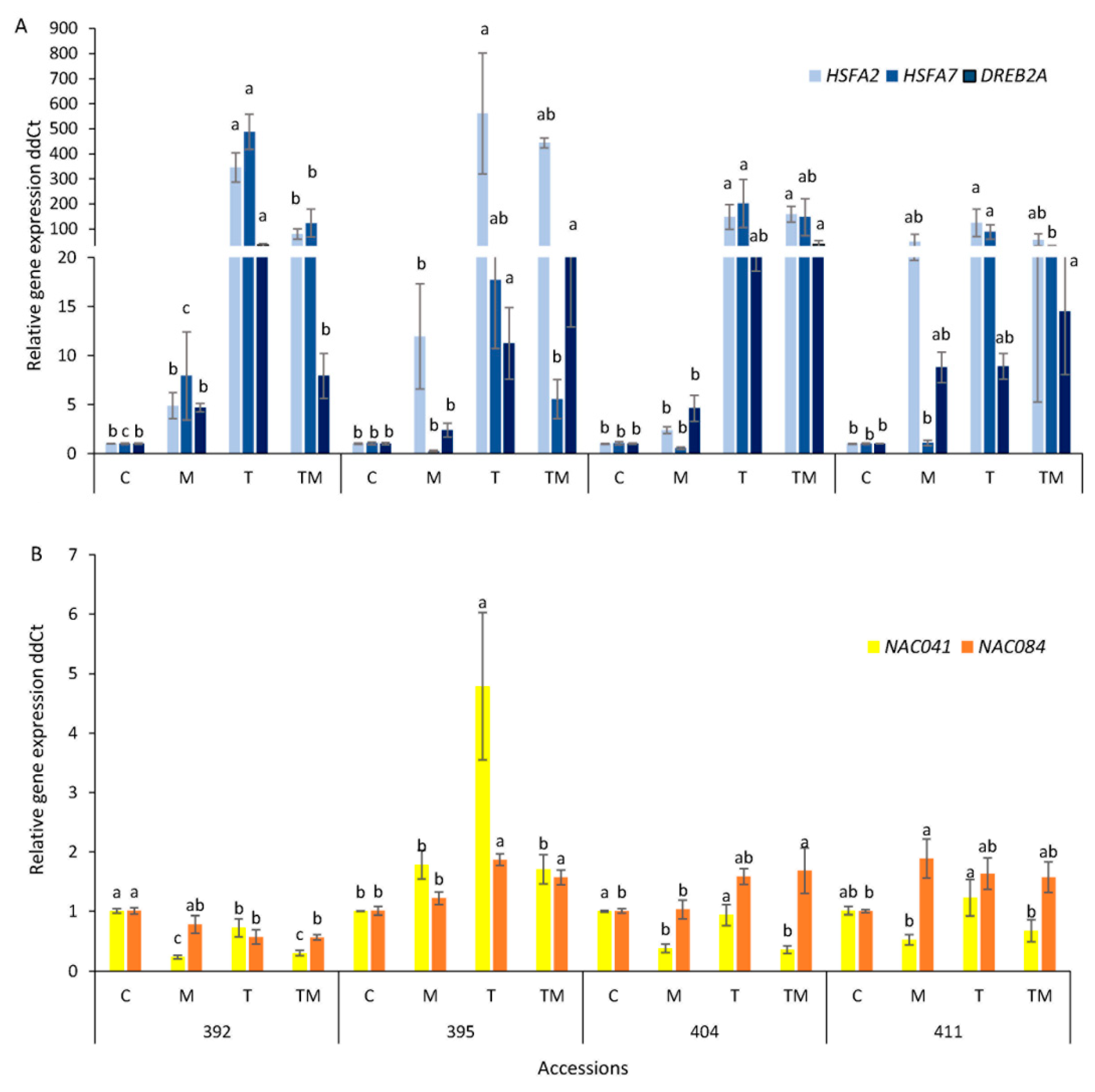
2.7. Gene Expression Analysis
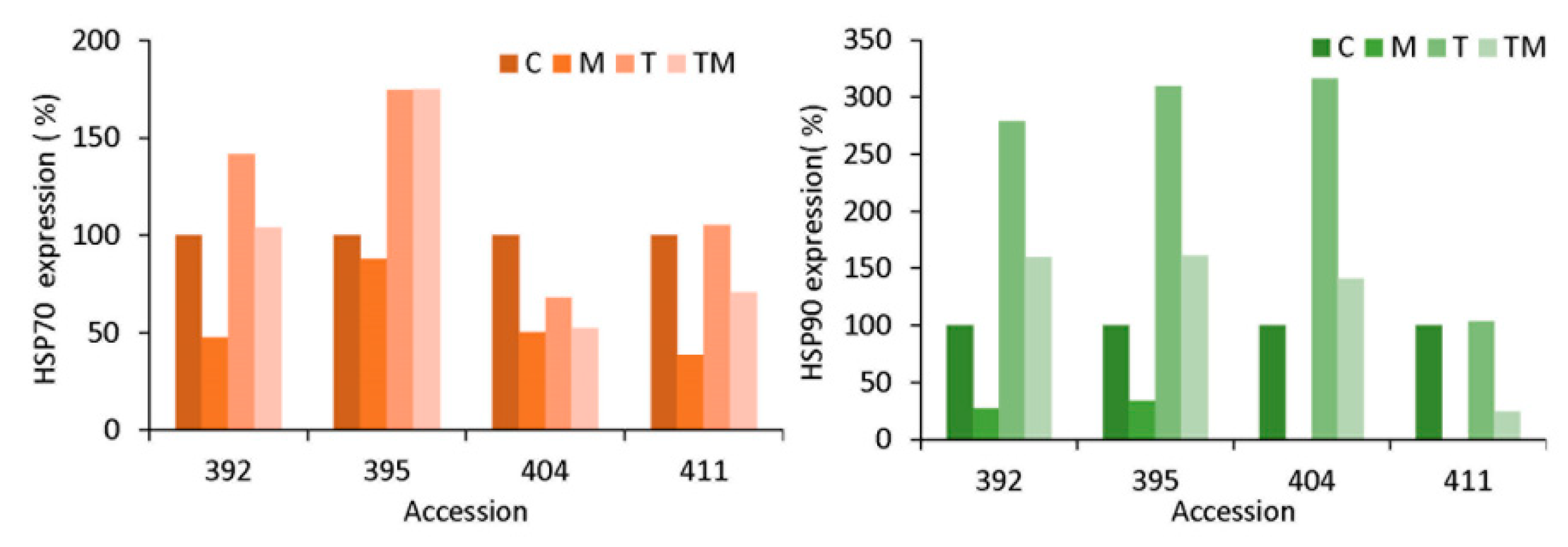
2.8. Protein Immmunodetection
2.9. Stress Response: Interactions of Accession and Stress Treatment
3. Discussion
3.1. Stress Markers
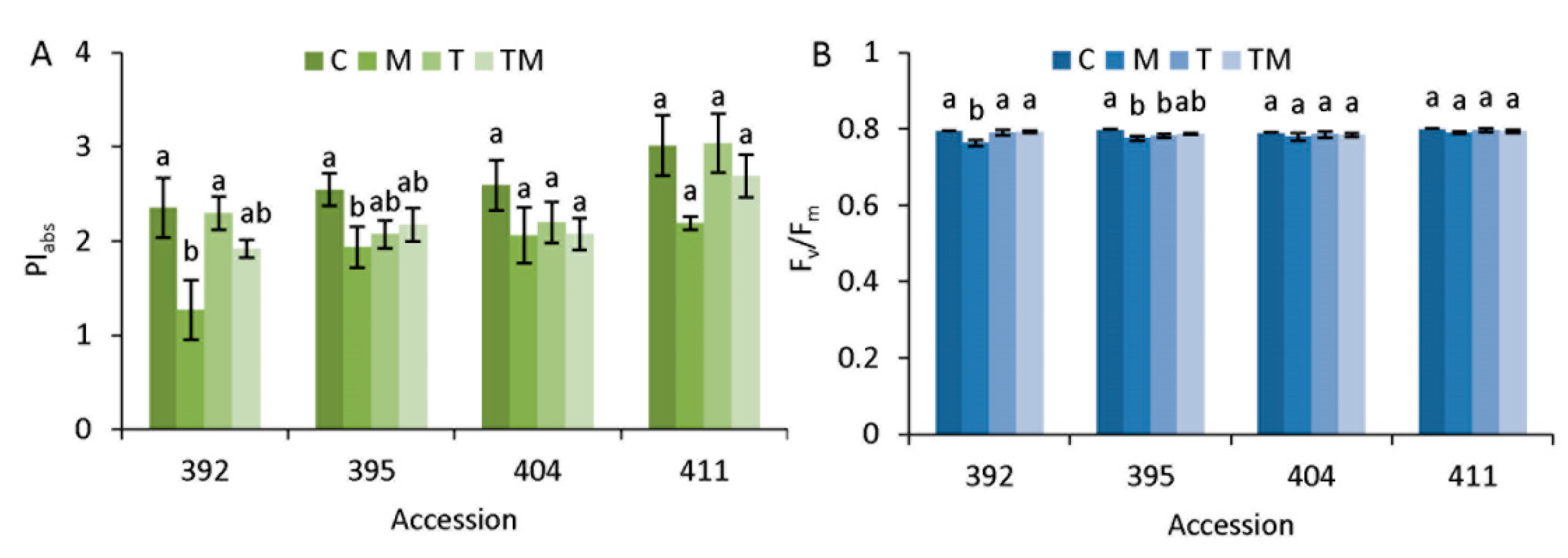
3.2. Photosynthetic Performance in Stress
3.3. Specialized Metabolite Accumulation under Stress Conditions
3.4. Soluble Sugar Fluctuations under Stress Conditions
3.5. Stress-Related Transcription Factor Gene Expression and Heat Shock Protein Accumulation
4. Materials and Methods
4.1. Plant Material
4.2. Plant Growth and Abiotic Stress Treatments
4.3. Photosynthetic Measurements
4.4. Biochemical Stress Markers
4.5. Protein Immmunodetection
4.6. Specialized Metabolites, Antioxidant Capacity and GSH
4.7. Sugar Analysis
4.8. Quantitative Real-Time PCR (RT-PCR) Analysis
4.9. Statistical Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Conflicts of Interest
References
- Chen, S.; Nelson, M.N.; Chèvre, A.-M.; Jenczewski, E.; Li, Z.; Mason, A.S. Trigenomic bridges for Brassica improvement. Crit. Rev. Plant Sci. 2011, 30, 524–547. [Google Scholar] [CrossRef]
- Šamec, D.; Pavlović, I.; Salopek-Sondi, B. White cabbage (Brassica oleracea var. capitata f. alba): Botanical, phytochemical and pharmacologicall overwiev. Phytochem. Rev. 2017, 16, 117–135. [Google Scholar] [CrossRef]
- Šamec, D.; Urlić, B.; Salopek-Sondi, B. Kale (Brassica oleracea var. acephala) as a superfood: Review of the scientific evidence behind the statement. Crit. Rev. Food Sci. Nutr. 2019, 59, 2411–2422. [Google Scholar] [CrossRef] [PubMed]
- Leisner, C.P. Review: Climate change impacts on food security- focus on perennial cropping systems and nutritional value. Plant Sci. 2020, 293, 110412. [Google Scholar] [CrossRef] [PubMed]
- Hassan, M.U.; Chattha, M.U.; Khan, I.; Chattha, M.B.; Barbanti, L.; Aamer, M.; Iqbal, M.M.; Nawaz, M.; Mahmood, A.; Ali, A.; et al. Heat stress in cultivated plants: Nature, impact, mechanisms, and mitigation strategies—A review. Plant Biosyst. 2021, 155, 211–234. [Google Scholar] [CrossRef]
- Corwin, D.L. Climate change impacts on soil salinity in agricultural areas. Eur. J. Soil Sci. 2021, 72, 842–862. [Google Scholar] [CrossRef]
- Kaushal, N.; Bhandari, K.; Kadambot, H.M.; Siddique, K.H.M.; Nayyar, H. Food crops face rising temperatures: An overview of responses, adaptive mechanisms, and approaches to improve heat tolerance. Cogent Food Agric. 2016, 2, 1134380. [Google Scholar] [CrossRef]
- Hirayama, T.; Shinozaki, K. Research on plant abiotic stress responses in the post-genome era: Past, present and future. Plant J. 2010, 61, 1041–1052. [Google Scholar] [CrossRef]
- Yoon, Y.; Seo, D.H.; Shin, H.; Kim, H.J.; Kim, C.M.; Jang, G. The Role of Stress-Responsive Transcription Factors in Modulating Abiotic Stress Tolerance in Plants. Agronomy 2020, 10, 788. [Google Scholar] [CrossRef]
- Nakashima, K.; Takasaki, H.; Mizoi, J.; Shinozaki, K.; Yamaguchi-Shinozaki, K. NAC transcription factors in plant abiotic stress responses. Biochim. Biophys. Acta 2012, 1819, 97–103. [Google Scholar] [CrossRef]
- Pavlović, I.; Petřík, I.; Tarkowska, D.; Lepeduš, H.; Vujčić, V.; Radić Brkanac, S.; Novák, O.; Salopek-Sondi, B. Correlations between phytohormones and drought tolerance in selected Brassica crops: Chinese cabbage, white cabbage and kale. Int. J. Mol. Sci. 2018, 19, 2866. [Google Scholar] [CrossRef] [PubMed]
- Pavlović, I.; Mlinarić, S.; Tarkowská, D.; Oklestková, J.; Novák, O.; Lepeduš, H.; Vujčić Bok, V.; Radić Brkanac, S.; Strnad, M.; Salopek-Sondi, B. Early Brassica crops responses to salinity stress: A Comparative Analysis between Chinese cabbage, White cabbage and Kale. Front. Plant Sci. 2019, 10, 450. [Google Scholar] [CrossRef] [PubMed]
- USDA; National Agricultural Statistics Service. Census of Agriculture. 2012. Available online: https://www.agcensus.usda.gov/Publications/2012/Full_Report/Volume_1,_Chapter_1_US/st99_1_038_038.pdf (accessed on 18 June 2015).
- Batelja, K.; Goreta Ban, S.; Žanić, K.; Miloš, B.; Dumičić, G.; Matotan, Z. Svojstva autohtonih populacija raštike (Brassica oleracea L. var. acephala) hrvatskog priobalja. Poljoprivreda 2009, 2, 1–7. [Google Scholar]
- Agrimonti, C.; Lauro, M.; Visioli, G. Smart agriculture for food quality: Facing climate change in the 21st century. Crit. Rev. Food Sci. Nutr. 2021, 61, 971–981. [Google Scholar] [CrossRef] [PubMed]
- Linić, I.; Šamec, D.; Grúz, J.; Vujčić Bok, V.; Strnad, M.; Salopek-Sondi, B. Involvement of phenolic acids in short-term adaptation to salinity stress is species specific among Brassicaceae. Plants 2019, 8, 155. [Google Scholar] [CrossRef] [PubMed]
- Pavlović, I.; Pěnčík, A.; Novák, O.; Vujčić, V.; Radić Brkanac, S.; Lepeduš, H.; Strnad, M.; Salopek-Sondi, B. Short-term salinity stress response of Brassica rapa caused changes in auxin metabolism. Plant Physiol. Biochem. 2018, 125, 74–84. [Google Scholar] [CrossRef]
- Beacham, A.M.; Hand, P.; Pink, D.A.C.; Monaghan, J.M. Analysis of Brassica oleracea early stage abiotic stress responses reveals tolerance in multiple crop types and for multiple sources of stress. J. Sci. Food Agric. 2017, 97, 5271–5277. [Google Scholar] [CrossRef]
- Lohani, N.; Jain, D.; Singh, M.B.; Bhalla, P.L. Engineering multiple abiotic stress tolerance in Canola, Brassica napus. Front. Plant Sci. 2020, 11, 3. [Google Scholar] [CrossRef]
- Singh, B.; Sran, A.S.; Sohi, G.S. Innovative strategies to develop abiotic and biotic stress tolerance in Mustard (Brassicaceae). In Brassica Breeding and Biotechnology; Islam, A.K.M.A., Hossain, M.A., Islam, A.K.M.M., Eds.; IntechOpen: London, UK, 2021; Available online: https://www.intechopen.com/chapters/75023 (accessed on 25 May 2022). [CrossRef]
- Dikšaitytė, A.; Viršilė, A.; Žaltauskaitė, J.; Januškaitienė, I.; Juozapaitienė, G. Growth and photosynthetic responses in Brassica napus differ during stress and recovery periods when exposed to combined heat, drought and elevated CO2. Plant Physiol. Biochem. 2019, 142, 59–72. [Google Scholar] [CrossRef]
- Zhang, H.; Zhao, Y.; Zhu, J.-K. Thriving under stress: How plants balance growth and the stress response. Dev. Cell 2020, 55, 529–543. [Google Scholar] [CrossRef]
- Petrić, I.; Šamec, D.; Karalija, E.; Salopek-Sondi, B. Beneficial microbes and molecules for mitigation of soil salinity in Brassica species: A Review. Soil Syst. 2022, 6, 18. [Google Scholar] [CrossRef]
- Hayat, S.; Mir, B.A.; Wani, A.S.; Hasan, S.A.; Irfan, M.; Ahmad, A. Screening of salt-tolerant genotypes of Brassica juncea based on photosynthetic attributes. J. Plant. Interact. 2011, 6, 53–60. [Google Scholar] [CrossRef]
- Mohamed, I.A.A.; Shalby, N.; Bai, C.; Qin, M.; Agami, R.A.; Jie, K.; Wang, B.; Zhou, G. Stomatal and photosynthetic traits are associated with investigating sodium chloride tolerance of Brassica napus L. cultivars. Plants 2020, 9, 62. [Google Scholar] [CrossRef] [PubMed]
- Hossain, M.A.; Bhattacharjee, S.; Armin, S.-M.; Qian, P.; Xin, W.; Li, H.-Y.; Burritt, D.J.; Fujita, M.; Tran, L.-S.P. Hydrogen peroxide priming modulates abiotic oxidative stress tolerance: Insights from ROS detoxification and scavenging. Front. Plant Sci. 2015, 6, 420. [Google Scholar] [CrossRef]
- Liu, W.-C.; Song, R.-F.; Zheng, S.-Q.; Zhang, B.-L.; Gao, X.; Lu, Y.-T. Coordination of plant growth and abiotic stress responses by tryptophan synthase β subunit 1 through modulation of tryptophan and ABA homeostasis in Arabidopsis. Mol. Plant 2022, 15, 973–990. [Google Scholar] [CrossRef]
- Liu, W.-C.; Song, R.-F.; Qiu, Y.-M.; Zheng, S.-Q.; Li, T.-T.; Wu, Y.; Song, C.-P.; Lu, Y.-T.; Yuan, H.-M. Sulfenylation of ENOLASE2 facilitates H2O2-conferred freezing tolerance in Arabidopsis. Dev. Cell 2022, 57, 1883–1898. [Google Scholar] [CrossRef]
- Khan, A.; Anwar, Y.; Hasan, M.M.; Iqbal, A.; Ali, M.; Alharby, H.F.; Hakeem, K.R.; Hasanuzzaman, M. Attenuation of drought stress in Brassica seedlings with exogenous application of Ca2+ and H2O2. Plants 2017, 6, 20. [Google Scholar] [CrossRef]
- Lee, B.-R.; La, V.H.; Park, S.-H.; Mamun, M.A.; Bae, D.-W.; Kim, T.-H. H2O2—Responsive hormonal status involves oxidative burst signaling and proline metabolism in rapeseed leaves. Antioxidants 2022, 11, 566. [Google Scholar] [CrossRef]
- Kalaji, H.M.; Jajoo, A.; Oukarroum, A.; Brestic, M.; Zivcak, M.; Samborska, I.A.; Cetner, M.D.; Łukasik, I.; Goltsev, V.; Ladle, R.J. Chlorophyll a fluorescence as a tool to monitor physiological status of plants under abiotic stress conditions. Acta Physiol. Plant. 2016, 38, 3–11. [Google Scholar] [CrossRef]
- Moradpour, M.; Abdullah, S.N.A.; Namasivayam, P. The impact of heat stress on morpho-physiological response and expression of specific genes in the heat stress-responsive transcriptional regulatory network in Brassica oleracea. Plants 2021, 10, 1064. [Google Scholar] [CrossRef]
- Rodríguez, V.M.; Soengas, P.; Alonso-Villaverde, V.; Sotelo, T.; Cartea, M.E.; Velasco, P. Effect of temperature stress on the early vegetative development of Brassica oleracea L. BMC Plant Biol. 2015, 15, 145. [Google Scholar] [CrossRef] [PubMed]
- Soengas, P.; Rodríguez, V.M.; Velasco, P.; Cartea, M.E. Effect of temperature stress on antioxidant defenses in Brassica oleracea. ACS Omega 2018, 3, 5237–5243. [Google Scholar] [CrossRef] [PubMed]
- Huang, R.; Liu, Z.; Xing, M.; Yang, Y.; Wu, X.; Liu, H.; Liang, W. Heat stress suppresses Brassica napus seed oil accumulation by inhibition of photosynthesis and BnWRI1 pathway. Plant Cell Physiol. 2019, 60, 1457–1470. [Google Scholar] [CrossRef] [PubMed]
- Elferjani, R.; Soolanayakanahally, R. Canola responses to drought, heat, and combined stress: Shared and specific effects on carbon assimilation, seed yield, and oil composition. Front. Plant Sci. 2018, 9, 1224. [Google Scholar] [CrossRef] [PubMed]
- Šamec, D.; Karalija, E.; Šola, I.; Vujčić Bok, V.; Salopek-Sondi, B. The role of polyphenols in abiotic stress response: The influence of molecular structure. Plants 2021, 10, 118. [Google Scholar] [CrossRef]
- Kourani, M.; Mohareb, F.; Rezwan, F.I.; Anastasiadi, M.; Hammond, J.P. Genetic and physiological responses to heat stress in Brassica napus. Front. Plant Sci. 2022, 13, 832147. [Google Scholar] [CrossRef]
- Essoh, A.P.; Monteiro, F.; Pena, A.R.; Pais, M.S.; Moura, M.; Romeiras, M.M. Exploring glucosinolates diversity in Brassicaceae: A genomic and chemical assessment for deciphering abiotic stress tolerance. Plant Physiol. Biochem. 2020, 150, 151–161. [Google Scholar] [CrossRef]
- Fareen, S.; Mohammad, Y.; Mohammad, F.; Ahmad, F.; Shamsul, H. Role of sugars under abiotic stress. Plant Physiol. Biochem. 2016, 109, 54–61. [Google Scholar] [CrossRef]
- Viana, V.E.; Aranha, B.C.; Busanello, C.; Maltzahn, L.E.; Panozzo, L.E.; De Oliveira, A.C.; Rombaldi, C.V.; Pegoraro, C. Metabolic profile of canola (Brassica napus L.) seedlings under hydric, osmotic and temperature stresses. Plant Stress 2022, 3, 100059. [Google Scholar] [CrossRef]
- Benkeblia, N. Insights on fructans and resistance of plants to drought stress. Front. Sustain. Food Syst. 2022, 6, 827758. [Google Scholar] [CrossRef]
- Keller, K.; Rodrigues, C.M.; Neuhaus, H.E.; Pommerrenig, B. Improved resource allocation and stabilization of yield under abiotic stress. J. Plant Physiol. 2021, 257, 153336. [Google Scholar] [CrossRef] [PubMed]
- Priya, M.; Dhanker, O.P.; Siddique, K.H.M.; Rao, B.H.; Nair, R.M.; Pandey, S.; Singh, S.; Varshney, R.K.; Prasad, P.V.V.; Nayyar, H. Drought and heat stress-related proteins: An update about their functional relevance in imparting stress tolerance in agricultural crops. Theor. Appl. Genet. 2019, 132, 1607–1638. [Google Scholar] [CrossRef] [PubMed]
- Guo, M.; Liu, J.-H.; Ma, X.; Luo, D.-X.; Gong, Z.-H.; Lu, M.-H. The Plant Heat Stress Transcription Factors (HSFs): Structure, regulation, and function in response to abiotic stresses. Front. Plant Sci. 2016, 7, 114. [Google Scholar] [CrossRef]
- Lohani, N.; Golicz, A.A.; Singh, M.B.; Prem, L.; Bhalla, P.L. Genome-wide analysis of the Hsf gene family in Brassica oleracea and a comparative analysis of the Hsf gene family in B. oleracea, B. rapa and B. napus. Funct. Integr. Genomics 2019, 19, 515–531. [Google Scholar] [CrossRef]
- Zhu, X.; Wang, Y.; Liu, Y.; Zhou, W.; Yan, B.; Yang, J.; Shen, Y. Overexpression of BcHsfA1 transcription factor from Brassica campestris improved heat tolerance of transgenic tobacco. PLoS ONE 2018, 13, e0207277. [Google Scholar] [CrossRef]
- Dong, X.; Yi, H.; Lee, J.; Nou, I.-S.; Han, C.-T.; Hur, Y. Global gene-expression analysis to identify differentially expressed genes critical for the heat stress response in Brassica rapa. PLoS ONE 2015, 10, e0130451. [Google Scholar] [CrossRef] [PubMed]
- Hasanuzzaman, M.; Nahar, K.; Alam, M.M.; Roychowdhury, R.; Fujita, M. Physiological, biochemical, and molecular mechanisms of heat stress tolerance in plants. Int. J. Mol. Sci. 2013, 14, 9643–9684. [Google Scholar] [CrossRef] [PubMed]
- Mishra, D.; Shekhar, S.; Singh, D.; Chakraborty, S.; Chakraborty, N. Heat shock proteins and abiotic stress tolerance in plants. In Regulation of Heat Shock Protein Responses; Asea, A.A.A., Kaur, P., Eds.; Springer: Dordrecht, The Netherlands, 2018. [Google Scholar] [CrossRef]
- Haq, S.; Khan, A.; Ali, M.; Khattak, A.M.; Gai, W.-X.; Zhang, H.-X.; Wei, A.M.; Gong, Z.-H. Heat shock proteins: Dynamic biomolecules to counter plant biotic and abiotic stresses. Int. J. Mol. Sci. 2019, 20, 5321. [Google Scholar] [CrossRef]
- Liu, T.; Song, X.; Duan, W.; Huang, Z.; Liu, G.; Li, Y.; Hou, X. Genome-wide analysis and expression patterns of NAC Transcription Factor Family under different developmental stages and abiotic stresses in Chinese Cabbage. Plant Mol. Biol. Rep. 2014, 32, 1041–1056. [Google Scholar] [CrossRef]
- Zhong, H.; Guo, Q.-Q.; Chen, L.; Ren, F.; Wang, Q.-Q.; Zheng, Y.; Li, X.-B. Two Brassica napus genes encoding NAC transcription factors are involved in response to high-salinity stress. Plant Cell Rep. 2012, 31, 1991–2003. [Google Scholar] [CrossRef]
- Ying, L.; Chen, H.; Cai, W. BnNAC485 is involved in abiotic stress responses andflowering time in Brassica napus. Plant Physiol. Biochem. 2014, 79, 77–87. [Google Scholar] [CrossRef] [PubMed]
- Strasser, R.J.; Tsimilli-Michael, M.; Srivastava, A. Analysis of the chlorophyll a fluorescence transient. In Chlorophyll a Fluorescence: A Signature of Photosynthesis; Papageorgiou, G.C., Govinjee, Eds.; Springer: Dordrecht, The Netherlands, 2004; pp. 321–362. [Google Scholar] [CrossRef]
- Mátai, A.; Hideg, É. A comparison of colorimetric assays detecting hydrogen peroxide in leaf extracts. Anal. Methods 2017, 9, 2357–2360. [Google Scholar] [CrossRef]
- Bradford, M.M. A rapid and sensitive method for the quantitation of microgram quantities of protein utilizing the principle of protein-dye binding. Anal. Biochem. 1976, 72, 248–254. [Google Scholar] [CrossRef]
- Aebi, H. Catalase in vitro. Meth. Enzymol. 1984, 105, 121–126. [Google Scholar] [CrossRef]
- Draper, H.H.; Hadley, M. Malondialdehyde detemination as index of lipid peroxidation. Meth. Enzymol. 1990, 186, 421–431. [Google Scholar] [CrossRef]
- Staples, R.C.; Stahmann, M.A. Changes in proteins and several enzymes in susceptible bean leaves after infection by the bean rust fungus. Phytopathology 1964, 54, 760–764. [Google Scholar]
- Laemmli, U.K. Cleavage of structural proteins during the assembly of the head of bacteriophage T4. Nature 1970, 227, 680–685. [Google Scholar] [CrossRef]
- Singleton, V.L.; Rossi, J.A. Colorimetry of total phenolics with phosphomolybdic-phosphotungstic acid reagents. Am. J. Enol. Viticul. 1965, 16, 144–158. [Google Scholar]
- Woisky, R.G.; Salatino, A. Analysis of propolis: Some parameters and procedures for chemical quality control. J. Apicul. Res. 1998, 37, 99–105. [Google Scholar] [CrossRef]
- Ishida, M.; Nagata, M.; Ohara, T.; Kakizaki, T.; Hatakeyama, K.; Nishio, T. Small variation of glucosinolate composition in Japanese cultivars of radish (Raphanus sativus L.) requires simple quantitative analysis for breeding of glucosinolate component. Breed. Sci. 2012, 62, 63–70. [Google Scholar] [CrossRef] [Green Version]
- Brand-Williams, W.; Cuvelier, M.E.; Berset, C.L.W.T. Use of a free radical method to evaluate antioxidant activity. LWT-Food Sci. Tech. 1995, 28, 25–30. [Google Scholar] [CrossRef]
- Malar, J.; Chairman, K.; Singh, A.R.J.; Vanmathi, J.S.; Balasubramanian, A.; Vasanthi, K. Antioxidative activity of different parts of the plant Lepidium sativum Linn. Biotech. Rep. 2014, 3, 95–98. [Google Scholar] [CrossRef]
- Maness, N. Extraction and analysis of soluble carbohydrates. In Plant Stress Tolerance; Sunkar, R., Ed.; Methods in Molecular Biology; Humana Press: New York, NY, USA, 2010; Volume 639. [Google Scholar] [CrossRef]
- Doyle, J.J.; Doyle, J.L. A rapid DNA isolation procedure for small quantities of fresh leaf tissue. Phytochem. Bull. 1987, 19, 11–15. [Google Scholar]
- Škiljaica, A.; Jagić, M.; Vuk, T.; Leljak Levanić, D.; Bauer, N.; Markulin, L. Evaluation of reference genes for RT-qPCR gene expression analysis in Arabidopsis thaliana exposed to elevated temperatures. Plant Biol. 2022, 24, 367–379. [Google Scholar] [CrossRef]
- Livak, K.J.; Schmittgen, T.D. Analysis of relative gene expression data using real-time quantitative PCR and the 2−ΔΔCT method. Methods 2001, 25, 402–408. [Google Scholar] [CrossRef]
- Gu, Z.; Eils, R.; Schlesner, M. Complex heatmaps reveal patterns and correlations in multidimensional genomic data. Bioinformatics 2016, 32, 2847–2849. [Google Scholar] [CrossRef]
- Sammut, C.; Webb, G.I. Leave-One-Out Cross-Validation. In Encyclopedia of Machine Learning; Sammut, C., Webb, G.I., Eds.; Springer: Boston, MA, USA, 2011; pp. 600–601. [Google Scholar] [CrossRef]











{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Acc. | City | Country | Coordinates | |
|---|---|---|---|---|
| 202 | Kaštelir | Croatia | 45°18′21.9708″ N | 13°41′11.7018″ E |
| 206 | Labinci | Croatia | 45°17′38.3244″ N | 13°41′12.6636″ E |
| 379 | Fuškulin | Croatia | 45°10′57.3492″ N | 13°38′32.5608″ E |
| 380 | Island Iž 2. | Croatia | 44°0′56.74″ N | 15°8′21.19″ E |
| 381 | Preko, island Ugljan | Croatia | 44°4′45.5916″ N | 15°11′14.5212″ E |
| 383 | Vrgorac | Croatia | 43°12′4.3806″ N | 17°23′16.0326″ E |
| 384 | Konavle, Pavlje Brdo | Croatia | 42°30′55.0296″ N | 18°20′50.5494″ E |
| 385 | Vitina, Mostar | Bosnia and Herzegovina | 43°14′33.5178″ N | 17°28′29.3694″ E |
| 386 | Vrgorac | Croatia | 43°11′28.1904″ N | 17°21′53.643″ E |
| 387 | Ponikve, Pelješac | Croatia | 42°50′52.983″ N | 17°37′36.786″ E |
| 388 | Topići, Baška voda | Croatia | 43°22′0.0912″ N | 16°57′41.2698″ E |
| 390 | Island Lošinj | Croatia | 44°34′53.097″ N | 14°24′21.9018″ E |
| 391 | Ugljan, islland Ugljan | Croatia | 44°8′0.1392″ N | 15°6′8.7804″ E |
| 392 | Dubrovnik | Croatia | 42°38′40.8264″ N | 18°7′21.2592″ E |
| 393 | Mostar | Bosnia and Herzegovina | 43°21′35.7186″ N | 17°49′7.269″ E |
| 394 | Oključina, island Vis | Croatia | 43°4′17.7348″ N | 16°6′36.1296″ E |
| 395 | Mostar | Bosnia and Herzegovina | 43°20′25.2564″ N | 17°43′28.1274″ E |
| 396 | Island Lošinj | Croatia | 44°41′9.942″ N | 14°22′12.864″ E |
| 397 | Vrgorac | Croatia | 43°12′25.7106″ N | 17°21′36.8496” E |
| 398 | Vrgorac, Prapatnice | Croatia | 43°13′43.85″ N | 17°20′59.03″ E |
| 399 | Čarsko polje, island Korčula | Croatia | 42°55′59.8362″ N | 16°56′6.0828″ E |
| 401 | Zavalatica, island Korčula | Croatia | 42°55′7.0968″ N | 16°56′9.4272″ E |
| 402 | Katuni | Croatia | 43°27′45.39″ N | 16°53′10.39″ E |
| 403 | Drinovci | Croatia | 43°21′33.0696″ N | 17°19′23.9874″ E |
| 404 | Blato na Cetini, Omiš | Croatia | 43°28′34.87″ N | 16°49′7.39″ E |
| 405 | Opuzen | Croatia | 43°0′19.28″ N | 17°33′34.69″ E |
| 406 | Babino polje, isl. Mljet | Croatia | 42°44′22.63″ N | 17°30′2.93″ E |
| 408 | Poreč-Pištan | Croatia | 45°13′29.33″ N | 13°37′39.76” E |
| 411 | Island Iž 1. | Croatia | 44°2′37.248″ N | 15°6′43.671″ E |
| 413 | Tomislav grad | Bosnia and Herzegovina | 43°43′7.39″ N | 17°13′25.07″ E |
| 414 | Srijane | Croatia | 43°31′20.94″ N | 16°41′14.45″ E |
| 416 | Vinjani donji | Croatia | 43°26′28.23″ N | 17°14′25.66″ E |
| 423 | Kreševo | Croatia | 43°29′14.65″ N | 16°52′35.42″ E |
| Parameter | Source | SS | Df | MS | F | p | Significance Level |
|---|---|---|---|---|---|---|---|
| Total phenols | A | 61.02 | 3 | 20.34 | 14.07 | 0.00000 | *** |
| T | 12.80 | 3 | 4.27 | 2.95 | 0.03868 | * | |
| A×T | 18.82 | 9 | 2.09 | 1.45 | 0.18598 | ns | |
| Total flavonoids | A | 9.42 | 3 | 3.14 | 42.69 | 0.00000 | *** |
| T | 34.45 | 3 | 11.48 | 156.08 | 0.00000 | *** | |
| A×T | 5.52 | 9 | 0.61 | 8.34 | 0.00000 | *** | |
| Antioxidative activity | A | 76.93 | 3 | 25.64 | 13.92 | 0.00000 | *** |
| T | 73.82 | 3 | 24.61 | 13.35 | 0.00000 | *** | |
| A×T | 51.94 | 9 | 5.77 | 3.13 | 0.00327 | ** | |
| Total glucosinolates | A | 183.15 | 3 | 61.05 | 30.19 | 0.00000 | *** |
| T | 1137.48 | 3 | 379.16 | 187.5 | 0.00000 | *** | |
| A×T | 111.58 | 9 | 12.40 | 6.13 | 0.00000 | *** | |
| MDA | A | 4.23 | 3 | 1.41 | 4.12 | 0.01404 | * |
| T | 32.87 | 3 | 10.96 | 31.99 | 0.00000 | *** | |
| A×T | 9.58 | 9 | 1.06 | 3.11 | 0.00847 | ** | |
| CAT activity | A | 2561.07 | 3 | 853.69 | 71.86 | 0.00000 | *** |
| T | 742.76 | 3 | 247.59 | 20.84 | 0.00000 | *** | |
| A×T | 545.15 | 9 | 60.57 | 5.10 | 0.00027 | *** | |
| H2O2 content | A | 8.28 | 3 | 2.76 | 3.14 | 0.03864 | * |
| T | 17.37 | 3 | 5.79 | 6.59 | 0.00136 | ** | |
| A×T | 14.83 | 9 | 1.65 | 1.88 | 0.09231 | ns | |
| GSH | A | 0.85 | 3 | 0.28 | 10.45 | 0.00001 | *** |
| T | 1.87 | 3 | 0.62 | 22.99 | 0.00000 | *** | |
| A×T | 0.99 | 9 | 0.11 | 4.06 | 0.00034 | *** | |
| Fv/Fm | A | 0.00 | 3 | 0.00 | 3.40 | 0.02667 | * |
| T | 0.00 | 3 | 0.00 | 6.10 | 0.00156 | ** | |
| A×T | 0.00 | 9 | 0.00 | 1.00 | 0.42690 | ns | |
| PIabs | A | 4.56 | 3 | 1.52 | 7.68 | 0.00034 | *** |
| T | 3.51 | 3 | 1.17 | 5.92 | 0.00184 | ** | |
| A×T | 1.40 | 9 | 0.16 | 0.79 | 0.62964 | ns | |
| Total sugars | A | 35.56 | 3 | 11.85 | 5.45 | 0.00204 | ** |
| T | 25.22 | 3 | 8.41 | 3.87 | 0.01296 | * | |
| A×T | 44.17 | 9 | 4.91 | 2.26 | 0.02835 | * | |
| A | 643.29 | 3 | 214.43 | 38.88 | 0.00000 | *** | |
| Proline content | T | 2011.56 | 3 | 670.52 | 121.59 | 0.00000 | *** |
| A×T | 284.31 | 9 | 31.59 | 5.73 | 0.00003 | *** | |
| A | 41.05 | 3 | 13.68 | 7.04 | 0.00015 | *** | |
| Root growth | T | 598.51 | 3 | 199.50 | 102.68 | 0.00000 | *** |
| A×T | 104.58 | 9 | 11.62 | 5.98 | 0.00000 | *** | |
| NAC041 | A | 2.63 | 3 | 0.88 | 12.63 | 0.00000 | *** |
| T | 1.46 | 3 | 0.49 | 7.00 | 0.00031 | *** | |
| A×T | 0.68 | 9 | 0.08 | 1.09 | 0.37708 | ns | |
| NAC084 | A | 19.42 | 3 | 6.47 | 15.64 | 0.00000 | *** |
| T | 0.79 | 3 | 0.26 | 0.64 | 0.59454 | ns | |
| A×T | 3.05 | 9 | 0.34 | 0.82 | 0.60024 | ns | |
| HSFA2 | A | 4363.49 | 3 | 1454.5 | 4.75 | 0.00424 | ** |
| T | 38,958.62 | 3 | 12,986.21 | 42.41 | 0.00000 | *** | |
| A×T | 14,531.79 | 9 | 1614.64 | 5.27 | 0.00001 | *** | |
| HSFA7 | A | 3390.37 | 3 | 1130.12 | 26.89 | 0.00000 | *** |
| T | 7988.41 | 3 | 2662.80 | 63.36 | 0.00000 | *** | |
| A×T | 8587.73 | 9 | 954.19 | 22.70 | 0.00000 | *** | |
| DREB2A | A | 50.25 | 3 | 16.75 | 0.69 | 0.55866 | ns |
| T | 1884.68 | 3 | 628.23 | 26.01 | 0.00000 | *** | |
| A×T | 928.47 | 9 | 103.16 | 4.27 | 0.00015 | *** |
| Gene | Primer Name | Sequence 5′->3′ |
|---|---|---|
| DREB2A | qB-DREB2A-Fw | TTTGATGTTTCTGAGCTTCTTGG |
| Bra009112 | qB-DREB2A-Rev | CATTGTCTCCCAGGCATTGG |
| HSFA2 | qB-HSFA2-Fw | ATGAATGTGATGATGGAAGATGGT |
| Bra000557 | qB-HSFA2-Rev | CTGCCCCAATCCAACGGTG |
| HSFA7A | qB-HSFA7A-Fw | TCTGAGACAGCAGCAACAAAC |
| Bra012828 | qB-HSFA7A-Rev | CTGGAGTAGCTGATACAGAAAC |
| NAC041 | qB-NAC041-Fw | CGAAGACGACAACAAGAGTGC |
| Bra021856 | qB-NAC041-Rev | GAGTCACATTCAAATCGCAGC |
| NAC084 | qB-NAC084-Fw | AGGAAGAAGACAGAGGAAACC |
| Bra006229 | qB-NAC084-Rev | GCTGAGGTAGGAGGAGATG |
| OGIO | qB-OGIO-Fw | CAGTATCGTAGCTGAGGTAGC |
| Bra028284 | qB-OGIO-Rev | AGAACGGAACACATACTTGACTC |
| PUX | qB-PUX-Fw | CAAACCCAAAGAGGTTGTTGC |
| Bra026205 | qB-PUX-Rev | TCATGTCGTTGTCTTCCAAGG |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Bauer, N.; Tkalec, M.; Major, N.; Talanga Vasari, A.; Tokić, M.; Vitko, S.; Ban, D.; Ban, S.G.; Salopek-Sondi, B. Mechanisms of Kale (Brassica oleracea var. acephala) Tolerance to Individual and Combined Stresses of Drought and Elevated Temperature. Int. J. Mol. Sci. 2022, 23, 11494. https://doi.org/10.3390/ijms231911494
Bauer N, Tkalec M, Major N, Talanga Vasari A, Tokić M, Vitko S, Ban D, Ban SG, Salopek-Sondi B. Mechanisms of Kale (Brassica oleracea var. acephala) Tolerance to Individual and Combined Stresses of Drought and Elevated Temperature. International Journal of Molecular Sciences. 2022; 23(19):11494. https://doi.org/10.3390/ijms231911494
Chicago/Turabian StyleBauer, Nataša, Mirta Tkalec, Nikola Major, Ana Talanga Vasari, Mirta Tokić, Sandra Vitko, Dean Ban, Smiljana Goreta Ban, and Branka Salopek-Sondi. 2022. "Mechanisms of Kale (Brassica oleracea var. acephala) Tolerance to Individual and Combined Stresses of Drought and Elevated Temperature" International Journal of Molecular Sciences 23, no. 19: 11494. https://doi.org/10.3390/ijms231911494