
Regional Differences in Ca2+ Signaling and Transverse-Tubules across Left Atrium from Adult Sheep
 , ,
, ,
Abstract
:
{kind=link}
{kind=link}
{kind=link}
{kind=link}
1. Introduction
2. Results and Discussion
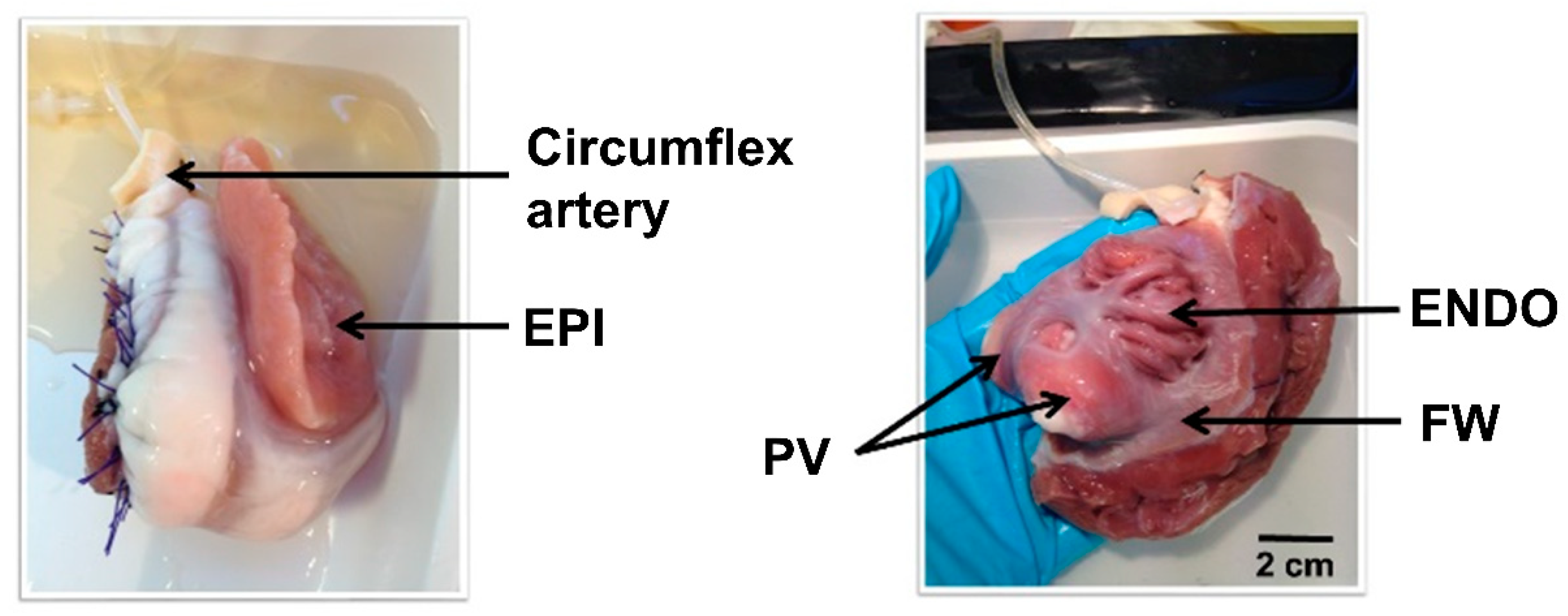
2.1. Regional Left Atria Myocyte Isolation
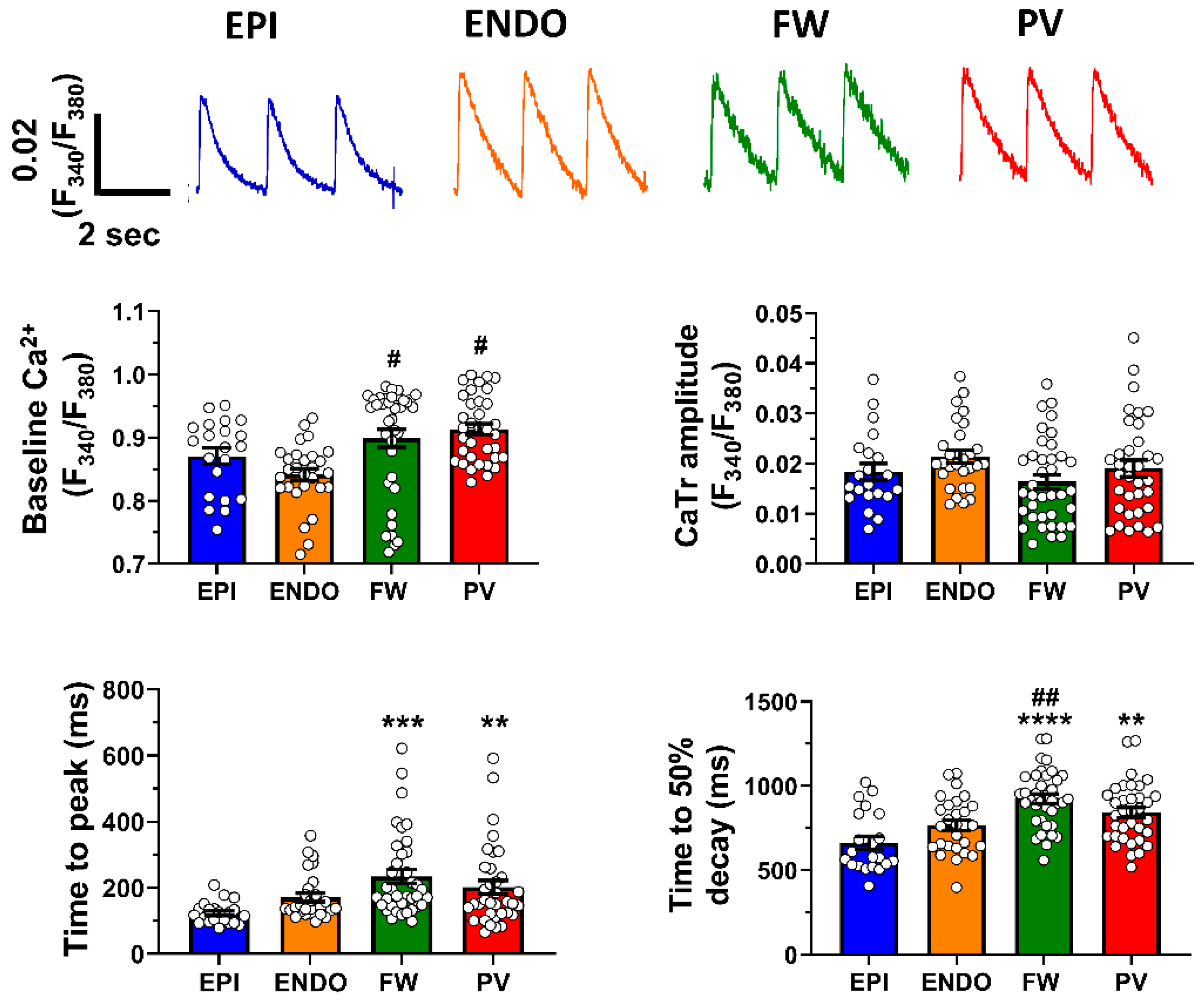
2.2. Ca2+ Signaling Heterogeneity over the Left Atrium
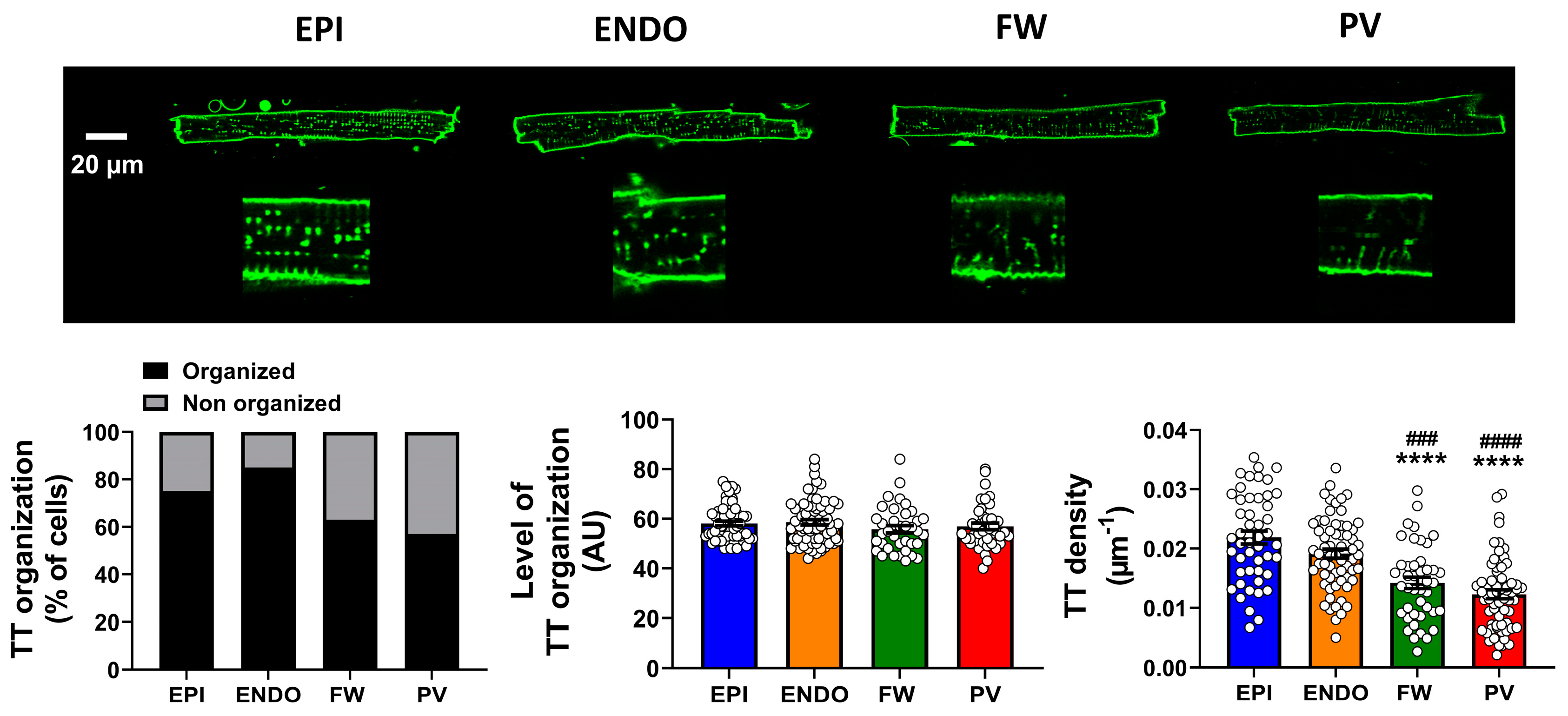
2.3. Heterogeneity of T-Tubule Network across the Left Atrium
3. Materials and Methods
3.1. Animal Ethics
3.2. Left Atria Cardiomyocyte Isolation
3.3. Whole Cell Ca2+ Transient Recording
3.4. Cell Membrane Staining
3.5. Rapid 2D Ca2+ Imaging
3.6. Data Analysis
3.7. Statistics
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Bers, D.M. Cardiac excitation-contraction coupling. Nature 2002, 415, 198–205. [Google Scholar] [CrossRef] [PubMed]
- Fabiato, A. Calcium-induced release of calcium from the cardiac sarcoplasmic reticulum. Am. J. Physiol. 1983, 245, C1–C14. [Google Scholar] [CrossRef]
- Brette, F.; Orchard, C. T-tubule function in mammalian cardiac myocytes. Circ. Res. 2003, 92, 1182–1192. [Google Scholar] [CrossRef] [PubMed]
- Dibb, K.M.; Clarke, J.D.; Horn, M.A.; Richards, M.A.; Graham, H.K.; Eisner, D.A.; Trafford, A.W. Characterization of an extensive transverse tubular network in sheep atrial myocytes and its depletion in heart failure. Circ. Heart Fail. 2009, 2, 482–489. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Brandenburg, S.; Kohl, T.; Williams, G.S.; Gusev, K.; Wagner, E.; Rog-Zielinska, E.A.; Hebisch, E.; Dura, M.; Didie, M.; Gotthardt, M.; et al. Axial tubule junctions control rapid calcium signaling in atria. J. Clin. Investig. 2016, 126, 3999–4015. [Google Scholar] [CrossRef] [Green Version]
- Schotten, U.; Verheule, S.; Kirchhof, P.; Goette, A. Pathophysiological mechanisms of atrial fibrillation: A translational appraisal. Physiol. Rev. 2011, 91, 265–325. [Google Scholar] [CrossRef]
- Setterberg, I.E.; Le, C.; Frisk, M.; Li, J.; Louch, W.E. The Physiology and Pathophysiology of T-Tubules in the Heart. Front. Physiol. 2021, 12, 718404. [Google Scholar] [CrossRef]
- Lenaerts, I.; Bito, V.; Heinzel, F.R.; Driesen, R.B.; Holemans, P.; D’Hooge, J.; Heidbuchel, H.; Sipido, K.R.; Willems, R. Ultrastructural and functional remodeling of the coupling between Ca2+ influx and sarcoplasmic reticulum Ca2+ release in right atrial myocytes from experimental persistent atrial fibrillation. Circ. Res. 2009, 105, 876–885. [Google Scholar] [CrossRef] [Green Version]
- Macquaide, N.; Tuan, H.T.; Hotta, J.; Sempels, W.; Lenaerts, I.; Holemans, P.; Hofkens, J.; Jafri, M.S.; Willems, R.; Sipido, K.R. Ryanodine receptor cluster fragmentation and redistribution in persistent atrial fibrillation enhance calcium release. Cardiovasc. Res. 2015, 108, 387–398. [Google Scholar] [CrossRef] [Green Version]
- Martins, R.P.; Kaur, K.; Hwang, E.; Ramirez, R.J.; Willis, B.C.; Filgueiras-Rama, D.; Ennis, S.R.; Takemoto, Y.; Ponce-Balbuena, D.; Zarzoso, M.; et al. Dominant frequency increase rate predicts transition from paroxysmal to long-term persistent atrial fibrillation. Circulation 2014, 129, 1472–1482. [Google Scholar] [CrossRef]
- Arora, R.; Aistrup, G.L.; Supple, S.; Frank, C.; Singh, J.; Tai, S.; Zhao, A.; Chicos, L.; Marszalec, W.; Guo, A.; et al. Regional distribution of T-tubule density in left and right atria in dogs. Heart Rhythm 2017, 14, 273–281. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gadeberg, H.C.; Bond, R.C.; Kong, C.H.T.; Chanoit, G.P.; Ascione, R.; Cannell, M.B.; James, A.F. Heterogeneity of T-Tubules in Pig Hearts. PLoS ONE 2016, 11, e0156862. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Haissaguerre, M.; Jais, P.; Shah, D.C.; Takahashi, A.; Hocini, M.; Quiniou, G.; Garrigue, S.; Le Mouroux, A.; Le Metayer, P.; Clementy, J. Spontaneous initiation of atrial fibrillation by ectopic beats originating in the pulmonary veins. N. Engl. J. Med. 1998, 339, 659–666. [Google Scholar] [CrossRef] [Green Version]
- Okamoto, Y.; Takano, M.; Ohba, T.; Ono, K. Arrhythmogenic coupling between the Na+–Ca2+ exchanger and inositol 1,4,5-triphosphate receptor in rat pulmonary vein cardiomyocytes. J. Mol. Cell. Cardiol. 2012, 52, 988–997. [Google Scholar] [CrossRef]
- Pasqualin, C.; Yu, A.; Malécot, C.O.; Gannier, F.; Cognard, C.; Godin-Ribuot, D.; Morand, J.; Bredeloux, P.; Maupoil, V. Structural heterogeneity of the rat pulmonary vein myocardium: Consequences on intracellular calcium dynamics and arrhythmogenic potential. Sci. Rep. 2018, 8, 3244. [Google Scholar] [CrossRef] [Green Version]
- Schüttler, D.; Bapat, A.; Kääb, S.; Lee, K.; Tomsits, P.; Clauss, S.; Hucker, W.J. Animal Models of Atrial Fibrillation. Circ. Res. 2020, 127, 91–110. [Google Scholar] [CrossRef] [PubMed]
- Gomez, A.M.; Valdivia, H.H.; Cheng, H.; Lederer, M.R.; Santana, L.F.; Cannell, M.B.; McCune, S.A.; Altschuld, R.A.; Lederer, W.J. Defective excitation-contraction coupling in experimental cardiac hypertrophy and heart failure. Science 1997, 276, 800–806. [Google Scholar] [CrossRef]
- Despa, S.; Brette, F.; Orchard, C.H.; Bers, D.M. Na/Ca exchange and Na/K-ATPase function are equally concentrated in transverse tubules of rat ventricular myocytes. Biophys. J. 2003, 85, 3388–3396. [Google Scholar] [CrossRef] [Green Version]
- Daniels, R.E.; Haq, K.T.; Miller, L.S.; Chia, E.W.; Miura, M.; Sorrentino, V.; McGuire, J.J.; Stuyvers, B.D. Cardiac expression of ryanodine receptor subtype 3; a strategic component in the intracellular Ca(2+) release system of Purkinje fibers in large mammalian heart. J. Mol. Cell Cardiol. 2017, 104, 31–42. [Google Scholar] [CrossRef]
- Landstrom, A.P.; Dobrev, D.; Wehrens, X.H.T. Calcium Signaling and Cardiac Arrhythmias. Circ. Res. 2017, 120, 1969–1993. [Google Scholar] [CrossRef]
- Patterson, E.; Lazzara, R.; Szabo, B.; Liu, H.; Tang, D.; Li, Y.-H.; Scherlag, B.J.; Po, S.S. Sodium-Calcium Exchange Initiated by the Ca2+Transient: An Arrhythmia Trigger Within Pulmonary Veins. J. Am. Coll. Cardiol. 2006, 47, 1196–1206. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rietdorf, K.; Masoud, S.; McDonald, F.; Sanderson, M.J.; Bootman, M.D. Pulmonary vein sleeve cell excitation–contraction-coupling becomes dysynchronized by spontaneous calcium transients. Biochem. Soc. Trans. 2015, 43, 410–416. [Google Scholar] [CrossRef]
- Lang, D.; Medvedev, R.Y.; Ratajczyk, L.; Zheng, J.; Yuan, X.; Lim, E.; Han, O.Y.; Valdivia, H.H.; Glukhov, A.V. Region-specific distribution of transversal-axial tubule system organization underlies heterogeneity of calcium dynamics in the right atrium. Am. J. Physiol. Heart Circ. Physiol. 2022, 322, H269–H284. [Google Scholar] [CrossRef] [PubMed]
- Pasqualin, C.; Gannier, F.; Malécot, C.O.; Bredeloux, P.; Maupoil, V. Automatic quantitative analysis of t-tubule organization in cardiac myocytes using ImageJ. Am. J. Physiol. Cell Physiol. 2015, 308, C237–C245. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Orchard, C.H.; Bryant, S.M.; James, A.F. Do t-tubules play a role in arrhythmogenesis in cardiac ventricular myocytes? J. Physiol. 2013, 591, 4141–4147. [Google Scholar] [CrossRef]



Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Cros, C.; Douard, M.; Chaigne, S.; Pasqualin, C.; Bru-Mercier, G.; Recalde, A.; Pascarel-Auclerc, C.; Hof, T.; Haïssaguerre, M.; Hocini, M.; et al. Regional Differences in Ca2+ Signaling and Transverse-Tubules across Left Atrium from Adult Sheep. Int. J. Mol. Sci. 2023, 24, 2347. https://doi.org/10.3390/ijms24032347
Cros C, Douard M, Chaigne S, Pasqualin C, Bru-Mercier G, Recalde A, Pascarel-Auclerc C, Hof T, Haïssaguerre M, Hocini M, et al. Regional Differences in Ca2+ Signaling and Transverse-Tubules across Left Atrium from Adult Sheep. International Journal of Molecular Sciences. 2023; 24(3):2347. https://doi.org/10.3390/ijms24032347
Chicago/Turabian StyleCros, Caroline, Matthieu Douard, Sebastien Chaigne, Come Pasqualin, Gilles Bru-Mercier, Alice Recalde, Caroline Pascarel-Auclerc, Thomas Hof, Michel Haïssaguerre, Meleze Hocini, and et al. 2023. "Regional Differences in Ca2+ Signaling and Transverse-Tubules across Left Atrium from Adult Sheep" International Journal of Molecular Sciences 24, no. 3: 2347. https://doi.org/10.3390/ijms24032347