
HLA-G and Recurrent Pregnancy Loss

Abstract
:1. Introduction
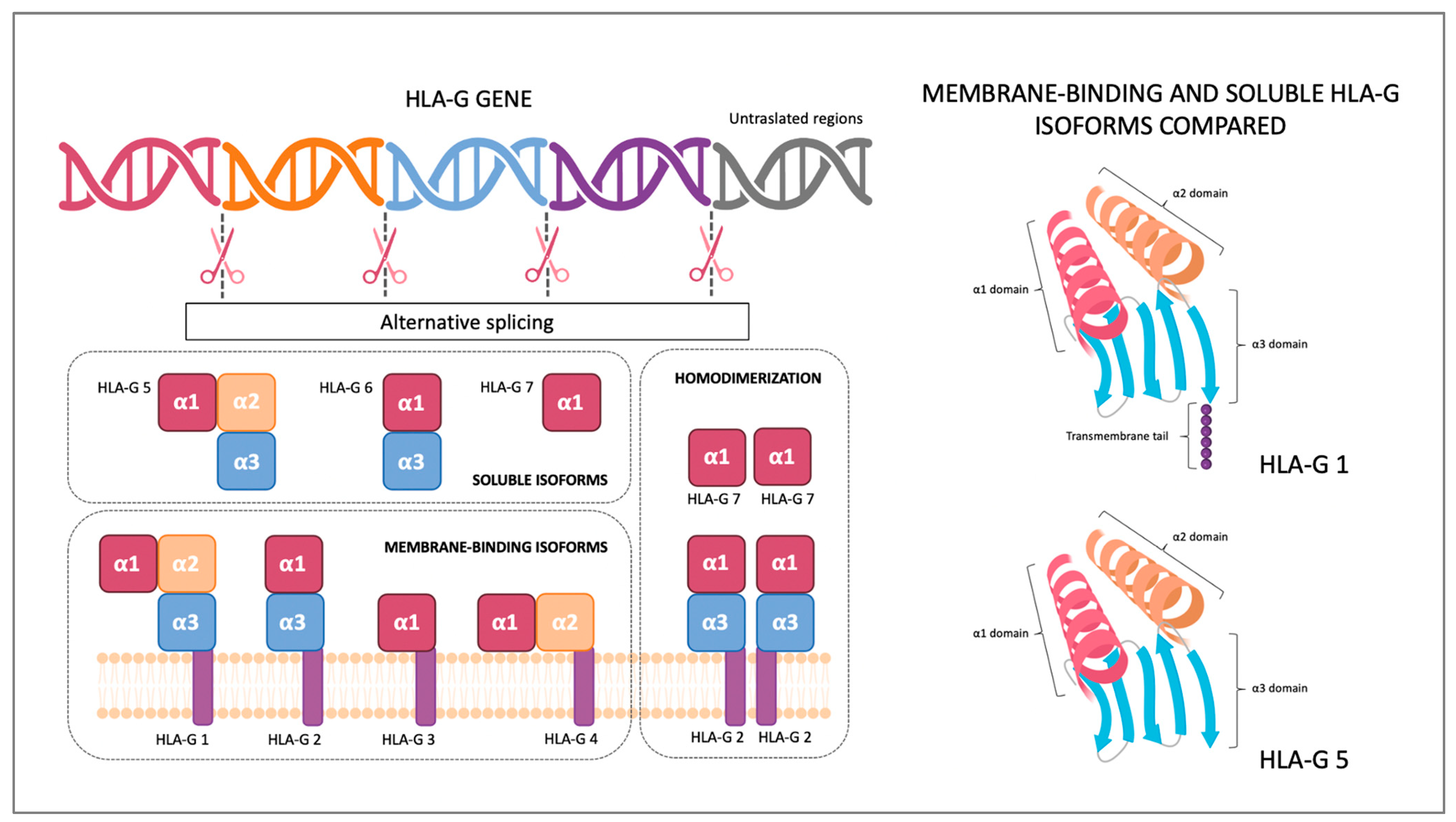
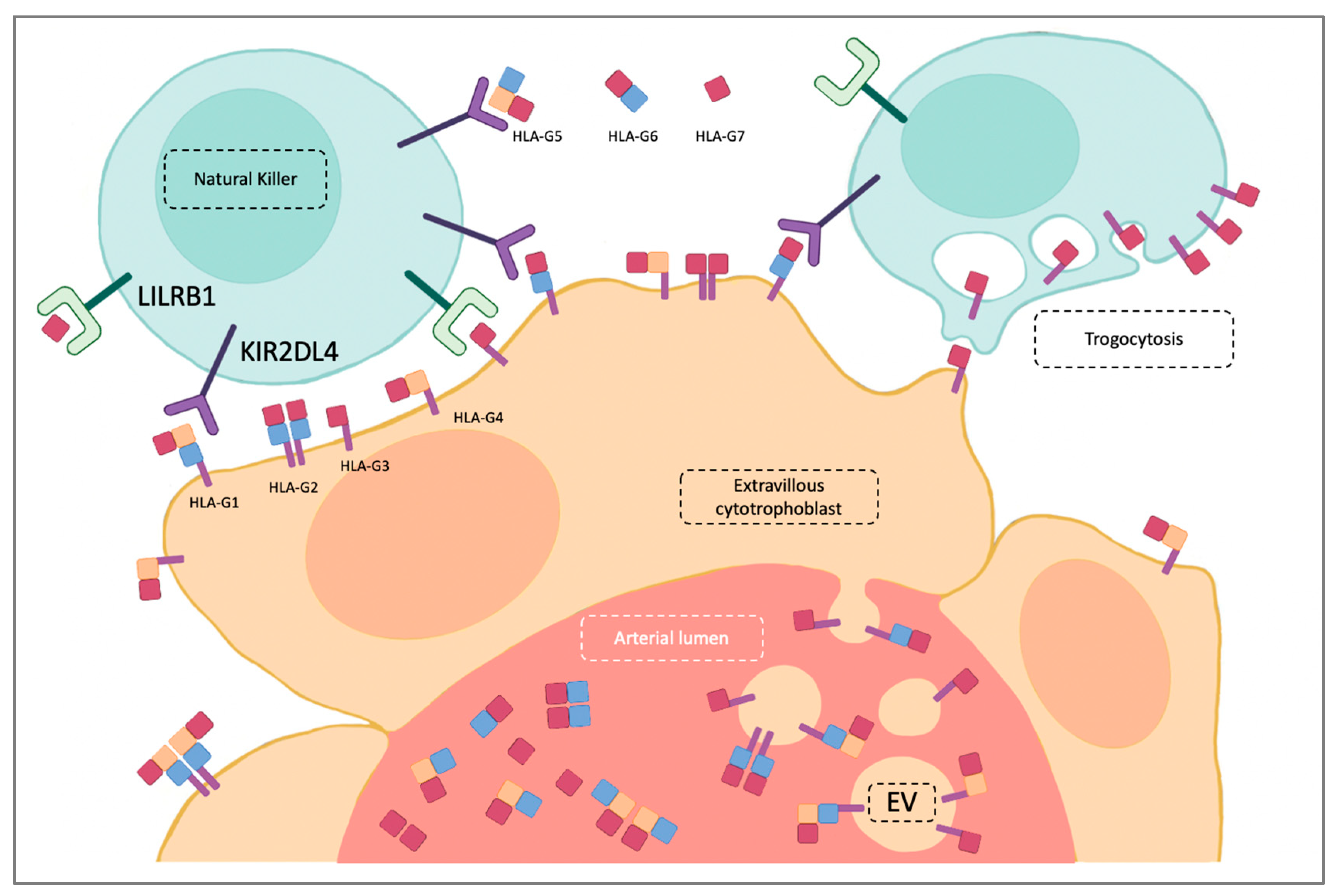
2. HLA-G Structure and Receptors
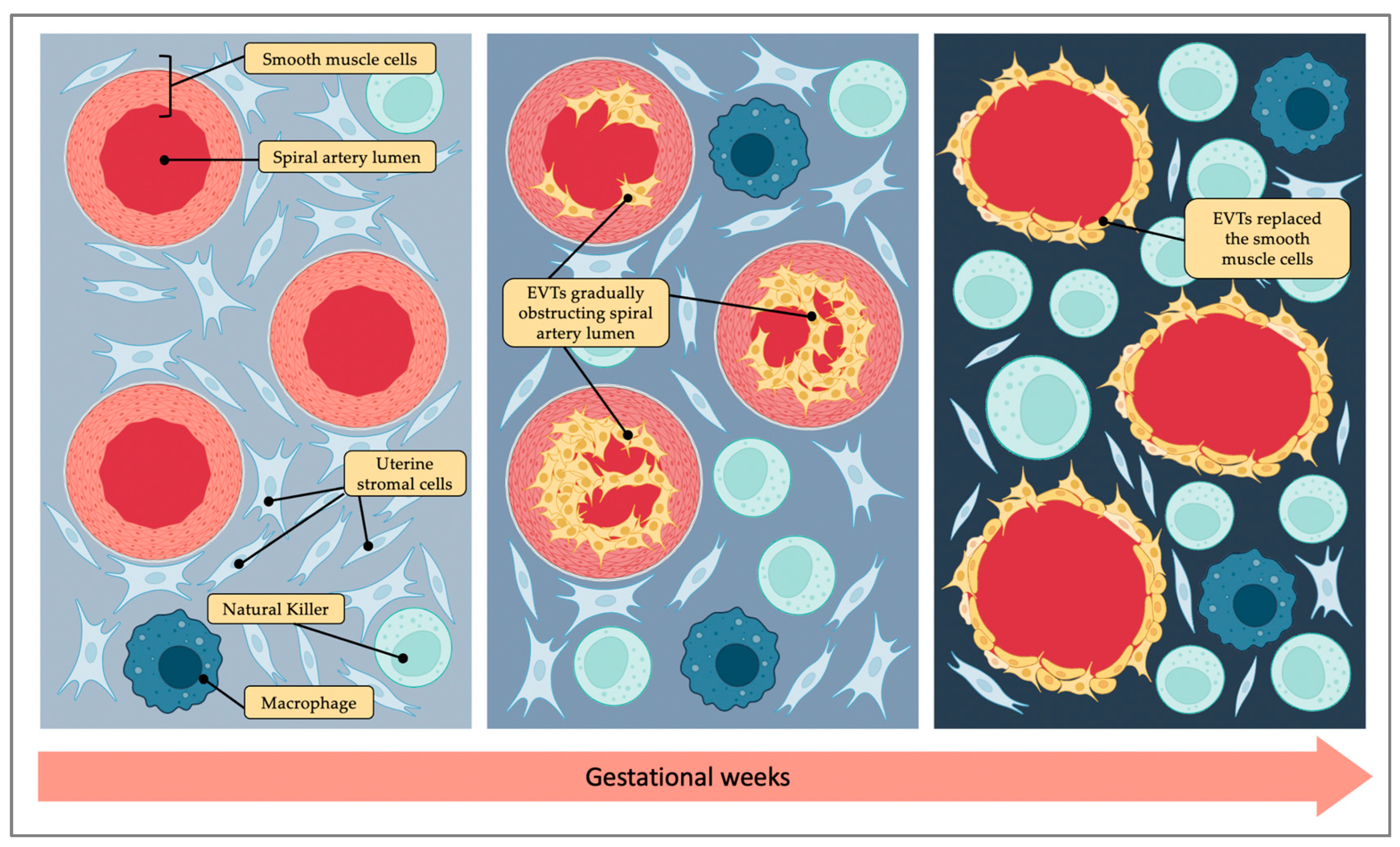
3. Placentation
4. HLA-G Role in Physiological Pregnancy
5. HLA-G in Recurrent Pregnancy Loss
6. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Kovats, S.; Main, E.K.; Librach, C.; Stubblebine, M.; Fisher, S.J.; DeMars, R. A class I antigen, HLA-G, expressed in human trophoblasts. Science 1990, 248, 220–223. [Google Scholar] [CrossRef] [PubMed]
- Martín-Villa, J.M.; Vaquero-Yuste, C.; Molina-Alejandre, M.; Juarez, I.; Suárez-Trujillo, F.; López-Nares, A.; Palacio-Gruber, J.; Barrera-Gutiérrez, L.; Fernández-Cruz, E.; Rodríguez-Sainz, C.; et al. HLA-G: Too Much or Too Little? Role in Cancer and Autoimmune Disease. Front. Immunol. 2022, 13, 796054. [Google Scholar] [CrossRef] [PubMed]
- Rotondo, J.C.; Mazziotta, C.; Lanzillotti, C.; Tognon, M.; Martini, F. Epigenetic Dysregulations in Merkel Cell Polyomavirus-Driven Merkel Cell Carcinoma. Int. J. Mol. Sci. 2021, 22, 11464. [Google Scholar] [CrossRef] [PubMed]
- ESHRE Guideline Group on RPL; Bender Atik, R.; Christiansen, O.B.; Elson, J.; Kolte, A.M.; Lewis, S.; Middeldorp, S.; Nelen, W.; Peramo, B.; Quenby, S.; et al. ESHRE guideline: Recurrent pregnancy loss. Hum. Reprod. Open 2018, 2018, hoy004. [Google Scholar]
- Van Dijk, M.M.; Kolte, A.M.; Limpens, J.; Kirk, E.; Quenby, S.; van Wely, M.; Goddijn, M. Recurrent pregnancy loss: Diagnostic workup after two or three pregnancy losses? A systematic review of the literature and meta-analysis. Hum. Reprod. Updat. 2020, 26, 356–367. [Google Scholar] [CrossRef] [Green Version]
- Li, Y.H.; Marren, A. Recurrent pregnancy loss: A summary of international evidence-based guidelines and practice. Aust. J. Gen. Pr. 2018, 47, 432–436. [Google Scholar] [CrossRef] [Green Version]
- Jauniaux, E.; Farquharson, R.G.; Christiansen, O.B.; Exalto, N. Evidence-based guidelines for the investigation and medical treat-ment of recurrent miscarriage. Hum. Reprod. 2006, 21, 2216–2222. [Google Scholar] [CrossRef] [Green Version]
- Dai, Y.; Liu, J.; Yuan, E.; Li, Y.; Shi, Y.; Zhang, L. Relationship Among Traditional Semen Parameters, Sperm DNA Fragmentation, and Unexplained Recurrent Miscarriage: A Systematic Review and Meta-Analysis. Front. Endocrinol. 2022, 12, 1844. [Google Scholar] [CrossRef]
- D’Ippolito, S.; Ticconi, C.; Tersigni, C.; Garofalo, S.; Martino, C.; Lanzone, A.; Scambia, G.; Di Simone, N. The pathogenic role of autoantibodies in recurrent pregnancy loss. Am. J. Reprod. Immunol. 2020, 83, e13200. [Google Scholar] [CrossRef]
- Christiansen, O.B. The epidemiology of recurrent pregnancy loss. In Recurrent Pregnancy Loss: Causes, Controversies and Treatment, 3rd ed.; Carp, H.J.A., Ed.; CRC Press: Boca Raton, FL, USA, 2020; pp. 1–12. [Google Scholar]
- Snell, G.D.; Higgins, G.F. Alleles at the histocompatibility-2 locus in the mouse as determined by tumor transplantation. Genetics 1951, 36, 306–310. [Google Scholar] [CrossRef]
- Neefjes, J.; Jongsma, M.L.M.; Paul, P.; Bakke, O. Towards a systems understanding of MHC class I and MHC class II antigen presentation. Nat. Rev. Immunol. 2011, 11, 823–836. [Google Scholar] [PubMed]
- Arnaiz-Villena, A.; Juarez, I.; Suarez-Trujillo, F.; López-Nares, A.; Vaquero, C.; Palacio-Gruber, J.; Martin-Villa, J.M. HLA-G: Function, polymorphisms and pathology. Int. J. Immunogenet. 2021, 48, 172–192. [Google Scholar] [CrossRef] [PubMed]
- Nardi Fda, S.; König, L.; Wagner, B.; Giebel, B.; Santos Manvailer, L.F.; Rebmann, V. Soluble monomers, dimers and HLA-G-expressing extracellular vesicles: The three dimensions of structural complexity to use HLA-G as a clinical biomarker. HLA 2016, 88, 77–86. [Google Scholar] [PubMed]
- Carter, A. Unique Aspects of Human Placentation. Int. J. Mol. Sci. 2021, 22, 8099. [Google Scholar] [CrossRef] [PubMed]
- Maltepe, E.; Fisher, S.J. Placenta: The forgotten organ. Annu. Rev. Cell Dev. Biol. 2015, 31, 523–552. [Google Scholar] [CrossRef] [PubMed]
- Burton, G.; Woods, A.; Jauniaux, E.; Kingdom, J. Rheological and physiological consequences of conversion of the maternal spiral arteries for uteroplacental blood flow during human pregnancy. Placenta 2009, 30, 473–482. [Google Scholar]
- Fajersztajn, L.; Veras, M.M. Hypoxia: From Placental Development to Fetal Programming. Birth Defects Res. 2017, 109, 1377–1385. [Google Scholar] [CrossRef]
- Xu, X.; Zhou, Y.; Wei, H. Roles of HLA-G in the maternal-fetal immune microenvironment. Front. Immunol. 2020, 11, 592010. [Google Scholar] [CrossRef]
- Rouas-Freiss, N.; Moreau, P.; LeMaoult, J.; Papp, B.; Roux, D.T.-L.; Carosella, E.D. Role of the HLA-G immune checkpoint molecule in pregnancy. Hum. Immunol. 2021, 82, 353–361. [Google Scholar] [CrossRef]
- Aisagbonhi, O.; Morris, G.P. Human Leukocyte Antigens in Pregnancy and Preeclampsia. Front. Genet. 2022, 13, 884275. [Google Scholar] [CrossRef]
- Turco, M.Y. Trophoblast organoids as a model for maternal-fetal interactions during human placentation. Nature 2018, 564, 263–267. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Singer, G.; Kurman, R.J.; McMaster, M.T.; Shih, I.-M. HLA-G Immunoreactivity is specific for intermediate trophoblast in gestational trophoblastic disease and can serve as a useful marker in differential diagnosis. Am. J. Surg. Pathol. 2002, 26, 914–920. [Google Scholar] [CrossRef] [PubMed]
- Nagamatsu, T.; Fujii, T.; Yamashita, T.; Miki, A.; Kanai, T.; Kusumi, M.; Osuga, Y.; Kozuma, S.; Taketani, Y. Hypoxia does not reduce HLA-G expression on extravillous cytotrophoblasts. J. Reprod. Immunol. 2004, 63, 85–95. [Google Scholar] [CrossRef] [PubMed]
- Mouillot, G.; Marcou, C.; Zidi, I.; Guillard, C.; Sangrouber, D.; Carosella, E.D.; Moreau, P. Hypoxia modulates HLA-G gene expression in tumor cells. Hum. Immunol. 2007, 68, 277–285. [Google Scholar] [CrossRef]
- Mori, A.; Nishi, H.; Sasaki, T.; Nagamitsu, Y.; Kawaguchi, R.; Okamoto, A.; Kuroda, M.; Isaka, K. HLA-G expression is regulated by miR-365 in trophoblasts under hypoxic conditions. Placenta 2016, 45, 37–41. [Google Scholar] [CrossRef]
- Soares, M.J.; Iqbal, K.; Kozai, K. Hypoxia and Placental Development. Birth Defects Res. 2017, 109, 1309–1329. [Google Scholar] [CrossRef]
- Smith, H.J. Antigenicity of carcinogen-induced and spontaneous tumours in inbred mice Br. J. Cancer 1966, 20, 831–837. [Google Scholar] [CrossRef] [Green Version]
- Zhang, X.; Wei, H. Role of Decidual Natural Killer Cells in Human Pregnancy and Related Pregnancy Complications. Front. Immunol. 2021, 12, 3421. [Google Scholar] [CrossRef]
- Xie, M.; Li, Y.; Meng, Y.-Z.; Xu, P.; Yang, Y.-G.; Dong, S.; He, J.; Hu, Z. Uterine Natural Killer Cells: A Rising Star in Human Pregnancy Regulation. Front. Immunol. 2022, 13, 2733. [Google Scholar] [CrossRef]
- Drury, J.A.; Parkin, K.L.; Coyne, L.; Giuliani, E.; Fazleabas, A.T.; Hapangama, D.K. The Dynamic Changes in the Number of Uterine Natural Killer Cells Are Specific to the Eutopic But Not to the Ectopic Endometrium in Women and in a Baboon Model of Endometriosis. Reprod. Biol. Endocrinol. 2018, 16, 67. [Google Scholar] [CrossRef]
- Santoni, A.; Carlino, C.; Stabile, H.; Gismondi, A. REVIEW ARTICLE: Mechanisms underlying recruitment and accumulation of decidual nk cells in uterus during pregnancy. Am. J. Reprod. Immunol. 2008, 59, 417–424. [Google Scholar] [CrossRef] [PubMed]
- Sentman, C.L.; Meadows, S.K.; Wira, C.R.; Eriksson, M. Recruitment of uterine NK Cells: Induction of CXC chemokine ligands 10 and 11 in human endometrium by estradiol and progesterone. J. Immunol. 2004, 173, 6760–6766. [Google Scholar] [CrossRef] [PubMed]
- Lash, G.E.; Otun, H.A.; Innes, B.A.; Percival, K.; Searle, R.F.; Robson, S.C.; Bulmer, J.N. Regulation of extravillous trophoblast invasion by uterine natural killer cells is dependent on gestational age. Hum. Reprod. 2010, 25, 1137–1145. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhuang, B.; Shang, J.; Yao, Y. HLA-G: An Important Mediator of Maternal-Fetal Immune-Tolerance. Front. Immunol. 2021, 12, 744324. [Google Scholar] [CrossRef]
- Yang, F.; Zheng, Q.; Jin, L. Dynamic Function and Composition Changes of Immune Cells During Normal and Pathological Pregnancy at the Maternal-Fetal Interface. Front. Immunol. 2019, 10, 2317. [Google Scholar] [CrossRef] [Green Version]
- Ashkar, A.A.; Croy, B.A. Interferon-g contributes to the normalcy of murine pregnancy. Biol. Reprod. 1999, 61, 493–502. [Google Scholar] [CrossRef] [Green Version]
- Blanco, O.; Tirado, I.; Muñoz-Fernández, R.; Abadía-Molina, A.C.; García-Pacheco, J.M.; Peña, J.; Olivares, E.G. Human decidual stromal cells express HLA-G: Effects of cytokines and decid-ualization. Hum. Reprod. 2008, 23, 144–152. [Google Scholar] [CrossRef] [Green Version]
- Gu, Y.; Jow, G.M.; Moulton, B.C.; Lee, C.; Sensibar, J.A.; Park-Sarge, O.K.; Chen, T.J.; Gibori, G. Apoptosis in decidual tissue regression and reorganization. Endocrinology 1994, 135, 1272–1279. [Google Scholar] [CrossRef]
- Lim, K.J.; Odukoya, O.A.; Ajjan, R.A.; Li, T.-C.; Weetman, A.P.; Cooke, I.D. The role of T-helper cytokines in human reproduction. Fertil. Steril. 2000, 73, 136–142. [Google Scholar] [CrossRef]
- Fonseca, B.M.; Cunha, S.C.; Gonçalves, D.; Mendes, A.; Braga, J.; Correia-Da-Silva, G.; Teixeira, N.A. Decidual NK Cell-Derived Conditioned Medium from Miscarriages Affects Endometrial Stromal Cell Decidualisation: Endocannabinoid Anandamide and Tumour Necrosis Factor-α Crosstalk. Hum. Reprod. 2020, 35, 265–274. [Google Scholar] [CrossRef]
- Caumartin, J.; Favier, B.; Daouya, M.; Guillard, C.; Moreau, P.; Carosella, E.D.; LeMaoult, J. Trogocytosis-based generation of suppressive NK cells. EMBO J. 2007, 26, 1423–1433. [Google Scholar] [PubMed] [Green Version]
- Ferreira, L.M.; Meissner, T.B.; Tilburgs, T.; Strominger, J.L. HLA-G: At the Interface of Maternal–Fetal Tolerance. Trends Immunol. 2017, 38, 272–286. [Google Scholar] [PubMed]
- Tilburgs, T.; Evans, J.H.; Crespo, Â.C.; Strominger, J.L. The HLA-G cycle provides for both NK tolerance and immunity at the maternal-fetal interface. Proc. Natl. Acad. Sci. USA 2015, 112, 13312–13317. [Google Scholar]
- Kotze, D.; Kruger, T.F.; Lombard, C.; Padayachee, T.; Keskintepe, L.; Sher, G. The effect of the biochemical marker soluble human leukocyte antigen G on pregnancy outcome in assisted reproductive technology—A multicenter study. Fertil. Steril. 2013, 100, 1303–1309. [Google Scholar]
- Morandi, F.; Ferretti, E.; Bocca, P.; Prigione, I.; Raffaghello, L.; Pistoia, V. A novel mechanism of soluble HLA-G mediated immune modulation: Downregulation of T cell chemokine receptor expression and impairment of chemotaxis. PLoS ONE 2010, 5, e11763. [Google Scholar]
- Li, P.; Wang, N.; Zhang, Y.; Wang, C.; Du, L. HLA-G/sHLA-G and HLA-G-Bearing Extracellular Vesicles in Cancers: Potential Role as Biomarkers. Front. Immunol. 2021, 12, 791535. [Google Scholar]
- Erebmann, V.; König, L.; Nardi, F.D.S.; Wagner, B.; Manvailer, L.F.; Horn, P.A. The Potential of HLA-G-Bearing Extracellular Vesicles as a Future Element in HLA-G Immune Biology. Front. Immunol. 2016, 7, 173. [Google Scholar]
- Cho, K.; Kook, H.; Kang, S.; Lee, J. Study of immune-tolerized cell lines and extracellular vesicles inductive environment promoting continuous expression and secretion of HLA-G from semiallograft immune tolerance during pregnancy. J. Extracell. Vesicles 2020, 9, 1795364. [Google Scholar]
- Kshirsagar, S.; Alam, S.; Jasti, S.; Hodes, H.; Nauser, T.; Gilliam, M.; Billstrand, C.; Hunt, J.; Petroff, M. Immunomodulatory molecules are released from the first trimester and term placenta via exosomes. Placenta 2012, 33, 982–990. [Google Scholar]
- Wu, H.-M.; Chen, L.-H.; Hsu, L.-T.; Lai, C.-H. Immune Tolerance of Embryo Implantation and Pregnancy: The Role of Human Decidual Stromal Cell- and Embryonic-Derived Extracellular Vesicles. Int. J. Mol. Sci. 2022, 23, 13382. [Google Scholar]
- Murrieta-Coxca, J.M.; Fuentes-Zacarias, P.; Ospina-Prieto, S.; Markert, U.R.; Morales-Prieto, D.M. Synergies of Extracellular Vesicles and Microchimerism in Promoting Immunotolerance During Pregnancy. Front. Immunol. 2022, 13, 837281. [Google Scholar] [PubMed]
- Hedlund, M.; Stenqvist, A.-C.; Nagaeva, O.; Kjellberg, L.; Wulff, M.; Baranov, V.; Mincheva-Nilsson, L. Human placenta expresses and secretes NKG2D ligands via exosomes that down-modulate the cognate receptor expression: Evidence for immunosuppressive function. J. Immunol. 2009, 183, 340–351. [Google Scholar] [PubMed] [Green Version]
- Grange, C.; Camussi, G. Immunosuppressive role of extracellular vesicles: HLA-G, an important player. Ann. Transl. Med. 2017, 5, 223. [Google Scholar] [PubMed]
- Dimitriadis, E.; Menkhorst, E.; Saito, S.; Kutteh, W.H.; Brosens, J.J. Recurrent pregnancy loss. Nat. Rev. Dis. Prim. 2020, 6, 98. [Google Scholar] [PubMed]
- Hu, L.; He, D.; Zeng, H. Association of parental HLA-G polymorphisms with soluble HLA-G expressions and their roles on recurrent implantation failure: A systematic review and meta-analysis. Front. Immunol. 2022, 13, 988370. [Google Scholar]
- Alegre, E.; Díaz-Lagares, A.; LeMaoult, J.; López-Moratalla, N.; Carosella, E.D.; González, A. Maternal antigen presenting cells are a source of plasmatic HLA-G during pregnancy: Longitudinal study during pregnancy. Hum. Immunol. 2007, 68, 661–667. [Google Scholar]
- Zidi, I.; Rizzo, R.; Bouaziz, A.; Laaribi, A.B.; Zidi, N.; Di Luca, D.; Tlili, H.; Bortolotti, D. sHLA-G1 and HLA-G5 levels are decreased in Tunisian women with multiple abortion. Hum. Immunol. 2016, 77, 342–345. [Google Scholar]
- Bae, S.-C.; Lee, Y.H. Association of HLA-G polymorphisms with systemic lupus erythematosus and correlation between soluble HLA-G levels and the disease: A meta-analysis. Z. Rheumatol. 2021, 80, 96–102. [Google Scholar]
- de Carvalho, J.F.; de Oliveira, R.M.; Rodrigues, C.E.; Glezer, A.; Bonfá, E.; Pereira, R.M. Heparin increases HLA-G levels in primary antiphospholipid syndrome. Clin. Dev. Immunol. 2012, 2012, 232390. [Google Scholar]
- Klitkou, L.; Dahl, M.; Hviid, T.V.F.; Djurisic, S.; Piosik, Z.M.; Skovbo, P.; Møller, A.M.; Steffensen, R.; Christiansen, O.B. Human leukocyte antigen (HLA)-G during pregnancy part I: Correlations between maternal soluble HLA-G at midterm, at term, and umbilical cord blood soluble HLA-G at term. Hum. Immunol. 2015, 76, 254–259. [Google Scholar]
- Krop, J.; Van Der Keur, C.; Kapsenberg, J.; Hollander, F.D.; Van Der Hoorn, M.; Heidt, S.; Claas, F.; Eikmans, M. Soluble HLA-G blood levels are not increased during ongoing pregnancy in women with a history of recurrent pregnancy loss. J. Reprod. Immunol. 2022, 153, 103665. [Google Scholar] [CrossRef]
- Madduru, D.; Dirsipam, K.; Goli, M.; Ramana Devi, V.; Jahan, P. Association of reduced maternal sHLA-G5 isoform levels and elevated TNF-α/IL-4 cytokine ratio with Recurrent Pregnancy Loss: A study on South Indian women. Scand. J. Immunol. 2021, 94, e13095. [Google Scholar] [CrossRef]
- Craenmehr, M.H.C.; Nederlof, I.; Cao, M.; Drabbels, J.J.M.; Spruyt-Gerritse, M.J.; Anholts, J.D.H.; Kapsenberg, H.M.; Stegehuis, J.A.; van der Keur, C.; Fasse, E.; et al. Increased HLA-G Expression in Term Placenta of Women with a History of Recurrent Miscarriage Despite Their Genetic Predisposition to Decreased HLA-G Levels. Int. J. Mol. Sci. 2019, 20, 625. [Google Scholar]
- Persson, G.; Picard, C.; Marin, G.; Isgaard, C.; Stæhr, C.S.; Molinari, N.; Chiaroni, J.; Lebech, M.; Hviid, T.V.F.; Di Cristofaro, J. Maternal HLA Ib Polymorphisms in Pregnancy Allo-Immunization. Front. Immunol. 2021, 12, 657217. [Google Scholar] [PubMed]
- Mayr, C. What Are 3’ UTRs Doing? Cold Spring Harb. Perspect. Biol. 2019, 11, a034728. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Castelli, E.C.; de Almeida, B.S.; Muniz, Y.C.N.; Silva, N.S.B.; Passos, M.R.S.; Souza, A.S.; Page, A.E.; Dyble, M.; Smith, D.; Aguileta, G.; et al. HLA-G genetic diversity and evolutive aspects in worldwide populations. Sci. Rep. 2021, 11, 23070. [Google Scholar] [PubMed]
- Hviid, T.V.F.; Rizzo, R.; Christiansen, O.B.; Melchiorri, L.; Lindhard, A.; Baricordi, O.R. HLA-G and IL-10 in serum in relation to HLA-G genotype and polymorphisms. Immunogenetics 2004, 56, 135–141. [Google Scholar] [CrossRef] [PubMed]
- Monti, M.; Lupoli, R.; Sosa Fernandez, L.M.; Cirillo, F.; Di Minno, M.N.D. Association of human leukocyte antigen-G 14 bp poly-morphism with recurrent pregnancy loss in European countries: A meta-analysis of literature studies. Fertil. Steril. 2019, 112, 577–585. [Google Scholar] [CrossRef] [PubMed]
- Kalotra, V.; Lall, M.; Verma, I.C.; Kaur, A. TheHLA-G14 bp insertion/deletion polymorphism and its association with soluble HLA-G levels in women with recurrent miscarriages. HLA 2018, 91, 167–174. [Google Scholar] [CrossRef] [PubMed]
- Dietrich, B.; Kunihs, V.; Haider, S.; Pollheimer, J.; Knöfler, M. 3-Dimensional JEG-3 choriocarcinoma cell organoids as a model for trophoblast expansion and differentiation. Placenta 2021, 104, 243–246. [Google Scholar]
- Zhu, X.-M.; Han, T.; Wang, X.-H.; Li, Y.-H.; Yang, H.-G.; Luo, Y.-N.; Yin, G.-W.; Yao, Y.-Q. Overexpression of miR-152 leads to reduced expression of human leukocyte antigen-G and increased natural killer cell mediated cytolysis in JEG-3 cells. Am. J. Obstet. Gynecol. 2010, 202, 592.e1–592.e7. [Google Scholar] [CrossRef] [PubMed]
- Michita, R.T.; Zambra, F.M.B.; Fraga, L.R.; Sanseverino, M.T.V.; Callegari-Jacques, S.M.; Vianna, P.; Chies, J.A.B. A tug-of-war between tolerance and rejection—New evidence for 3′UTR HLA-G haplotypes influence in recurrent pregnancy loss. Hum. Immunol. 2016, 77, 892–897. [Google Scholar] [CrossRef] [PubMed]
- Bai, W.; Lin, Z.; Ye, J.; Zhou, L.; Xi, J.; Cai, W. Frequency of HLA-G UTR-1/UTR-3/UTR-7 in women with unexplained recurrent spontaneous abortion. Exp. Ther. Med. 2022, 24, 729. [Google Scholar] [CrossRef] [PubMed]
- Castelli, E.C.; Veiga-Castelli, L.C.; Yaghi, L.; Moreau, P.; Donadi, E.A. Transcriptional and posttranscriptional regulations of the HLA-G gene. J. Immunol. Res. 2014, 2014, 734068. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dias, F.C.; Bertol, B.C.; Poras, I.; Souto, B.M.; Mendes-Junior, C.T.; Castelli, E.C.; Gineau, L.; Sabbagh, A.; Rouas-Freiss, N.; Carosella, E.D.; et al. The genetic diversity within the 1.4 kb HLA-G 5’ upstream regulatory region moderately impacts on cellular microenvironment responses. Sci. Rep. 2018, 8, 5652. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Agrawal, D.; Prakash, S.; Misra, M.K.; Phadke, S.R.; Agrawal, S. Implication of HLA-G 5′ upstream regulatory region polymorphisms in idiopathic recurrent spontaneous abortions. Reprod. Biomed. Online 2015, 30, 82–91. [Google Scholar] [CrossRef] [Green Version]
- Tang, Y.; Liu, H.; Li, H.; Peng, T.; Gu, W.; Li, X. Hypermethylation of the HLA-G promoter is associated with preeclampsia. Mol. Hum. Reprod. 2015, 21, 736–744. [Google Scholar] [CrossRef] [Green Version]
- Moslehi, A.; Hashemi-Beni, B.; Moslehi, A.; Akbari, M.A.; Adib, M. The effect of progesterone and 17-β estradiol on mem-brane-bound HLA-G in adipose derived stem cells. Korean J. Physiol. Pharmacol. 2016, 20, 341. [Google Scholar] [CrossRef]
- Yie, S.-M.; Li, L.-H.; Li, G.-M.; Xiao, R.; Librach, C.L. Progesterone enhances HLA-G gene expression in JEG-3 choriocarcinoma cells and human cytotrophoblasts in vitro. Hum. Reprod. 2006, 21, 46–51. [Google Scholar] [CrossRef] [Green Version]
- Akhter, A.; Faridi, R.M.; Das, V.; Pandey, A.; Naik, S.; Agrawal, S. In vitro up-regulation of HLA-G using dexamethasone and hydrocortisone in first-trimester trophoblast cells of women experiencing recurrent miscarriage. Tissue Antigens 2012, 80, 126–135. [Google Scholar] [CrossRef]
- Hakam, M.S.; Miranda-Sayago, J.M.; Hayrabedyan, S.; Todorova, K.; Spencer, P.S.; Jabeen, A.; Barnea, E.R.; Fernandez, N. Preimplantation Factor (PIF) Promotes HLA-G, -E, -F, -C Expression in JEG-3 Choriocarcinoma Cells and Endogenous Progesterone Activity. Cell Physiol. Biochem. 2017, 43, 2277–2296. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, G.; Zhang, R.; Li, C.; Chen, A. Evaluation of the effect of low molecular weight heparin in unexplained recurrent pregnancy loss: A meta-analysis of randomized controlled trials. J. Matern. Neonatal. Med. 2022, 35, 7601–7608. [Google Scholar] [CrossRef] [PubMed]
- Zhang, J.; Li, H.; Fan, B.; Xu, W.; Zhang, X. Extracellular vesicles in normal pregnancy and pregnancy-related diseases. J. Cell. Mol. Med. 2020, 24, 4377–4388. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Buca, D.; Bologna, G.; D’Amico, A.; Cugini, S.; Musca, F.; Febbo, M.; D’Arcangelo, D.; Buca, D.; Simeone, P.; Liberati, M.; et al. Extracellular Vesicles in Feto–Maternal Crosstalk and Pregnancy Disorders. Int. J. Mol. Sci. 2020, 21, 2120. [Google Scholar] [CrossRef] [Green Version]
- Tersigni, C.; Lucchetti, D.; Franco, R.; Colella, F.; Neri, C.; Crispino, L.; Sgambato, A.; Lanzone, A.; Scambia, G.; Vatish, M.; et al. Circulating Placental Vesicles Carry HLA-DR in Pre-Eclampsia: A New Potential Marker of the Syndrome. Front. Immunol. 2021, 12, 717879. [Google Scholar]
- Tersigni, C.; Redman, C.; Dragovic, R.; Tannetta, D.; Scambia, G.; DI Simone, N.; Sargent, I.; Vatish, M. HLA-DR is aberrantly expressed at feto-maternal interface in pre-eclampsia. J. Reprod. Immunol. 2018, 129, 48–52. [Google Scholar] [CrossRef]
- Jiang, L.; Fei, H.; Jin, X.; Liu, X.; Yang, C.; Li, C.; Chen, J.; Yang, A.; Zhu, J.; Wang, H.; et al. Extracellular Vesicle-Mediated Secretion of HLA-E by Trophoblasts Maintains Pregnancy by Regulating the Metabolism of Decidual NK Cells. Int. J. Biol. Sci. 2021, 17, 4377–4395. [Google Scholar] [CrossRef]
- King, A.; Allan, D.S.; Bowen, M.; Powis, S.J.; Joseph, S.; Verma, S.; Hiby, S.E.; McMichael, A.J.; Loke, Y.W.; Braud, V.M. HLA-E is expressed on trophoblast and interacts with CD94/NKG2 receptors on decidual NK cells. Eur. J. Immunol. 2000, 30, 1623–1631. [Google Scholar]
- Rajaratnam, N.; Ditlevsen, N.E.; Sloth, J.K.; Bæk, R.; Jørgensen, M.M.; Christiansen, O.B. Extracellular Vesicles: An Important Biomarker in Recurrent Pregnancy Loss? J. Clin. Med. 2021, 10, 2549. [Google Scholar] [CrossRef]
- Jørgensen, M.M.; Bæk, R.; Sloth, J.; Varming, K.; Christiansen, O.B.; Ditlevsen, N.E.; Rajaratnam, N. Treatment with intravenous im-munoglobulin increases the level of small EVs in plasma of pregnant women with recurrent pregnancy loss. J. Reprod. Immunol. 2020, 140, 103128. [Google Scholar] [CrossRef]
- Yang, Y.; Wang, W.; Weng, J.; Li, H.; Ma, Y.; Liu, L.; Ma, W. Advances in the study of HLA class Ib in maternal-fetal immune tolerance. Front. Immunol. 2022, 13, 976289. [Google Scholar] [CrossRef] [PubMed]
- Beneventi, F.; Locatelli, E.; De Amici, M.; Martinetti, M.; Spinillo, A. Soluble HLA-G concentrations in obese women during preg-nancy and in cord blood. J. Reprod. Immunol. 2017, 119, 31–37. [Google Scholar] [CrossRef] [PubMed]
- Amodio, G.; Gregori, S. HLA-G Genotype/Expression/Disease Association Studies: Success, Hurdles, and Perspectives. Front. Immunol. 2020, 11, 1178. [Google Scholar] [CrossRef] [PubMed]




{kind=link}
{kind=link}
{kind=link}
| Role of HLA-G in Physiological Pregnancy | Role of HLA-G in Recurrent Pregnancy Loss |
|---|---|
|
|
| Physiological Pregnancy | Recurrent Pregnancy Loss |
|---|---|
|
|
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Barbaro, G.; Inversetti, A.; Cristodoro, M.; Ticconi, C.; Scambia, G.; Di Simone, N. HLA-G and Recurrent Pregnancy Loss. Int. J. Mol. Sci. 2023, 24, 2557. https://doi.org/10.3390/ijms24032557
Barbaro G, Inversetti A, Cristodoro M, Ticconi C, Scambia G, Di Simone N. HLA-G and Recurrent Pregnancy Loss. International Journal of Molecular Sciences. 2023; 24(3):2557. https://doi.org/10.3390/ijms24032557
Chicago/Turabian StyleBarbaro, Greta, Annalisa Inversetti, Martina Cristodoro, Carlo Ticconi, Giovanni Scambia, and Nicoletta Di Simone. 2023. "HLA-G and Recurrent Pregnancy Loss" International Journal of Molecular Sciences 24, no. 3: 2557. https://doi.org/10.3390/ijms24032557