
Diagnostic Utility of TSSC3 and RB1 Immunohistochemistry in Hydatidiform Mole
 , , , and
, , , and
Abstract
:1. Introduction
2. Results
2.1. Demographic and Clinical Characteristics
2.2. Immunohistochemical Study of Paternal- and Maternal-Imprinted Genes
2.2.1. Paternally Imprinted Genes
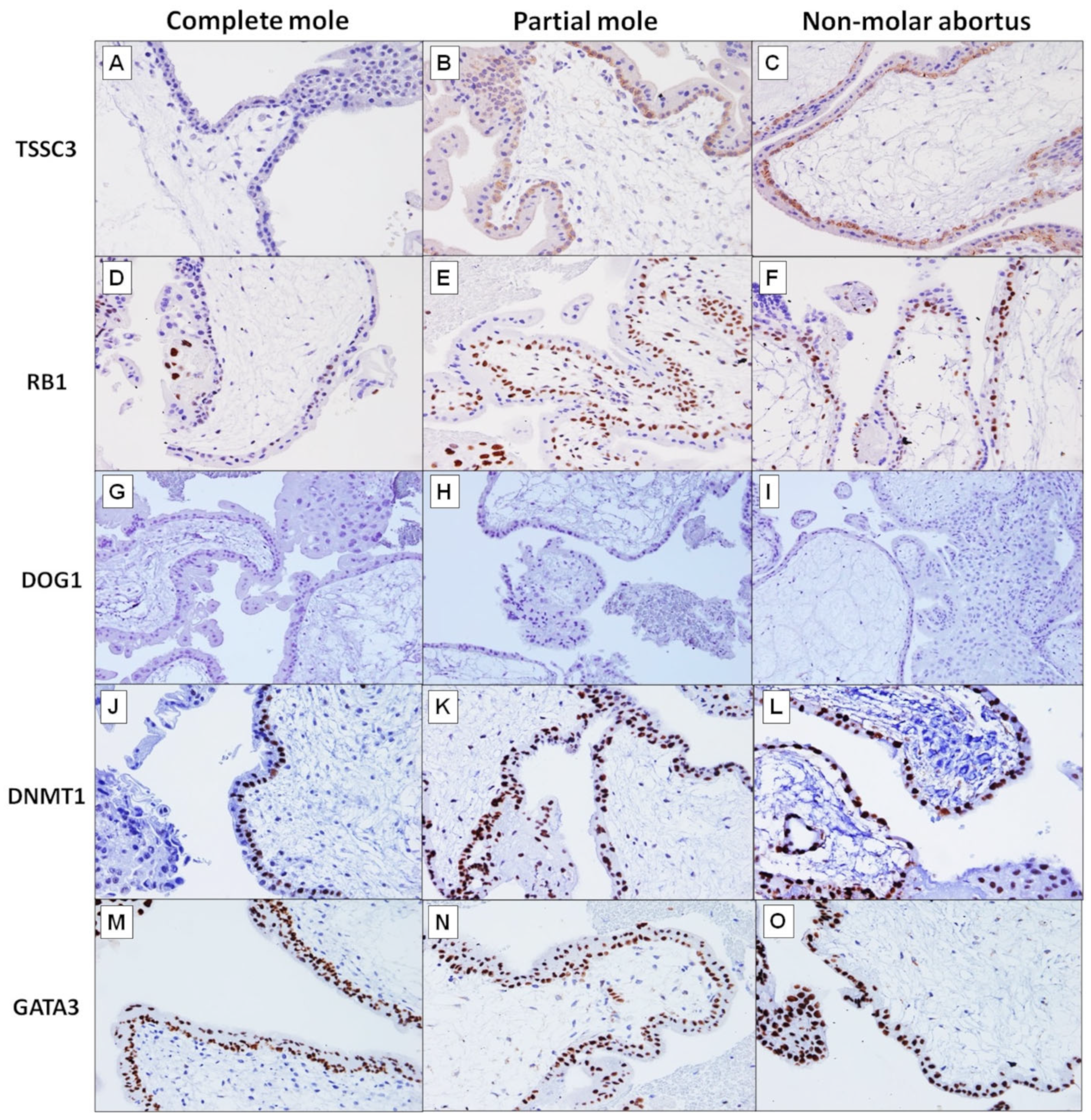
TSSC3 Immunohistochemistry
RB1 Immunohistochemistry
DOG1 Immunohistochemistry
2.2.2. Maternally Imprinted Genes
DNMT1 Immunohistochemistry
GATA3 Immunohistochemistry
2.3. TSSC3 and RB1 Immunohistochemistry in Equivocal Cases
3. Discussion
4. Materials and Methods
4.1. Formalin-Fixed Paraffin-Embedded Hydatidiform Mole Tissue Samples
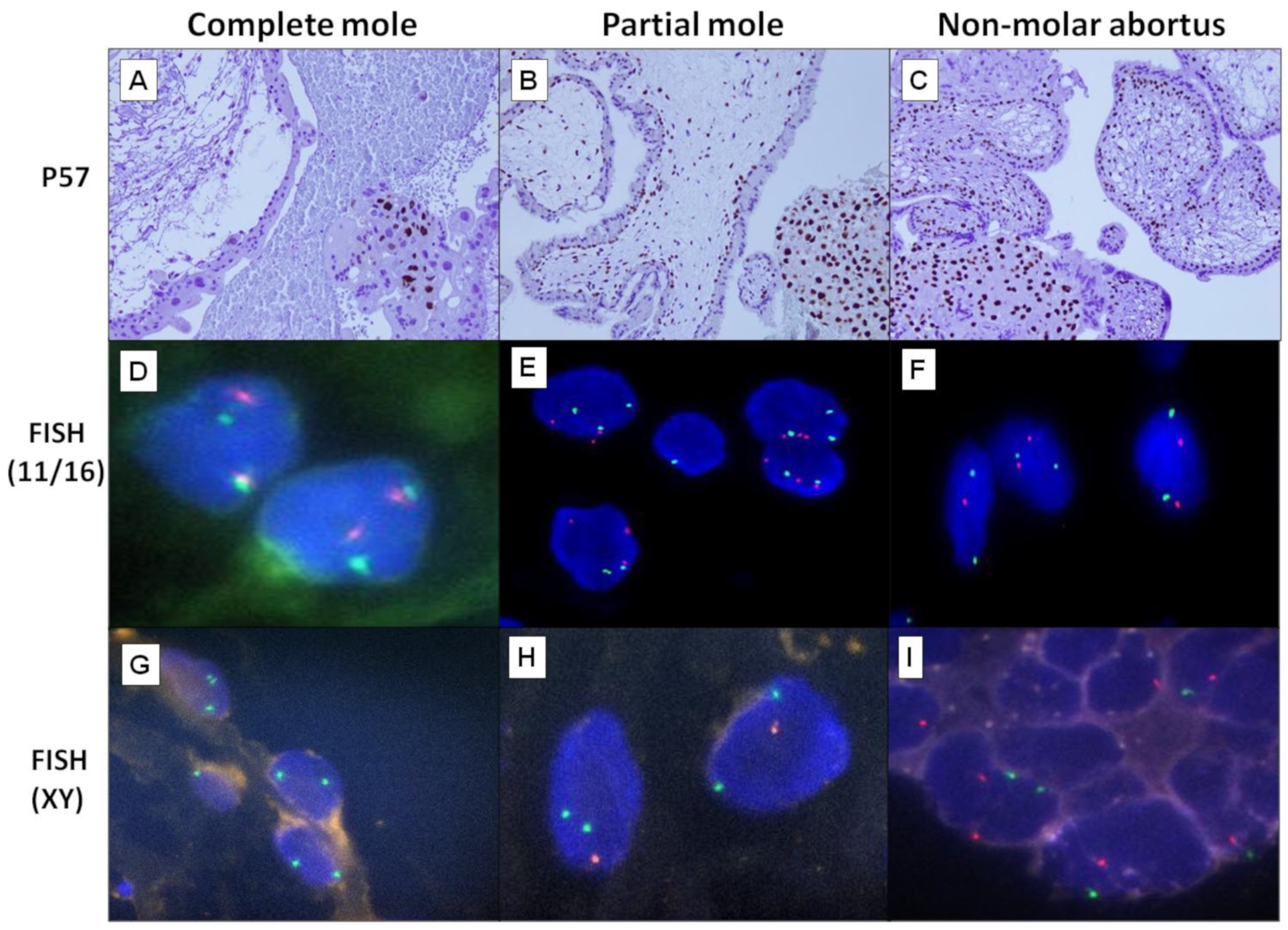
4.2. DNA Ploidy Study by Fluorescence In Situ Hybridization
4.3. Immunohistochemistry of Paternally and Maternally Imprinted Genes
4.4. Statistical Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Yamamoto, E.; Trinh, T.D.; Sekiya, Y.; Tamakoshi, K.; Nguyen, X.P.; Nishino, K.; Niimi, K.; Kotani, T.; Kajiyama, H.; Shibata, K.; et al. The management of hydatidiform mole using prophylactic chemotherapy and hysterectomy for high-risk patients decreased the incidence of gestational trophoblastic neoplasia in Vietnam: A retrospective observational study. Nagoya J. Med. Sci. 2020, 82, 183–191. [Google Scholar]
- Tham, B.W.L.; Everard, J.E.; Tidy, J.A.; Drew, D.; Hancock, B.W. Gestational trophoblastic disease in the Asian population of Northern England and North Wales. BJOG 2003, 110, 555–559. [Google Scholar] [CrossRef]
- Zainal, N.; Kampan, N.C.; Rose, I.M.; Ghazali, R.; Shafiee, M.N.; Yussoff, N.H.; Tamil, A.; Jamil, M.A.; Hussin, N.H. Complementary role of p57kip2 immunostaining in diagnosing hydatidiform mole subtypes. Horm. Mol. Biol. Clin. Investig. 2021, 42, 311–316. [Google Scholar] [CrossRef] [PubMed]
- Nirmala, C.K.; Nor Azlin, M.I.; Harry, S.R.; Lim, P.S.; Shafiee, M.N.; Nur Azurah, A.G.; Omar, M.H.; Hatta, M.D. Outcome of molar pregnancies in Malaysia: A tertiary centre experience. J. Obstet. Gynaecol. 2013, 33, 191–193. [Google Scholar] [CrossRef] [PubMed]
- Wong, Y.P.; Chia, W.K.; Selimin, A.; Chia, P.Y.; Mustangin, M.; Shuib, S.; Khong, T.Y.; Tan, G.C. Diagnostic utility of p57 immunohistochemistry and DNA ploidy analysis by fluorescence in situ hybridisation in hydatidiform moles. Malays. J. Pathol. 2021, 43, 341–351. [Google Scholar] [PubMed]
- Prabhu, I.K.; Rosenbaum, A. Hydatidiform Mole in a Patient With a Concern for Neoplasia: A Case Report. Cureus 2020, 12, e10319. [Google Scholar] [CrossRef] [PubMed]
- Madi, J.M.; Braga, A.; Paganella, M.P.; Litvin, I.E.; Wendland, E.M. Accuracy of p57KIP2 compared with genotyping to diagnose complete hydatidiform mole: A systematic review and meta-analysis. BJOG 2018, 125, 1226–1233. [Google Scholar] [CrossRef] [Green Version]
- Ronnett, B.M. Hydatidiform moles: Ancillary techniques to refine diagnosis. Arch. Pathol. Lab. Med. 2018, 142, 1485–1502. [Google Scholar] [CrossRef] [Green Version]
- Castrillon, D.H.; Sun, D.; Weremowicz, S.; Fisher, R.A.; Crum, C.P.; Genest, D.R. Discrimination of complete hydatidiform mole from its mimics by immunohistochemistry of the paternally imprinted gene product p57KIP2. Am. J. Surg. Pathol. 2001, 25, 1225–1230. [Google Scholar] [CrossRef]
- Tycko, B. Imprinted genes in placental growth and obstetric disorders. Cytogenet. Genome Res. 2006, 113, 271–278. [Google Scholar] [CrossRef]
- Hayati, A.R.; Tan, G.C. Clinicopathologic and immunohistochemical differences in complete and partial hydatidiform moles in a multiracial Malaysian population. Int. J. Gynecol. Pathol. 2005, 24, 277–285. [Google Scholar] [CrossRef] [PubMed]
- Kihara, M.; Matsui, H.; Seki, K.; Nagai, Y.; Wake, N.; Sekiya, S. Genetic origin and imprinting in hydatidiform moles. Comparison between DNA polymorphism analysis and immunoreactivity of p57KIP2. J. Reprod. Med. 2005, 50, 307–312. [Google Scholar] [PubMed]
- Osterheld, M.C.; Caron, L.; Chaubert, P.; Meagher-Villemure, K. Combination of immunohistochemistry and ploidy analysis to assist histopathological diagnosis of molar diseases. Clin. Med. Pathol. 2008, 1, 61–67. [Google Scholar] [CrossRef] [PubMed]
- Khooei, A.; Atabaki Pasdar, F.; Fazel, A.; Mahmoudi, M.; Nikravesh, M.R.; Khaje Delui, M.; Pourheydar, B. Ki-67 expression in hydatidiform moles and hydropic abortions. Iran. Red. Crescent. Med. J. 2013, 15, 590–594. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lund, H.; Nielsen, S.; Grove, A.; Vyberg, M.; Sunde, L. p57 in Hydatidiform Moles: Evaluation of Antibodies and Expression in Various Cell Types. Appl. Immunohistochem. Mol. Morphol. 2020, 28, 694–701. [Google Scholar] [CrossRef] [PubMed]
- McConnell, T.G.; Murphy, K.M.; Hafez, M.; Vang, R.; Ronnett, B.M. Diagnosis and subclassification of hydatidiform moles using p57 immunohistochemistry and molecular genotyping: Validation and prospective analysis in routine and consultation practice settings with development of an algorithmic approach. Am. J. Surg. Pathol. 2009, 33, 805–817. [Google Scholar] [CrossRef] [Green Version]
- Furtado, L.V.; Paxton, C.N.; Jama, M.A.; Tripp, S.R.; Wilson, A.R.; Lyon, E.; Jarboe, E.A.; Thaker, H.M.; Geiersbach, K.B. Diagnostic utility of microsatellite genotyping for molar pregnancy testing. Arch. Pathol. Lab. Med. 2013, 137, 55–63. [Google Scholar] [CrossRef] [Green Version]
- Imprinted Genes: By Species. Available online: https://www.geneimprint.com/site/genes-by-species.Homo+sapiens.imprinted-M (accessed on 9 March 2023).
- Xing, D.; Adams, E.; Huang, J.; Ronnett, B.M. Refined diagnosis of hydatidiform moles with p57 immunohistochemistry and molecular genotyping: Updated analysis of a prospective series of 2217 cases. Mod. Pathol. 2021, 34, 961–982. [Google Scholar] [CrossRef]
- Bajrami, E.; Spiroski, M. Genomic Imprinting. Open Access Maced. J. Med. Sci. 2016, 4, 181–184. [Google Scholar] [CrossRef] [Green Version]
- Isles, A.R.; Davies, W.; Wilkinson, L.S. Genomic imprinting and the social brain. Philos. Trans. R Soc. Lond. B Biol. Sci. 2006, 361, 2229–2237. [Google Scholar] [CrossRef] [Green Version]
- Plasschaert, R.N.; Bartolomei, M.S. Genomic imprinting in development, growth, behavior and stem cells. Development 2014, 141, 1805–1813. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hui, P.; Buza, N.; Murphy, K.M.; Ronnett, B.M. Hydatidiform Moles: Genetic Basis and Precision Diagnosis. Annu. Rev. Pathol. 2017, 12, 449–485. [Google Scholar] [CrossRef]
- Creeth, H.D.J.; McNamara, G.I.; Isles, A.R.; John, R.M. Imprinted genes influencing the quality of maternal care. Front. Neuroendocrinol. 2019, 53, 100732. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fisher, R.A.; Hodges, M.D.; Newlands, E.S. Familial recurrent hydatidiform mole: A review. J. Reprod. Med 2004, 49, 595–601. [Google Scholar] [PubMed]
- Murdoch, S.; Djuric, U.; Mazhar, B.; Seoud, M.; Khan, R.; Kuick, R.; Bagga, R.; Kircheisen, R.; Ao, A.; Ratti, B.; et al. Mutations in NALP7 cause recurrent hydatidiform moles and reproductive wastage in humans. Nat. Genet 2006, 38, 300–302. [Google Scholar] [CrossRef]
- Ngan, H.Y.S.; Seckl, M.J.; Berkowitz, R.S.; Xiang, Y.; Golfier, F.; Sekharan, P.K.; Lurain, J.R.; Massuger, L. Update on the diagnosis and management of gestational trophoblastic disease. Int. J. Gynaecol. Obstet. 2018, 143 (Suppl. 2), 79–85. [Google Scholar] [CrossRef] [Green Version]
- Sebire, N.J.; Foskett, M.; Fisher, R.A.; Rees, H.; Seckl, M.; Newlands, E. Risk of partial and complete hydatidiform molar pregnancy in relation to maternal age. BJOG 2002, 109, 99–102. [Google Scholar] [CrossRef]
- Cavaliere, A.; Ermito, S.; Dinatale, A.; Pedata, R. Management of molar pregnancy. J. Prenat. Med. 2009, 3, 15–17. [Google Scholar]
- Wong, Y.P.; Tan, G.C.; Aziz, S.; Pongprakyun, S.; Ismail, F. Beta-human Chorionic Gonadotropin-secreting Lung Adenocarcinoma. Malays. J. Med. Sci. 2015, 22, 76–80. [Google Scholar]
- Mondal, S.K.; Mandal, S.; Bhattacharya, S.; Panda, U.K.; Ray, A.; Ali, S.M. Expression of p57 immunomarker in the classification and differential diagnosis of partial and complete hydatidiform moles. J. Lab. Physicians 2019, 11, 270–274. [Google Scholar] [CrossRef]
- López, C.L.; Figueira Gouvêa, A.L.; Rodrigues, F.R.; Braga, A.; Valente Machado, M.D.; Lopes, V.S. Human epidermal growth factor receptor 2 fluorescence in situ hybridization and P57KIP2 immunohistochemistry using tissue microarray: Improving histopathological subtyping of hydatidiform mole. Placenta 2020, 99, 166–172. [Google Scholar] [CrossRef] [PubMed]
- Salas, M.; John, R.; Saxena, A.; Barton, S.; Frank, D.; Fitzpatrick, G.; Higgins, M.J.; Tycko, B. Placental growth retardation due to loss of imprinting of Phlda2. Mech. Dev. 2004, 121, 1199–1210. [Google Scholar] [CrossRef] [PubMed]
- Wrzeska, M.; Rejduch, B. Genomic imprinting in mammals. J. Appl. Genet. 2004, 45, 427–433. [Google Scholar]
- Piedrahita, J.A. The role of imprinted genes in fetal growth abnormalities. Birth Defects Res. A Clin. Mol. Teratol. 2011, 91, 682–692. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Saxena, A.; Frank, D.; Panichkul, P.; Van den Veyver, I.B.; Tycko, B.; Thaker, H. The product of the imprinted gene IPL marks human villous cytotrophoblast and is lost in complete hydatidiform mole. Placenta 2003, 24, 835–842. [Google Scholar] [CrossRef] [PubMed]
- Lee, M.P.; Feinberg, A.P. Genomic imprinting of a human apoptosis gene homologue, TSSC3. Cancer Res 1998, 58, 1052–1056. [Google Scholar]
- Frank, D.; Fortino, W.; Clark, L.; Musalo, R.; Wang, W.; Saxena, A.; Li, C.M.; Reik, W.; Ludwig, T.; Tycko, B. Placental overgrowth in mice lacking the imprinted gene Ipl. Proc. Natl. Acad. Sci. USA 2002, 99, 7490–7495. [Google Scholar] [CrossRef] [Green Version]
- Takao, T.; Asanoma, K.; Tsunematsu, R.; Kato, K.; Wake, N. The maternally expressed gene Tssc3 regulates the expression of MASH2 transcription factor in mouse trophoblast stem cells through the AKT-Sp1 signaling pathway. J. Biol. Chem. 2012, 287, 42685–42694. [Google Scholar] [CrossRef] [Green Version]
- Kato, H.; Matsuda, T.; Hirakawa, T.; Ueda, K.; Inoue, T.; Miyanari, Y.; Asanoma, K.; Nakano, H.; Wake, N. Differential diagnosis between complete and partial mole by TSSC3 antibody completely correlates to DNA diagnosis. Diagn. Mol. Pathol. 2005, 14, 164–169. [Google Scholar] [CrossRef]
- Thaker, H.M.; Berlin, A.; Tycko, B.; Goldstein, D.P.; Berkowitz, R.S.; Castrillon, D.H.; Genest, D.R. Immunohistochemistry for the imprinted gene product IPL/PHLDA2 for facilitating the differential diagnosis of complete hydatidiform mole. J. Reprod. Med. 2004, 49, 630–636. [Google Scholar]
- Kanber, D.; Berulava, T.; Ammerpohl, O.; Mitter, D.; Richter, J.; Siebert, R.; Horsthemke, B.; Lohmann, D.; Buiting, K. The human retinoblastoma gene is imprinted. PLoS Genet. 2009, 5, e1000790. [Google Scholar] [CrossRef] [PubMed]
- Buiting, K.; Kanber, D.; Horsthemke, B.; Lohmann, D. Imprinting of RB1 (the new kid on the block). Brief Funct. Genomics 2010, 9, 347–353. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rahat, B.; Thakur, S.; Hamid, A.; Bagga, R.; Kaur, J. Association of aberrant methylation at promoter regions of tumor suppressor genes with placental pathologies. Epigenomics 2016, 8, 767–787. [Google Scholar] [CrossRef] [PubMed]
- Matsuoka, S.; Thompson, J.S.; Edwards, M.C.; Bartletta, J.M.; Grundy, P.; Kalikin, L.M.; Harper, J.W.; Elledge, S.J.; Feinberg, A.P. Imprinting of the gene encoding a human cyclin-dependent kinase inhibitor, p57KIP2, on chromosome 11p15. Proc. Natl. Acad. Sci. USA 1996, 93, 3026–3030. [Google Scholar] [CrossRef] [Green Version]
- Miettinen, M.; Wang, Z.F.; Lasota, J. DOG1 antibody in the differential diagnosis of gastrointestinal stromal tumors: A study of 1840 cases. Am. J. Surg. Pathol. 2009, 33, 1401–1408. [Google Scholar] [CrossRef] [PubMed]
- Lezcano, C.; Ho, J.; Seethala, R.R. Sox10 and DOG1 Expression in Primary Adnexal Tumors of the Skin. Am. J. Dermatopathol. 2017, 39, 896–902. [Google Scholar] [CrossRef]
- Mirkovic, J.; Elias, K.; Drapkin, R.; Barletta, J.A.; Quade, B.; Hirsch, M.S. GATA3 expression in gestational trophoblastic tissues and tumours. Histopathology 2015, 67, 636–644. [Google Scholar] [CrossRef]
- Banet, N.; Gown, A.M.; Shih, I.M.; Kay Li, Q.; Roden, R.B.; Nucci, M.R.; Cheng, L.; Przybycin, C.G.; Nasseri-Nik, N.; Wu, L.S.; et al. GATA-3 expression in trophoblastic tissues: An immunohistochemical study of 445 cases, including diagnostic utility. Am. J. Surg. Pathol. 2015, 39, 101–108. [Google Scholar] [CrossRef] [Green Version]
- Robaina, M.C.; Mazzoccoli, L.; Arruda, V.O.; Reis, F.R.S.; Apa, A.G.; de Rezende, L.M.M.; Klumb, C.E. Deregulation of DNMT1, DNMT3B and miR-29s in Burkitt lymphoma suggests novel contribution for disease pathogenesis. Exp. Mol. Pathol. 2015, 98, 200–207. [Google Scholar] [CrossRef]
- Mukhopadhyay, A.; Ravikumar, G.; Meraaj, H.; Dwarkanath, P.; Thomas, A.; Crasta, J.; Thomas, T.; Kurpad, A.V.; Sridhar, T.S. Placental expression of DNA methyltransferase 1 (DNMT1): Gender-specific relation with human placental growth. Placenta 2016, 48, 119–125. [Google Scholar] [CrossRef]
- Sharifah, N.A.; Zakaria, Z.; Chia, W.K. FISH analysis using PPAR γ-specific probes for detection of PAX8-PPAR γ translocation in follicular thyroid neoplasms. Methods. Mol. Biol. 2013, 952, 187–196. [Google Scholar] [PubMed]



{kind=link}
{kind=link}
| Demographic | CM n (%) | PM n (%) | NMA n (%) | p Value CM vs. PM |
|---|---|---|---|---|
| Age range (years old) | 21–51 | 24–38 | 21–42 | |
| Mean (SD) | 34.9 (8.841) | 31.3 (4.03) | 33.7 (6.828) | p = 0.4567 |
| 20–29 years old | 9 (31.0%) | 5 (33.3%) | 5 (29.4%) | |
| 30–39 years old | 12 (41.4%) | 10 (66.7%) | 7 (41.2%) | |
| 40 years old and above | 8 (27.6%) | 0 (0%) | 5 (29.4%) | |
| Gestational age range (weeks) | 7–15 | 9–18 | 8–12 | |
| Mean (SD) a | 10.60 (2.415) | 13.17 (3.125) | 10.57 (1.284) | p = 0.0247 * |
| β-hCG levels range (mIU/mL) | 22,683–1,000,000 | 6985–369,109 | <1.2–25,888 | |
| Mean (SD) b | 234,024 (250,856) | 102,174 (135,505) | 10,124 (12,563) | p = 0.0227 * |
| Ethnicity | ||||
| Malay | 23 (79.3%) | 15 (100%) | 15 (88.2%) | |
| Chinese | 3 (10.3%) | 0 (0%) | 2 (11.8%) | |
| Others (Punjabi, Caucasian) | 3 (10.3%) | 0 (0%) | 0 (0%) |
| TSSC3 | CM | PM | NMA | p Value CM vs. PM | p Value CM vs. NMA | |
|---|---|---|---|---|---|---|
| CT | Positive | 9 | 15 | 17 | <0.0001 * | <0.0001 * |
| Negative | 20 | 0 | 0 | |||
| VSC | Positive | 0 | 2 | 0 | 0.111 | 1.0 |
| Negative | 29 | 13 | 17 | |||
| ST | Positive | 0 | 0 | 0 | 1.0 | 1.0 |
| Negative | 29 | 15 | 17 | |||
| IT | Positive | 2 | 5 | 4 | 0.1656 | 0.0358 |
| Negative | 27 | 10 | 12 | |||
| DC | Positive | 23 | 12 | 12 | 0.2785 | 0.2785 |
| Negative | 1 | 3 | 3 | |||
| RB1 | ||||||
| CT | Positive | 3 | 15 | 16 | <0.0001 * | <0.0001 * |
| Negative | 26 | 0 | 1 | |||
| VSC | Positive | 1 | 0 | 0 | 1.0 | 1.0 |
| Negative | 28 | 15 | 17 | |||
| ST | Positive | 0 | 0 | 0 | 1.0 | 1.0 |
| Negative | 29 | 15 | 17 | |||
| IT | Positive | 29 | 15 | 17 | 1.0 | 1.0 |
| Negative | 0 | 0 | 0 | |||
| DC | Positive | 22 | 13 | 13 | 1.0 | 1.0 |
| Negative | 3 | 2 | 1 |
| DNMT1 | CM | PM | NMA | p Value CM vs. PM | p Value CM vs. NMA | |
|---|---|---|---|---|---|---|
| CT | Positive | 29 | 15 | 17 | 1.0 | 1.0 |
| Negative | 0 | 0 | 0 | |||
| VSC | Positive | 12 | 5 | 2 | 0.7477 | 0.0487 * |
| Negative | 17 | 10 | 15 | |||
| ST | Positive | 15 | 6 | 7 | 0.535 | 0.552 |
| Negative | 14 | 9 | 10 | |||
| IT | Positive | 27 | 14 | 13 | 1.0 | 0.1744 |
| Negative | 2 | 1 | 4 | |||
| DC | Positive | 13 | 8 | 8 | 1.0 | 1.0 |
| Negative | 10 | 7 | 5 | |||
| GATA3 | ||||||
| CT | Positive | 28 | 15 | 16 | 1.0 | 1.0 |
| Negative | 1 | 0 | 0 | |||
| VSC | Positive | 0 | 0 | 0 | 1.0 | 1.0 |
| Negative | 29 | 15 | 17 | |||
| ST | Positive | 27 | 15 | 16 | 0.5402 | 0.5313 |
| Negative | 2 | 0 | 0 | |||
| IT | Positive | 29 | 15 | 17 | 1.0 | 1.0 |
| Negative | 0 | 0 | 0 | |||
| DC | Positive | 1 | 8 | 2 | 0.0008 * | 0.547 |
| Negative | 23 | 7 | 13 |
| Sample No. | PM002 | PM003 | PM009 | PM019 |
|---|---|---|---|---|
| Maternal age (years) | 30 | 30 | 38 | 27 |
| Gestational age (weeks) | NA | 8 | 18 | Not available |
| β-hCG level (mIU/mL) | 184,108.50 | 181,113.20 | 33,906 | Not available |
| Preliminary diagnosis based on histomorphological features alone | ? Partial mole | Uncertain | ? Partial mole | ? Partial mole |
| p57 immunohistochemistry (Percentage of positivity) | 100% | 50% | 60% | 100% |
| DNA Ploidy Status | 2n, XX | 2n, XX | 3n, XXX | 2n, XX |
| Diagnosis (histomorphology + p57 + DNA ploidy study) | Non-molar abortus | Uncertain | Partial mole | Non-molar abortus |
| TSSC3 immunohistochemistry (Intensity/percentage) | Positive 3+/20% | Positive 2+/50% | Positive 3+/50% | Positive 3+/40% |
| RB1 immunohistochemistry (Percentage) | Positive 50% | Negative 40% | Positive 100% | Positive 100% |
| Final diagnosis | Non-molar abortus | Non-molar abortus | Partial mole | Non-molar abortus |
| TSSC3 and RB1 remarks | Reinforced the diagnosis | The p57 staining was equivocal in this case. TSSC3 showed 50% positivity; it is likely a non-molar abortus. Except for the rare possibility of a diploid biparental complete mole or a retention of maternal DNA. | Reinforced the diagnosis | Reinforced the diagnosis |
| Primary Antibodies [Clone Code] | Manufacturer | Catalog Number | Imprinted Gene | Expression Site | Control | Dilution Factor |
|---|---|---|---|---|---|---|
| P57 Rabbit monoclonal [EP2515Y] | Abcam, Cambridge, UK | AB75974 | Paternal | Nucleus | Placenta | 1:500 |
| RB Rabbit monoclonal [EPR17512] | Abcam, Cambridge, UK | AB181616 | Paternal | Nucleus | Tonsil | 1:1500 |
| TSSC3 Rabbit polyclonal | Abcam, Cambridge, UK | AB234669 | Paternal | Cytoplasm | Placenta | 1:300 |
| DOG1 Rabbit monoclonal (SP31) | Cell Marque, California, USA | 244R-14 | Paternal | Cytoplasm | GIST | 1:500 |
| DNMT1 Rabbit monoclonal [EPR18453] | Abcam, Cambridge, UK | AB188453 | Maternal | Nucleus | Tonsil | 1:100 |
| GATA3 Mouse monoclonal (L50-823) | Cell Marque, California, USA | 390M-14 | Maternal | Nucleus | Placenta | 1:500 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Chia, W.K.; Chia, P.Y.; Abdul Aziz, N.H.; Shuib, S.; Mustangin, M.; Cheah, Y.K.; Khong, T.Y.; Wong, Y.P.; Tan, G.C. Diagnostic Utility of TSSC3 and RB1 Immunohistochemistry in Hydatidiform Mole. Int. J. Mol. Sci. 2023, 24, 9656. https://doi.org/10.3390/ijms24119656
Chia WK, Chia PY, Abdul Aziz NH, Shuib S, Mustangin M, Cheah YK, Khong TY, Wong YP, Tan GC. Diagnostic Utility of TSSC3 and RB1 Immunohistochemistry in Hydatidiform Mole. International Journal of Molecular Sciences. 2023; 24(11):9656. https://doi.org/10.3390/ijms24119656
Chicago/Turabian StyleChia, Wai Kit, Pik Yuen Chia, Nor Haslinda Abdul Aziz, Salwati Shuib, Muaatamarulain Mustangin, Yoke Kqueen Cheah, Teck Yee Khong, Yin Ping Wong, and Geok Chin Tan. 2023. "Diagnostic Utility of TSSC3 and RB1 Immunohistochemistry in Hydatidiform Mole" International Journal of Molecular Sciences 24, no. 11: 9656. https://doi.org/10.3390/ijms24119656